Introduction
Chronic obstructive pulmonary disease (COPD) is a
common respiratory disorder that causes the death of >3 million
individuals worldwide every year (1). COPD will be the third leading cause
of death in the world by 2030 (2). COPD is characterized by persistent
irreversible airflow limitation due to the loss of elastic fibers
from small airways and alveolar walls (3). The limitation is related to the
chronic inflammatory response of the respiratory tract and lungs to
harmful particles or gases (4).
There is increasing evidence that persistent airflow limitation is
closely related to airway remodeling, which occurs mainly in small
airways with an inner diameter of <2 mm (5–7).
Pulmonary fibroblasts are closely related to COPD
airway remodeling. Fibrosis-related factors produced by pulmonary
fibroblasts, such as collagen fibers, elastic fibers, reticular
fibers and constantly secreted matrix collagen, are increased
during COPD airway remodeling, which affects the
protease-antiproteinase balance (8). Various cytokines and inflammatory
mediators produced by fibroblasts also enhance the tissue
inflammation (9). Pulmonary
fibroblasts have been shown to contribute to airway remodeling in
COPD through synthesis and secretion of the main components of the
extracellular matrix (9).
Pulmonary fibroblasts are transformed into pulmonary myofibroblasts
activated by TGF-β1 (10,11). Myofibroblast differentiation,
characterized by ECM production and α-smooth muscle actin
expression, is one of the primary molecular mechanisms of airway
remodeling in COPD (12).
However, the mechanisms of fibroblasts in COPD remain poorly
understood.
Cell death may serve a vital role in the
pathogenesis of COPD (13).
Increased cell death can be observed during destruction of lung
tissue in both humans and mice (14,15). Pyroptosis, mediated by the
activation of caspase-1 and the inflammasome, is a unique form of
inflammatory cell death (16).
Activated caspase-1 proteolytically cleaves inactive pro-IL-1β and
pro-IL-18 into the mature inflammatory cytokines IL-1β and IL-18,
which induce the inflammasome activation-dependent pyroptotic cell
death, which results in the production of inflammatory chemokines
and TNF-α (17). Gasdermin D
(GSDMD), a substrate of caspase-1, has been identified as the
pyroptosis executioner (18).
Inflammasomes, important products of the inflammatory reaction,
serve a vital role in pyroptosis. NLR pyrin domain containing
protein 3 (NLRP3) is one of the most widely studied inflammasomes
and contains a caspase recruitment domain (19). Inflammasome activators, including
extracellular ATP, reactive oxygen species and damage-associated
molecular patterns, are also increased in the airways of patients
with COPD (20). Pyroptosis
induced by triggering receptor expressed on myeloid cells
1-mediated activation of the NLRP3 inflammasome aggravates COPD
development (21). In addition,
cigarette smoke extract-induced pyroptosis is also involved in the
progression of COPD (22). These
results suggest a potential critical role of pyroptosis in COPD
progression.
Long non-coding RNAs (lncRNAs) are noncoding RNAs
longer than 200 nucleotides with limited coding potential. They
serve crucial roles in various biological processes, including cell
apoptosis, invasion, proliferation, migration and therapeutic
responsiveness, by regulating gene expression (23). Importantly, lncRNAs drive multiple
important disease phenotypes in different ways (24), such as RNA decay, transcription
regulation, microRNA (miRNA/miR) sponging and epigenetic
modification (25). One of the
major mechanisms of lncRNAs is functioning as competing endogenous
RNAs (ceRNAs) in a regulatory network, including lncRNAs, miRNAs
and target mRNAs (26). ceRNAs
compete for miRNA binding and block miRNA-mediated target gene
silencing (27).
lncRNAs are widely involved in the pathogenesis of
COPD. The lncRNA small nuclear RNA host gene 5 (SNHG5) ameliorates
the effects of cigarette smoke extract on the proliferation,
apoptosis and inflammation of a COPD cell model via the
miR-132/PTEN axis in COPD (28),
while lncRNA MIR155HG promotes the polarization of M1 macrophage
and the release of pro-inflammatory cytokines in COPD (29). Furthermore, long intergenic
non-protein coding RNA 987 mitigates COPD by regulating
lipopolysaccharide (LPS)-caused cell apoptosis, oxidative stress,
inflammation and autophagy via the Let-7b-5p/sirtuin1 axis
(30). Growth arrest-specific 5
(GAS5), a well-documented lncRNA, is an important factor associated
with cell proliferation and death in various diseases (31,32). GAS5 acts as a ceRNA to sponge
miR-223-3p in certain diseases. For example, GAS5 promotes the
microglial inflammatory response in Parkinson's disease by
targeting the miR-223-3p/NLRP3 pathway, and low expression levels
of GAS5 injure myocardial cells and are associated with the
progression of chronic heart failure by regulating miR-223-3p
(33,34). Previous studies suggest that GAS5
serves a vital role in cell death (35–38). Nevertheless, it has not been
elucidated if lncRNA GAS5 is involved in cell death in COPD.
The present study investigated the effect of lncRNA
GAS5 on a cell model of COPD, as well as whether lncRNA GAS5 is
involved in pyroptosis and the underlying molecular mechanisms.
Materials and methods
Cell culture
The MRC-5 human lung fibroblast cell line (American
Type Culture Collection) was cultured in DMEM (cat. no. 12430054;
Thermo Fisher Scientific, Inc.) supplemented with 10% FBS (cat. no.
10100147; Thermo Fisher Scientific, Inc.) and 1%
penicillin/streptomycin (cat. no. 15070063; Thermo Fisher
Scientific, Inc.). The cells were grown in 75-cm2 flask
at 37°C in a humidified atmosphere with 5% CO2 and were
passaged 1:3 using 0.25% trypsin (cat. no. 1672868; Thermo Fisher
Scientific, Inc.) when they reached 80–90% confluence. LPS was
purchased from Sigma-Aldrich; Merck KGaA (cat. no. L2880). MRC-5
cells were stimulated with 10 µg/ml LPS for 48 h at 37°C (39) in the present study. Cells not
treated with LPS were used as Control.
Western blotting
Proteins were extracted from MRC-5 cells by RIPA
lysis buffer (Beyotime Institute of Biotechnology; cat. no. P0013B)
and quantified using BCA. Proteins (30 µg per lane) were separated
by SDS-PAGE on a 10% gel and subsequently transferred onto a PVDF
membrane, which were blocked with 5% skimmed milk for 1 h at room
temperature. The following primary antibodies were employed
overnight at 4°C: Rabbit anti-caspase-1 (1:500 dilution; cat. no.
BM4291; Boster Biological Technology), rabbit anti-pro-caspase-1
(1:1,000 dilution; cat. no. ab179515; Abcam), rabbit anti-IL-18
(1:500 dilution; cat. no. M00124; Boster Biological Technology),
rabbit anti-IL-1β (1:500 dilution; cat. no. A1112; ABclonal Biotech
Co., Ltd.), rabbit anti-cleaved N-terminal GSDMD (1:1,000 dilution;
cat. no. ab215203; Abcam) and mouse anti-GAPDH (1:5,000 dilution;
cat. no. A00227-1; Boster Biological Technology). Horseradish
peroxidase-conjugated goat anti-rabbit/mouse IgG (1:5,000 dilution;
cat. no. BM2002 and BA1070; Boster Biological Technology) was used
as a secondary antibody for 1 h at room temperature. Immunoreactive
protein bands were detected by ECL hypersensitive chemiluminescence
kit (cat. no. P0018M; Beyotime Institute of Biotechnology) with an
Odyssey Scanning System (version 3.0, LI-COR Biosciences).
The third-generation lentiviral vector
construction and lentiviral infection
pcDNA3.1 (+) GAS5 overexpression vector (OE-GAS5)
were designed and constructed based on human GAS5 full-length
coding protein sequences (GenBank accession number NC_000001) by
Shanghai GeneChem Co., Ltd. Then, 20 µM OE-GAS5 or overexpression
negative control lentivirus (NC) vectors were transfected into
MRC-5 cells with Lipofectamine® 3000 (Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. MRC-5
cells were transfected at 37°C for 8 h and were used in subsequent
experiments 48 h post-transfection.
Quantitative PCR (qPCR)
Total RNA was isolated from MRC-5 cells using
TRIzol® (cat. no. 15596026; Thermo Fisher Scientific,
Inc.). The concentration and quality of RNA were determined using
an ND-2000 Spectrophotometer. The extracted RNA was
reverse-transcribed into cDNA using a HiScript Reverse
Transcriptase kit (Vazyme Biotech Co., Ltd, China). Thermocycling
conditions for RT-PCR were 25°C for 5 min, 50°C for 45 min and 85°C
for 2 min, followed by holding at 4°C. quantitative PCR (qPCR) was
performed with the KAPA SYBR FAST qPCR kit (Kapa Biosystems) using
a SimpliAmp™ PCR System (Thermo Fisher Scientific, Inc.). The
thermocycling conditions were as follows: 95°C for 5 min, followed
by 40 cycles at 95°C for 10 sec, 60°C for 30 sec and 72°C for 30
sec. The primers for qPCR were designed using Primer 5.0 (Premier
Biosoft International). The primer pairs used were as follows:
GAPDH (gene accession no. AF275320) forward,
5′-TGACTTCAACAGCGACACCCA-3′ and reverse,
5′-CACCCTGTTGCTGTAGCCAAA-3′; GAS5 (gene accession no. NR_152533)
forward, 5′-GTGTCCCCAAGGAAGGATGA-3′ and reverse,
5′-GTAGTCAAGCCGACTCTCCA-3′; NLRP3 (gene accession no. AB120959)
forward, 5′-GCTGGCATCTGGATGAGGAA-3′ and reverse,
5′-GTGTGTCCTGAGCCATGGAA-3′; hsa-miR-223-3p-RT,
5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTGGGGT-3′;
hsa-miR-223-3p forward, 5′-TTGTCAGTTTGTCAA-3′ and reverse,
5′-CAGTGCAGGGTCCGAGGT-3′; hsa-miR-223-3p mimic,
5′-UGUCAGUUUGUCAAAUACCCC-3′; hsa-miR-223-3p inhibitor,
5′-GGGGUAUUUGACAAACUGACA-3′; hsa-miRNA mimic NC,
5′-UUUGUACUACACAAAAGUACUG-3′; hsa-miRNA inhibitor NC,
5′-CAGUACUUUUGUGUAGUACAAA-3′; hsa-U6 forward,
5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′.
GAPDH and small nuclear RNA U6 were used as internal references to
normalize the expression of mRNA and miRNA, respectively. The
samples were analyzed in triplicate. The data were quantified using
the 2−ΔΔCq method (40).
Immunofluorescence analyses
Immunofluorescence staining of MRC-5 cells was
performed as previously described (41). MRC-5 cells plated onto cover
glasses were fixed with 4% paraformaldehyde for 15 min and then
blocked in 5% BSA for 1 h both at room temperature. Fixed MRC-5
cells were incubated at 4°C overnight with rabbit anti-caspase-1,
followed by incubation with anti-rabbit IgG 594 secondary
antibodies (1:200 dilution; cat. no. ab150080; Abcam) for 1 h at
37°C. The images were acquired using a fluorescence microscope
(Nikon E800; Nikon Corporation) equipped with a digital camera
(1200F; Nikon Corporation) and image acquisition software
(NIS-Elements AR 3.2; Nikon Corporation). A total of three
independent samples in each group were included for quantitative
analysis in the present study.
Apoptosis analysis
MRC-5 cells were treated with 10 µg/ml LPS for 48 h.
The cells were subjected to a TUNEL assay according to the
manufacturer's instructions (Dead End Fluorometric TUNEL System;
Promega Corporation). Briefly, treated MRC-5 cells were fixed with
4% paraformaldehyde phosphate at 4°C for 10 min. Subsequently, the
cells were rinsed briefly with PBS, and then permeabilized with
0.1% Triton X-100 for 2 min on ice. Then, the cells were incubated
with TUNEL reagent in the dark for 1 h at 37°C. The cells were
rinsed with PBS and counterstained with 10 µg/ml DAPI at 37°C for 5
min. After the sections were sealed with neutral resin (cat. no.
96949-21-2; Sinopharm Chemical Reagent Co., Ltd.), fluorescence in
three random fields of view was measured using a fluorescence
microscope (Nikon E800; Nikon Corporation) equipped with a digital
camera (1200F; Nikon Corporation) and image acquisition software
(NIS-Elements AR 3.2, Nikon Corporation).
Cell Counting kit-8 (CCK-8)
MRC-5 cells were routinely cultured in 96-well
plates (4×104 cells/well) for 24 h. Subsequently, 10 µl
Cell Counting kit-8 (CCK-8) solution (Dojindo Laboratories, Inc.)
was added to the medium and the cells were incubated for 2 h at
37°C with 5% CO2. The optical density value was measured
at a wavelength of 450 nm with a Spectrafluor microreader plate
(Molecular Devices, LLC). These experiments were repeated three
times.
Bioinformatics analysis
StarBase database v2.0 (https://starbase.sysu.edu.cn/index.php/) was used to
predict the binding site of GAS5 and miR-223-3p. miR-223-3p and
NLRP3 target site was predicted using RNAhybrid v2.2 (https://bibiserv.cebitec.uni-bielefeld.de/rnahybrid).
Luciferase activity assay
The binding of GAS5 with miR-223-3p was validated
using dual-luciferase reporter assay. Hsa-miR-223-3p mimics and
hsa-miR-223-3p inhibitor were designed and synthesized by Shanghai
GenePharma Co., Ltd. as follows: Mimics,
5′-UGUCAGUUUGUCAAAUACCCC-3′; mimic NC,
5′-UUUGUACUACACAAAAGUACUG-3′; miR-223-3p inhibitor,
5′-GGGGUAUUUGACAAACUGACA-3′ and inhibitor NC,
5′-CAGUACUUUUGUGUAGUACAAA-3′. Wild-type (WT) and mutant (MUT) GAS5
or NLRP3 3′ untranslated regions were overexpressed and then cloned
into the luciferase reporter vector pmirGLO (Promega Corporation).
293T cells were cultured at 37°C overnight to 70–80% confluence,
then ~2×105 cells were seeded into 24-well plates. After
culturing for 24 h, 293T cells were co-transfected with luciferase
plasmids and miR-223-3p mimics using Lipofectamine® 3000
(Thermo Fisher Scientific, Inc.). After 48 h, luciferase activity
was determined using a Dual-Luciferase Reporter Assay kit (cat. no.
FR201-01; TransGen Biotech Co., Ltd.) by Eppendorf BioSpectrometer
fluorescence (BioPhotometer® D30; Eppendorf), and
firefly luciferase activity was normalized against Renilla
luciferase activity.
Lactate dehydrogenase (LDH) assay
LDH release was measured using an LDH Cytotoxicity
Assay kit (cat. no. C0017; Beyotime Institute of Biotechnology) as
previously described (42).
Supernatants from Control, NC and OE-GAS5 MRC-5 cells were
collected and 10% (vol/vol) LDH release reagent was added. LDH
release was measured at an optical density of 490 nm using the
Eppendorf BioSpectrometer fluorescence. Cytotoxicity was calculated
as % × (experimental LDH release-cell spontaneous LDH
release)/(maximum LDH release-cell spontaneous LDH release).
ELISA
Suitably treated MRC-5 cells were cultured for 24 h
in DMEM with 10% FBS. The supernatant was collected to detect the
levels of IL-2, IL-6, IL-10 and TNF-α using their respective kits
according to the manufacturer's instructions. IL-2 Human ELISA kit
(cat. no. EH2IL22), IL-6 Human ELISA kit (cat. no. EH2IL6), IL-10
Human ELISA kit (cat. no. EHIL10) and TNF-α Human ELISA kit (cat.
no. BMS223HS) were purchased from Thermo Fisher Scientific, Inc.
The calibration curves were plotted on semi-log papers, and the
optical density values of samples were calculated from the standard
curve for three assays.
RNA immunoprecipitation (RIP)
To validate the interaction between GAS5 and
miR-223-3p binding proteins, the Magna RIP kit (cat. no. 17–700;
Millipore) was used as previously described (43). MRC-5 cells (1×107 in
each group) were washed with cold PBS and harvested with cell
scrapers. MRC-5 cells were lysed using RIP Lysis Buffer (150 mM
KCl, 25 mM Tris pH 7.4, 5 mM EDTA, 0.5% NP40, 100 µl for per IP
reaction) supplemented with RNase and protease cocktail inhibitor,
followed by centrifugation at 12,000 × g and 4°C for 1 min. The
supernatants were collected and incubated with 5 µg Argonaute-2
antibody (1:1,000, cat. no. 233727; Abcam) or control IgG to
capture the RNAs used for qPCR. Magnetic beads for
immunoprecipitation were prepared according to the kit
instructions. The RNA binding protein-RNA complexes were
immunoprecipitated with premade magnetic beads at 4°C overnight
with agitation. Then the immunocomplex was precipitated using
protein G Dynabeads (40 µl for per IP reaction). After washing the
beads with ice-cold RIP Wash Buffer from the Magna RIP kit, the RNA
binding proteins were digested with proteinase K at 55°C for 30 min
with shaking. The purified RNA was isolated with TRIzol®
reagent (cat. no. 15596018; Thermo Fisher Scientific, Inc.) and
reverse transcribed, and the relative gene expression of GAS5 and
miR-223-3p was measured by qPCR, which was performed as described
above.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism software (8.0; GraphPad Software, Inc.). Each experiment was
repeated three times. Data are presented as the mean ± standard
deviation. Unpaired Student's t-test was used for two group
comparisons and one-way ANOVA followed by Duncan's post hoc test
was used for multiple comparisons. P<0.05 was considered to
indicate a statistically significant difference.
Results
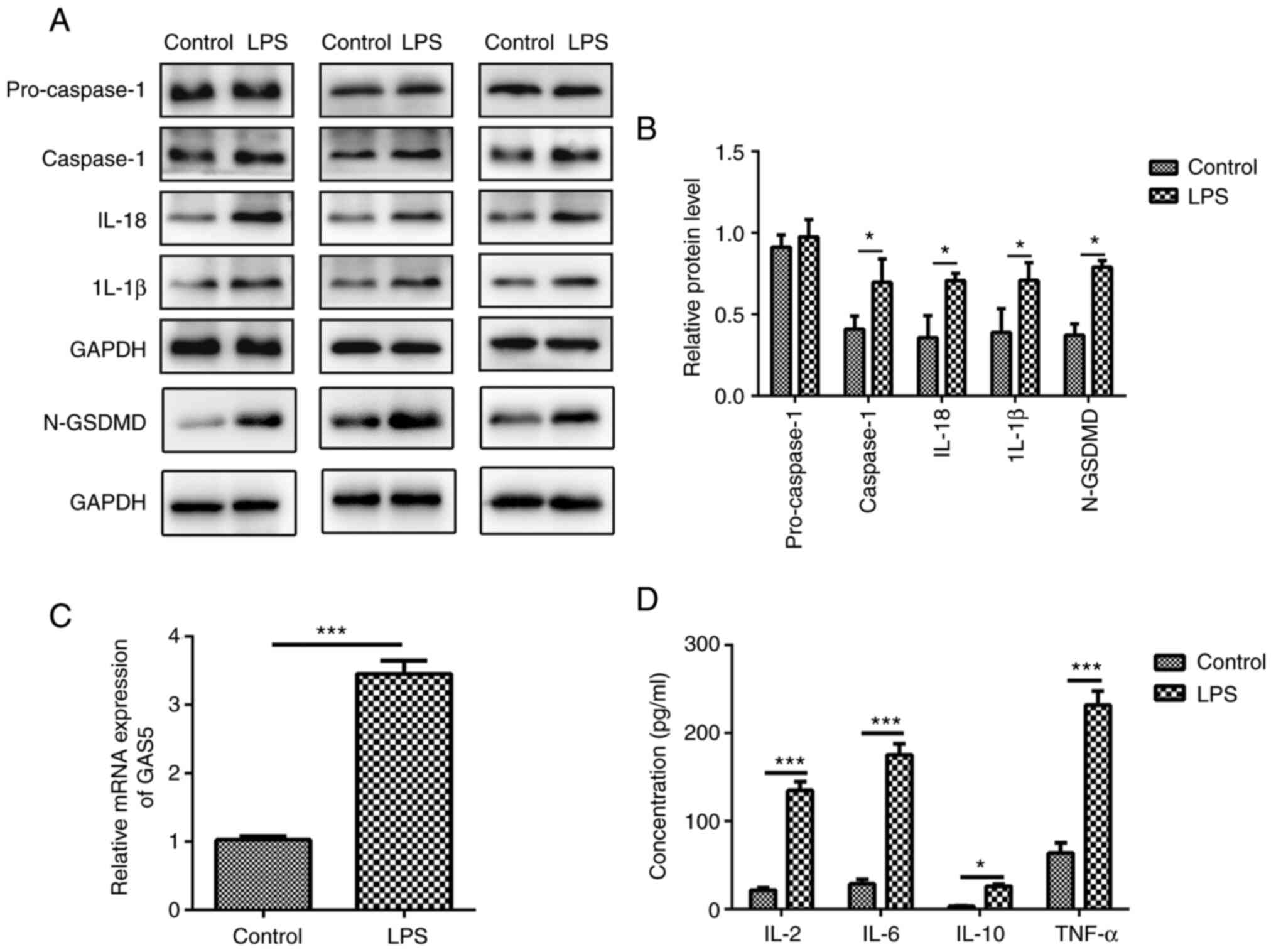
MRC-5 cells display characteristic
features of pyroptosis after LPS treatment
Pyroptosis is a unique form of inflammatory cell
death, which depends on the activation of caspase-1, IL-1β and
IL-18 (16). To elucidate the
relationship between COPD and pyroptosis, the present study
revealed that after 10 µg/ml LPS treatment, the expression levels
of caspase-1, IL-1β, IL-18 and cleaved N-terminal GSDMD were
increased in MRC-5 human lung fibroblast cells (Fig. 1A and B).
LPS induces GAS5 expression
lncRNA GAS5, a well-known tumor suppressor lncRNA,
aggravates apoptosis and inflammation in several human diseases,
including myocardial infarction and multiple sclerosis (7,44,45). However, whether lncRNA GAS5 is
involved in LPS-induced pyroptosis of MRC-5 cells is not fully
understood. The level of GAS5 was significantly increased
(3.31±0.48) after LPS treatment in MRC-5 cells compared with that
in the control group (1.05±0.09) (Fig. 1C; P<0.001), indicating that
GAS5 is involved in the pyroptosis of MRC-5 cells. Additionally,
LPS exposure markedly increased the levels of pro-inflammatory
factors IL-2, IL-6, IL-10 and TNF-α in MRC-5 cells (Fig. 1D). The results demonstrated that
lncRNA GAS5 might be associated with the release of inflammatory
factors that induce pyroptosis.
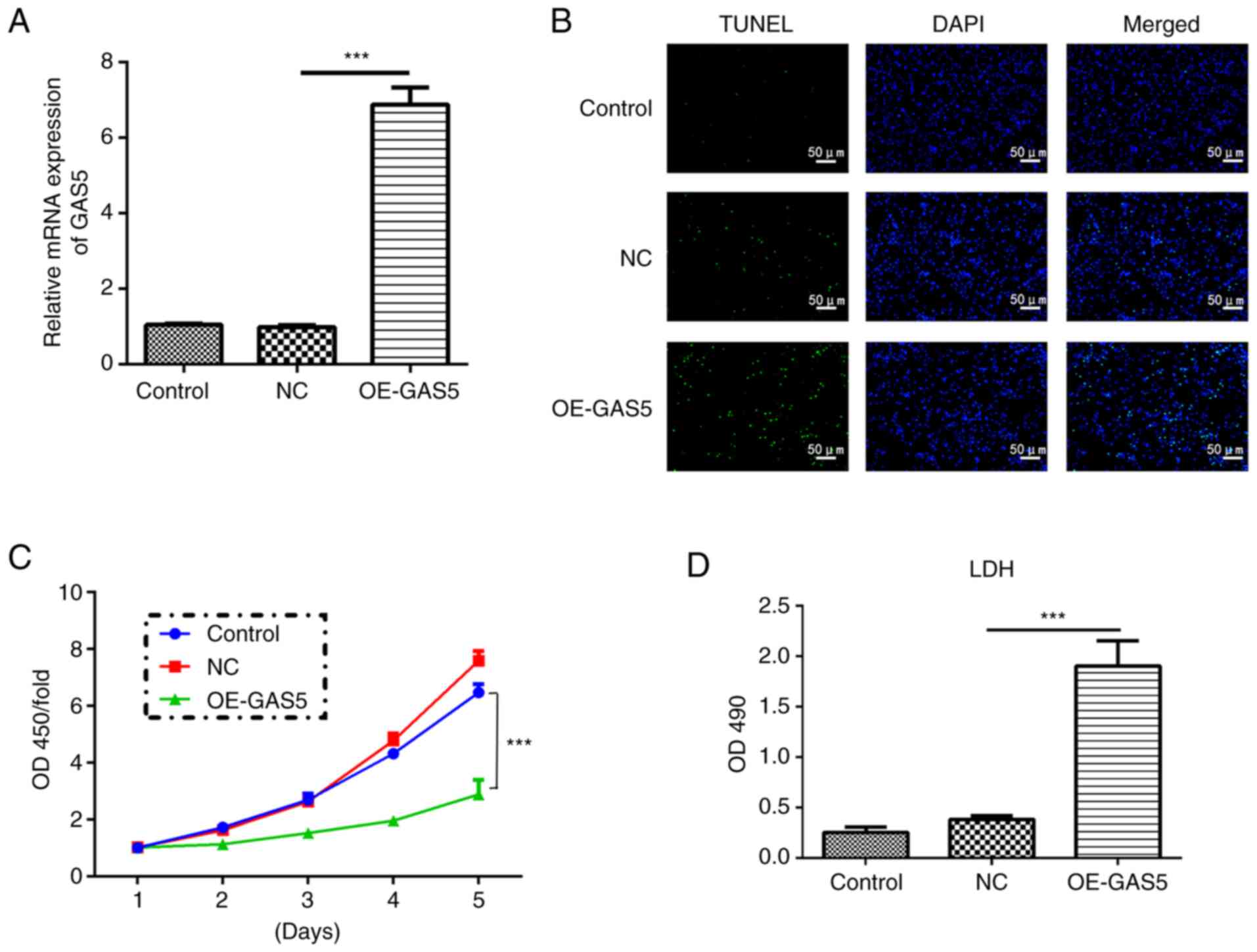
Upregulation of lncRNA GAS5 induces
pyroptosis
To explore the role of lncRNA GAS5 in pyroptosis,
lncRNA GAS5 was first overexpressed (OE-GAS5) and it was
demonstrated that the mRNA expression level of the lncRNA GAS5
group was 7-fold higher than that of the control and negative
control (NC) groups (Fig. 2A).
Upregulation of GAS5 efficiently induced cell death assessed by a
TUNEL assay, while it significantly inhibited cell proliferation at
the fifth day as examined by a CCK-8 assay in MRC-5 cells (Fig. 2B and C). In addition, highly
expressed GAS5 significantly enhanced the release of LDH, an
inflammatory response marker, suggesting an increased inflammatory
response (Fig. 2D).
Overexpression of lncRNA GAS5 significantly increased the protein
levels of caspase-1, IL-1β, IL-18 and cleaved N-terminal GSDMD
(Fig. 3A and B).
Immunofluorescence analyses revealed that upregulation of lncRNA
GAS5 promoted the expression of caspase-1 in MRC-5 cells (Fig. 3C). Furthermore, overexpression of
GAS5 significantly promoted the expression of the pro-inflammatory
factors IL-2, IL-6, IL-10 and TNF-α (Fig. 3D). Additionally, overexpression of
lncRNA GAS5 notably increased the level of NLRP3 (Fig. 3E), one of the key elements in the
NLRP3 inflammasome complex (17).
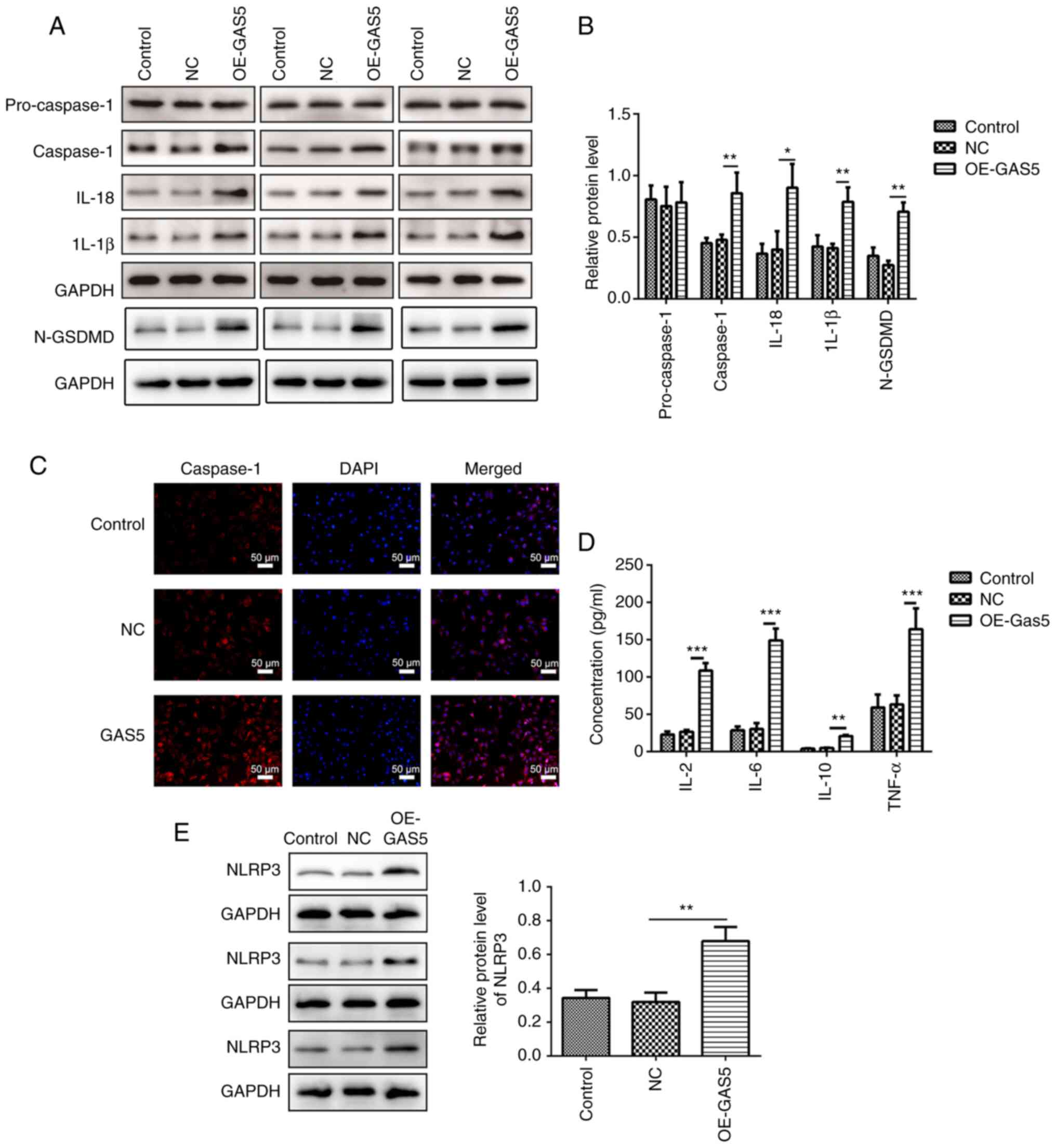 | Figure 3.Overexpression of lncRNA GAS5 induces
characteristic features of pyroptosis in MRC-5 cells. (A) Protein
expression levels of pro-caspase-1, caspase-1, IL-1β, IL-18 and
N-GSDMD in GAS5-overexpressing MRC-5 cells were measured using
western blotting and three sets of representative blots are shown.
(B) Semi-quantification of gene expression normalized to GAPDH. (C)
Immunofluorescence staining showing the levels of caspase-1 in
GAS5-overexpressing MRC-5 cells. Scale bar, 50 µm. (D) Quantitative
measurement of IL-2, IL-6, IL-10 and TNF-α in GAS5-overexpressing
MRC-5 cells using ELISA. (E) Protein expression levels of NLRP3 in
GAS5-overexpressing MRC-5 cells were measured using western
blotting. Semi-quantification of NLRP3 expression normalized to
GAPDH and three sets of representative blots are shown. The results
are presented as the mean ± SD (n=3; *P<0.05, **P<0.01,
***P<0.001 vs. control). N-GSDMD, cleaved N-terminal gasdermin
D; GAS5, growth arrest-specific 5; lncRNA, long non-coding RNA;
NLRP3, NLR family pyrin domain containing 3; NC, negative control;
OE, overexpression. |
GAS5 positively regulates NLRP3
expression by sponging miR-223-3p
To examine the mechanisms by which GAS5 positively
regulated NLRP3 expression, it was predicted that GAS5 could serve
as a miRNA sponge. The StarBase database identified that GAS5 can
bind to miR-223-3p (Fig. 4A). The
miRNA mimics and inhibitors of miR-223-3p were transfected into
MRC-5 cells to achieve ectopic miRNA expression, and the results
revealed that miR-223-3p expression was increased by miR-223-3p
mimics and suppressed by miR-223-3p inhibitors (Fig. 4B). Luciferase reporter assays were
used to detect whether GAS5 binds miR-223-3p directly. A decrease
in luciferase reporter activity was determined in 293T cells when
NLRP3 was co-transfected with miR-223-3p mimics, but not with the
negative control (Fig. 4C).
Subsequently, the RIP assay using an antibody against argonaute
RISC catalytic component 2 or IgG antibody showed that GAS5-WT
directly interacted with miR-223-3p (Fig. 4D).
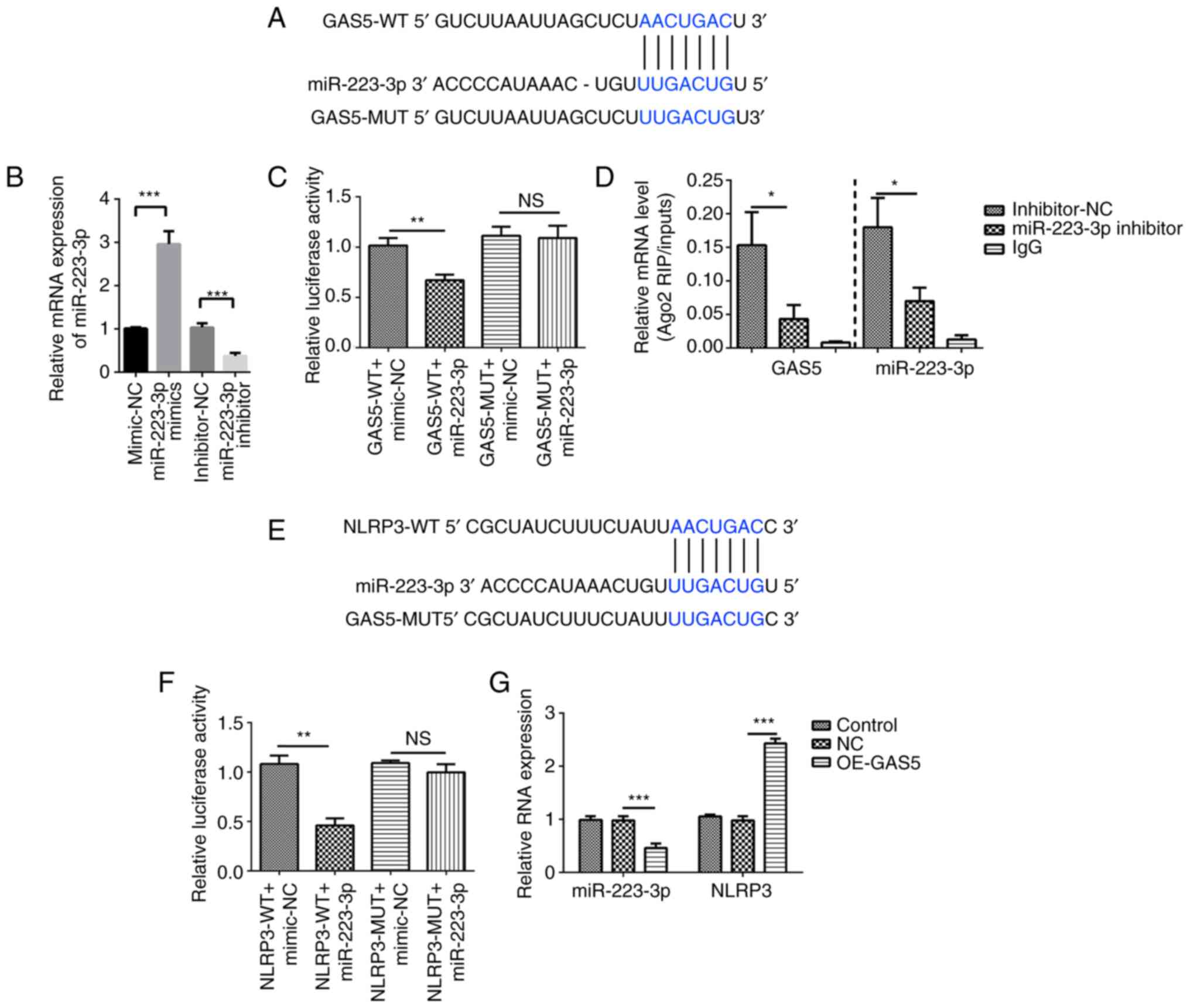 | Figure 4.GAS5 targets the miR-223-3p/NLRP3
axis. (A) Bioinformatics analysis was used to predict the binding
site of GAS5 to miR-223-3p. (B) Transfection efficiency of
miR-223-3p mimics and inhibitor was analyzed using qPCR. (C) GAS5
binds to the putative target sites of miR-223-3p as verified using
a luciferase assay. (D) RIP assay followed by qPCR was used to
determine GAS5 and miR-223-3p expression. (E) Bioinformatics
analysis was used to predict the binding site of miR-223-3p to
NLRP3. (F) miR-223-3p binds to the putative target sites of NLRP3
as verified using a luciferase assay. (G) Expression levels of
miR-223-3p and NLRP3 in GAS5-overexpressing MRC-5 cells were
measured using qPCR. The results are presented as the mean ± SD
(n=3; *P<0.05, **P<0.01, ***P<0.001 vs. control). GAS5,
growth arrest-specific 5; NLRP3, NLR family pyrin domain containing
3; miR-223-3p, microRNA 223-3p; 3′ UTR, 3′ untranslated region;
Ago2, argonaute RISC catalytic component 2; OE, overexpression; NC,
GAS5 overexpression negative control lentivirus vectors; RIP, RNA
immunoprecipitation; qPCR, quantitative PCR; WT, wild-type; MUT,
mutant; NS, no significant difference. |
Furthermore, RNAhybrid 2.2 showed that miR-223-3p
targets NLRP3 (Fig. 4E). A dual
luciferase reporter assay showed decreased luciferase activity in
293T cells co-transfected with miR-223-3p and NLRP3-WT compared
with that in cells co-transfected with miR-223-3p and NLRP3-MUT,
which verified the binding between NLRP3 and miR-223-3p. However,
the relative luciferase activity of the NLRP3-MUT co-transfected
with miR-223-3p group was not markedly changed compared with that
of the NLRP3-MUT co-transfected with miRNA-NC group (Fig. 4F). Furthermore, overexpression of
GAS5 significantly inhibited miR-223-3p expression and increased
that of NLRP3 as confirmed by qPCR (Fig. 4G). Thus, it was concluded that
GAS5 induced pyroptosis in MRC-5 cells by targeting
miR-223-3p/NLRP3.
Discussion
Aberrant expression of lncRNAs is involved in
several pulmonary disorders, revealing the association of lncRNAs
with the pathogenesis of these diseases (46). GAS5, a well-studied lncRNA, is
involved in the progression and prognosis of multiple cancers
(47–49). It has been demonstrated that GAS5
expression is notably increased in asthma to control the regulatory
T/T helper 17 cells balance and glucocorticoid activity, while it
is decreased in non-small cell lung cancer (NSCLC) to regulate the
PTEN/PI3K/AKT signaling pathway (50–52). Nonetheless, to the best of our
knowledge, no studies on the expression or function of GAS5 in COPD
have been published. In the present study, increased GAS5
expression was verified in COPD model cells. Furthermore, it was
observed that the overexpression of GAS5 induced MRC-5 cell death
that was caused by the activation of caspase-1 and inflammasomes.
Thus, it was concluded that GAS5 is involved in the regulation of
pyroptosis in COPD.
COPD is a multifactorial, multi-mechanical disease
with high morbidity and mortality (53), the pathogenesis of which involves
numerous pathophysiological processes, such as the inflammatory
response, oxidative stress, apoptosis, protease/antiproteinase
imbalance, production of autoantibodies, alteration in cell
proliferation, and cellular aging (54). Pyroptosis, characterized by DNA
damage and membrane rupture, is known as a unique form of
caspase-1-dependent cell death (16). Caspase-1 regulates the splicing of
inactive pro-IL-1β and pro-IL-18 into mature inflammatory cytokines
IL-1β and IL-18 to mediate pyroptosis (55). In the present study, it was
revealed that LPS treatment of MRC-5 cells significantly induced
caspase-1, IL-18 and IL-1β gene expression, indicating that
pyroptosis is involved in the development of COPD. However, the
specific mechanisms remain unclear.
GAS5, one of the most well-established
tumor-suppressive lncRNAs, is associated with cell death in
multiple human malignancies (35,56,57). GAS5 regulates cell growth arrest
and induces apoptosis through p53- or E2 factor 1-dependent
pathways in NSCLC (58). GAS5
promotes vascular smooth muscle cell growth arrest and apoptosis in
vascular remodeling (44).
Exosomal GAS5 silencing reduces the apoptosis of macrophages and
vascular endothelial cells in atherosclerosis (57). Overexpression of GAS5 decreases
the level of IL-18 and induces the apoptosis of fibroblast-like
synoviocytes (59). Additionally,
GAS5 overexpression decreases cell inflammatory responses and
apoptosis in LPS-treated MLE-12 cells (60). To the best of our knowledge, at
present, there is no evidence illustrating the exact role of GAS5
in COPD.
In the present study, it was revealed that GAS5
expression was significantly increased in LPS-treated MRC-5 cells,
suggesting that GAS5 might be associated with the pathogenesis of
COPD. At the same time, upregulation of GAS5 induced the release of
IL-2, IL-6, IL-10 and TNF-α, which is different from the function
of GAS5 in MLE-12 cells and fibroblast-like synoviocytes, in which
overexpression of GAS5 decreases cell inflammatory responses
(57,59). The aforementioned results
indicated that GAS5 served opposite roles in inflammatory responses
in different diseases. IL-10 exhibits diverse effects in the immune
response. On the one hand, IL-10 is an anti-inflammatory cytokine
that suppresses the expression or function of multiple inflammatory
cytokines, including IFN-γ, TNF-α and IL-6. On the other hand,
IL-10 can enhance immune events, such as immunoglobulin production
by B cells, the cytotoxicity of natural killer cells and
CD8+ T cells (61,62). An increased IL-10 concentration
has been identified in the blood samples of patients with COPD
(63), which might lead to
immunosuppression and reduce the inflammatory response in COPD
progression. Additionally, the present study also revealed that
overexpression of GAS5 promoted death and inhibited proliferation
by activating caspase-1, IL-1β, IL-18 and NLRP3 in MRC-5 cells.
Caspase-1 activation is mediated by inflammasomes, including NLRP3,
NLR family pyrin domain containing 1b (NLRP1b) and NLR family card
domain containing 4 (NLRC4) (64). In the present study, it was only
demonstrated that overexpression of GAS5 increased the level of
NLRP3 without further investigating if lncRNA GAS5 promotes
pyroptosis by regulating NLRP1b and NLRC4 inflammasomes, which
should be investigated in the future.
lncRNAs have been demonstrated to regulate gene
expression as ceRNAs (65). This
means lncRNAs can regulate the target gene expression by
competitively binding to the response elements within miRNA. For
example, lncRNA H19 promotes acute lymphoblastic leukemia by
sponging miR326 to reduce BCL-2 expression (66). Long intergenic non coding RNA 1184
increases the proliferation and invasion of colorectal cancer by
targeting the miR-331/HER2-p signaling pathway (67). Furthermore, ceRNA networks have
also been reported to regulate the progression of COPD. lncRNA
SNHG5, cancer susceptibility 2 and RP11-86H7.1 have all been
reported to serve as miRNA sponges and keep miRNA away from its
target genes through ceRNA networks to serve important roles in the
progression of COPD (28,68,69). However, to the best of our
knowledge, at present, no study has reported the role of GAS5 in
COPD. It needs to be explored whether GAS5 can serve as a ceRNA in
COPD. In the present study, a series of experiments demonstrated
that GAS5 could directly bind to miR-223-3p. At present, there is
only one report that miR-223-3p is increased in patients with COPD
(70). In contrast to this
conclusion, the present results are in line with existing knowledge
suggesting that miR-223-3p acts as a tumor suppressor in lung
squamous cell carcinoma (71),
and colon (72), prostate
(73) and ovarian cancer
(74). This divergence may be due
to the difference of in vitro and in vivo
experimental environments and the influence of the qualitative
difference across cell types. Additionally, it was also revealed
that miR-223-3p could directly bind to NLRP3, which is in line with
previous reports regarding the role of miR-223-3p (75). Therefore, it was concluded that
GAS5 could regulate COPD progression by targeting the
miR-223-3p/NLRP3 axis.
These findings provide knowledge regarding the
pathogenesis of COPD. The present study provided evidence that
lncRNA GAS5 may function as a regulator of pyroptosis in COPD by
targeting the miR-223-3p/NLRP3 axis. GAS5 overexpression promoted
cell death via the regulation of caspase-1, IL-18 and IL-1β
expression in MRC-5 cells. To the best of our knowledge, the
present study was the first to validate the role of GAS5 during
COPD progression. However, the present study only upregulated
lncRNA GAS5 in in vitro experiment to support the above
conclusion. More evidence is required to validate the predictions
and conclusions of the present study, for example, in in
vivo as well as clinical studies.
There are certain limitations of the present study.
For example, the in vitro experiments were only performed in
MRC-5 cells, and thus, further experiments should be performed in
more cell lines to confirm the findings reported in the present
study. Additionally, LPS-treated MRC-5 cells can also be used as a
model for other diseases, such as pneumonia and acute respiratory
distress syndrome (46,76). Thus, the results in the present
study need to be verified in improved COPD models. Due to the
limited experimental conditions, the role and mechanisms of GAS5 in
COPD have not been confirmed in any animal model or human tissue
samples. Thus, there are still some deficiencies in the present
study, and a lot of work is required in the future.
In conclusion, the results of the present study
suggested that lncRNA GAS5 expression was significantly increased
in a COPD cell model. Overexpression of GAS5 promoted pyroptosis by
targeting miR-223-3p/NLRP3 in MRC-5 cells. These findings suggest
that GAS5 serves an important role in the progression of COPD.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural Science
Foundation of China (grant nos. 81660013 and 81860015).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YC and YD were responsible for the conception of the
present study. RM, YC and YD confirm the authenticity of all the
raw data. RM and JL conducted the experiments, and analyzed and
interpreted the data. JL performed the statistical analysis. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mouronte-Roibas C, Leiro-Fernandez V,
Fernandez-Villar A, Botana-Rial M, Ramos-Hernandez C and
Ruano-Ravina A: COPD, emphysema and the onset of lung cancer. A
systematic review. Cancer Lett. 382:240–244. 2016. View Article : Google Scholar
|
|
2
|
Lopez-Campos JL, Tan W and Soriano JB:
Global burden of COPD. Respirology. 21:14–23. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rabe KF and Watz H: Chronic obstructive
pulmonary disease. Lancet. 389:1931–1940. 2017. View Article : Google Scholar
|
|
4
|
Singh D, Agusti A, Anzueto A, Barnes PJ,
Bourbeau J, Celli BR, Criner GJ, Frith P, Halpin DMG, Han M, et al:
Global strategy for the diagnosis, management, and prevention of
chronic obstructive lung disease: The GOLD science committee report
2019. Eur Respir J. 53:19001642019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhou L, Tao Y, Li H, Niu Y, Li L, Kan H,
Xie J and Chen R: Acute effects of fine particulate matter
constituents on cardiopulmonary function in a panel of COPD
patients. Sci Total Environ. 770:1447532021. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hirano T, Matsunaga K, Oishi K, Doi K,
Harada M, Suizu J, Murakawa K, Chikumoto A, Ohteru Y, Matsuda K, et
al: Abundant TNF-LIGHT expression in the airways of patients with
asthma with persistent airflow limitation: Association with
nitrative and inflammatory profiles. Respir Investig. 59:651–660.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bettoncelli G, Blasi F, Brusasco V,
Centanni S, Corrado A, De Benedetto F, De Michele F, Di Maria GU,
Donner CF, Falcone F, et al: The clinical and integrated management
of COPD. Sarcoidosis Vasc Diffuse Lung Dis. 31 (Suppl 1):S3–S21.
2014.
|
|
8
|
Salazar LM and Herrera AM: Fibrotic
response of tissue remodeling in COPD. Lung. 189:101–109. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Krimmer DI, Burgess JK, Wooi TK, Black JL
and Oliver BG: Matrix proteins from smoke-exposed fibroblasts are
pro-proliferative. Am J Respir Cell Mol Biol. 46:34–39. 2012.
View Article : Google Scholar
|
|
10
|
Di T, Yang Y, Fu C, Zhang Z, Qin C, Sai X,
Liu J, Hu C, Zheng M, Wu Y and Bian T: Let-7 mediated airway
remodelling in chronic obstructive pulmonary disease via the
regulation of IL-6. Eur J Clin Invest. 51:e134252021. View Article : Google Scholar
|
|
11
|
Spanjer AI, Baarsma HA, Oostenbrink LM,
Jansen SR, Kuipers CC, Lindner M, Postma DS, Meurs H, Heijink IH,
Gosens R and Königshoff M: TGF-beta-induced profibrotic signaling
is regulated in part by the WNT receptor frizzled-8. FASEB J.
30:1823–1835. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Barnes PJ: Small airway fibrosis in COPD.
Int J Biochem Cell Biol. 116:1055982019. View Article : Google Scholar
|
|
13
|
Henson PM, Vandivier RW and Douglas IS:
Cell death, remodeling, and repair in chronic obstructive pulmonary
disease? Proc Am Thorac Soc. 3:713–717. 2006. View Article : Google Scholar
|
|
14
|
Conlon TM, John-Schuster G, Heide D,
Pfister D, Lehmann M, Hu Y, Ertuz Z, Lopez MA, Ansari M, Strunz M,
et al: Inhibition of LTbetaR signalling activates WNT-induced
regeneration in lung. Nature. 588:151–156. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li T, Fanning KV, Nyunoya T, Chen Y and
Zou C: Cigarette smoke extract induces airway epithelial cell death
via repressing PRMT6/AKT signaling. Aging (Albany NY).
12:24301–24317. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Riegman M, Sagie L, Galed C, Levin T,
Steinberg N, Dixon SJ, Wiesner U, Bradbury MS, Niethammer P,
Zaritsky A and Overholtzer M: Ferroptosis occurs through an osmotic
mechanism and propagates independently of cell rupture. Nat Cell
Biol. 22:1042–1048. 2020. View Article : Google Scholar
|
|
17
|
Cho SJ, Hong KS, Jeong JH, Lee M, Choi
AMK, Stout-Delgado HW and Moon JS: DROSHA-dependent AIM2
inflammasome activation contributes to lung inflammation during
idiopathic pulmonary fibrosis. Cells. 8:9382019. View Article : Google Scholar
|
|
18
|
Shi J, Gao W and Shao F: Pyroptosis:
Gasdermin-mediated programmed necrotic cell death. Trends Biochem
Sci. 42:245–254. 2017. View Article : Google Scholar
|
|
19
|
Cao R, Fang D, Wang J, Yu Y, Ye H, Kang P,
Li Z, Wang H and Gao Q: ALDH2 overexpression alleviates high
glucose-induced cardiotoxicity by inhibiting nlrp3 inflammasome
activation. J Diabetes Res. 2019:48579212019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Franklin BS, Bossaller L, De Nardo D,
Ratter JM, Stutz A, Engels G, Brenker C, Nordhoff M, Mirandola SR,
Al-Amoudi A, et al: The adaptor ASC has extracellular and
‘prionoid’ activities that propagate inflammation. Nat Immunol.
15:727–737. 2014. View
Article : Google Scholar
|
|
21
|
Wang L, Chen Q, Yu Q, Xiao J and Zhao H:
TREM-1 aggravates chronic obstructive pulmonary disease development
via activation NLRP3 inflammasome-mediated pyroptosis. Inflamm Res.
70:971–980. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang MY, Jiang YX, Yang YC, Liu JY, Huo
C, Ji XL and Qu YQ: Cigarette smoke extract induces pyroptosis in
human bronchial epithelial cells through the ROS/NLRP3/caspase-1
pathway. Life Sci. 269:1190902021. View Article : Google Scholar
|
|
23
|
Xue C, Chen C, Gu X and Li L: Progress and
assessment of lncRNA DGCR5 in malignant phenotype and immune
infiltration of human cancers. Am J Cancer Res. 11:1–13.
2021.PubMed/NCBI
|
|
24
|
Schmitt AM and Chang HY: Long noncoding
RNAs in cancer pathways. Cancer Cell. 29:452–463. 2016. View Article : Google Scholar
|
|
25
|
Peng WX, Koirala P and Mo YY:
LncRNA-mediated regulation of cell signaling in cancer. Oncogene.
36:5661–5667. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhong Y, Du Y, Yang X, Mo Y, Fan C, Xiong
F, Ren D, Ye X, Li C, Wang Y, et al: Circular RNAs function as
ceRNAs to regulate and control human cancer progression. Mol
Cancer. 17:792018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Song YX, Sun JX, Zhao JH, Yang YC, Shi JX,
Wu ZH, Chen XW, Gao P, Miao ZF and Wang ZN: Non-coding RNAs
participate in the regulatory network of CLDN4 via ceRNA mediated
miRNA evasion. Nat Commun. 8:2892017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shen Q, Zheng J, Wang X, Hu W and Jiang Y
and Jiang Y: LncRNA SNHG5 regulates cell apoptosis and inflammation
by miR-132/PTEN axis in COPD. Biomed Pharmacother. 126:1100162020.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li N, Liu Y and Cai J: LncRNA MIR155HG
regulates M1/M2 macrophage polarization in chronic obstructive
pulmonary disease. Biomed Pharmacother. 117:1090152019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang Y, Chen J, Chen W, Liu L, Dong M, Ji
J, Hu D and Zhang N: LINC00987 ameliorates COPD by regulating
LPS-induced cell apoptosis, oxidative stress, inflammation and
autophagy through Let-7b-5p/SIRT1 axis. Int J Chron Obstruct Pulmon
Dis. 15:3213–3225. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ye K, Wang S, Zhang H, Han H, Ma B and Nan
W: Long noncoding RNA GAS5 suppresses cell growth and
epithelial-mesenchymal transition in osteosarcoma by regulating the
miR-221/ARHI pathway. J Cell Biochem. 118:4772–4781. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lyu K, Xu Y, Yue H, Li Y, Zhao J, Chen L,
Wu J, Zhu X, Chai L, Li C, et al: Long noncoding RNA GAS5 acts as a
tumor suppressor in laryngeal squamous cell carcinoma via miR-21.
Cancer Manag Res. 11:8487–8498. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu W, Zhang L, Geng Y, Liu Y and Zhang N:
Long noncoding RNA GAS5 promotes microglial inflammatory response
in parkinson's disease by regulating NLRP3 pathway through sponging
miR-223-3p. Int Immunopharmacol. 85:1066142020. View Article : Google Scholar
|
|
34
|
Li G, Du P, Qiang X, Jin D, Liu H, Li B
and Guo J: Low-expressed GAS5 injure myocardial cells and
progression of chronic heart failure via regulation of miR-223-3P.
Exp Mol Pathol. 117:1045292020. View Article : Google Scholar
|
|
35
|
He X, Wang S, Li M, Zhong L, Zheng H, Sun
Y, Lai Y, Chen X, Wei G, Si X, et al: Long noncoding RNA GAS5
induces abdominal aortic aneurysm formation by promoting smooth
muscle apoptosis. Theranostics. 9:5558–5576. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mazar J, Rosado A, Shelley J, Marchica J
and Westmoreland TJ: The long non-coding RNA GAS5 differentially
regulates cell cycle arrest and apoptosis through activation of
BRCA1 and p53 in human neuroblastoma. Oncotarget. 8:6589–6607.
2017. View Article : Google Scholar
|
|
37
|
Li D and He S: MAGE3 and Survivin
activated dendritic cell immunotherapy for the treatment of
non-small cell lung cancer. Oncol Lett. 15:8777–8783. 2018.
|
|
38
|
Chen L, Yang H, Xiao Y, Tang X, Li Y, Han
Q, Fu J, Yang Y and Zhu Y: LncRNA GAS5 is a critical regulator of
metastasis phenotype of melanoma cells and inhibits tumor growth in
vivo. Onco Targets Ther. 9:4075–4087. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chu H, Qu X, Wang F, Chang J, Cheng R,
Song X, Chen T and Zhang G: MicroRNA-206 promotes
lipopolysaccharide-induced inflammation injury via regulation of
IRAK1 in MRC-5 cells. Int Immunopharmacol. 73:590–598. 2019.
View Article : Google Scholar
|
|
40
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liu C, Wang F, Wang B, Wu T, Wang Y, Huo
W, Zhang S, Su Y, Liu J, Liu Y and Yu J: Pseudolaric acid B induces
apoptosis in human rhabdomyosarcoma RD cells. Oncol Lett.
20:3582020. View Article : Google Scholar
|
|
42
|
Lian L, Xue J, Li W, Ren J, Tang F, Liu Y,
Xue F and Dai J: VscF in T3SS1 helps to translocate VPA0226 in
vibrio parahaemolyticus. Front Cell Infect Microbiol.
11:6524322021. View Article : Google Scholar
|
|
43
|
Stoll L, Rodriguez-Trejo A, Guay C, Brozzi
F, Bayazit MB, Gattesco S, Menoud V, Sobel J, Marques AC, Veno MT,
et al: A circular RNA generated from an intron of the insulin gene
controls insulin secretion. Nat Commun. 11:56112020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tang R, Mei X, Wang YC, Cui XB, Zhang G,
Li W and Chen SY: LncRNA GAS5 regulates vascular smooth muscle cell
cycle arrest and apoptosis via p53 pathway. Biochim Biophys Acta
Mol Basis Dis. 1865:2516–2525. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Xie MY and Hou LJ: LncRNA GAS5 aggravates
myocardial infarction by sponging miR-26b. Int J Cardiol.
331:2102021. View Article : Google Scholar
|
|
46
|
Tang X, Wang T, Qiu C, Zheng F, Xu J and
Zhong B: Long non-coding RNA (lncRNA) CRNDE regulated
lipopolysaccharides (LPS)-induced MRC-5 inflammation injury through
targeting MiR-141. Med Sci Monit. 26:e9209282020. View Article : Google Scholar
|
|
47
|
Ni W, Yao S, Zhou Y, Liu Y, Huang P, Zhou
A, Liu J, Che L and Li J: Long noncoding RNA GAS5 inhibits
progression of colorectal cancer by interacting with and triggering
YAP phosphorylation and degradation and is negatively regulated by
the m(6)A reader YTHDF3. Mol Cancer. 18:1432019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Filippova EA, Fridman MV, Burdennyy AM,
Loginov VI, Pronina IV, Lukina SS, Dmitriev AA and Braga EA: Long
noncoding RNA GAS5 in breast cancer: Epigenetic mechanisms and
biological functions. Int J Mol Sci. 22:68102021. View Article : Google Scholar
|
|
49
|
Yang X, Xie Z, Lei X and Gan R: Long
non-coding RNA GAS5 in human cancer. Oncol Lett. 20:2587–2594.
2020. View Article : Google Scholar
|
|
50
|
Qiu YY, Wu Y, Lin MJ, Bian T, Xiao YL and
Qin C: LncRNA-MEG3 functions as a competing endogenous RNA to
regulate Treg/Th17 balance in patients with asthma by targeting
microRNA-17/RORgammat. Biomed Pharmacother. 111:386–394. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Keenan CR, Schuliga MJ and Stewart AG:
Pro-inflammatory mediators increase levels of the noncoding RNA
GAS5 in airway smooth muscle and epithelial cells. Can J Physiol
Pharmacol. 93:203–206. 2015. View Article : Google Scholar
|
|
52
|
Cao L, Chen J, Ou B, Liu C, Zou Y and Chen
Q: GAS5 knockdown reduces the chemo-sensitivity of non-small cell
lung cancer (NSCLC) cell to cisplatin (DDP) through regulating
miR-21/PTEN axis. Biomed Pharmacother. 93:570–579. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yang Y, Jin X, Jiao X, Li J, Liang L, Ma
Y, Liu R and Li Z: Advances in pharmacological actions and
mechanisms of flavonoids from traditional chinese medicine in
treating chronic obstructive pulmonary disease. Evid Based
Complement Alternat Med. 2020:88711052020. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yao H and Rahman I: Current concepts on
oxidative/carbonyl stress, inflammation and epigenetics in
pathogenesis of chronic obstructive pulmonary disease. Toxicol Appl
Pharmacol. 254:72–85. 2011. View Article : Google Scholar
|
|
55
|
Li F, Xu D, Hou K, Gou X, Lv N, Fang W and
Li Y: Pretreatment of indobufen and aspirin and their combinations
with clopidogrel or ticagrelor alleviates inflammasome mediated
pyroptosis via inhibiting NF-κB/NLRP3 pathway in ischemic stroke. J
Neuroimmune Pharmacol. 16:835–853. 2021. View Article : Google Scholar
|
|
56
|
Fawzy MS, Toraih EA, Ageeli EA,
Al-Qahtanie SA, Hussein MH and Kandil E: Noncoding RNAs orchestrate
cell growth, death and drug resistance in renal cell carcinoma.
Epigenomics. 12:199–219. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Chen L, Yang W, Guo Y, Chen W, Zheng P,
Zeng J and Tong W: Exosomal lncRNA GAS5 regulates the apoptosis of
macrophages and vascular endothelial cells in atherosclerosis. PLoS
One. 12:e01854062017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Shi X, Sun M, Liu H, Yao Y, Kong R, Chen F
and Song Y: A critical role for the long non-coding RNA GAS5 in
proliferation and apoptosis in non-small-cell lung cancer. Mol
Carcinog. 54 (Suppl 1):E1–E12. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ma C, Wang W and Li P: LncRNA GAS5
overexpression downregulates IL-18 and induces the apoptosis of
fibroblast-like synoviocytes. Clin Rheumatol. 38:3275–3280. 2019.
View Article : Google Scholar
|
|
60
|
Li J and Liu S: LncRNA GAS5 suppresses
inflammatory responses and apoptosis of alveolar epithelial cells
by targeting miR-429/DUSP1. Exp Mol Pathol. 113:1043572020.
View Article : Google Scholar
|
|
61
|
Nagata K and Nishiyama C: IL-10 in mast
cell-mediated immune responses: Anti-inflammatory and
proinflammatory roles. Int J Mol Sci. 22:49722021. View Article : Google Scholar
|
|
62
|
Castellucci M, Rossato M, Calzetti F,
Tamassia N, Zeminian S, Cassatella MA and Bazzoni F: IL-10 disrupts
the Brd4-docking sites to inhibit LPS-induced CXCL8 and TNF-alpha
expression in monocytes: Implications for chronic obstructive
pulmonary disease. J Allergy Clin Immunol. 136:781–791. 2015.
View Article : Google Scholar
|
|
63
|
Wei B and Li CS: Changes in
Th1/Th2-producing cytokines during acute exacerbation chronic
obstructive pulmonary disease. J Int Med Res. 46:3890–3902. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
He Y, Hara H and Núñez G: Mechanism and
regulation of NLRP3 inflammasome activation. Trends Biochem Sci.
41:1012–1021. 2016. View Article : Google Scholar
|
|
65
|
Zheng W, Chu Q and Xu T: The long
noncoding RNA NARL regulates immune responses via microRNA-mediated
NOD1 downregulation in teleost fish. J Biol Chem. 296:1004142021.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Mofidi M, Rahgozar S and Pouyanrad S:
Increased level of long non coding RNA H19 is correlated with the
downregulation of miR-326 and BCL-2 genes in pediatric acute
lymphoblastic leukemia, a possible hallmark for leukemogenesis. Mol
Biol Rep. 48:1531–1538. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Sui YX, Zhao DL, Yu Y and Wang LC: The
role, function, and mechanism of long intergenic noncoding RNA1184
(linc01184) in colorectal cancer. Dis Markers. 2021:88979062021.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Liu P, Zhang H, Zeng H, Meng Y, Gao H,
Zhang M and Zhao L: LncRNA CASC2 is involved in the development of
chronic obstructive pulmonary disease via targeting miR-18a-5p/IGF1
axis. Ther Adv Respir Dis. 15:175346662110280722021. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Zhao J, Pu J, Hao B, Huang L, Chen J, Hong
W, Zhou Y, Li B and Ran P: LncRNA RP11-86H7.1 promotes airway
inflammation induced by TRAPM2.5 by acting as a ceRNA of miRNA-9-5p
to regulate NFKB1 in HBECS. Sci Rep. 10:115872020. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Roffel MP, Maes T, Brandsma CA, van den
Berge M, Vanaudenaerde BM, Joos GF, Brusselle GG, Heijink IH and
Bracke KR: MiR-223 is increased in lungs of patients with COPD and
modulates cigarette smoke-induced pulmonary inflammation. Am J
Physiol Lung Cell Mol Physiol. 321:L1091–L1104. 2021. View Article : Google Scholar
|
|
71
|
Luo P, Wang Q, Ye Y, Zhang J, Lu D, Cheng
L, Zhou H, Xie M and Wang B: MiR-223-3p functions as a tumor
suppressor in lung squamous cell carcinoma by miR-223-3p-mutant p53
regulatory feedback loop. J Exp Clin Cancer Res. 38:742019.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Chai B, Guo Y, Cui X, Liu J, Suo Y, Dou Z
and Li N: MiR-223-3p promotes the proliferation, invasion and
migration of colon cancer cells by negative regulating PRDM1. Am J
Transl Res. 11:4516–4523. 2019.PubMed/NCBI
|
|
73
|
Wei Y, Yang J, Yi L, Wang Y, Dong Z, Liu
Z, Ou-yang S, Wu H, Zhong Z, Yin Z, et al: MiR-223-3p targeting
SEPT6 promotes the biological behavior of prostate cancer. Sci Rep.
4:75462014. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Fang G, Liu J, Wang Q, Huang X, Yang R,
Pang Y and Yang M: MicroRNA-223-3p regulates ovarian cancer cell
proliferation and invasion by targeting SOX11 expression. Int J Mol
Sci. 18:12082017. View Article : Google Scholar
|
|
75
|
Long FQ, Kou CX, Li K, Wu J and Wang QQ:
MiR-223-3p inhibits rTp17-induced inflammasome activation and
pyroptosis by targeting NLRP3. J Cell Mol Med. 24:14405–14414.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Xie Y, Qian Y, Wang Y, Liu K and Li X:
Mechanical stretch and LPS affect the proliferation, extracellular
matrix remodeling and viscoelasticity of lung fibroblasts. Exp Ther
Med. 20:52020. View Article : Google Scholar : PubMed/NCBI
|