Introduction
Atherosclerosis (AS) is a chronic progressive
inflammatory disease (1).
Inflammation and cell death are two key pathological mechanisms of
AS (2). In general, cell death
may occur via apoptosis, autophagy and necrosis; however, other
forms of cell death have been identified, including pyroptosis
(3). Pyroptosis is a form of
inflammasome-mediated cell death that is dependent on the
activation of caspase-1. The maturation of pro-IL-1β and pro-IL-18
is induced by caspase-1 cleavage (4). Pyroptosis has been indicated to be
associated with the death of human macrophages due to oxidative
damage, suggesting that pyroptosis has an important role in AS
development (5).
Pyroptosis is a type of programmed cell death that
accompanies an inflammatory response (3). Previous studies have reported that
inflammation, and even pyroptosis, have an important role in the
progression of cardiovascular diseases, including atherosclerosis,
diabetic cardiomyopathy, ischemia-reperfusion injury, heart failure
and myocardial infarction (6,7).
Pyroptosis differs from apoptosis and necrosis in that it mainly
manifests as inflammasome formation, caspase and gasdermin
activation and the formation of multiple protein holes in the cell
membrane, leading to the rapid loss of cell membrane integrity and
the release of a large number of pro-inflammatory factors (8). Consequently, pyroptosis may have a
prominent role in AS-related inflammation. However, it has remained
elusive whether it is possible to target and regulate pyroptotic
cells in atherosclerotic lesions to treat AS.
Endothelial cell (EC) dysfunction and even death are
a key and initial stage in the development of AS (9). Vascular endothelial dysfunction is
the first step in a complex and multifaceted process that
ultimately leads to the formation of a plaque, as well as the
formation of atherosclerotic lesions and their complications
(10). Studies have indicated
that caspase-1 activation in ECs is able to promote endothelial
activation and monocyte recruitment, leading to AS (11). Curcumin (Fig. 1A), a polyphenolic compound, is the
principal curcuminoid derived from the rhizomes of turmeric
(12); it is able inhibit
lipopolysaccharide-induced NOD-, LRR- and pyrin domain-containing
protein 3 (NLRP3) inflammasome, thereby inhibiting the activation
of IL-1β and caspase-1 (13,14). In addition, curcumin also exhibits
antioxidant and anti-tumor properties (15–17).
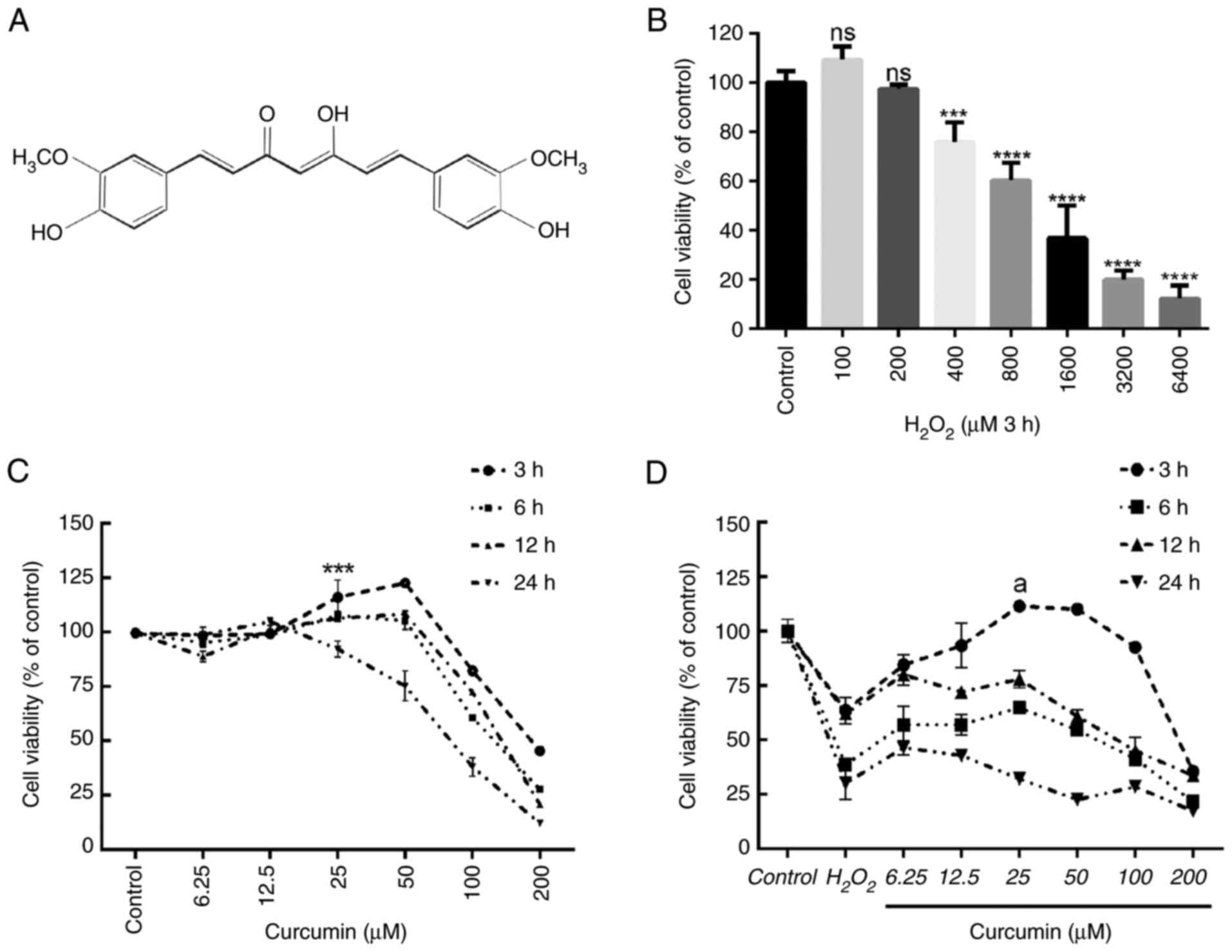 | Figure 1.Curcumin suppresses
H2O2-induced viability impairment of HUVECs.
The effects of increasing concentrations of
H2O2 and curcumin on HUVEC viability were
determined. (A) Curcumin contains a variety of functional groups,
including the β-diketo group, carbon-carbon double bonds and phenyl
rings containing varying amounts of hydroxyl and methoxy
substituents. Viability of HUVECs treated with increasing doses of
(B) H2O2 (0–6,400 µM) for 3 h and (C)
curcumin (0–200 µM) for 3, 6, 12 or 24 h, and (D) different
concentrations of curcumin (0–200 µM) for 3, 6, 12 or 24 h after 3
h of pretreatment with H2O2 (800 µM). Cell
viability was detected using an MTT assay. Values are expressed as
the mean ± standard deviation of three independent experiments.
***P<0.001, ****P<0.0001 vs. the control group;
aP<0.001 vs. the H2O2 group;
ns, no significance. HUVECs, human umbilical vein endothelial
cells. |
Therefore, it is conceivable that EC dysfunction and
atherogenesis may be associated with inflammation-related death
pathways. However, whether the effect of curcumin on EC function is
related to pyroptosis remains to be elucidated. In the present
study, the effects of curcumin on
H2O2-induced pyroptosis of human umbilical
vein ECs (HUVECs) were explored and a caspase-1 inhibitor (VX-765)
and NLRP3 inhibitor (MCC950) were used to corroborate the results.
The role of curcumin in EC dysfunction was also explored.
Materials and methods
Cell culture and treatment agents
The HUVEC line (immortal cells; no. C-003-5C) was
purchased from The Cell Bank of the Type Culture Collection of the
Chinese Academy of Sciences and cultured in RPMI-1640 medium (cat.
no. C11875500BT, Gibco; Thermo Fisher Scientific, Inc.), 10% FBS
(cat. no. FSP500; Excell Biotechnology Co., Ltd.) and 1%
penicillin/streptomycin at 37°C in a humidified atmosphere with 5%
CO2. Curcumin and H2O2 were
purchased from MilliporeSigma. VX-765 and MCC950 were purchased
from APExBIO Technology LLC. For experiments involving
pharmacological reagents, according to the different experimental
groups, HUVECs were treated with PBS (control),
H2O2 (800 µM) for 3 h, curcumin (25 µM) for 3
h, VX-765 (10 µM) for 1 h or MCC950 (10 µM) for 2 h.
Cell viability assay
An MTT assay (cat. no. C0009S; Beyotime Institute of
Biotechnology) was performed to evaluate the viability of HUVECs
according to the manufacturer's protocol. A total of
2×103 HUVECs per well were seeded in 96-well plates in
complete medium and treated with different concentrations of
curcumin (0–200 µM) for 3–24 h or H2O2
(0–6,400 µM) for 3–24 h. Next, 10 µl MTT reagent solution was added
to each well and the cells were incubated for a further 4 h at
37°C. The absorbance was measured at 490 nm using a Gen5 microplate
reader (BioTek Instruments).
Hematoxylin and eosin (H&E)
staining
The coverslip was placed in a six-well plate and
HUVECs (2×105 cells/well) were added to it and cultured
for 12 h. After HUVECs were attached to the coverslip, the cells
were fixed with 4% paraformaldehyde at room temperature (25°C) for
30 min. The cells were stained with hematoxylin solution included
in a kit (cat. no. C0105S; Beyotime Institute of Biotechnology) at
room temperature for 3 min and then placed under running tap water
at room temperature for 5 min. The cells were next stained in
working eosin Y solution from the kit at room temperature for 2
min. A drop of neutral balsam mounting medium was placed over the
cells on each slide and the coverslip was added. The slides were
then observed using a microscope (Axiolab 5; Carl Zeiss Suzhou Co.,
Ltd.).
DAPI/propidium iodide (PI) fluorescent
staining
In order to assess pyroptosis, cells were
double-stained with DAPI and PI (cat. no. G1012; Wuhan Servicebio
Technology Co., Ltd.). HUVECs (2×105 cells/well) were
cultured in a 6-well plate. HUVECs were treated with
H2O2 for 3 h and then either treated with 25
µM curcumin for 3 h or left untreated (control), whereas for the
VX-765 group, HUVECs were pretreated with caspase-1 inhibitor
VX-765 (10 µM) for 1 h and then incubated with
H2O2 (800 µM) for 3 h. Following treatment,
the cells in each group were washed with PBS three times and
stained with 50 µl PI (100 µg/ml) for 30 min and 50 µl DAPI for 5
min at 4°C in the dark. The stained cells were examined under a
fluorescent microscope (DS-Ri2; Nikon Corporation) at a
magnification of ×400.
Immunofluorescence
Immunofluorescence was performed to detect the
expression of caspase-1 in HUVECs. In brief, the cells were fixed
with 4% paraformaldehyde at room temperature for 30 min,
permeabilized with 0.6% Triton X-100 at room temperature for 0.5 h
and then blocked with goat serum (cat. no. C0265; Beyotime
Institute of Biotechnology) at room temperature for 0.5 h.
Subsequently, the cells were incubated with an anti-caspase-1
antibody (cat. no. ARG30330; dilution, 1:2,000; Arigo
Biolaboratories) or an anti-αvβ3 antibody (cat. no. ab190147;
dilution, 1:2,000; Abcam) at 4°C overnight, followed by incubation
with an FITC-conjugated secondary antibody (cat. no. GB22303;
dilution, 1:2,000; Wuhan Servicebio Technology Co., Ltd.) or a
CY3-conjugated secondary antibody (cat. no. GB21301; dilution,
1:2,000; Wuhan Servicebio Technology Co., Ltd.) in the dark at room
temperature for 1 h. The nuclei were stained with DAPI at room
temperature for 20 min. The cells were then imaged under a
fluorescence microscope (FV300; Olympus Corporation).
Analysis of DNA fragmentation
TUNEL staining was performed to detect DNA
fragmentation of HUVECs. In brief, HUVECs (2×105
cells/well) were cultured on coverslips in a 6-well plate.
Following the designated treatments, cells were fixed with 4%
paraformaldehyde and permeabilized at room temperature for 30 min
with 0.1% Triton X-100, followed by incubation with TUNEL reaction
mixture (cat. no. G1502; Wuhan Servicebio Technology Co., Ltd.) at
37°C in the dark for 1 h. Cells were then stained at room
temperature for 5 min with DAPI and examined under a fluorescent
microscope (DS-Ri2; Nikon Corporation).
ELISA
The relative concentrations of IL-1β and IL-18 in
the culture medium following the treatment of HUVECs with
H2O2, curcumin or VX-765 were determined
using specific ELISA kits (cat nos. EHC127.96 and EHC002b.96;
Neobioscience Technology Co., Ltd.), according to the
manufacturer's protocols.
Cytotoxicity assay
HUVECs were treated with curcumin, VX-765, MCC950
and H2O2. Cytotoxicity was determined by
measuring the lactate dehydrogenase (LDH) released from cells using
an LDH assay kit (cat. no. C0016; Beyotime Institute of
Biotechnology) according to the manufacturer's protocol. The
absorbance was determined at a wavelength of 450 nm using a
spectrophotometric microplate reader.
Western blot analysis
Cells were collected and proteins were extracted
using RIPA lysis buffer (cat. no. CW2333S; CoWin Biosciences), and
protease and phosphatase inhibitors were added. Protein
concentrations were determined using a BCA Protein Assay kit
(Bio-Rad Laboratories, Inc.). After loading 30 µg protein per lane,
proteins were separated using SDS-PAGE (10% separation gel and 5%
stacking gel) and transferred onto PVDF membranes (cat. no.
ISEQ00010; MilliporeSigma) followed by blocking with 5% skimmed
milk (cat. no. P0216; Beyotime Institute of Biotechnology.) at room
temperature for 2 h. Subsequently, the membranes were incubated
with primary antibodies against NLRP3 (cat. no. 19771-1-AP;
dilution, 1:1,000; ProteinTech Group, Inc.); apoptosis-associated
speck-like protein containing a caspase recruitment domain (ASC;
cat. no. 10500-1-AP; dilution, 1:1,000; ProteinTech Group, Inc.);
caspase-1, gasdermin D (GSDMD) and IL-1β (cat. no. ARG30330;
dilution, 1:1,000; Arigo Biolaboratories); and ET-1 (cat. no.
12191-1-AP; dilution, 1:1,000; ProteinTech Group, Inc.) at 4°C
overnight. Following washing with Tris-buffered saline with 0.1%
Tween-20 detergent (TBST) three times, the membranes were incubated
with an HRP-conjugated anti-rabbit or anti-mouse IgG secondary
antibody (cat. no. GB23301 and GB23303; dilution, 1:5,000; Wuhan
Servicebio Technology Co., Ltd.) for 1 h. Following washing with
TBST three times, the immunoreactive bands were detected using
chemiluminescence reagent (cat. no. CW0049S; CoWin Biosciences) and
visualized using a chemiluminescence gel imager (5200MuIti; Tanon
Science and Technology Co., Ltd.). The relative intensity was then
measured and analyzed using ImageJ 1.43u software (National
Institutes of Health).
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 8.0 software (GraphPad Software, Inc.) and values are
expressed as the mean ± standard deviation. Differences between
groups were determined using one-way ANOVA and Dunnett's
multiple-comparisons post-hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Curcumin alleviates
H2O2-induced EC injury
First, the effects of different concentrations of
H2O2 (0, 200, 400, 800, 1,600, 3,200 and
6,400 µM) on HUVEC viability were examined after 3 h. According to
the results of the MTT analysis, 800 µM H2O2
treatment for 3 h was selected as the optimal concentration and
time and used in the following experiments (Fig. 1B). Cell viability was measured
after incubation with various concentrations of curcumin (0–200 µM)
for different durations (3–24 h). Concentrations of >50 µM had
an obvious toxic effect, with longer treatments leading to higher
toxicity (Fig. 1C). Similarly, 25
µM curcumin was used to incubate HUVECs for different durations
(3–24 h) to determine cell viability following pretreatment with
H2O2 for 3 h. Curcumin treatment restored
H2O2-induced cell damage after 3 h. However,
longer treatments did not increase cell viability (Fig. 1D). Therefore, treatment with 25 µM
curcumin for 3 h was selected as the optimal concentration and time
and was used in subsequent experiments.
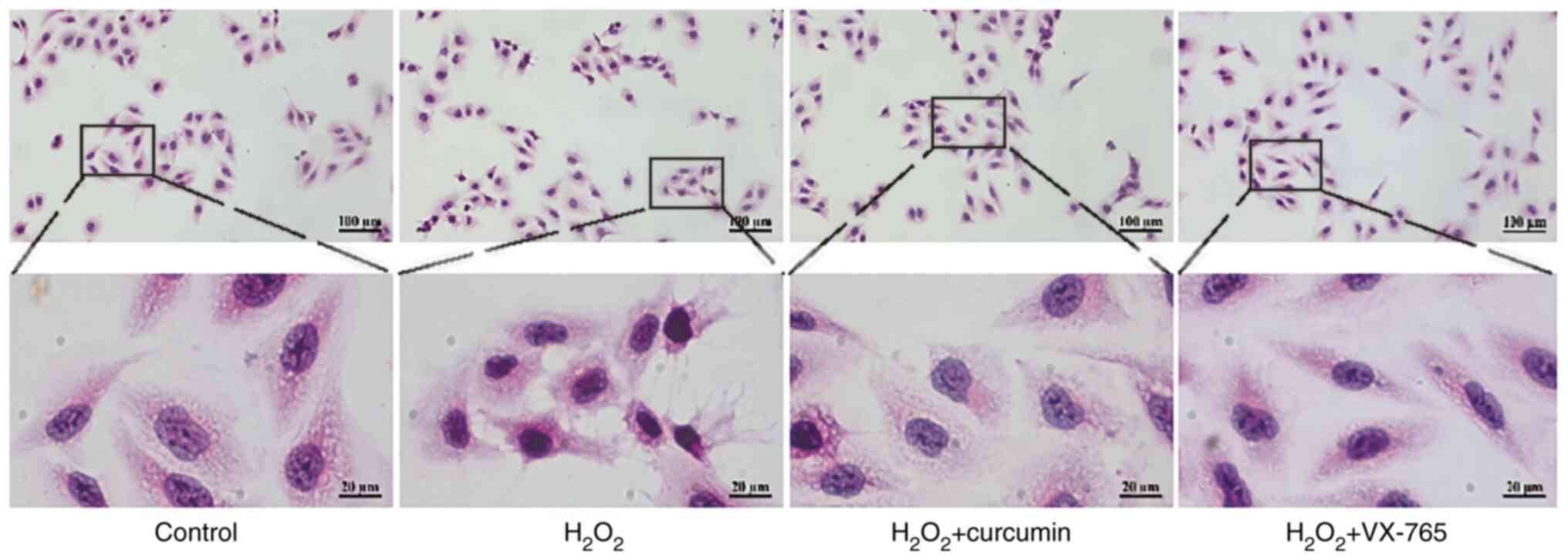
To determine the role of curcumin during
H2O2-induced HUVEC injury, H&E staining
was performed on HUVECs. Cells were divided into the control,
H2O2, curcumin and VX-765 pre-treatment
groups. The results indicated that H2O2
treatment led to rupture of the cell membrane of HUVECs and
karyopyknosis (Fig. 2). In
comparison, curcumin reduced H2O2-induced
HUVEC damage.
Curcumin suppresses
H2O2-induced HUVEC pyroptosis
Pyroptosis is uniquely dependent on the activation
of caspase-1, which is able to process cytokines IL-1β and IL-18
into their active forms and then induce pyroptotic cell death. To
elucidate the association between curcumin and
H2O2-induced pyroptosis, a variety of
experiments were performed. Cells were divided into a control,
H2O2, curcumin and VX-765 pre-treatment
groups. Western blot analysis revealed that caspase-1, GSDMD and
IL-1β were all activated and enhanced in
H2O2-treated HUVECs (Fig. 3A). Caspase-1/TUNEL double staining
was performed in HUVECs (Fig.
3B), indicating that the number of caspase-1 and TUNEL-positive
cells were both markedly increased in the presence of
H2O2 in HUVECs, and the number of caspase-1
and TUNEL-positive cells were both markedly decreased in the
presence of curcumin in HUVECs. Treatment of HUVECs with
H2O2 significantly increased the release of
LDH (Fig. 3C) and the proportion
of cells with PI-positive staining was markedly improved (Fig. 3D). However, this phenomenon was
counteracted by curcumin treatment. Similar changes in IL-1β and
IL-18 expression were observed in the H2O2-,
curcumin- and VX-765-treated and untreated HUVECs (Fig. 3E and F). The results indicated
that the selective inhibitor of caspase-1 (VX-765) reduced the
level of activated caspase-1 and inhibited the maturation of GSDMD
and IL-1β. This indicated that the
H2O2-induced pyroptosis of HUVECs was
dependent on caspase-1.
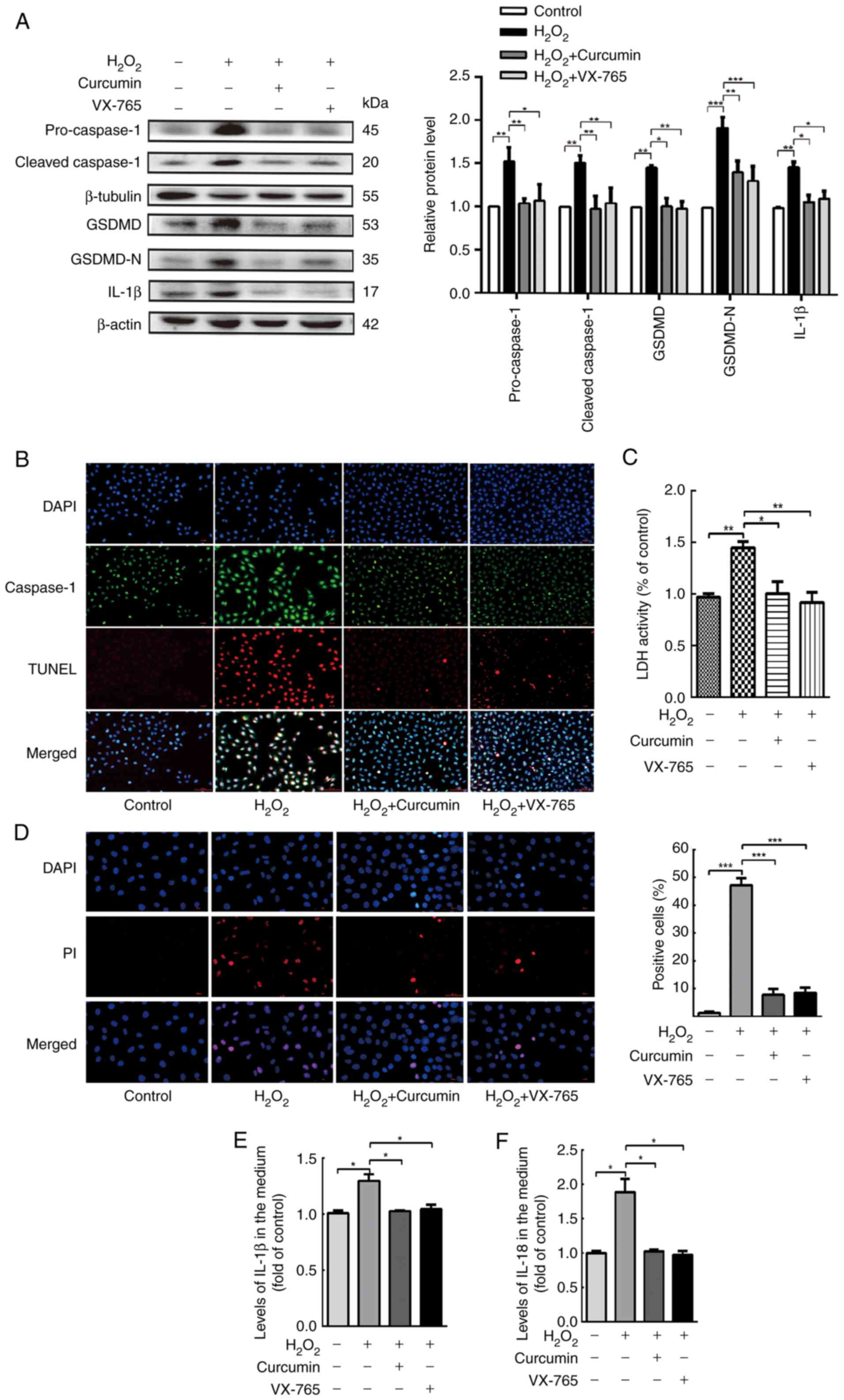 | Figure 3.Curcumin suppresses
H2O2-induced pyroptosis of HUVECs. HUVECs
were treated with H2O2 for 3 h and then
either treated with 25 µM curcumin for 3 h or left untreated
(control), whereas for the VX-765 group, HUVECs were pretreated
with caspase-1 inhibitor VX-765 (10 µM) for 1 h and then incubated
with H2O2 (800 µM) for 3 h. (A) Western blot
analysis revealed that the protein levels of pro-caspase-1,
caspase-1, pro-GSDMD, GSDMD and IL-1β were increased in HUVECs
following treatment with H2O2 for 3 h. VX-765
and curcumin inhibited the protein expression of pro-caspase-1,
caspase-1, pro-GSDMD, GSDMD and IL-1β. β-actin or β-tubulin was
used as an internal control. (B) Compared with the
H2O2 group, caspase-1 (green) and TUNEL (red)
double-positive cells were decreased in the presence of curcumin or
VX-765. The nuclei were stained blue with DAPI (magnification,
×400; scale bar, 50 µm). (C) The relative release of LDH in
H2O2-treated HUVECs was increased, while that
in curcumin- and VX-765-treated HUVECs was decreased (n=3). (D) The
percentage of PI-positive HUVECs (red) was increased following
H2O2 treatment, while it decreased following
curcumin and VX-765 treatment (left, representative images; right,
quantification of PI-positive cells; magnification, ×400; scale
bar, 50 µm). Relative concentration of (E) IL-1β and (F) IL-18 in
the culture medium following the treatment of HUVECs with
H2O2, curcumin or VX-765, as determined by
ELISA. Values are expressed as the mean ± standard deviation from
three independent experiments. *P<0.05, **P<0.01 and
***P<0.001 as indicated. HUVECs, human umbilical vein
endothelial cells; GSDMD, gasdermin D; LDH, lactate dehydrogenase;
PI, propidium iodide. |
Curcumin alleviates NLRP3 inflammasome
activation, which is involved in pyroptosis
Caspase-1 activation requires a protein complex
known as the inflammasome. The NLRP3 inflammasome is the most
extensively studied type of inflammasome. An increase in the
protein levels of cleaved caspase-1 and mature IL-1β/18 is a
hallmark of NLRP3 inflammasome activation. NLRP3 recruits caspase-1
through ASC, allowing activated caspase-1 to cleave pro-IL-1β and
pro-GSDMD, thus leading to the maturation of IL-1β and GSDMD.
Therefore, the expression levels of NLRP3, ASC, caspase-1, GSDMD
and IL-1β in HUVECs were measured following treatment with
curcumin. In addition, the NLRP3 inhibitor MCC950 was used
(4).
As presented in Fig.
4A, the inflammasome-associated protein levels following
treatment with H2O2 were measured using
western blot analysis. NLRP3 and ASC, as well as the core component
of pyroptosis, cleaved caspase-1, were markedly upregulated by
H2O2. Conversely, curcumin treatment
significantly blocked NLRP3 inflammasome activation. Furthermore,
curcumin and MCC950 weakened the ability of
H2O2 to induce HUVEC pyroptosis, as evidenced
by the decrease in the number of TUNEL and caspase-1
double-positive cells (Fig. 4B).
Curcumin and MCC950 also abrogated the release of LDH and the
increase in PI-positive cells, indicating inhibition of cell lysis
and pyroptotic cell death by curcumin (Fig. 4C and D). Similarly, curcumin and
MCC950 also significantly suppressed the expression of caspase-1
and the production of IL-1β and IL-18 (Fig. 4E and F), along with the inhibition
of caspase-1-dependent cell death.
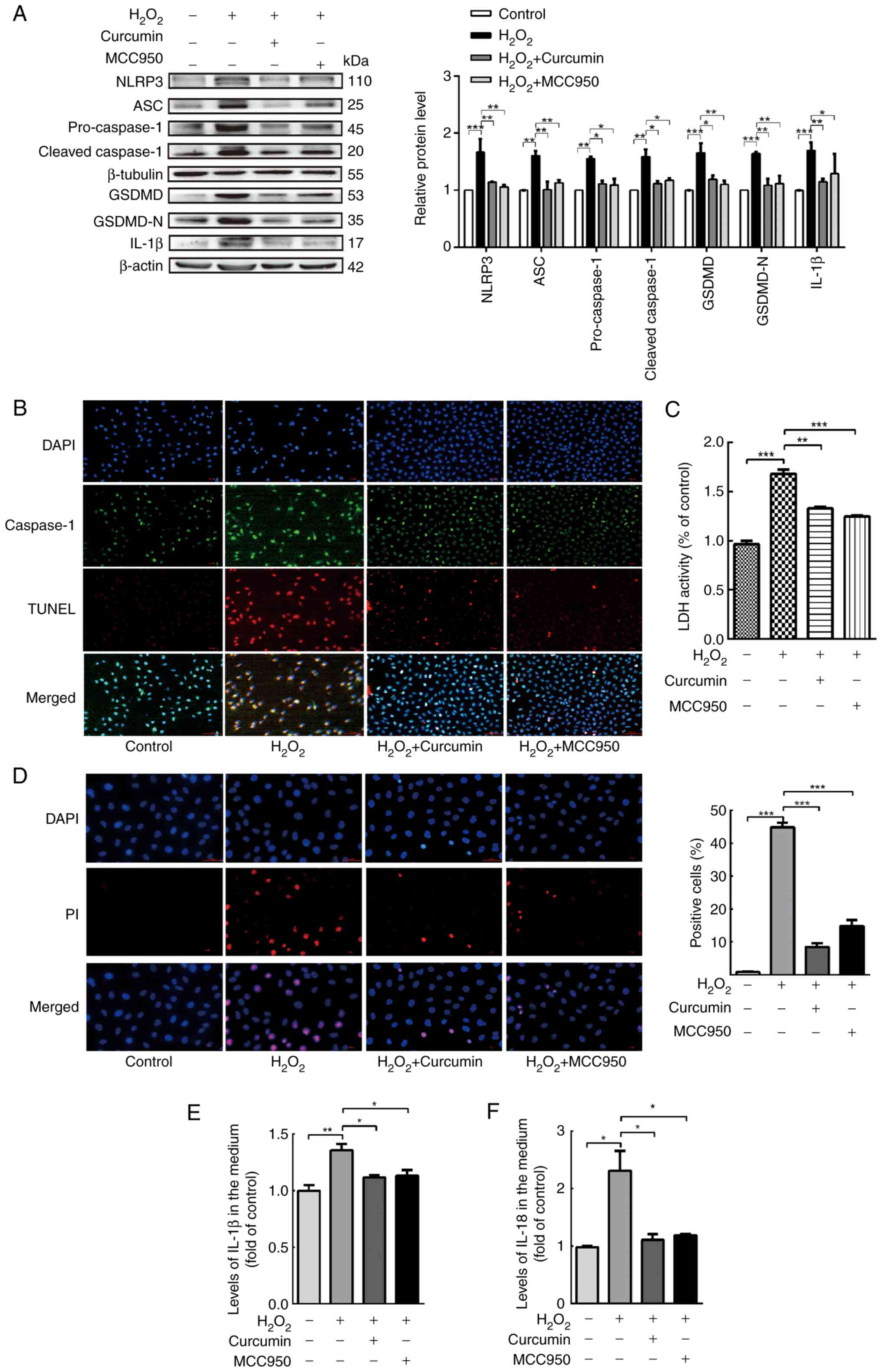 | Figure 4.Curcumin alleviates NLRP3
inflammasome activation, which is involved in pyroptosis. HUVECs
were treated with H2O2 for 3 h and then
either treated with 25 µM curcumin for 3 h or left untreated
(control), whereas for the MCC95 group, HUVECs were pretreated with
NLRP3 inhibitor (MCC950; 10 µM) for 2 h and then incubated with
H2O2 (800 µM) for 3 h. (A) Western blot
analysis indicated that the protein levels of NLRP3, ASC,
pro-caspase-1, caspase-1, pro-GSDMD, GSDMD and IL-1β were
upregulated in HUVECs following treatment with
H2O2 for 3 h and that MCC950 and curcumin
inhibited the protein expression of NLRP3, ASC, pro-caspase-1,
caspase-1, pro-GSDMD, GSDMD and IL-1β. β-Actin or β-tubulin was
used as an internal control. (B) As compared with the
H2O2 group, caspase-1 (green) and TUNEL (red)
double-positive cells were decreased in the presence of curcumin or
MCC950. The nuclei were stained blue with DAPI (magnification,
×400; scale bar, 50 µm). (C) The relative release of LDH by
H2O2-treated HUVECs was increased, while that
of curcumin- and MCC950-treated HUVECs was decreased (n=3). (D) The
percentage of PI (red)-positive H2O2-treated
HUVECs was increased, while that among curcumin- and MCC950-treated
cells was decreased (left, representative images; right,
quantification of PI-positive cells) (magnification, ×400; scale
bar, 50 µm). Relative concentration of (E) IL-1β and (F) IL-18 in
the culture medium following H2O2, curcumin
or MCC950 treatment of HUVECs, as determined by ELISA. Values are
expressed as the mean ± standard deviation of three independent
experiments. *P<0.05, **P<0.01, ***P<0.001. HUVECs, human
umbilical vein endothelial cells; NLRP3, NOD-, LRR- and pyrin
domain-containing protein 3; GSDMD, gasdermin D; ASC,
apoptosis-associated speck-like protein containing a caspase
recruitment domain; LDH, lactate dehydrogenase; PI, propidium
iodide. |
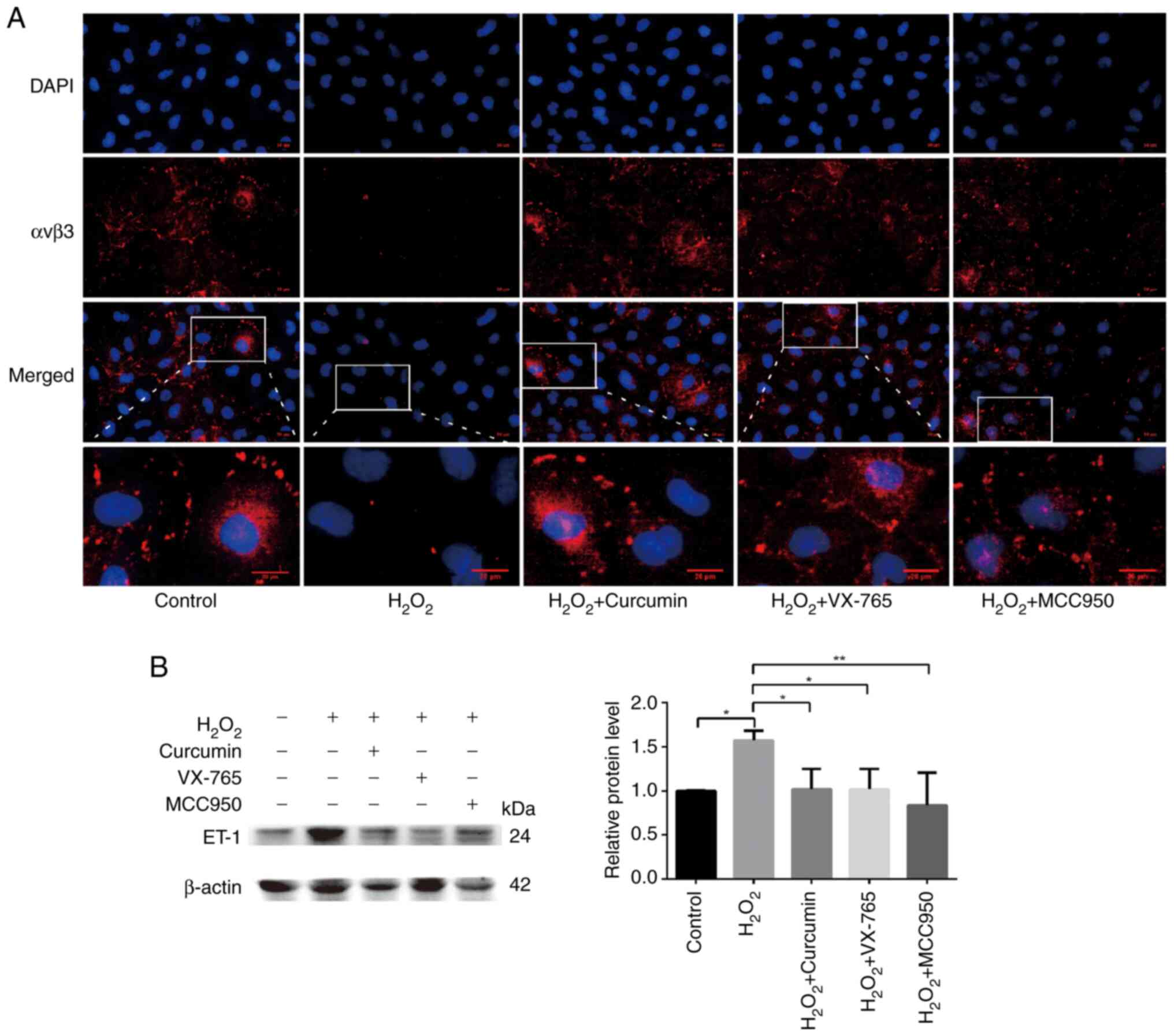
Curcumin improves
H2O2-induced functional damage of HUVECs
Endothelial cell adhesion molecules, including
integrin αvβ3 and E-selectin, are involved in angiogenesis
(18). A previous study reported
that decreased expression of αvβ3 in atherosclerotic mice led to
decreased recruitment of endothelial progenitor cells (EPCs)
(19); αvβ3 has an important role
in the EPC-led restoration of the biological function of ECs.
Endothelin (ET)-1 is a potent vasoconstrictor peptide produced and
released primarily by ECs. In addition to its vasoregulatory
properties, ET-1 overexpression is associated with the development
and progression of atherosclerosis and is generally considered to
be an atherogenic peptide (20,21). Therefore, immunofluorescence was
used to detect the expression levels of αvβ3 and western blot
analysis to quantify ET-1-related protein levels. Cells were
divided into the control, H2O2, curcumin,
VX-765 pre-treatment and MCC950 pre-treatment groups.
H2O2 significantly inhibited the expression
of αvβ3 (green), which was restored following the addition of
curcumin (Fig. 5A). At the same
time, high ET-1 expression was observed in the
H2O2 treatment group, while low ET-1
expression was present in the curcumin, inhibitor and control
groups (Fig. 5B). Curcumin was
thus further confirmed to improve
H2O2-induced cell damage.
Discussion
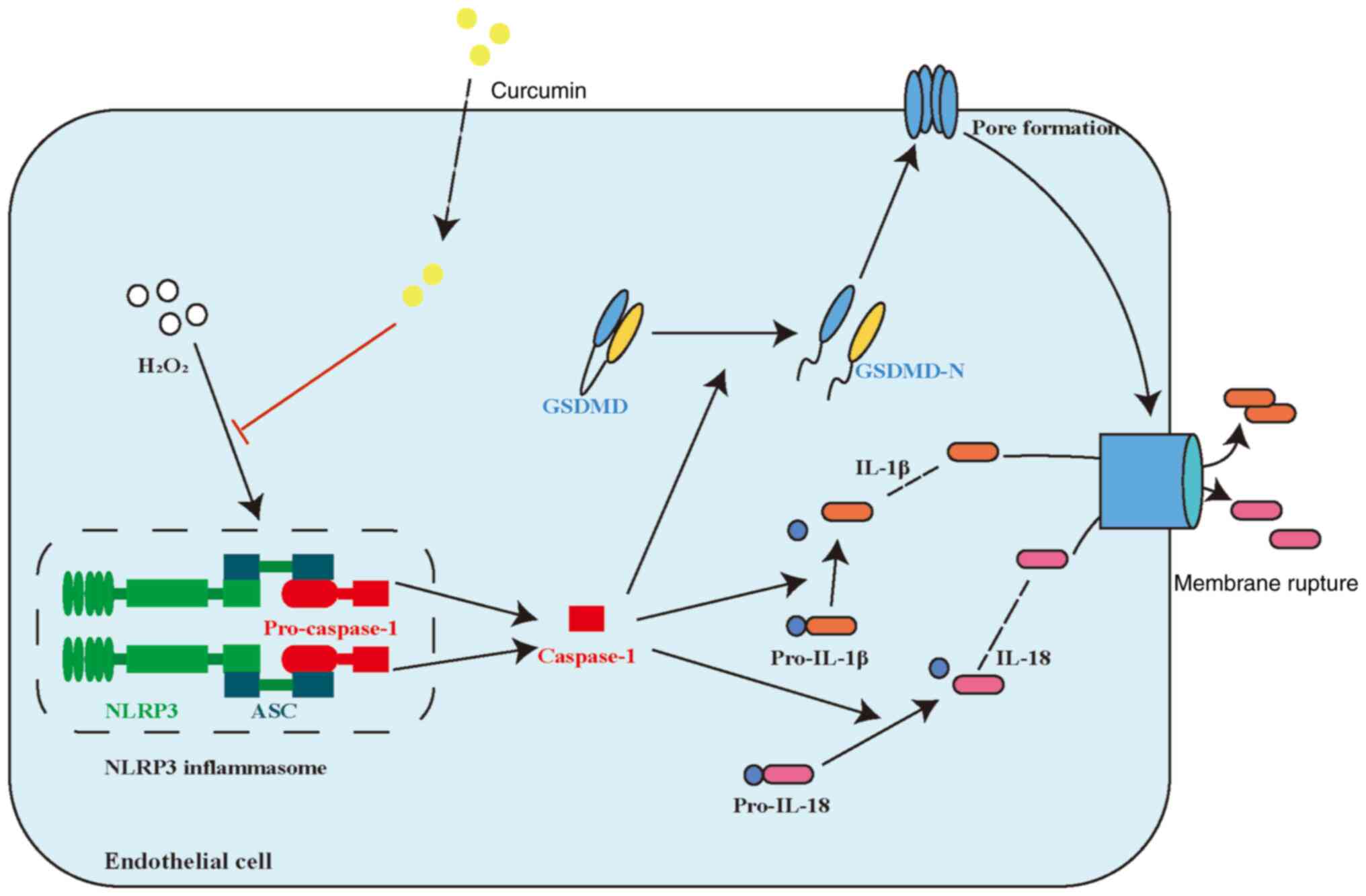
The results of the present study suggested that
curcumin has an important role in restoring
H2O2-induced pyroptosis in HUVECs. Curcumin
inhibited both the H2O2-induced and NLRP3
inflammasome activation pathways in HUVECs. Curcumin ameliorated
pyroptosis of HUVECs by inhibiting NLRP3 inflammasome activation.
These results suggested that curcumin may be a promising
therapeutic agent for AS. A schematic summary of the proposed
mechanism through which curcumin modulates the function of HUVECs
challenged with H2O2 is provided in Fig. 6.
Reactive oxygen species (ROS) are products of a
one-electron reduction of a type of oxygen in the body (22). H2O2 is able
to undergo a spontaneous conversion to OH through the Fenton
reaction (22). Vascular wall
cells are directly damaged by oxidative stress and EC damage is a
key driver of early AS (22).
Excessive accumulation of ROS in mitochondria is able to cause
programmed cell death (23).
Studies have indicated that ROS, as an important molecular upstream
regulator of the NLRP3 inflammasome, is able to activate the NLRP3
inflammasome (6,24). In the present study,
H2O2 was used to create a model of oxidative
damage in HUVECs and the effect of curcumin on the
H2O2-induced pyroptosis of HUVECs was
explored. The results suggested that curcumin significantly reduced
H2O2-induced pyroptosis, inflammasome
activation and release of inflammatory factors (including IL-1β and
IL-18). In addition, The AMP-activated protein kinase (AMPK)
pathway was reported to prevent and treat AS by promoting
cholesterol efflux, inhibiting inflammation and accelerating fatty
acid oxidation (25). Studies
have also indicated that the activation of the AMPK and sirtuin 1
(SIRT1) pathway not only inhibits processes of AS by inhibiting
oxidative stress and apoptosis in ECs, but also has an effective
protective role in a variety of inflammation-related diseases
(26,27). However, whether curcumin is able
to reduce oxidative stress by activating the MAPK/SIRT1 pathway
remains elusive and should be further studied.
Integrins not only mediate physical cell adhesion
but also initiate signaling events, alone or in combination with
growth factor receptor-mediated signals, to promote basic cell
functions such as cell migration, proliferation, survival and
differentiation. The integrin αvβ3 is able to bind to multiple
ligands in an Arg-Gly-Asp-dependent manner (28). It also enhances the biological
functions of ECs by mediating the binding of EPCs to ECs and
promotes the phosphorylation of VEGF receptor 2 and activation of
the ERK1/2/MAPK signaling pathway (29). In addition, tissue ET-1 levels are
significantly increased in the early and late stages of coronary
artery disease and are associated with its severity (30). A study indicated that ET-1 tissue
immunoreactivity predicts AS progression in patients with chronic
kidney disease (31). Consistent
with the findings of that study, the present results indicated that
curcumin restores αvβ3 expression and inhibits ET-1 overexpression.
Although ET-1-mediated EC dysfunction may be involved in the
development of AS (21), the
direct effects of ET-1 on EC function remain elusive.
Previous studies have demonstrated that curcumin has
multiple biological activities (15–17). In the present study, it was
confirmed that curcumin has significant anti-inflammatory and
anti-pyroptosis effects. However, although curcumin has a wide
range of potential beneficial pharmacological activities, numerous
questions regarding the fate of the compound in mammalian organisms
remain unanswered. Further studies are required to improve the
current understanding of curcumin and promote its application in
human diseases.
In conclusion, the present study proved for the
first time that curcumin is able to inhibit
H2O2-induced inflammation and pyroptosis in
HUVECs. Therefore, curcumin is a potential drug for the treatment
of AS. However, further studies are required to verify the
mechanisms through which curcumin inhibits NLRP3 to improve EC
pyroptosis. In addition, the role of curcumin in angiogenesis
mediated by EC adhesion molecule αvβ3, thereby stabilizing
atherosclerotic plaques and reducing adverse cardiovascular events,
remains to be further experimentally explored.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural Science
Foundation of Hunan Province (grant nos. 2018JJ2346 and
2018JJ2348).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YY and SL conceptualized the study. YY, CZ and SL
designed the study. YY, LY, QZ and YL carried out the experiments
and curated the data. YY, QZ, YH and YL analyzed the data. YY wrote
the manuscript. CZ and YH reviewed and edited the manuscript. YY
and SL checked and approved the authenticity of the raw data. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lusis AJ: Atherosclerosis. Nature.
407:233–241. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Welsh P, Grassia G, Botha S, Sattar N and
Maffia P: Targeting inflammation to reduce cardiovascular disease
risk: A realistic clinical prospect? Br J Pharmacol. 174:3898–3913.
2017. View Article : Google Scholar
|
|
3
|
Miao EA, Leaf IA, Treuting PM, Mao DP,
Dors M, Sarkar A, Warren SE, Wewers MD and Aderem A:
Caspase-1-induced pyroptosis is an innate immune effector mechanism
against intracellular bacteria. Nat Immunol. 11:1136–1142. 2010.
View Article : Google Scholar
|
|
4
|
Miao EA, Rajan JV and Aderem A:
Caspase-1-induced pyroptotic cell death. Immunol Rev. 243:206–214.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lin J, Shou X, Mao X, Dong J, Mohabeer N,
Kushwaha KK, Wang L, Su Y, Fang H and Li D: Oxidized low density
lipoprotein induced caspase-1 mediated pyroptotic cell death in
macrophages: implication in lesion instability? PLoS One.
8:e621482013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li N, Zhou H, Wu H, Wu Q, Duan M, Deng W
and Tang Q: STING-IRF3 contributes to lipopolysaccharide-induced
cardiac dysfunction, inflammation, apoptosis and pyroptosis by
activating NLRP3. Redox Biol. 24:1012152019. View Article : Google Scholar
|
|
7
|
Yang F, Qin Y, Lv J, Wang Y, Che H, Chen
X, Jiang Y, Li A, Sun X, Yue E, et al: Silencing long non-coding
RNA Kcnq1ot1 alleviates pyroptosis and fibrosis in diabetic
cardiomyopathy. Cell Death Dis. 9:10002018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen X, He WT, Hu L, Li J, Fang Y, Wang X,
Xu X, Wang Z, Huang K and Han J: Pyroptosis is driven by
non-selective gasdermin-D pore and its morphology is different from
MLKL channel-mediated necroptosis. Cell Res. 26:1007–1020. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mollace V, Gliozzi M, Musolino V, Carresi
C, Muscoli S, Mollace R, Tavernese A, Gratteri S, Palma E, Morabito
C, et al: Oxidized LDL attenuates protective autophagy and induces
apoptotic cell death of endothelial cells: Role of oxidative stress
and LOX-1 receptor expression. Int J Cardiol. 184:152–158. 2015.
View Article : Google Scholar
|
|
10
|
Jensen HA and Mehta JL: Endothelial cell
dysfunction as a novel therapeutic target in atherosclerosis.
Expert Rev Cardiovasc Ther. 14:1021–1033. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yin Y, Li X, Sha X, Xi H, Li YF, Shao Y,
Mai J, Virtue A, Lopez-Pastrana J, Meng S, et al: Early
hyperlipidemia promotes endothelial activation via a
caspase-1-sirtuin 1 pathway. Arterioscler Thromb Vasc Biol.
35:804–816. 2015. View Article : Google Scholar
|
|
12
|
Prasad S, Gupta SC, Tyagi AK and Aggarwal
BB: Curcumin, a component of golden spice: from bedside to bench
and back. Biotechnol Adv. 32:1053–1064. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yin H, Guo Q, Li X, Tang T, Li C, Wang H,
Sun Y, Feng Q, Ma C, Gao C, et al: Curcumin suppresses IL-1β
secretion and prevents inflammation through inhibition of the NLRP3
inflammasome. J Immunol. 200:2835–2846. 2018. View Article : Google Scholar
|
|
14
|
Saeedi-Boroujeni A, Mahmoudian-Sani MR,
Bahadoram M and Alghasi A: COVID-19: A case for inhibiting NLRP3
inflammasome, suppression of inflammation with curcumin? Basic Clin
Pharmacol Toxicol. 128:37–45. 2021. View Article : Google Scholar
|
|
15
|
Liang WF, Gong YX, Li HF, Sun FL, Li WL,
Chen DQ, Xie DP, Ren CX, Guo XY, Wang ZY, et al: Curcumin activates
ROS signaling to promote pyroptosis in hepatocellular carcinoma
HepG2 cells. In Vivo. 35:249–257. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kocaadam B and Sanlier N: Curcumin, an
active component of turmeric (Curcuma longa), and its
effects on health. Crit Rev Food Sci Nutr. 57:2889–2895. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Menon VP and Sudheer AR: Antioxidant and
anti-inflammatory properties of curcumin. Adv Exp Med Biol.
595:105–125. 2007. View Article : Google Scholar
|
|
18
|
Seguin J, Nicolazzi C, Mignet N, Scherman
D and Chabot GG: Vascular density and endothelial cell expression
of integrin alpha v beta 3 and E-selectin in murine tumours. Tumour
Biol. 33:1709–1717. 2012. View Article : Google Scholar
|
|
19
|
Filippi A, Constantin A, Alexandru N,
Voicu G, Constantinescu CA, Rebleanu D, Fenyo M, Simionescu D,
Simionescu A, Manduteanu I and Georgescu A: Integrins α4β1 and αVβ3
are reduced in endothelial progenitor cells from diabetic
dyslipidemic mice and may represent new targets for therapy in
aortic valve disease. Cell Transplant. 29:9636897209462772020.
View Article : Google Scholar
|
|
20
|
Brewster LM, Garcia VP, Levy MV,
Stockelman KA, Goulding A, DeSouza NM, Greiner JJ, Hijmans JG and
DeSouza CA: Endothelin-1-induced endothelial microvesicles impair
endothelial cell function. J Appl Physiol (1985). 128:1497–1505.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mathew V, Hasdai D and Lerman A: The role
of endothelin in coronary atherosclerosis. Mayo Clin Proc.
71:769–777. 1996. View Article : Google Scholar
|
|
22
|
Förstermann U, Xia N and Li H: Roles of
vascular oxidative stress and nitric oxide in the pathogenesis of
atherosclerosis. Circ Res. 120:713–735. 2017. View Article : Google Scholar
|
|
23
|
Schroder K and Tschopp J: The
inflammasomes. Cell. 140:821–832. 2010. View Article : Google Scholar
|
|
24
|
Zhao Y, Wang Z, Feng D, Zhao H, Lin M, Hu
Y, Zhang N, Lv L, Gao Z, Zhai X, et al: p66Shc contributes to liver
fibrosis through the regulation of mitochondrial reactive oxygen
species. Theranostics. 9:1510–1522. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang Q, Xu J, Ma Q, Liu Z, Sudhahar V, Cao
Y, Wang L, Zeng X, Zhou Y, Zhang M, et al: PRKAA1/AMPKα1-driven
glycolysis in endothelial cells exposed to disturbed flow protects
against atherosclerosis. Nat Commun. 9:46672018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chan SH, Hung CH, Shih JY, Chu PM, Cheng
YH, Lin HC, Hsieh PL and Tsai KL: Exercise intervention attenuates
hyperhomocysteinemia-induced aortic endothelial oxidative injury by
regulating SIRT1 through mitigating NADPH oxidase/LOX-1 signaling.
Redox Biol. 14:116–125. 2018. View Article : Google Scholar
|
|
27
|
Thornton CC, Al-Rashed F, Calay D, Birdsey
GM, Bauer A, Mylroie H, Morley BJ, Randi AM, Haskard DO, Boyle JJ
and Mason JC: Methotrexate-mediated activation of an
AMPK-CREB-dependent pathway: A novel mechanism for vascular
protection in chronic systemic inflammation. Ann Rheum Dis.
75:439–448. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Felding-Habermann B, Silletti S, Mei F,
Siu CH, Yip PM, Brooks PC, Cheresh DA, O'Toole TE, Ginsberg MH and
Montgomery AM: A single immunoglobulin-like domain of the human
neural cell adhesion molecule L1 supports adhesion by multiple
vascular and platelet integrins. J Cell Biol. 139:1567–1581. 1997.
View Article : Google Scholar
|
|
29
|
Hao D, Xiao W, Liu R, Kumar P, Li Y, Zhou
P, Guo F, Farmer DL, Lam KS, Wang F and Wang A: Discovery and
characterization of a potent and specific peptide ligand targeting
endothelial progenitor cells and endothelial cells for tissue
regeneration. ACS Chem Biol. 12:1075–1086. 2017. View Article : Google Scholar
|
|
30
|
Lerman A, Holmes DR Jr, Bell MR, Garratt
KN, Nishimura RA and Burnett JC Jr: Endothelin in coronary
endothelial dysfunction and early atherosclerosis in humans.
Circulation. 92:2426–2431. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Noshad H, Argani H, Nezami N, Ghojazadeh
M, Zomorrodi A, Bohlouli A, Bonyadi MR, Fakhrjou A, Ghorbanihaghjo
A, Gharedaghi A, et al: Arterial atherosclerosis in patients with
chronic kidney disease and its relationship with serum and tissue
endothelin-1. (corrected). Iran J Kidney Dis. 3:203–209.
2009.PubMed/NCBI
|