Introduction
Preeclampsia (PE) occurs in 4–5% of pregnancies
worldwide, accounting for a large number of maternal complications
and contributing to a high rate of maternal morbidity (1). PE is characterized by the occurrence
of hypertension and proteinuria after 20 weeks of gestation in a
patient without a previous history of hypertension (2). In addition, patients with a clinical
history of PE are at an increased risk of developing PE in
subsequent pregnancies (3). At
present, research is focused on further defining and characterizing
PE; however, the etiology of PE remains to be elucidated. Moreover,
further investigations into predicting PE development are required
(2,4).
PE is an inflammatory disease. Previous studies have
reported that an altered immune system response and excessive
inflammation can contribute to the development of PE (5,6).
Inflammatory responses at the utero-placental interface were
associated with aberrant extravillous trophoblast invasion during
placentation in transgenic preeclamptic rat models and human PE
(7,8). Several in vitro studies
previously suggested that exposure to inflammatory stimuli can
trigger the secretion of proinflammatory cytokines from trophoblast
cells to mediate PE (9,10). Inflammation occurs in healthy
pregnancies in women, yet increased levels of inflammation are
often a characteristic of PE (11). Regular nurse appointments are
critical for discovering complications in pregnancy (12). Checks during regular nurse
appointments include blood pressure measurement, PE diagnosis and
surveillance, and determination of optimal timing of delivery
(13).
Pentraxin 3 (PTX3), a member of the c-reactive
protein family, is a long pentraxin protein and a pro-inflammatory
marker (14). Under inflammatory
conditions, PTX3 is secreted by vascular endothelial cells and
monocytes (15). A number of
previous studies have reported elevated expression levels of PTX3
in the serum of pregnant women with PE (16–19). However, the specific role of PTX3
in gestational trophoblast cells is yet to be fully elucidated. It
has previously been reported that PTX3 is induced by interleukin
(IL)-1β in human endometrial stem cells (20). Mature IL-1β cytokine is a key
pro-inflammatory cytokine involved in the induction of a placental
inflammatory response (21).
Thus, the present study aimed to explore the effects of PTX3 on the
proliferation and invasion of trophoblasts, and the effects of
IL-1β on PTX3 expression.
Materials and methods
Cell culture and treatment
HTR-8/SV neo and JEG3 cells were provided by Procell
Life Science & Technology Co., Ltd. Cells were grown in
DMEM/nutrient mixture F12 (DMEM/F12; HyClone; Cytiva) with 10% FBS
(Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin-streptomycin at 37°C with 5% CO2.
Untransfected HTR-8/SV neo and JEG3 cells were used as the control
group. The vector overexpressing PTX3 (Ov-PTX3) at a concentration
of 20 µM was purchased from Wuhan GeneCreate Biological Engineering
Co., Ltd. by utilizing pcDNA3.1 vector containing the full-length
PTX3 cDNA sequence, regarding the empty vector (Ov-NC) as the
corresponding negative control. These plasmids were transfected
into HTR-8/SV neo and JEG3 cells (1×106 cells/well) by
using Lipofectamine® 3000 (cat. no. L3000015;
Invitrogen; Thermo Fisher Scientific, Inc.) at 37°C for 48 h.
Transfection efficacy was evaluated by performing reverse
transcription-quantitative (RT-q)PCR and western blot analysis 48 h
post-transfection. When ~80% confluence was reached, cells were
cultured in serum-starved DMEM/F12 with or without IL-1β (R&D
Systems, Inc.) or 1 µg/ml IL-1 receptor antagonist (IL-1Ra; Swedish
Orphan Biovitrum AB) for 24, 48 or 72 h at 37°C. IL-1Ra was used to
pretreat cells at 37°C for 1 h before transfection.
Cell Counting Kit-8 (CCK-8) assay
For the detection of cell proliferation, cell
suspensions were inoculated in a 96-well plate at 5×103
cells/well and incubated at 37°C with 5% CO2. CCK-8
reagent (Shanghai Yeasen Biotechnology Co., Ltd.) was subsequently
added into each well, and cells were cultured for a further 3 h.
The absorbance was read using a microplate reader at 450 nm.
RT-qPCR assay
Total RNA was extracted from cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). Total RNA was reverse transcribed into cDNA by using the
iScript™ cDNA synthesis kit (Bio-Rad Laboratories, Inc.) in
accordance with the manufacturer's protocol. qPCR was performed
using SYBR Premix Ex Taq (Takara Biotechnology Co., Ltd.) on a
7900HT Fast Real-Time PCR System (Applied Biosystems; Thermo Fisher
Scientific, Inc.) with the following thermocycling conditions:
Initial denaturation at 95°C for 2 min; followed by 40 cycles of
95°C for 20 sec and 72°C for 30 sec. The sequences of the primers
designed were: PTX3 forward, 5′-CATCTCCTTGCGATTCTGTTTTG-3′ and
reverse, 5′-CCATTCCGAGTGCTCCTGA-3′; MMP2 forward,
5′-TACAGGATCATTGGCTACACACC-3′ and reverse,
5′-GGTCACATCGCTCCAGACT-3′; MMP9 forward,
5′-TGTACCGCTATGGTTACACTCG-3′ and reverse,
5′-GGCAGGGACAGTTGCTTCT-3′; and GAPDH forward,
5′-GGAGCGAGATCCCTCCAAAAT-3′ and reverse,
5′-GGCTGTTGTCATACTTCTCATGG-3′. The relative mRNA expression of
target genes was calculated using the 2−ΔΔCq method
(22) and data were normalized to
the GAPDH expression levels detected in the same sample.
Western blot analysis
The treated cells were harvested, lysed in RIPA
lysis buffer (Beyotime Institute of Biotechnology) and proteins
were quantified using a BCA kit (Thermo Fisher Scientific, Inc.).
An equal amount of protein lysates (20 µg) were separated via 10%
SDS-PAGE and then transferred to PVDF membranes. The membranes were
subsequently blocked with 5% non-fat milk for 1.5 h at room
temperature and incubated with primary antibodies overnight at 4°C
at dilutions of 1:1,000, according to the manufacturer's protocol.
Primary antibodies for PTX3 (cat. no. ab90806), MMP2 (cat. no.
ab92536) and MMP9 (ab76003) were obtained from Abcam. After washing
with PBS three times, the membranes were incubated with
HRP-conjugated goat anti-rabbit secondary antibody (A0208) at room
temperature for 1 h (Beyotime Institute of Biotechnology). Proteins
bands were visualized using enhanced chemiluminescence (Thermo
Fisher Scientific, Inc.). Band intensity was semi-quantified using
ImageJ software (version 1.48v; National Institutes of Health).
Cell cycle detection via flow
cytometry
Flow cytometry was conducted to determine the cell
distribution in phases. Cells were collected 48 h
post-transfection, washed with PBS and fixed in 70% ethanol at
−20°C. Following overnight fixation, cells were washed with PBS and
stained with propidium iodide (Beckman Coulter, Inc.) for 30 min at
37°C. Cell cycle analysis was performed by using an Attune™ NxT
Acoustic Focusing Cytometer (Invitrogen; Thermo Fisher Scientific,
Inc.).
Transwell invasion assay
Transwell 24-well culture plates with a 8-µm pore
insert precoated with Matrigel (BD Biosciences) at 37°C for 30 min
were obtained. A total of 3×104 cells in 100 ml FBS-free
DMEM/F12 were placed in the upper chamber. DMEM/F12 containing 20%
FBS was added to the lower chamber. After 24 h of incubation at
37°C, cells were fixed with 4% paraformaldehyde for 20 min at 37°C
and then stained with crystal violet for 30 min at 37°C. The
stained cells were photographed under an inverted light microscope
(magnification, ×100; Olympus Corporation). ImageJ software
(version 1.48v; National Institutes of Health) was used to
calculate the numbers of cells invaded in five random images.
Statistical analysis
All experiments were repeated independently three
times. Data are indicated as the mean ± standard deviation. The
statistical differences among multiple groups were measured by
one-way analysis of variance followed by Tukey's post hoc test.
Graph Prism 6.0 software (GraphPad Software, Inc.) was used for
data analysis. P<0.05 was considered to indicate a statistically
significant difference.
Results
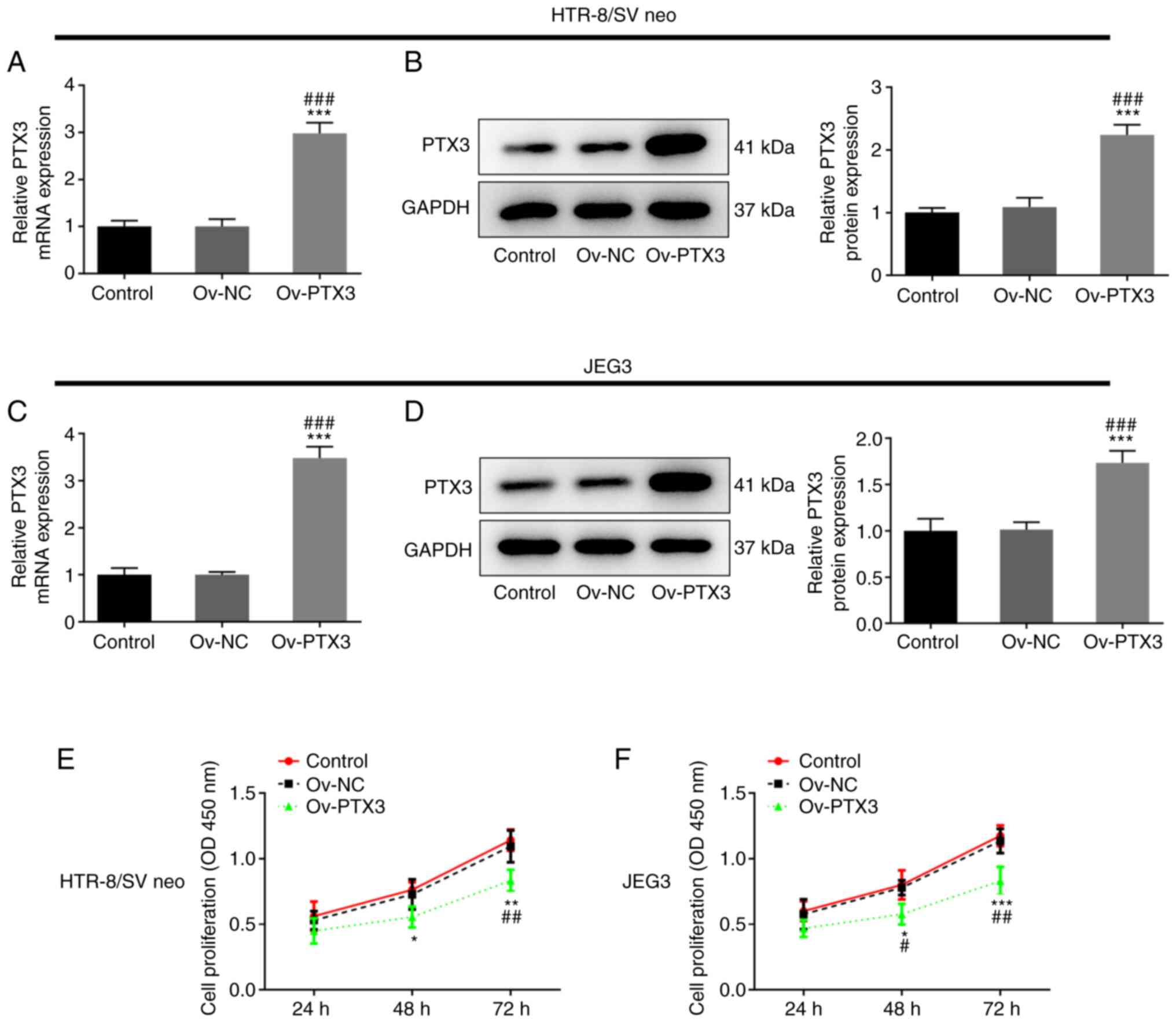
Overexpression of PTX3 inhibits the
proliferation of HTR-8/SV neo and JEG3 cells
It has previously been reported that PTX3 is
elevated in the plasma of patients with PE (16–19). Thus, PTX3 was overexpressed in
HTR-8/SV neo and JEG3 cells. As presented in Fig. 1A and B, PTX3 expression was
significantly upregulated in the Ov-PTX3 group compared with the
control or Ov-NC group (P<0.001) in HTR-8/SV neo cells.
Consistently, Ov-PTX3 transfection also led to upregulated PTX3
expression in JEG3 cells when compared with the control or Ov-NC
group (P<0.001; Fig. 1C and
D). A key characteristic of PE development is altered
trophoblast proliferation (23);
thus, the proliferation of HTR-8/SV neo and JEG3 cells was detected
following PTX3 overexpression. Notably, the proliferation of
HTR-8/SV neo (P<0.05, P<0.01; Fig. 1E) and JEG3 (P<0.05, P<0.01,
P<0.001; Fig. 1F) cells was
significantly reduced following PTX3 overexpression. These results
revealed that overexpression of PTX3 inhibited the proliferation of
HTR-8/SV neo and JEG3 cells.
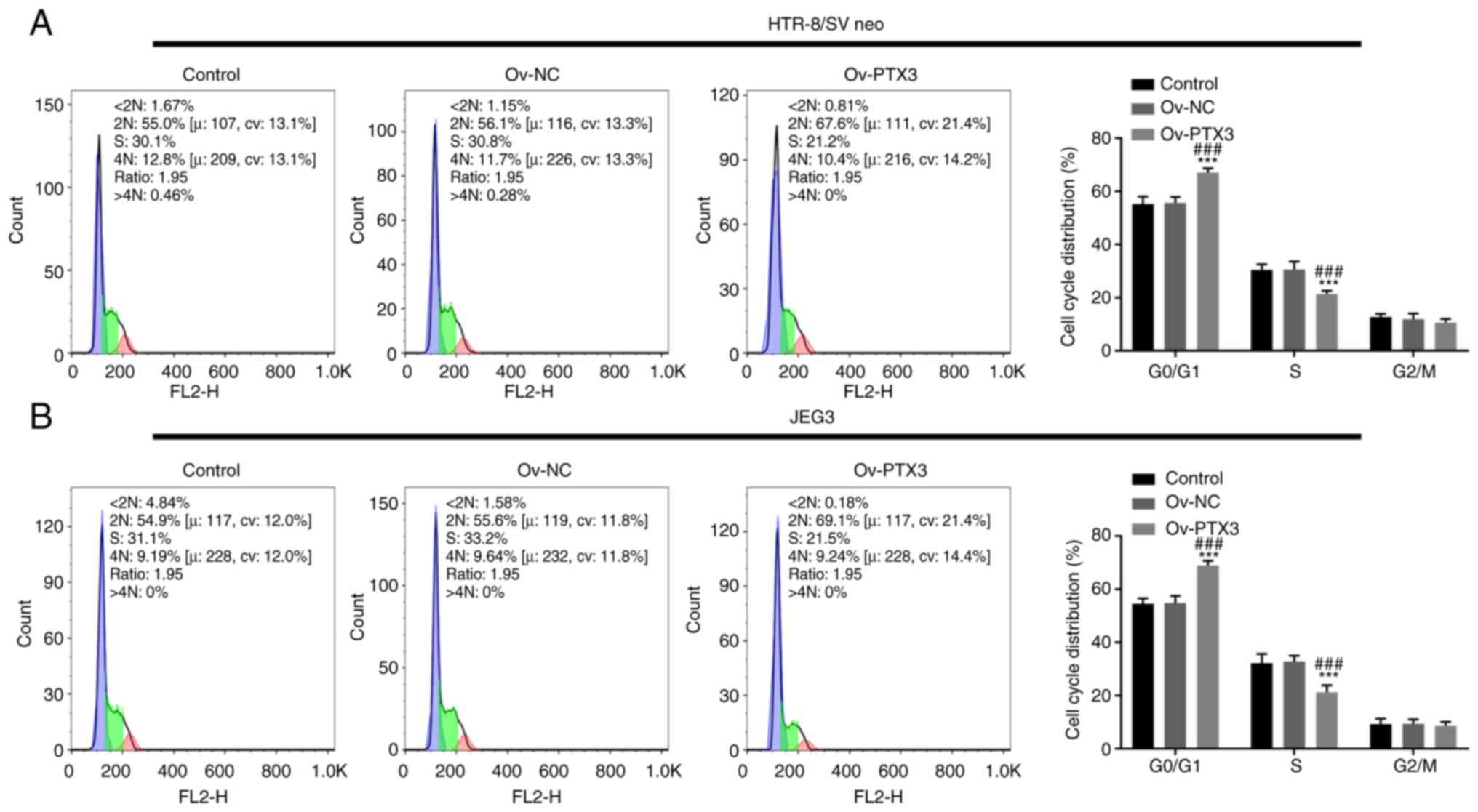
Overexpression of PTX3 inhibits the
cell cycle promotion and invasion of HTR-8/SV neo and JEG3
cells
Following PTX3 overexpression, the
G0/G1 phase of HTR-8/SV neo and JEG3 cells
was prolonged, while the S phase was reduced compared with the
control or Ov-NC group, suggesting that the cell cycle was arrested
(P<0.001; Fig. 2A and B).
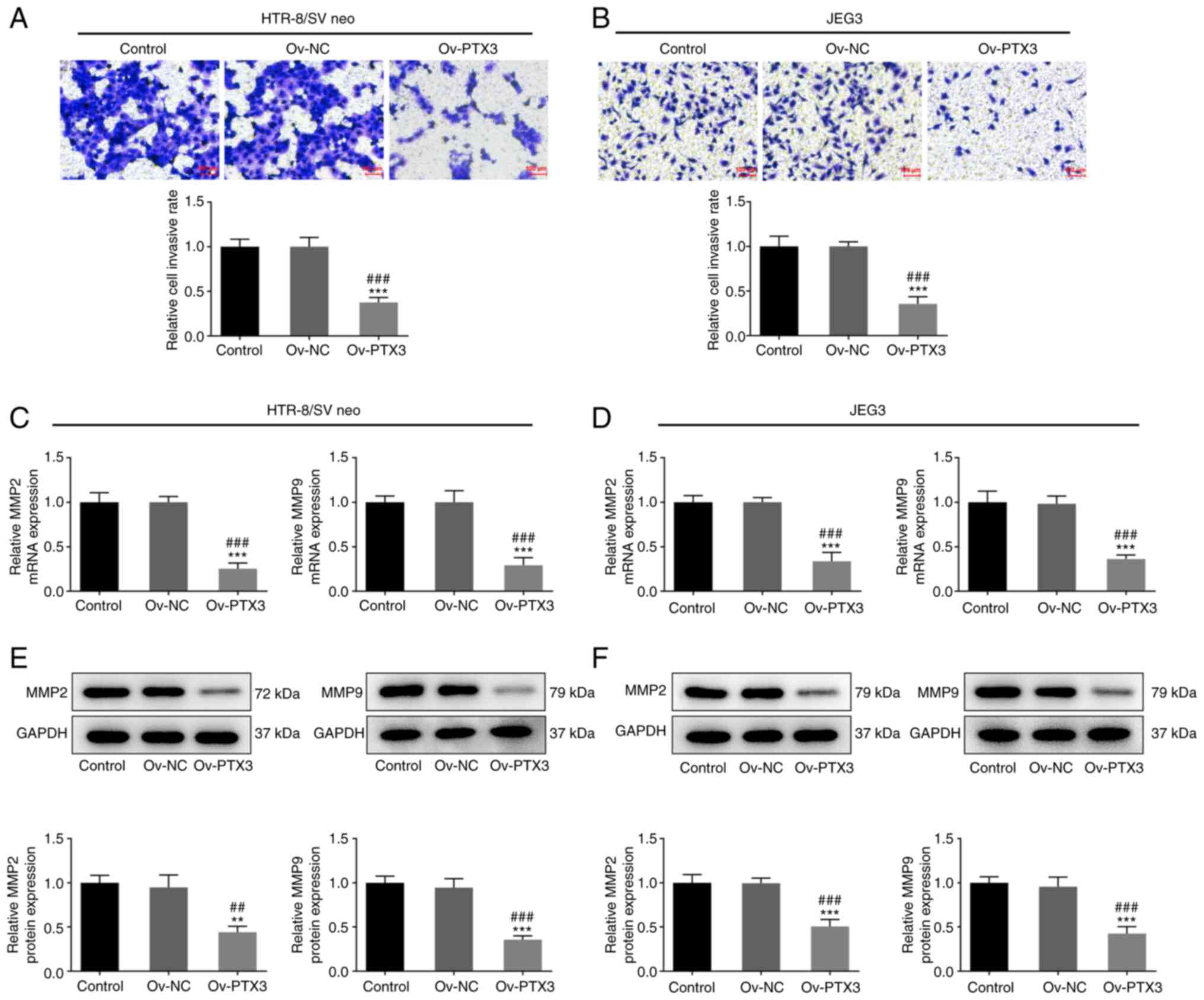
Furthermore, the invasion of HTR-8/SV neo and JEG3 cells in the
Ov-PTX3 group was decreased compared with the control or Ov-NC
group (P<0.001; Fig. 3A and
B). MMPs, which are zinc-dependent proteases involved in the
uterine and vascular tissue remodeling during healthy pregnancy
(24,25), are associated with the invasion of
trophoblasts (26). Thus, the
expression levels of MMP2 and MMP9 were detected following PTX3
overexpression. Notably, expression levels of MMP2 and MMP9 mRNA
were reduced following PTX3 overexpression in both HTR-8/SV neo
(P<0.001; Fig. 3C) and JEG3
(P<0.001; Fig. 3D) cells
compared with the control or Ov-NC group. Meanwhile, significantly
upregulated MMP2 and MMP9 protein expression was observed in
HTR-8/SV neo (P<0.01, P<0.001; Fig. 3E) and JEG3 (P<0.001; Fig. 3F) cells compared with the control
or Ov-NC group. Collectively, these results demonstrated that PTX3
overexpression inhibited the cell cycle promotion and invasion of
HTR-8/SV neo and JEG3 cells.
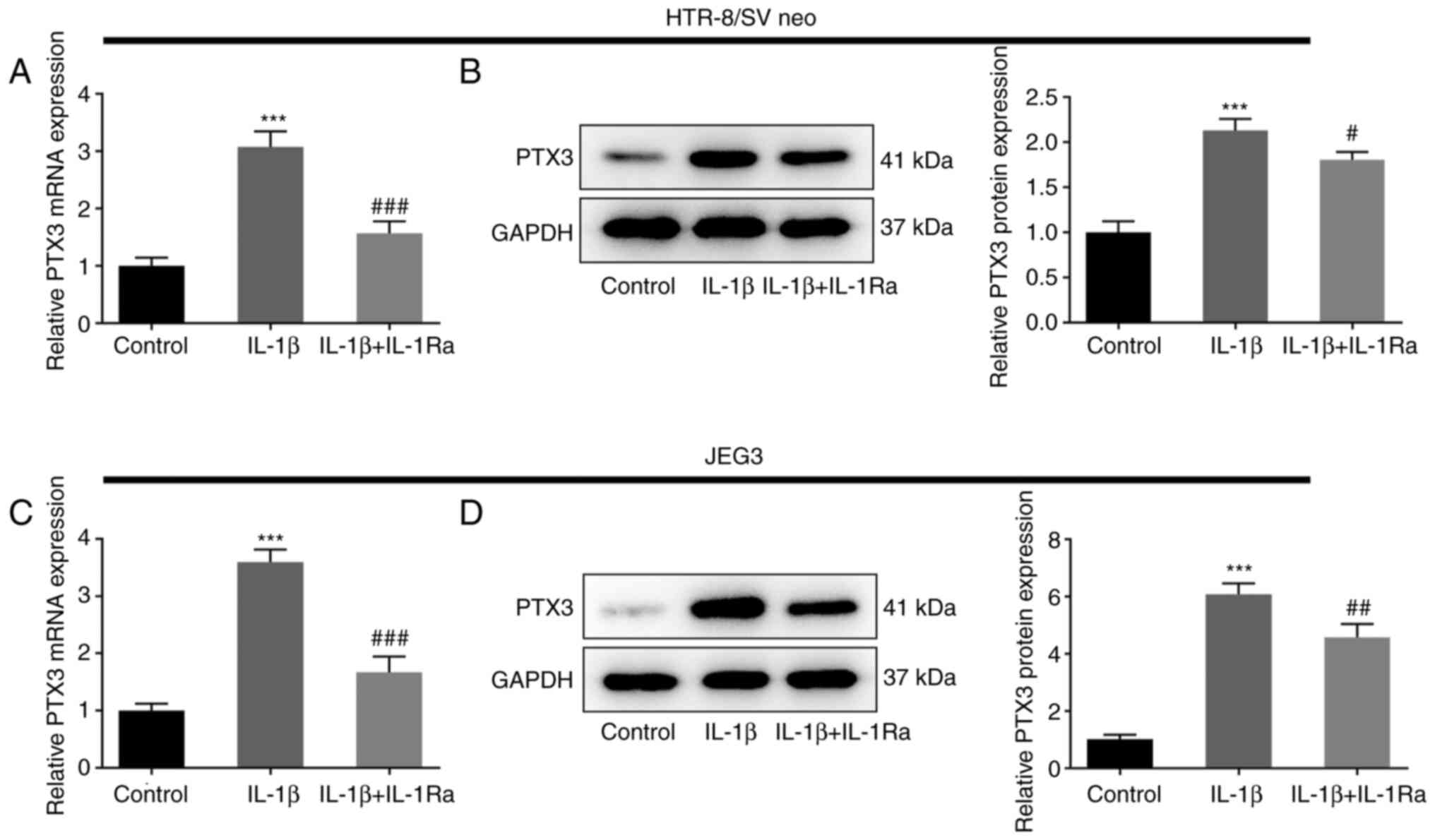
IL-1β induces the expression of PTX3
in HTR-8/SV neo and JEG3 cells, which is suppressed by the IL-1β
antagonist
IL-1β, an essential inflammatory cytokine involved
in the mediation of inflammatory responses, promotes the
differentiation and motility of extravillous trophoblasts (27). Thus, the effects of IL-1β on the
functions of extravillous trophoblasts were detected following PTX3
overexpression in HTR-8/SV neo and JEG3 cells. Following treatment
with IL-1β, the expression of PTX3 was significantly increased in
HTR-8/SV neo and JEG3 cells, compared with the control group
(P<0.001). However, co-treatment with IL-1β and IL-1Ra
attenuated the expression of PTX3, compared with cells treated with
IL-1β alone (P<0.05, P<0.01, P<0.001; Fig. 4A-D). Collectively, these results
revealed that IL-1β induced the expression of PTX3 in HTR-8/SV neo
and JEG3 cells, which was suppressed following treatment with an
IL-1β antagonist.
IL-1Ra reverses the inhibitory effects
of PTX3 overexpression on proliferation, cell cycle, and invasion
of HTR-8/SV neo and JEG3 cells
As the results of the present study demonstrated
that IL-1Ra inhibited the promoting effects of PTX3 overexpression
in HTR-8/SV neo and JEG3 cells, both cell lines were treated with
Ov-PTX3 and IL-1Ra to detect any changes in the activities of
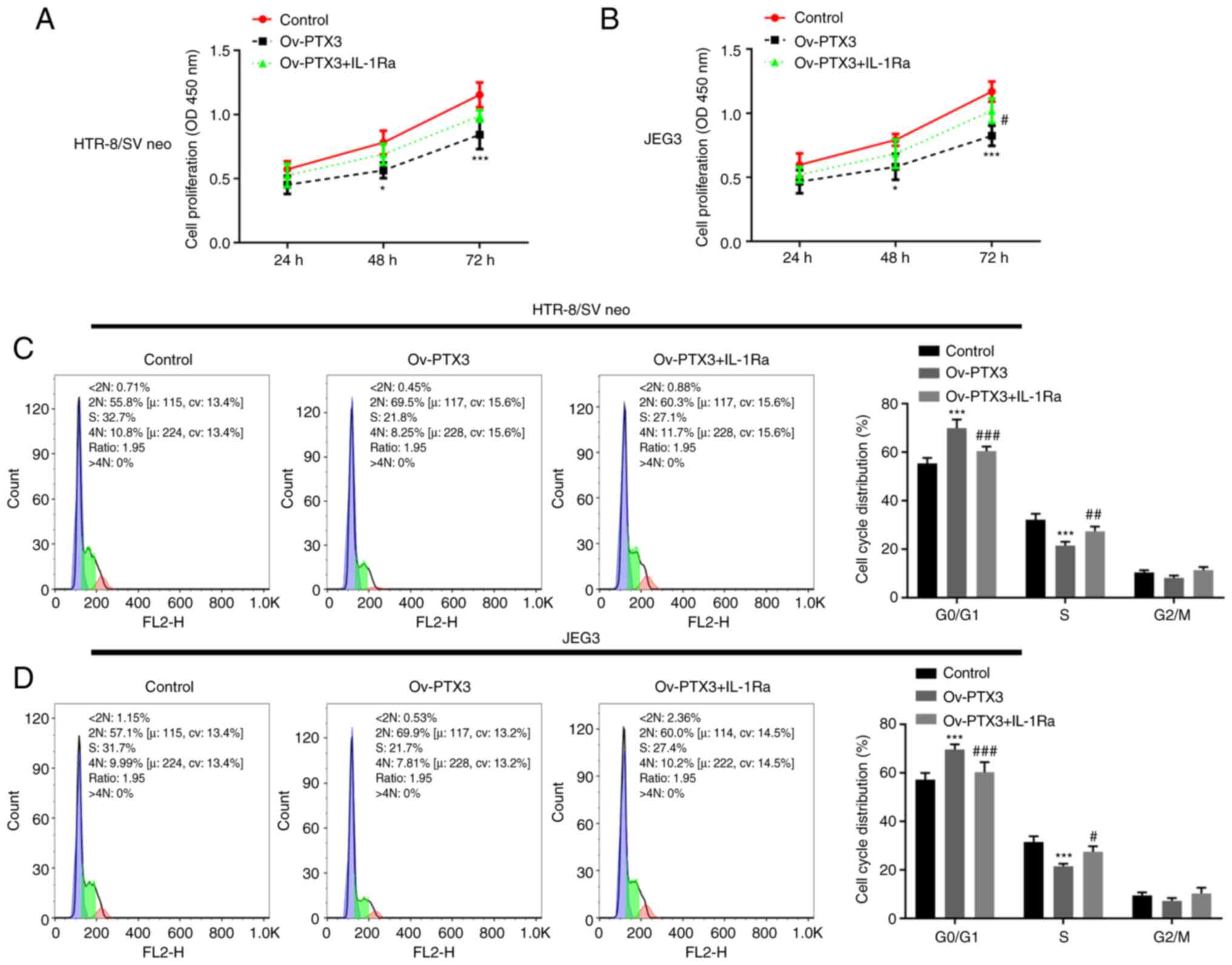
cells. Results of the CCK-8 analysis indicated that IL-1Ra
treatment increased the levels of proliferation suppressed by
Ov-PTX3 (P<0.05; Fig. 5A and
B). Moreover, compared with the Ov-PTX3 group, treatment with
IL-1Ra in HTR-8/SV neo and JEG3 cells caused a shortened
G0/G1 phase and prolonged S phase, indicated
by the results of flow cytometry (P<0.05, P<0.01, P<0.001;
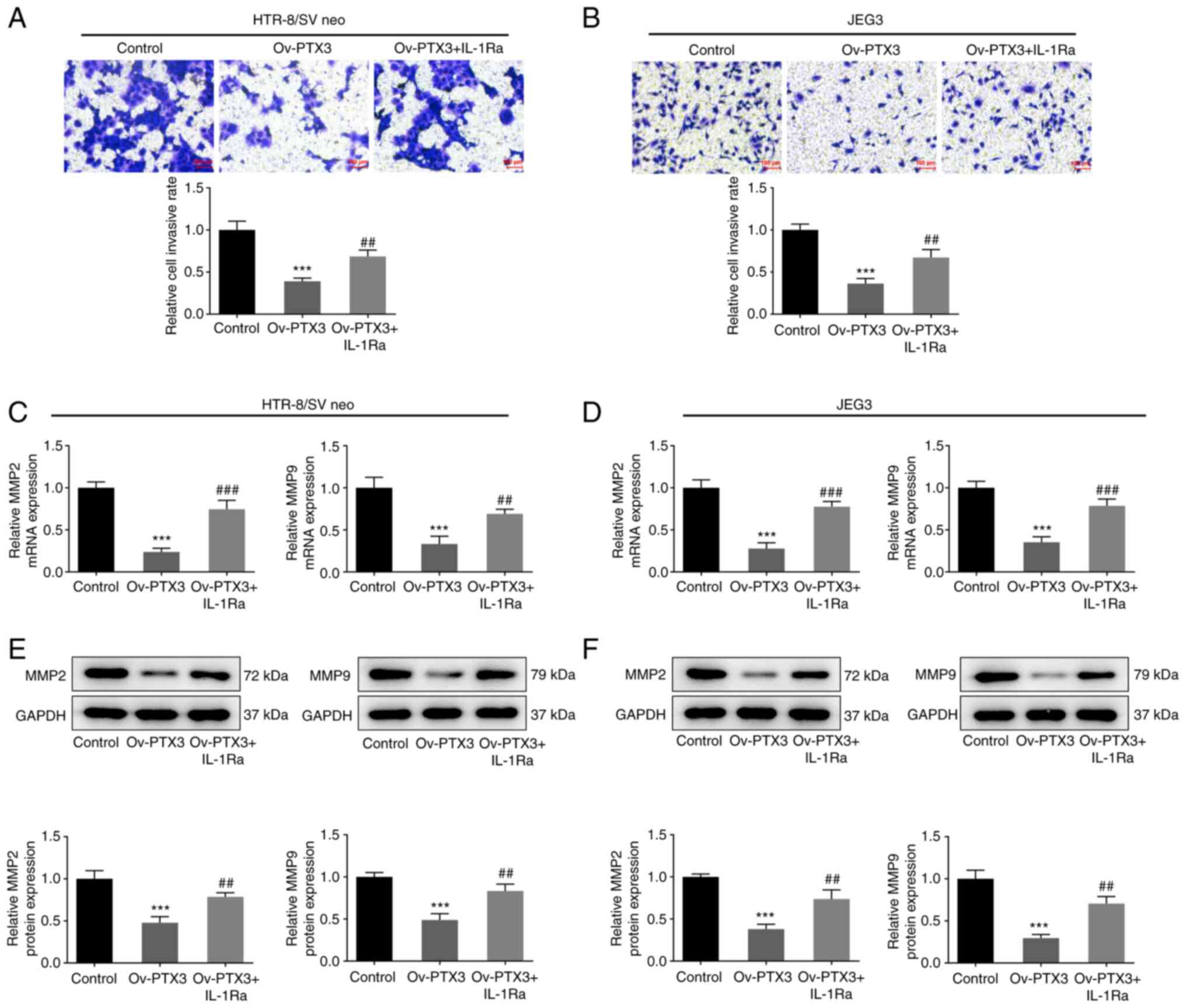
Fig. 5C and D). Furthermore, the
levels of invasion in both HTR-8/SV neo (P<0.01; Fig. 6A) and JEG3 (P<0.01; Fig. 6B) cells were increased following
treatment with IL-1Ra and Ov-PTX3, compared with the Ov-PTX3 group.
Consistently, the mRNA expression of MMP2 and MMP9 in HTR-8/SV neo
(P<0.01, P<0.001; Fig. 6C)
and JEG3 (P<0.001; Fig. 6D)
cells was significantly elevated following treatment with IL-1Ra
and Ov-PTX3, compared with the Ov-PTX3 group. The MMP2 and MMP9
protein expression levels exhibited the same trends in HTR-8/SV neo
(P<0.01; Fig. 6E) and JEG3
(P<0.001; Fig. 6F) cells
following treatment with IL-1Ra and Ov-PTX3, compared with the
Ov-PTX3 group. These findings suggested that IL-1Ra reversed the
inhibitory effects of PTX3 overexpression on proliferation, cell
cycle and invasion of HTR-8/SV neo and JEG3 cells.
Discussion
The results of the present study demonstrated that
compared with the control group, the Ov-PTX3 group in both HTR-8/SV
neo and JEG3 cells exhibited lower proliferation, invasion and cell
cycle promotion of trophoblasts in PE. Results of a previous study
demonstrated high levels of PTX3 expression in a number of
diseases, including inflammatory diseases. Moreover, high PTX3
plasma levels were observed in mice fed with a high-fat diet, which
contributed to the development of obesity (28). High expression levels of PTX3 were
also revealed in critically ill patients and those who were
infected with sepsis or septic shock, which were associated with
high rates of mortality (29,30). Thus, PTX3 has the ability to act
as a successful biological marker for the diagnosis of disease
mortality and severity, in contrast with other representative
markers (29,31).
Although investigations into PE are ongoing, it is
classified into two subtypes: Early-onset PE (delivery before 34
weeks of gestation) and late-onset PE (delivery after 34 weeks of
gestation) (32). This disease is
characterized by inadequate trophoblast invasion in the uterus,
poor remodeling of the spiral arteries and redundant trophoblast
apoptosis in the placenta (33).
A growing body of evidence suggests that MMP2 and MMP9 play a role
in endometrial tissue remodeling during the menstrual cycle and
pregnancy (34,35). MMP2 and MMP9 are abundantly
expressed in invading extravillous trophoblast cells, and the
expression of MMP2 and MMP9 is highly related to trophoblast cell
invasiveness (36,37). Additionally, it is likely that
other MMPs may also be involved in the progression of PE, therefore
the expression of additional MMPs will be examined in further
studies. Thus, the alteration of MMP2 and MMP9 expression was
investigated in the present study; mRNA and protein levels of MMP2
and MMP9 in HTR-8/SV neo and JEG3 cells were suppressed following
PTX2 overexpression, highlighting a role of PTX3 in trophoblast
migration and invasion in PE development.
PTX3 expression is induced by stimuli, including
inflammatory cytokines, Toll-like receptor agonists, distinct
microbial moieties and a number of microorganisms (38). Furthermore, the IL-1β pathway
plays an essential role in pregnancy (39). The results of a previous study
demonstrated that TNF-α and IL-1β are crucial in abnormal
extravillous trophoblast invasion in PE (40). It has been reported that plasma
level of IL-1β is significantly increased in preeclampsia (41). Luppi and Deloia (42) also demonstrated that the
spontaneous intracellular synthesis of IL-1β in monocytes of
preeclamptic women was higher than in normal pregnant and
non-pregnant women. As PE is an inflammatory disease in which PTX3
is highly expressed, it was hypothesized in the current study that
IL-1β played a role in inducing the expression of PTX3 for the
promotion of PE development. Thus, IL-1β was used to induce both
HTR-8/SV neo and JEG3 cells in the present study. The expression
level of PTX3 was significantly increased following IL-1β
treatment, which was partially reversed by the further addition of
IL-1Ra, suggesting that there may be an incomplete inhibitory
effect of IL-1Ra or additional pathways induced by IL-1β may be
implicated. This result was consistent with those of a previous
study, which highlighted that a single cytokine or a single element
of the immune system would not account for all normal trophoblastic
activities (43). IL-1Ra binds to
IL-1R1 with a high affinity and inhibits IL-1α or IL-1β function
(44). In the present study,
based on the result that IL-1Ra treatment reduced PTX3 level in
IL-1β-induced HTR-8/SV neo and JEG3 cells, we speculated that
IL-1Ra treatment would also reduce the level of PTX3 in
PTX3-overexpressing cells. In the subsequent experiments, both
HTR-8/SV neo and JEG3 cells were treated with Ov-PTX3 and IL-1Ra,
and cell proliferation was subsequently increased, compared with
those treated with Ov-PTX3 alone. Furthermore, Ov-PTX3-induced
decreases in invasion and cell cycle promotion were restored
following treatment with IL-1Ra. Since IL-1Ra binds to IL-1R1
(common receptor for both IL-1β and IL-1α) (45), it may inhibit the function of
IL-1β and IL-1α cytokines. Therefore, in future studies, whether
IL-1α also affects PTX3 expression and PE development may require
further research.
However, this experiment has some limitations. In
this study, the effect of PTX3 overexpression on cell cycle
pathways and caspase activity were not explored, which is an aim of
our future studies. Besides, lacking the use of additional groups
(mutant PTX3, Ov-NC + IL-1Ra or IL-Rα alone treatment groups) and
further mechanistic experiments to unravel the pathways mediating
the effect of PTX3 on PE development (including proliferation,
invasion and cell cycle of trophoblasts) are potential limitations
of the present study. Additionally, this study only discussed the
effects of PTX3 on PE in HTR-8/SV neo and JEG3 cells by PTX3
overexpression, but had no the data concerning PTX3 silencing or
clinical data to show the relationship between PTX3, IL-1β
expression and PE development, which is also a potential limitation
of this paper.
Collectively, the results of the present study and
previous investigations using in vitro models revealed that
the elevated expression levels of PTX3 induced by IL-1β inhibited
the proliferation, invasion and cell cycle of trophoblasts, thus
triggering the progression of PE. Therefore, PTX3 knockout by
targeting IL-1β may lead to the prevention of PE development.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XW and JZ designed the study and analyzed the data.
JJ performed the experiments. XW and JZ drafted the manuscript and
interpreted the data. XW and JJ confirm the authenticity of all the
raw data. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hauth JC, Ewell MG, Levine RJ, Esterlitz
JR, Sibai B, Curet LB, Catalano PM and Morris CD: Pregnancy
outcomes in healthy nulliparas who developed hypertension. Calcium
for preeclampsia prevention study group. Obstet Gynecol. 95:24–28.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chappell LC, Cluver CA, Kingdom J and Tong
S: Pre-eclampsia. Lancet. 398:341–354. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Barton JR and Sibai BM: Prediction and
prevention of recurrent preeclampsia. Obstet Gynecol. 112:359–372.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Karumanchi SA: Angiogenic factors in
preeclampsia: From diagnosis to therapy. Hypertension.
67:1072–1079. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shaw J, Tang Z, Schneider H, Salje K,
Hansson SR and Guller S: Inflammatory processes are specifically
enhanced in endothelial cells by placental-derived TNF-alpha:
Implications in preeclampsia (PE). Placenta. 43:1–8. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Harmon AC, Cornelius DC, Amaral LM,
Faulkner JL, Cunningham MW Jr, Wallace K and LaMarca B: The role of
inflammation in the pathology of preeclampsia. Clin Sci (Lond).
130:409–419. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cotechini T, Komisarenko M, Sperou A,
Macdonald-Goodfellow S, Adams MA and Graham CH: Inflammation in rat
pregnancy inhibits spiral artery remodeling leading to fetal growth
restriction and features of preeclampsia. J Exp Med. 211:165–179.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fan M, Li X, Gao X, Dong L, Xin G, Chen L,
Qiu J and Xu Y: LPS induces preeclampsia-like phenotype in rats and
HTR8/SVneo cells dysfunction through TLR4/p38 MAPK pathway. Front
Physiol. 10:10302019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kadam L, Kilburn B, Baczyk D, Kohan-Ghadr
HR, Kingdom J and Drewlo S: Rosiglitazone blocks first trimester
in-vitro placental injury caused by NF-κB-mediated inflammation.
Sci Rep. 9:20182019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zaga-Clavellina V, Garcia-Lopez G,
Flores-Herrera H, Espejel-Nuñez A, Flores-Pliego A,
Soriano-Becerril D, Maida-Claros R, Merchant-Larios H and
Vadillo-Ortega F: In vitro secretion profiles of interleukin
(IL)-1beta, IL-6, IL-8, IL-10, and TNF alpha after selective
infection with Escherichia coli in human fetal membranes. Reprod
Biol Endocrinol. 5:462007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Poston L: Endothelial dysfunction in
pre-eclampsia. Pharmacol Rep. 58 (Suppl 1):S69–S74. 2006.PubMed/NCBI
|
|
12
|
Black KD and Morin KH: Development and
testing of the preeclampsia prenatal symptom-monitoring checklist
(PPSMC). J Nurs Meas. 22:14–28. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Anderson CM and Schmella MJ: CE:
Preeclampsia: Current approaches to nursing management. Am J Nurs.
117:30–38. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xiong Z, Wang X, Jiang S, Jin M and Chen
W: Association between pentraxin-3 and the risk of preeclampsia: A
meta-analysis. Medicine (Baltimore). 99:e207442020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Napoleone E, di Santo A, Peri G, Mantovani
A, de Gaetano G, Donati MB and Lorenzet R: The long pentraxin PTX3
up-regulates tissue factor in activated monocytes: Another link
between inflammation and clotting activation. J Leukoc Biol.
76:203–209. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Akhter T, Wikstrom AK, Larsson M, Larsson
A, Wikstrom G and Naessen T: Serum Pentraxin 3 is associated with
signs of arterial alteration in women with preeclampsia. Int J
Cardiol. 241:417–422. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ovayolu A, Turksoy VA, Ovayolu G, Ozek MA,
Dogan I and Karaman E: Analyses of interleukin-6, presepsin and
pentraxin-3 in the diagnosis and severity of late-onset
preeclampsia. J Matern Fetal Neonatal Med. 35:299–307. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Turkmen O, Mollaoglu S, Goynumer G and
Isbilen B: Plasma pentraxin 3 levels in preeclamptic patients. Clin
Exp Obstet Gynecol. 42:220–223. 2015.PubMed/NCBI
|
|
19
|
Hamad RR, Eriksson MJ, Berg E, Larsson A
and Bremme K: Impaired endothelial function and elevated levels of
pentraxin 3 in early-onset preeclampsia. Acta Obstet Gynecol Scand.
91:50–56. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Popovici RM, Krause MS, Jauckus J,
Germeyer A, Brum IS, Garlanda C, Strowitzki T and von Wolff M: The
long pentraxin PTX3 in human endometrium: Regulation by steroids
and trophoblast products. Endocrinology. 149:1136–1143. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Abrahams VM: The role of the Nod-like
receptor family in trophoblast innate immune responses. J Reprod
Immunol. 88:112–117. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kaya B, Nayki U, Nayki C, Ulug P, Oner G,
Gultekin E and Yildirim Y: Proliferation of trophoblasts and Ki67
expression in preeclampsia. Arch Gynecol Obstet. 291:1041–1046.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fathipour V, Khaki Z and Nassiri SM:
Evaluation of matrix metalloproteinases (MMP)-2 and MMP-9 activity
in serum and biochemical and hematological parameters in
spontaneous canine cutaneous tumors before and after surgical
treatment. Vet Res Forum. 9:19–26. 2018.PubMed/NCBI
|
|
25
|
Li W, Mata KM, Mazzuca MQ and Khalil RA:
Altered matrix metalloproteinase-2 and −9 expression/activity links
placental ischemia and anti-angiogenic sFlt-1 to uteroplacental and
vascular remodeling and collagen deposition in hypertensive
pregnancy. Biochem Pharmacol. 89:370–385. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shimonovitz S, Hurwitz A, Dushnik M,
Anteby E, Geva-Eldar T and Yagel S: Developmental regulation of the
expression of 72 and 92 kd type IV collagenases in human
trophoblasts: A possible mechanism for control of trophoblast
invasion. Am J Obstet Gynecol. 171:832–838. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Prutsch N, Fock V, Haslinger P, Haider S,
Fiala C, Pollheimer J and Knöfler M: The role of interleukin-1β in
human trophoblast motility. Placenta. 33:696–703. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bonacina F, Moregola A, Porte R, Baragetti
A, Bonavita E, Salatin A, Grigore L, Pellegatta F, Molgora M,
Sironi M, et al: Pentraxin 3 deficiency protects from the metabolic
inflammation associated to diet-induced obesity. Cardiovasc Res.
115:1861–1872. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Muller B, Peri G, Doni A, Torri V,
Landmann R, Bottazzi B and Mantovani A: Circulating levels of the
long pentraxin PTX3 correlate with severity of infection in
critically ill patients. Crit Care Med. 29:1404–1407. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mauri T, Bellani G, Patroniti N, Coppadoro
A, Peri G, Cuccovillo I, Cugno M, Iapichino G, Gattinoni L, Pesenti
A and Mantovani A: Persisting high levels of plasma pentraxin 3
over the first days after severe sepsis and septic shock onset are
associated with mortality. Intensive Care Med. 36:621–629. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mauri T, Coppadoro A, Bellani G, Bombino
M, Patroniti N, Peri G, Mantovani A and Pesenti A: Pentraxin 3 in
acute respiratory distress syndrome: An early marker of severity.
Crit Care Med. 36:2302–2308. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Huppertz B: The critical role of abnormal
trophoblast development in the etiology of preeclampsia. Curr Pharm
Biotechnol. 19:771–780. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Longtine MS, Chen B, Odibo AO, Zhong Y and
Nelson DM: Villous trophoblast apoptosis is elevated and restricted
to cytotrophoblasts in pregnancies complicated by preeclampsia,
IUGR, or preeclampsia with IUGR. Placenta. 33:352–359. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang X, Qi C and Lin J: Enhanced
expressions of matrix metalloproteinase (MMP)-2 and −9 and vascular
endothelial growth factors (VEGF) and increased microvascular
density in the endometrial hyperplasia of women with anovulatory
dysfunctional uterine bleeding. Fertil Steril. 93:2362–2367. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mishra B, Kizaki K, Koshi K, Ushizawa K,
Takahashi T, Hosoe M, Sato T, Ito A and Hashizume K: Expression of
extracellular matrix metalloproteinase inducer (EMMPRIN) and its
related extracellular matrix degrading enzymes in the endometrium
during estrous cycle and early gestation in cattle. Reprod Biol
Endocrinol. 8:602010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Su MT, Tsai PY, Tsai HL, Chen YC and Kuo
PL: MiR-346 and miR-582-3p-regulated EG-VEGF expression and
trophoblast invasion via matrix metalloproteinases 2 and 9.
Biofactors. 43:210–219. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Isaka K, Usuda S, Ito H, Sagawa Y,
Nakamura H, Nishi H, Suzuki Y, Li YF and Takayama M: Expression and
activity of matrix metalloproteinase 2 and 9 in human trophoblasts.
Placenta. 24:53–64. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kunes P, Holubcova Z, Kolackova M and
Krejsek J: Pentraxin 3(PTX 3): An endogenous modulator of the
inflammatory response. Mediators Inflamm. 2012:9205172012.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Equils O, Kellogg C, McGregor J, Gravett
M, Neal-Perry G and Gabay C: The role of the IL-1 system in
pregnancy and the use of IL-1 system markers to identify women at
risk for pregnancy complications†. Biol Reprod. 103:684–694. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Takamura S, Takahashi A, Inoue Y and
Teraki Y: Effects of tumor necrosis factor-alpha, interleukin-23
and interleukin-17A inhibitors on bodyweight and body mass index in
patients with psoriasis. J Dermatol. 45:1130–1134. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kocyigit Y, Atamer Y, Atamer A, Tuzcu A
and Akkus Z: Changes in serum levels of leptin, cytokines and
lipoprotein in pre-eclamptic and normotensive pregnant women.
Gynecol Endocrinol. 19:267–273. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Luppi P and Deloia JA: Monocytes of
preeclamptic women spontaneously synthesize pro-inflammatory
cytokines. Clin Immunol. 118:268–275. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Alanbay I, Coksuer H, Ercan CM, Ustun Y,
Pala G, Karasahin KE, Ozturk O, Kurt I and Baser I:
Chitotriosidase, interleukin-1 beta and tumor necrosis factor alpha
levels in mild preeclampsia. Arch Gynecol Obstet. 285:1505–1511.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hannum CH, Wilcox CJ, Arend WP, Joslin FG,
Dripps DJ, Heimdal PL, Armes LG, Sommer A, Eisenberg SP and
Thompson RC: Interleukin-1 receptor antagonist activity of a human
interleukin-1 inhibitor. Nature. 343:336–340. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Powers NE, Swartzwelter B, Marchetti C, de
Graaf DM, Lerchner A, Schlapschy M, Datar R, Binder U, Edwards CK
III, Skerra A and Dinarello CA: PASylation of IL-1 receptor
antagonist (IL-1Ra) retains IL-1 blockade and extends its duration
in mouse urate crystal-induced peritonitis. J Biol Chem.
295:868–882. 2020. View Article : Google Scholar : PubMed/NCBI
|