Introduction
Obesity is a complex multifactorial disease, defined
as excessive fat deposition in adipose tissue, which may be harmful
to health. The degree of obesity is measured by body mass index
(BMI). BMI is calculated using weight in kilograms divided by
height in meters squared. For adults, a BMI of 25.0-29.9
kg/m2 is defined as overweight and ≥30 kg/m2
as obese (1). The worldwide
prevalence of obesity tripled between 1975 and 2016. The World
Health Organization estimated obesity had a global prevalence rate
of 13 and 39% of people aged ≥18 years was overweight in 2016
(2). In Mexico, an increase in
the prevalence of overweight and obese subjects was reported in
different age groups in 2018 compared with 2016, while in adults
aged ≥20 years, the prevalence of overweight and obese subjects
were 73.0 and 30.5%, respectively (3).
Obesity is considered to be an epidemic worldwide.
This condition increases the risk of numerous types of disease,
including insulin resistance (IR), type 2 diabetes (T2D) and
cardiovascular disease (CVD). Obesity is associated with a
low-grade inflammation state where increased production and
secretion of inflammatory factors, such as TNFα, IL-1β and IL-6,
result in alteration of key steps in the insulin signaling pathway,
which leads to development of IR (4). In addition, the high fat content in
adipose tissue, which is characterized by increased palmitic acid
and lipopolysaccharides concentration, leads to mitochondrial
dysfunction, increased reactive oxygen species (ROS) production,
inflammation and therefore higher risk of IR (5,6).
Adipose tissue is responsible for production and
secretion of numerous adipokines, including leptin, adiponectin and
retinol-binding protein 4 (RBP4), which leads to development of
metabolic disorder such as IR, T2D, CVD, dyslipidemia and liver
steatosis (6,7). RBP4 is expressed in the liver and
adipose tissue. In the circulation, RBP4 is responsible for
transport of retinol from the liver to peripheral tissue where it
is metabolized to retinoic acid (RA). In addition, RBP4 induces
secretion of proinflammatory cytokines in macrophages and activates
antigen-presenting cells in adipose tissue (6,7).
RBP4 is an adipokine that is expressed and secreted in mature
adipocytes, as well as in adipocyte and preadipocyte cultures
(8). Epidemiological studies have
reported an association between high levels of RBP4 and development
of metabolic disease, including obesity, IR and T2D in humans
(9–12) and animals (13,14), suggesting the role of RBP4 as a
potential biomarker of inflammation and oxidative stress (OS). In
this context, the current review article analyzed the role of RBP4
in obesity and associated chronic alterations.
RBP4
RBP4 is a serum polypeptide of 201 amino acids and a
size of 21 kDa. It consists of an N-terminal loop, β-barrel core, α
helix and C-terminal loop. The β-barrel core is a structural part
that specifically hosts one molecule of retinol (15). By ensuring retention of this
hydrophobic metabolite in an aqueous medium, it allows retinol
transportation in blood (15).
This protein is encoded by the RBP4 gene located on
chromosome 10q23-24 in humans. It is synthesized primarily in the
liver and adipose tissue, and to a lesser extent in other organs,
such as lungs, kidney, testes, brain and retina (7).
The primary function of RBP4 is transport of
retinol, an active metabolite of vitamin A, from the liver to
target tissue, such as adipose tissue, the retina, brain,
epidermis, kidney, lung, sexual organs and inmune system cells
(15). Vitamin A is a fat-soluble
compound with key biological activity in maintenance of immune
function, maintenance of epithelial cell integrity, physiology of
vision, reproduction as well as lipid metabolism. The physiological
actions of vitamin A are mediated by its bioactive metabolite,
retinoic acid. The vitamin A is found as provitamin A in
carotenoids; these include β-carotene obtained from fruit and
vegetables. Vitamin A is also found as preformed retinoids in the
form of retinyl esters, these compounds include retinol (alcohol),
retinal (aldehyde), retinoic acid (irreversibly oxidized form of
retinol), which are derived from animal sources (16). Following intestinal absorption,
retinyl esters are transported in chylomicrons to the liver for
storage. When the body is deficient in vitamin A, retinyl esters
are hydrolyzed and released as retinol, which binds to RBP4 and is
secreted into the circulation (17).
In the circulation, the retinol-RBP4 complex binds
transthyretin (TTR) to form a tertiary retinol-RBP4-TTR complex.
Binding to TTR stabilizes the retinol-RBP4 complex, decreases RBP4
loss by renal filtration and allows RBP4 to be recycled following
retinol uptake into cells (7,18).
Retinol spontaneously dissociates from RBP4 due to its hydrophobic
nature. Retinol readily moves into cells by diffusion through the
plasma membrane (19).
Nevertheless, in certain tissue, such as retina, brain, spleen,
kidney, testis and adipose, the retinol-RBP4 complex is recognized
by vitamin A receptor stimulated by retinoic acid 6 (STRA6) that
transports retinol from binding protein RBP4 into cells (19,20). Intracellularly, phosphorylated
STRA6 associates directly with intracellular retinol-binding
protein (CRBP1), transferring extracellular retinol molecule from
RBP4 to intracellular CRBP1, which prevents retinol dissociation in
the aqueous medium of the cytosol. Following binding of CRBP1 to
retinol, the CRBP1 dissociates from STRA6 and delivers retinol to a
retinol-metabolizing enzyme, such as lecithinretinol
acyltransferase (19,21). This catalyzes transformation of
retinol to retinol esters, which are stored in the endoplasmic
reticulum (22). In the absence
of retinol, RBP4 is freely filtered by glomeruli due to its small
molecular size and partially reabsorbed in cells of the proximal
tubule, allowing its reuse (10).
Due to their chemical stability and hydrophobicity,
retinol esters serve an essential role in visual function, both for
color vision and adaptation in darkness. In the retina, retinol
esters are hydrolyzed, isomerized and oxidized by retinol isomerase
and 11-cis-retinol dehydrogenases to form 11-cis-retinal, which
binds to opsin protein to form the visual pigment rhodopsin
(22,23). The activated rhodopsin breaks down
in all-trans-retinal and opsin following exposure to a photon light
source. When a photon light source enters the retina it is absorbed
by rhodopsin; this energy produces the rupture of 11 cis-retinal,
causing its isomerization to its trans form and releasing opsin.
This isomerization induces generation of an electric signal in the
optic nerve, which is interpreted as vision (24,25).
In addition, retinol induces expression of multiple
genes involved in energy homeostasis, insulin response, fatty acid
metabolism and the gene that codes for RBP4. To accomplish this, it
must be converted to retinaldehyde by the enzyme retinol
dehydrogenase 10. Subsequently, retinaldehyde is oxidized to RA by
retinaldehyde dehydrogenases 1–3 (26–28). RA activates or suppresses gene
transcription via nuclear receptors, namely retinoic acid receptor
(RAR)α, RARβ and RARγ (28,29). In the absence of RA, RBP4
secretion is inhibited, which leads to its accumulation in the
endoplasmic reticulum until the RA is available (30,31).
RBP4 in obesity
In obesity, white adipose tissue secretes large
amounts of adipokines, including RBP4, which have local and
systemic effects (retinol homeostasis and transport) and contribute
to IR (15,32). The association between elevated
serum levels of RBP4 (analyzed by ELISA) and obesity/T2D was
reported by Yang et al (26) in the Chinese population in obese
patients with and without diabetes. However, no significant
association was noted between non-diabetic and diabetic groups,
suggesting that hyperglycemia was not associated with elevated
serum RBP4 levels (33). The
association between RBP4 and obesity and associated disease has
been investigated in humans and animal and cell culture models
(7,9,11,13,18,32–39). In a 10-year follow-up study of
3,445 Chinese school-age children (11–12 years), an association
between serum RBP4 levels and adverse cardiovascular risk,
metabolic syndrome (MetS), IR, hyperglycemia, hypertension and
hyperlipidemia was reported, suggesting that RBP4 may be an early
biomarker of MetS (11).
Moreover, elevated serum RBP4 concentrations, analyzed by ELISA,
were positively associated with BMI, waist-hip index, elevated
blood pressure (BP), triglycerides (TG), total cholesterol (TC)
(11) low-density lipoprotein
(LDL) (11,35), homeostatic model assessment of IR
(HOMA-IR) and elevated leptin levels. Furthermore, elevated RBP4
levels were associated with lower levels of high-density
lipoprotein (HDL) cholesterol, regardless of sex, age, puberty and
time of follow-up (11). Elevated
levels of RBP4 in serum are significantly associated with incidence
of MetS and levels of very low-density lipoprotein and LDL
(35).
Conversely, Korek et al (36) published a study of a Polish
population, which did not identify significant differences in serum
RBP4 concentration levels between obese (BMI: 32.05-39.51
kg/m2) and lean subjects (BMI: 21.14-24.21
kg/m2). However, a positive correlation was noted
between concentration of RBP4 in serum (analyzed by ELISA) and
levels of TG in obese and control groups (36). A study of obese Romanian children
reported that following adjustment for age, sex and BMI, blood
levels of RBP4 (analyzed by ELISA) were not associated with
characteristics of MetS (obesity, IR and hypercholesterolemia) body
composition (fat and muscle mass) or anthropometric profile
(height, BMI, abdominal circumference, tricipital skin-fold
thickness, waist circumference/height ratio) (32). Although controversial results have
been published, a correlation between elevated levels of RBP4 in
the blood and the incidence of IR, serum lipid levels, and
anthropometric parameters has been reported (9,11,32,35–39). Therefore, population studies
implementing different dietary plans have been performed to
evaluate the association between weight loss and concentration of
adipokines, including RBP4 (37,38,40). A study of Spanish women (age,
18–80 years) with obesity who consumed a calorie-restricted diet
with 20, 27 or 35% protein for 3 months aimed to evaluate weight
loss in these subjects; serum levels of RBP4 (analyzed by ELISA)
were decreased by 12.50, 3.56 and 17.50%, respectively. Women
following the 35% protein diet exhibited a 30% greater decrease in
RBP4 levels than those following the 20% protein diet (following
adjustment for weight loss). In addition, RBP4 levels were directly
associated with TG concentration. However, the association between
serum RBP4 and levels of other adipokines, such as leptin, was not
significant (37). Similar
results were demonstrated in an intervention study of obese Spanish
individuals (BMI ≥35 kg/m2) who were subjected to a very
low-calorie diet for 6 months (40). Dietary intervention induced a
significant decrease in serum levels of RBP4 and inflammatory
markers, such as TNFα and C-reactive protein (CRP), which suggested
attenuation of inflammation is associated with obesity (40).
Previous studies have yielded similar results
(38–40). A study of American women (age,
18–62 years) with obesity and MetS who followed a diet and physical
activity plan for two months to lose weight reported that serum
levels of RBP4, analyzed by ELISA, were significantly associated
with concentration of TC and TG (38). However, serum RBP4 levels were not
associated with changes in body composition (weight, BMI, waist,
trunk fat), suggesting that serum RBP4 levels were stable in this
study population (38). However,
a study of Japanese men (age, 48±2 years) who participated in a
physical training program for 3 months demonstrated that obese
individuals with physical training without calorie restriction
exhibited significantly decreased levels of cardiovascular risk
factors, such as TC, TG, LDL cholesterol, RBP4, leptin and IL-6
(39). Regression analysis
revealed that change in serum RBP4 levels were associated with
change in TG levels. This suggested that physical training
contributes to decrease in RBP4 levels, regardless of body mass and
caloric intake (39).
Other studies have shown a significant association
between elevated serum levels of RBP4 and hypertriglyceridemia
(11,36,41). Therefore, the association between
RBP4 and TG may be representative of the association between RBP4
and obesity. RBP4 increases expression levels of genes that promote
fatty acid synthesis in adipose tissue. This suggests that RBP4
expression is associated with visceral fat accumulation (42). Physiologically, high
concentrations of RBP4 increase transport of retinol in the blood,
which is catabolized to RA isomers in the liver. These isomers
serve as ligands for nuclear receptors, such as RAR and retinoid X
receptor (RXR), to regulate transcription of numerous genes,
including those associated with glucose and lipid metabolism
(37).
Furthermore, RXR forms heterodimers with nuclear
receptors, including liver X receptor (LXR), RAR and peroxisome
proliferator-activated receptors (PPARs), which serve key roles in
signal transduction. LXR is a key modulator of lipid metabolism and
inflammatory cell signaling and regulates cholesterol efflux in
macrophages. Cholesterol and its metabolites are natural ligands
that activate LXR. In addition, activation of stearoyl-coenzyme A
desaturase and elongase 6, which are expressed in adipogenic
tissue, including hepatic and adipose tissue, has been shown to
serve a key role in lipid metabolism. Both enzymes are regulated by
sterol regulatory element-binding protein 1 (SREBP1) (43). SREBP1 is a key transcription
factor involved in expression of lipogenic genes that is induced by
activation of LXR and RXR (43).
Increased lipid synthesis is associated with increased RBP4
secretion and synthesis. The latter process occurs in brown adipose
tissue, primarily via the RXR/PPAR heterodimer. Specifically, PPARγ
and PPARα are involved in this process; these are both activated by
cyclic AMP (cAMP) (44).
In addition, excessive synthesis of RBP4 in adipose
tissue contributes to the inflammatory process, which is
characteristic of obesity, via activation of Toll-like receptor
(TLR) 2 and the TLR4/myeloid differentiation factor 2 receptor
complex in macrophages (45).
TLRs possess an extracellular domain that recognizes
pathogen-associated molecular patterns, a transmembrane region and
a cytosolic toll/interleukin-1 receptor domain that interacts with
adaptor molecules, such as myeloid differentiation primary response
88 (MyD88) and TNFα receptor. These proteins induce activation of
JNK-dependent pathways and NF-κB by phosphorylating inhibitor of
NF-κB (IκB) kinase, which phosphorylates IκB for proteasomal
degradation (46). Following
degradation of IκB, NF-κB translocates to the nucleus and induces
expression levels of inflammatory mediators, such as TNFα, monocyte
chemotactic protein-1, INF-γ and IL-6, −1β, −2 and −12, that are
required for activation of macrophages and dendritic cells
(45). In adipocytes, TNFα
stimulates lipolysis (47). This
is associated with inflammation and serves a causal role in IR as
release of free fatty acids increases endogenous glucose production
and decreases muscle glucose uptake (48,49). The JNK pathway induces activation
of antigen-presenting cells, such as macrophages and dendritic
cells, resulting in proliferation of pro-inflammatory cluster of
differentiation 4 T cells, which promotes inflammation and systemic
IR (Fig. 1A and B) (6).
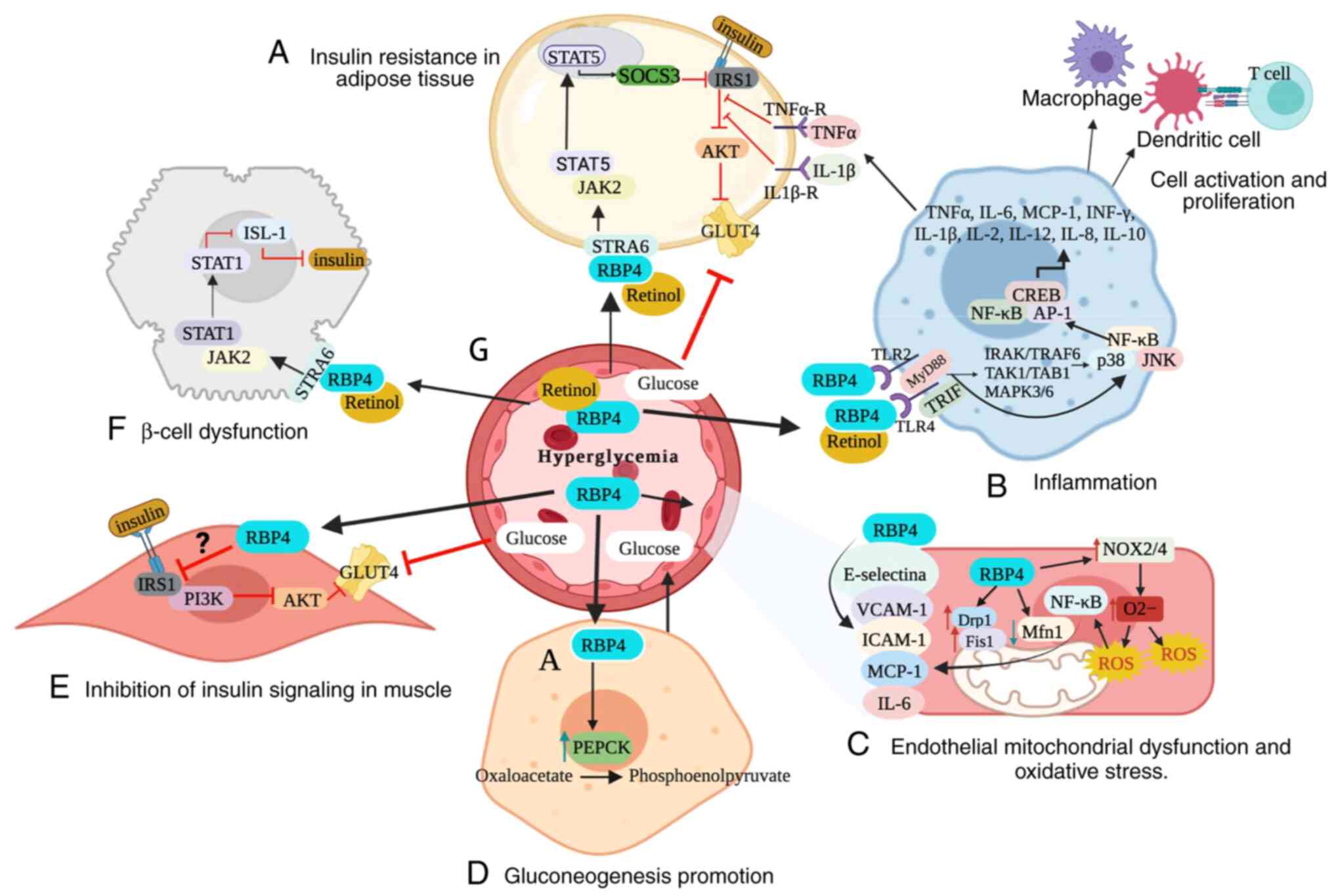 | Figure 1.Molecular mechanisms of RBP4 in
obesity and associated disease. (A) In adipocytes, the retinol-RBP4
complex directly inhibits insulin signaling by activating the
JAK2/STAT5/SOCS3 pathway, leading to IR. (B) In adipose tissue,
macrophages, retinol-RBP4 and RBP4 interact with TLR2 and TLR4/MD2
and downstream pathways of MyD88 and TRIF, releasing TNFα, IL-6,
MCP-1, INF-γ, IL-1β, IL-2, IL-12, IL-8 and IL-10. This results in
activation of the immune system and promotes an inflammatory state
as well as inhibition of insulin signaling. (C) RBP4 induces NF-κB
and NADPH oxidase-dependent endothelial inflammation, leading to
development of OS by mitochondrial dysfunction. (D) RBP4 increases
hepatic expression of PEPCK, thus increasing glucose production in
the liver. (E) In skeletal muscle, RBP4 is associated with high
serine 307 phosphorylation of IRS-1, which decreases its affinity
for PI3K and inhibits insulin signaling. (F) Retinol-RBP4 promotes
β-cell dysfunction via the JAK2/STAT1/ISL-1 pathway. (G)
Aforementioned mechanisms promote development of hyperglycemia.
PKB, Protein kinase B (PKB); AP-1, Activator protein 1; Drp1,
Dynamin-related protein 1; Fis1, Mitochondrial fission 1 protein;
GLUT 4, Glucose transporter type 4; ICAM-1, Intercellular Adhesion
Molecule 1; IRS1, Insulin receptor substrate-1; ISL-1, Insulin gene
enhancer protein; JAK2, Janus Kinase 2; MCP-1, Monocyte chemotactic
protein-1; MCP-1, Monocyte chemotactic protein-1; Mfn1,
Mitofusin-1; MyD88, Myeloid differentiation primary response 88;
NOX2, NADPH oxidase; PEPCK, Phosphoenolpyruvate carboxykinase;
RBP4, Retinol Binding Protein 4; ROS, reactive oxygen species;
SOCS3, Suppressor of cytokine signaling 3; TLR, Toll-like receptor;
TNFα-R, tumor necrosis factor α receptor; TRIF,
TIR-domain-containing adapter-inducing interferon-β; VCAM-1,
Vascular cell adhesion protein 1. |
In addition, activation of TLR causes activation of
the IL-1 receptor-associated kinase (IRAK) family (46). The IRAK1-IRAK2-IRAK4 complex binds
MyD88, thenIRAK4phosphorylates to IRAK1 at serine and threonine
residues, this event activates TNF-receptor-associated factor 6
(TRAF6), which subsequently activates TGF-β-activated protein
kinase 1 (TAK1). TAK1 forms a complex with TGF-β-activated kinase
1/MAP3K7 binding protein 1 (TAB1), TAB2 and TAB3 (46). Moreover, TAK1 activates MAPK3 and
MAPK6 to activate an alternative pathway that phosphorylates JNK
and p38 (46). In addition, RBP4
induces endothelial inflammation, which is dependent on NF-κB and
NADPH oxidase and is independent of retinol and STRA6. This
suggests that RBP4 exerts retinol-dependent and -independent
effects via activation of STRA6 and different receptors and
signaling pathways in different types of cell (50) (Fig.
1C).
RBP4 in IR
IR involves excessive secretion of adipokines,
including RBP4. RBP4 is considered to be involved in the
pathogenesis of obesity and development of IR. The retinol-RBP4
complex directly inhibit insulin signaling in adipocytes by
activating Janus kinase 2 (JAK2)/STAT5 signaling, which induces
suppressor of cytokine signaling (SOCS) 3 expression (an inhibitor
of insulin signaling); this mechanism is retinol-dependent and
requires the membrane receptor STRA6 (Fig. 1A) (42,51). In addition, the retinol-RBP4-TTR
complex is associated with development of IR in obese patients or
subjects with T2D (7,41). In a study of Indian adults of
similar age (36–46 years), ELISA was used to analyze serum levels
of RBP4 and TTR (52). The
subjects were stratified into three groups: Normal glucose
tolerance (NGT; n=90), impaired glucose tolerance (IGT; n=70) and
T2D (n=90). The mean serum RBP4 and TTR levels were higher in
subjects with T2D (RBP4, 13; TTR, 832 µg/ml), followed by subjects
with IGT (RBP4, 10.5; TTR, 720.0 µg/ml) and NGT (RBP4, 8.7; TTR,
551.5 µg/ml). RBP4 and TTR exhibited a significant association with
T2D regardless of other factors associated with IR, such as age,
sex, TG and HDL cholesterol. Serum RBP4 levels exhibited a
significant association with IR in all subjects, even in the NGT
group. However, TTR was not associated with IR (52).
By contrast, a study of 167 Thai participants (age,
35–66 years) without T2D or any other chronic disease, assessed
certain anthropometric parameters such as fasting blood glucose
(FBG), glycosylated hemoglobin (HbA1c), fasting insulin and serum
lipid levels, considered risk factors for the development of T2D
(41). The participants of the
aforementioned study were divided into normal (FBG <100 mg/dl)
and high T2D risk group (FBG ≥100 mg/dl). The high T2D risk group
exhibited increased TTR levels and HOMA-IR values, as well as
decreased serum RBP4 levels as determined by ELISA compared with
the normal T2D risk group. Significant positive correlations were
noted between elevated serum TG levels and RBP4, TTR and HOMA-IR
(41). Patients with primary
hypertriglyceridemia exhibit increased rates of non-esterified
fatty acid turnover and secretion into adipose tissue, supplying
excess fatty acids to the liver for TG synthesis (53). This increase suggests that
hypertriglyceridemic patients develop IR in adipose tissue.
Furthermore, RBP4 and TTR have been hypothesized to play a role in
the development of IR (52).
The role of RBP4 in development of IR was analyzed
by Yang et al (33) in
mice with adipose-specific deletion of glucose transporter-4
(adipose-GLUT4−/−), transgenic mice selectively
overexpressing GLUT4 in adipocytes (adipose-GLUT4-Tg),
RBP4-overexpressing mice (RBP4-Tg) and mice with deletion of RBP4
(RBP4−/−). The aforementioned study demonstrated
that adipose-GLUT4−/− mice developed IR in muscle
and liver tissue and exhibited increased levels of RBP4 mRNA and
protein in serum, as determined by western blot analysis. By
contrast, adipose-GLUT4-Tg mice exhibited decreased RBP4 levels in
adipose tissue. Moreover, RBP4−/− mice exhibited
improved insulin sensitivity, while RBP4-Tg mice developed IR.
Considering that RBP4 serves an essential role in the development
of IR, insulin signaling was analyzed in muscle and liver tissue of
RBP4−/− and RBP4-Tg mice; administration of
insulin increased PI3K activity in the muscle of control mice,
whereas its effect was decreased by 30% in RBP4-Tg mice compared
with wild type. By contrast, PI3K activity increased by 80% in
muscle tissue from RBP4−/− compared with control
mice. However, PI3K activity was not altered in the liver of
RBP4-Tg mice (33).
In the skeletal muscle of Wistar rats, RBP4 levels
decrease insulin-dependent glucose uptake (54). RBP4 is associated with impaired
insulin signaling, decreased affinity of insulin receptor substrate
(IRS) 1 for PI3K and decreased GLUT4 translocation to the cell
membrane, which decreases glucose uptake and increases IR in the
muscle (Fig. 1E) (33,55). In adipose tissue, the retinol-RBP4
complex interacts with its membrane receptor STRA6, which activates
the JAK2/STAT3/5 pathway, stimulating expression of SOCS3, an
inhibitor of insulin signaling. SOCS3 specifically inhibits binding
of PI3K to IRS in the PI3K/AKT pathway, leading to IR in adipose
tissue (Fig. 1A) (19). In addition, RBP4 increases hepatic
expression of phosphoenolpyruvate carboxykinase (PEPCK), a molecule
that participates in gluconeogenesis by catalyzing conversion of
oxaloacetate to phosphoenolpyruvate, which increases production of
glucose in the liver and decreases insulin activity to promote
development of hyperglycemia (Fig.
1D) (33,42). Biological processes involving
RBP4, such as inflammation in obesity, dyslipidemia, IR and
hyperglycemia, are key risk factors for development of T2D and
associated disease.
RBP4 in T2D and its comorbidities, such as
retinopathy, nephropathy and CVD
T2D is a chronic metabolic disorder characterized by
hyperglycemia, which results from defective insulin action. An
imbalance between insulin and insulin sensitivity leads to IR,
primarily affecting middle-aged and older adults who have prolonged
hyperglycemia due to obesity (56). An association between serum RBP4
levels in obese subjects and IR has been previously demonstrated by
factors (obesity, IR, OS, metabolic disorder) that contribute to
the development of T2D (9–14).
Involvement of RBP4 has been assessed in the pathogenesis of T2D. A
prospective population-based study of 1,011 Chinese participants
with prediabetes (age, 55.6±7.2 years) demonstrated that serum RBP4
levels (analyzed by ELISA) <31 (HR, 2.01; 95% CI, 1.31-3.09) and
>55 µg/ml (HR, 1.94; 95% CI, 1.32-2.93) are associated with
increased risk of incidence of T2D independent of adiposity and
HOMA-IR, suggesting that serum RBP4 levels may increase the risk of
T2D via pathways that do not involve IR (9). For example, higher serum RBP4 levels
may be involved in pathogenesis of β cell dysfunction (9).
Huang et al (57) reported that increased circulating
RBP4 levels are associated with progression of hyperglycemia in
db/db mice and inversely associated with insulinogenic index (index
that determinates the glucose stimulated insulin secretion). In
vitro, RBP4 inhibits glucose-stimulated insulin secretion
(GSIS) in a dose- and time-dependent manner in primary islets
isolated from C57BL/J mice and in a β cell line (INS-1E).
RBP4-transgenic overexpressing mice (RBP4-Tg; age, 8 weeks) exhibit
a dynamic decrease in GSIS prior to impairment of insulin
sensitivity and glucose tolerance. The aforementioned study also
showed in vivo that islets isolated from RBP4-Tg mice
exhibit significantly decreased levels of GSIS (57). In addition, STRA6 was expressed in
pancreatic β cells, which mediates the inhibitory effect of RBP4 on
insulin synthesis via activation of the JAK2/STAT1/insulin gene
enhancer protein-1 (ISL-1) pathway (58). ISL-1 is a transcription factor
involved in regulation of insulin synthesis and maintenance of
normal β cell function (57,58). Moreover, decreased circulating
RBP4 levels effectively restores β cell function and ameliorate
hyperglycemia in db/db mice. These findings indicated that β-cell
dysfunction is associated with increased RBP4 circulation levels
and hyperglycemia (Fig. 1F)
(58).
RBP4 is associated with multiple T2D comorbidities.
For example, plasma RBP4 levels analyzed by ELISA are significantly
higher in patients with diabetic retinopathy (DR), as well as those
with vision-threatening DR (VTDR) compared with those without DR or
VTDR (59). A meta-analysis
demonstrated an association between elevated RBP4 levels and
development of DR in subjects with T2D (60). The mechanisms underlying the
RBP4/DR axis may involve an association of plasma RBP4 levels with
adverse profile of inflammatory markers and OS, which is associated
with progression of DR (59) due
to the presence of a high levels of lipids in retinal tissue
(61); increased serum RBP4
levels, which induce inflammation in human retinal endothelial
cells and RBP4-mediated progression of DR via a unique and
independent proinflammatory mechanism that increases retinal levels
of IL-18 RNA and protein (60).
By contrast, diabetic nephropathy (DN) is a common
and serious microvascular complication of diabetes and one of the
leading causes of end-stage renal disease worldwide (62). It has been reported that serum
RBP4 levels (analyzed by ELISA) are significantly increased in
diabetic patients with macro- and microalbuminuria compared with
those in control and normoalbuminuria groups (63). Furthermore, serum RBP4 levels are
positively associated with clinical parameters, such as systolic
and diastolic blood pressure (S and DBP), glucose, HbA1c, HOMA-IR,
albumin/creatinine ratio (ACR) and TG levels. Moreover, serum RBP4
levels are inversely associated with glomerular filtration rate
(GFR) (63). A meta-analysis
(62) indicated that levels of
circulating RBP4 are significantly elevated in microalbuminuria and
macroalbuminuria groups compared with normal albuminuria subjects
with T2D. In addition, significantly increased RBP4 levels were
identified in T2D subjects with chronic kidney disease (CKD) and
low GFR and RBP4 levels were positively associated with ACR but
negatively associated with GFR (62). The latter inverse association has
been observed in subjects with prediabetes (9). Retinol homeostasis is mediated by GF
and reabsorption of RBP4 in the proximal tubule. Therefore,
decreased catabolism resulting from microvascular damage in the
kidney leads to a gradual increase in plasma RBP4 concentration in
subjects with DN compared with patients with T2D without DN
(62). The alterations in RBP4
metabolism during CKD are important for development of T2D as
patients with T2D are exposed to increased OS, which is associated
with endothelial dysfunction (63). Patients with T2D often exhibit
kidney dysfunction and therefore RBP4 may increase OS via its
action on polymorphonuclear leukocytes by interacting with TLR4 in
leucocytes and inducing inflammation (6,45,63).
The potential association between RBP4 and
cardiovascular events has also been evaluated. Circulating
adipokines and vascular function were assessed in 3,505
participants of the third-generation Framingham cohort, which were
free of CVD (mean age, 40 years; 53% female, 47% male). In the
aforementioned study, mean arterial pressure was positively
associated with serum RBP4, as determined by ELISA (64). Furthermore, in a study of Chinese
individuals with prehypertension (Pre-HT) (65), elevated serum levels of RBP4 were
noted in subjects with Pre-HT compared with subjects with normal
BP. Moreover, higher serum RBP4 levels were identified in obese
patients with Pre-HT compared with non-obese patients with Pre-HT.
Serum RBP4 levels, determined by ELISA, were positively correlated
with BMI, SBP and DBP (65).
However, a prospective study of 950 Chinese subjects with T2D
(follow-up, 22 years) indicated that higher serum RBP4 levels were
not significantly associated with decreased CVD mortality (66).
In addition, a prospective cohort study revealed
that RBP4 gene overexpression in transgenic mice (RBP4-Tg)
increases incidence of atherosclerotic events by inducing
macrophage-derived foam cell formation (67). In another study of diabetic rats
with atherosclerosis, serum RBP4 levels, analyzed by ELISA, were
positively associated with HOMA-IR, SBP, atherogenic index
(predictor of atherosclerosis) and fasting insulin, TG and CRP
levels (42). RBP4 stimulates
expression of proinflammatory molecules, such as vascular cell
adhesion protein-1, E-selectin, intercellular adhesion molecule-1,
monocyte chemotactic protein-1, IL-6, TNFα and CRP, which induce
endothelial inflammation (Fig.
1C) (18). RBP4-mediated
endothelial inflammation is independent of retinol and signaling
receptor and transporter of retinol STRA6 via the
NADPH/NF-κB-pathway. In addition, RBP4 has a key role in increasing
nitric oxide (NO) production due to stimulation of the PI3K/AKT/NO
synthase pathway and inhibition of extracellular signal-regulated
kinase 1/2 phosphorylation and insulin-induced endothelin-1
secretion, leading to vasodilatation (68).
RBP4 is involved in development of vascular and
endothelial inflammation following oxidative damage (50,63). OS serves a key role in the
pathophysiology of IR and diabetes; it decreases peripheral insulin
sensitivity via at least five key molecular mechanisms, including β
cell dysfunction, inflammatory response, GLUT-4 downregulation
and/or membrane localization, mitochondrial dysfunction and
impairment of normal insulin signaling pathways (69).
RBP4, hyperglycemia and OS
Elevated RBP4 levels promote hyperglycemia and IR by
inhibiting PI3K activity in skeletal muscle and increasing PEPCK
synthesis in the liver, which increases hepatic glucose production,
inhibits insulin signaling and impairs glucose uptake in skeletal
muscle cells (Fig. 1D and E)
(42,62). Hyperglycemia induces free radical
formation and impairs endogenous antioxidant defense systems via
different mechanisms, including increased polyol (sorbitol) and
hexosamine pathway signaling, increased advanced glycation
end-product formation and activation of protein kinase C isoforms.
OS is defined as a disturbance in prooxidant-antioxidant balance of
the cell in favor of the former (69,70). Excess production of ROS, such as
the superoxide anion (O2−) and hydroxyl free
radical and hydrogen peroxide, are harmful to cell components,
notably proteins, lipids and DNA, which damages the cell (70).
Adipokines are associated with numerous types of
OS-associated disease and pathological conditions (obesity,
inflammation, IR, diabetes). RBP4 promotes OS by impairing
endothelial mitochondrial function (Fig. 1C) (71). The association between RBP4 levels
and OS markers has been demonstrated by multiple studies (34,71,72). Codoñer-Franch et al
(34) demonstrated a positive
association between RBP4 and the inflammatory state in obesity and
OS; there was a positive association between RBP4 and urinary
8-isoprostane (a marker of lipid peroxidation damage) and a
negative association between RBP4 and decreased glutathione and
GFR, suggesting that OS was dependent on RBP4 levels.
Liu et al (72) investigated the association between
RBP4 and OS in a Chinese population; their results indicated a
positive association between RBP4 and 8-iso-prostaglandin F2α and
13-(S)-hydroxyoctadecadienoic acid (13-HODE), which are both
products of peroxidation of unsaturated fatty acids. The
aforementioned study established an association between RBP4 and OS
markers in humans (72). In a
study of human aortic endothelial cells (HAECs), the treatment of
40 µg/ml RBP4 increased mitochondrial O2−
production, which promoted mitochondrial dysfunction and increased
membrane potential. RBP4 suppresses mitofusin-1 protein expression
and enhances dynamin-related protein-1 and fission-1 protein
expression levels in HAECs (Fig.
1C), suggesting an impairment of mitochondrial fusion and
fission dynamics (73). Moreover,
mitochondrial damage induces endothelial apoptosis, whereas RBP4
stimulation suppresses PI3K/AKT signaling in HAECs (73).
In addition to endothelial dysfunction, development
of OS and elevated RBP4 levels may be due to certain OS products,
such as 13-HODE, which serve as a ligand for PPARγ, causing its
activation (74). In brown
adipose tissue cells that PPARγ and cAMP-mediated PPARα act on the
promoter of the RBP4 gene to positively regulate RBP4 expression
(44). This reveals a direct
association between certain OS markers and increased RBP4
levels.
Table I summarizes
studies that have analyzed the association between RBP4 and obesity
and associated chronic alterations in different populations
worldwide and in animal models, along with sex, age, sample size
and association between variables.
 | Table I.RBP4 is associated with obesity and
associated clinical/metabolic disorder and disease. |
Table I.
RBP4 is associated with obesity and
associated clinical/metabolic disorder and disease.
| First author/s,
year | Study subjects | Sex | Age | Sample size, n | Clinical/metabolic
disorder or disease | Association | (Refs.) |
|---|
| Fan, 2019 | Chinese
population | Male (n=295) and
female (n=716) | 56±7 years | 1,011 | HOMA-IR, TG, LDLc
in DT2 | + | (9) |
|
|
|
|
|
| TG | - |
|
| Li, 2018 | Chinese
population | Male (n=1754) and
female (n=1591) | 6–18 years | 3,345 | BMI, WC, BP, TG,
TC, LDLc, HOMA-IR, leptin, IR | + | (11) |
|
|
|
|
|
| HDLc | - |
|
| Boaghi, 2020 | Romanian
population | Male and female
(data not available) | 5–17 years | 213 | TG, HDLc in
obesity | - | (32) |
| Yang Q, 2005 | Chinese
population | NA | NA | NA | Obesity, DT2 | + | (33) |
| Yang Q, 2005 | Mouse model | Male | NA | 36 | IR, obesity,
DT2 | + | (33) |
| Wessel, 2019 | Dutch
population | Male (n=29) and
female (n=49) | >18 years | 78 | VLDL and LDLc in
MetS | + | (35) |
| Korek E, 2018 | Polish
population | Male (n=41) and
female (n=12) | 24–63 years | 53 | TG in obesity | + | (36) |
| Mateo-Gallego R,
2018 | Spanish
population | Female | 18–80 years | 76 | TG in obesity | + | (37) |
| Comerford KB,
2014 | US population | Female | NA | 35 | TC, TG in
obesity | + | (38) |
| Numao S, 2012 | Japanese
population | Male | 48±2 years | 29 | TG in obesity | + | (39) |
| Kwanbunjan K,
2018 | Thai
population | Male (n= 47) and
female (n=120) | 35–66 years | 167 | TTR, HOMA-IR and TG
in healthy individuals | + | (41) |
| Zhou, 2018 | Rat model | Male | 4 weeks | 46 | DA, TG, HOMA-IR,
SBP CRP in DT2 | + | (42) |
| Pandey, 2015 | Indian
population | Male (n=123) and
female (n=127) | 20 years | 250 | Obesity, IR,
DT2 | + | (52) |
| Huang, 2021 | Cell model
INS-1E | NA | NA | NA | β cell
dysfunction | + | (57) |
| Li JY, 2018 | Chinese
population | Male (n=155) and
female (n=132) | 62±7 years | 287 | DR, CVDR in
DT2 | + | (59) |
| Mahfouz MH,
2016 | Saudi Arabia
population | Male (n=74) and
female (n=76) | NA | 150 | HT, Glucose, HbA1c,
CRP in DN, DT2 | + | (63) |
|
|
|
|
| GFR | - |
|
| Zachariah,
2016 | US population | Male (n=1644) and
female (n=1861) | 40 years | 3,505 | Arterial
pressure | + | (64) |
| Zhang, 2017 | Chinese
population | Male (n=188) and
female (n=132) | NA | 320 | BMI, SBP, DBP in
Pre-HT | + | (65) |
| Liu, 2016 | Chinese
population | Male | NA | 950 | TG | + | (66) |
|
|
|
|
|
| GFR | - |
|
|
|
|
|
|
| Mortality in CVD in
DT2 | - |
|
| Liu Y, 2017 | Mouse model | NA | NA | NA |
Atherosclerosis | + | (67) |
RBP4 in cancer and obesity-associated
disease
RBP4 is associated with other types of
obesity-associated disease, including liver, bone and joint
pathology, sleep apnea, hepatocellular carcinoma, breast cancer,
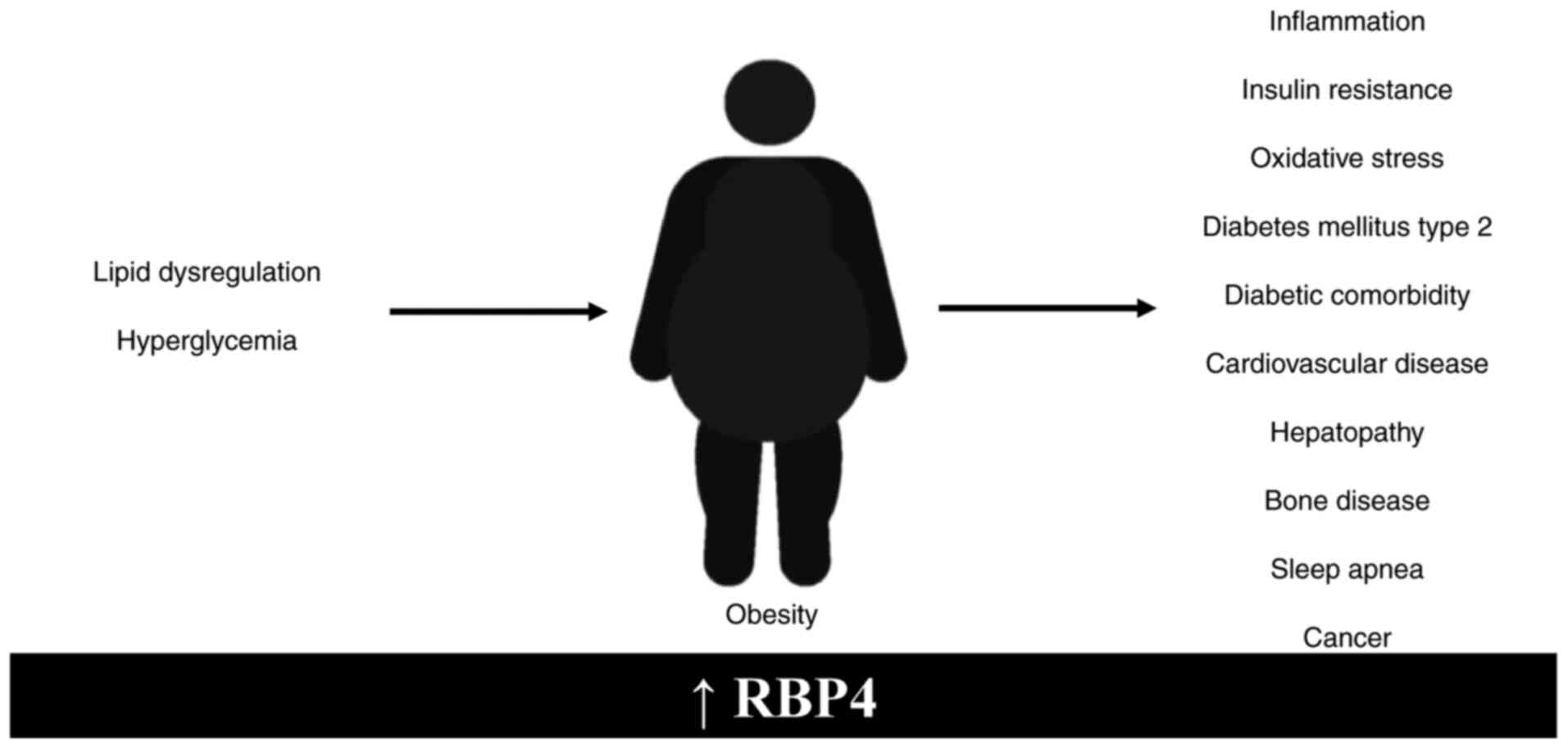
lung cancer and colorectal adenoma and ovarian cancer (75–90). Elevated levels of RBP4 have been
reported in Japanese patients with severe and moderate obstructive
sleep apnea (75). Despite the
lack of association of RBP4 levels with HOMA-IR or BP, a positive
association between visceral fat and TG has been noted (75). However, in Greek patients, no
correlation has been noted between RBP4 levels and sleep apnea,
anthropometric characteristics (BMI, neck circumference, waist
circumference, hip circumference, waist-to-hip ratio), glycemic
markers or lipid profile (76).
By contrast, high levels of RBP4 have been observed
in obese patients with non-alcoholic fatty liver disease (NAFLD)
with and without prediabetes (77). In a murine model, it has been
observed, that RBP4 and retinol levels were significantly decreased
in NAFLD (78). By contrast,
elevated concentrations of RBP4 in Chinese subjects have been
associated with development and progression of NAFLD (79). Elevated RBP4 levels are considered
a risk factor for progression of CKD in patients with NAFLD
(80).
Recent data have shown an association between RBP4
and bone and joint pathology (81–84). RBP4 levels are significantly
elevated in patients with psoriatic arthritis and psoriasis
(80). In addition, RBP4 is
secreted in osteoarthritic joints and is positively associated with
expression levels of other adipokines and extracellular matrix
metalloproteinases (MMPs) 1 and 3 (82). However, a Mendelian randomization
study found no association between RBP4 levels and osteoarthritis
(83).
RBP4 is associated with the incidence of numerous
types of cancer (84–89). RBP4 mRNA and protein expression is
decreased in hepatocellular carcinoma (HCC) but high in normal
tissue. Higher RBP4 expression is associated with improved overall
survival time in patients with HCC (84). RBP4 is associated with complement
and coagulation cascades, metabolic pathways, biosynthesis of
antibiotics and peroxisome proliferator-activated receptor
signaling and pyruvate metabolism pathways (84). Elevated levels of RBP4 in plasma,
tumor tissue, liver and abdominal fat are associated with greater
blood flow impairment and metastatic potential in patients with
breast cancer (85); conversely,
another study found that gene expression levels of RBP4 are
significantly decreased and associated with unfavorable prognosis
and pathogenesis of breast cancer (86). However, Tsakogiannis et al
(87) published a study in 2021
that did not demonstrate a significant association between RBP4 and
breast cancer. By contrast, high levels of serum RBP4 are
associated with increased risk of non-small cell lung cancer
(88). RBP4 has been proposed as
a potential biomarker for early detection of high-risk colorectal
cancer and adenoma (89).
Overexpression of RBP4 in ovarian cancer cells induces expression
levels of MMP2 and MMP9, as well as activation of the RhoA/Rock1
and cyclin D pathways, promoting migration and proliferation of
cancer cells (90). A schematic
representation of altered RBP4 levels in obesity and their
association with other types of pathology is shown in Fig. 2.
Clinical applications of RBP4 and future
perspectives
Currently, use of drugs that modulate RBP4 levels is
proposed for clinical improvement of patients with numerous types
of pathology (hepatic steatosis, obesity, IR, diabetes, atrophic
macular degeneration) (15,33,91–95). One of the first RBP4 agonist drugs
to be discovered was fenretinide. This synthetic retinoid increases
urinary excretion of RBP4 (33).
By binding to RBP4, it increases its hepatic secretion and
sterically alters formation of complexes with TTR, which increases
renal clearance of RBP4 and decreases its serum concentration
(15). In an in vivo model
of obesity induced by a high-fat diet, treatment with fenretinide
increases renal clearance of RBP4, decreases its serum levels and
improves IR and glucose intolerance in study mice (33). In addition, fenretinide has an
inhibitory effect on development of hepatic steatosis in a murine
model of obesity (91). However,
fenretinide treatment inhibits vitamin A synthesis in
β-carotene-fed mice, which is detrimental to visual function;
therefore, further understanding of the effect of this drug on
vitamin A homeostasis is required (96).
A1120 is a non-retinoid ligand for RBP4 that
disrupts the RBP4-TTR complex by inducing a conformational change
at the interaction interface, thereby decreasing serum RBP4
concentration and retinol levels (15). However, A1120 has low hepatic
stability and no effect on glucose levels has been observed in
animal models (93,97). As A1120 is not a retinoid and does
not interfere with vision, it may be a more favorable drug compared
with fenretinide (15).
BPN-14136 is another non-retinoid compound used to
treat age-associated atrophic macular degeneration and Stargardt's
disease. This drug disrupts the RBP4-TTR complex (15), decreases serum RBP4 levels and
prevents high-fat diet-induced obesity and hepatic steatosis in
mice overexpressing RBP4 in adipose tissue. It also decreases
bisretinoid synthesis without affecting vision of mice and induces
decreased CRP and levels of components involved in the complement
cascade in the retina (98).
Non-retinoid antagonists of RBP4 may represent a promising class of
compounds as potential therapeutic agents.
Use of thiazolidinediones (synthetic PPAR ligands)
has been proposed to inhibit expression of RBP4. For example,
rosiglitazone decreases RBP4 levels in the adipose tissue of
Glut4−/− mice (33). In addition, pioglitazone
significantly decreases levels of RBP4 in an obese rat model,
decreases body weight and improves insulin sensitivity (94). Finally, sitagliptin, a dipeptidyl
peptidase-4 inhibitor, decreases RBP4 expression and improves
insulin sensitivity in diabetic rats (95). However, thiazolidinediones are
associated with adverse events, such as weight gain and edema.
Conclusion
RBP4 is an adipokine associated with obesity
(32). RBP4 participates in lipid
metabolism by activating RXR and LXR receptors (37). In addition, RBP4 promotes
development of inflammation via release of proinflammatory
cytokines and activation of macrophages and dendritic cells via the
TLR4 and JNK pathways (45–47). Moreover, RBP4 serves a key role in
IR-mediated activation of the JAK2/STAT5 signaling pathway
(42,51). RBP4 also participates in
development of hyperglycemia in muscle via inhibition of PI3K
activity (19,33,54,55) and in liver via increased PEPCK
activity (33,42). It also regulates OS via
mitochondrial dysfunction (72,73). These processes participate in
development of obesity, RI, diabetes and associated complications,
such as retinopathy, nephropathy and CVD. RBP4 and its downstream
signaling contribute to development of obesity and associated
pathology, the regulation of this adipokine may be important for
clinical improvement of patients. Further research is needed to
develop an effective treatment to regulate levels of this adipokine
without generating adverse effects.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
YAFC, MER and EFA designed the review and wrote the
manuscript. CGC, JMMB and MIBB edited the manuscript. Data
authentication is not applicable. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Piché ME, Tchernof A and Després JP:
Obesity phenotypes, diabetes, and cardiovascular diseases. Circ
Res. 126:1477–1500. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
WHO: World Health Organization, . Obesity
and overweight. 2021 July 20;https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight
|
|
3
|
Shamah-Levy T, Vielma-Orozco E,
Heredia-Hernández O, Romero-Martínez M, Mojica-Cuevas J,
Cuevas-Nasu L and Rivera-Dommarco J: Encuesta Nacional de Salud y
Nutrición 2018–19: Resultados Nacionales. Pública IN: Instituto
Nacional de Salud Pública; 2020
|
|
4
|
Wu H and Ballantyne CM: Metabolic
inflammation and insulin resistance in obesity. Circ Res.
126:1549–1564. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang L, Hu J and Zhou H: Macrophage and
adipocyte mitochondrial dysfunction in obesity-induced metabolic
diseases. World J Mens Health. 39:606–614. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Moraes-Vieira PM, Yore MM, Dwyer PM, Syed
I, Aryal P and Kahn BB: RBP4 activates antigen-presenting cells,
leading to adipose tissue inflammation and systemic insulin
resistance. Cell Metab. 19:512–526. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nono Nankam PA and Blüher M:
Retinol-binding protein 4 in obesity and metabolic dysfunctions.
Mol Cell Endocrinol. 531:1113122021. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tsutsumi C, Okuno M, Tannous L, Piantedosi
R, Allan M, Goodman DS and Blaner WS: Retinoids and
retinoid-binding protein expression in rat adipocytes. J Biol Chem.
267:1805–1810. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fan J, Yin S, Lin D, Liu Y, Chen N, Bai X,
Ke Q, Shen J, You L, Lin X, et al: Association of serum
retinol-binding protein 4 Levels and the risk of incident type 2
diabetes in subjects with prediabetes. Diabetes Care. 42:1574–1581.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Grosjean F, Esposito P, Maccarrone R,
Libetta C, Dal Canton A and Rampino T: RBP4: A culprit for insulin
resistance in end stage renal disease that can be cleared by
hemodiafiltration. BioMed Res Int. Nov 23–2017.(Epub ahead of
print). View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li G, Esangbedo IC, Xu L, Fu J, Li L, Feng
D, Han L, Xiao X, Li M, Mi J, et al: Childhood retinol-binding
protein 4 (RBP4) levels predicting the 10-year risk of insulin
resistance and metabolic syndrome: The BCAMS study. Cardiovasc
Diabetol. 17:692018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Grzegorczyk EA, Harasim-Symbor E, Lukaszuk
B, Harasiuk D, Choromanska B, Mysliwiec P, Zendzian-Piotrowska M
and Chabowski A: Lack of pronounced changes in the expression of
fatty acid handling proteins in adipose tissue and plasma of
morbidly obese humans. Nutr Diabetes. 8:32018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen Y, Lv P, Du M, Liang Z, Zhou M and
Chen D: Increased retinol-free RBP4 contributes to insulin
resistance in gestational diabetes mellitus. Arch Gynecol Obstet.
296:53–61. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Abbas NAT and El Salem A: Metformin,
sitagliptin, and liraglutide modulate serum retinol-binding
protein-4 level and adipocytokine production in type 2 diabetes
mellitus rat model. Can J Physiol Pharmacol. 96:1226–1231. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Steinhoff JS, Lass A and Schupp M:
Biological functions of RBP4 and its relevance for human diseases.
Front Physiol. 12:6599772021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fairulnizal Md, Noh M, Devi Nair
Gunasegavan R and Mustar S: Vitamin A in health and disease.
Vitamin A. 2019. View Article : Google Scholar
|
|
17
|
Olsen T and Blomhoff R: Retinol, retinoic
acid, and retinol-binding protein 4 are differentially associated
with cardiovascular disease, type 2 diabetes, and obesity: An
overview of human studies. Adv Nutr. 11:644–666. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zabetian-Targhi F, Mahmoudi MJ, Rezaei N
and Mahmoudi M: Retinol binding protein 4 in relation to diet,
inflammation, immunity, and cardiovascular diseases. Adv Nutr.
6:748–762. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Noy N: Vitamin A in regulation of insulin
responsiveness: Mini review. Proc Nutr Soc. 75:212–215. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kawaguchi R, Yu J, Honda J, Hu J,
Whitelegge J, Ping P, Wiita P, Bok D and Sun H: A membrane receptor
for retinol binding protein mediates cellular uptake of vitamin A.
Science. 315:820–825. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Napoli JL: Functions of intracellular
retinoid binding-proteins. Subcell Biochem. 81:21–76. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Saari JC: Vitamin A metabolism in rod and
cone visual cycles. Annu Rev Nutr. 32:125–145. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dowling JE: Vitamin A: Its many roles-from
vision and synaptic plasticity to infant mortality. J Comp Physiol
A Neuroethol Sens Neural Behav Physiol. 206:389–399. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chelstowska S, Widjaja-Adhi MA, Silvaroli
JA and Golczak M: Molecular basis for vitamin A uptake and storage
in vertebrates. Nutrients. 8:6762016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wright CB, Redmond TM and Nickerson JM: A
history of the classical visual cycle. Prog Mol Biol Transl Sci.
134:433–448. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang FC, Xu F, Wang TN and Chen GX: Roles
of vitamin A in the regulation of fatty acid synthesis. World J
Clin Cases. 9:4506–4519. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Iskakova M, Karbyshev M, Piskunov A and
Rochette-Egly C: Nuclear and extranuclear effects of vitamin A. Can
J Physiol Pharmacol. 93:1065–1075. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ghyselinck NB and Duester G: Retinoic acid
signaling pathways. Development. 146:dev1675022019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Balmer JE and Blomhoff R: Gene expression
regulation by retinoic acid. J Lipid Res. 43:1773–1808. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Smith JE, Borek C, Gawinowicz MA and
Goodman DS: Structure-function relationships of retinoids in their
effects on retinol-binding protein metabolism in cultured H4II EC3
liver cells. Arch Biochem Biophys. 238:1–9. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bellovino D, Lanyau Y, Garaguso I, Amicone
L, Cavallari C, Tripodi M and Gaetani S: MMH cells: An in vitro
model for the study of retinol-binding protein secretion regulated
by retinol. J Cell Physiol. 181:24–32. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Boaghi A, Pop RM, Vasilache SL, Banescu C,
Hutanu A, Marginean OC and Pascanu IM: Plasma RBP4 level in
association with body composition, metabolic profile, STRA6 and
RBP4 gene polymorphisms in obese Romanian children. Diabetes Metab
Syndr Obes. 13:4643–4650. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang Q, Graham TE, Mody N, Preitner F,
Peroni OD, Zabolotny JM, Kotani K, Quadro L and Kahn BB: Serum
retinol binding protein 4 contributes to insulin resistance in
obesity and type 2 diabetes. Nature. 436:356–362. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Codoñer-Franch P, Mora-Herranz A,
Simó-Jordá R, Pérez-Rambla C, Boix-García L and Faus-Pérez A:
Retinol-binding protein 4 levels are associated with measures of
liver and renal function and oxidant/antioxidant status in obese
children. J Pediatr. 163:593–595. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wessel H, Saeed A, Heegsma J, Connelly MA,
Faber KN and Dullaart RPF: Plasma levels of retinol binding protein
4 relate to large VLDL and small LDL particles in subjects with and
without type 2 diabetes. J Clin Med. 8:17922019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Korek E, Gibas-Dorna M,
Chęcińska-Maciejewska Z, Krauss H, Łagiedo-Żelazowska M,
Kołodziejczak B and Bogdański P: Serum RBP4 positively correlates
with triglyceride level but not with BMI, fat mass and insulin
resistance in healthy obese and non-obese individuals. Biomarkers.
23:683–688. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mateo-Gallego R, Lamiquiz-Moneo I,
Perez-Calahorra S, Marco-Benedí V, Bea AM, Baila-Rueda L,
Laclaustra M, Peñalvo JL, Civeira F and Cenarro A: Different
protein composition of low-calorie diet differently impacts
adipokine profile irrespective of weight loss in overweight and
obese women. Nutr Metab Cardiovasc Dis. 28:133–142. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Comerford KB, Buchan W and Karakas SE: The
effects of weight loss on FABP4 and RBP4 in obese women with
metabolic syndrome. Horm Metab Res. 46:224–231. 2014.PubMed/NCBI
|
|
39
|
Numao S, Sasai H, Nomata Y, Matsuo T, Eto
M, Tsujimoto T and Tanaka K: Effects of exercise training on
circulating retinol-binding protein 4 and cardiovascular disease
risk factors in obese men. Obes Facts. 5:845–855. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
López-Domènech S, Abad-Jiménez Z,
Iannantuoni F, de Marañón AM, Rovira-Llopis S, Morillas C, Bañuls
C, Víctor VM and Rocha M: Moderate weight loss attenuates chronic
endoplasmic reticulum stress and mitochondrial dysfunction in human
obesity. Mol Metab. 19:24–33. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kwanbunjan K, Panprathip P, Phosat C,
Chumpathat N, Wechjakwen N, Puduang S, Auyyuenyong R, Henkel I and
Schweigert FJ: Association of retinol binding protein 4 and
transthyretin with triglyceride levels and insulin resistance in
rural thais with high type 2 diabetes risk. BMC Endocr Disord.
18:262018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhou W, Ye SD, Chen C and Wang W:
Involvement of RBP4 in diabetic atherosclerosis and the role of
vitamin D intervention. J Diabetes Res. 2018:73298612018.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Weiss K, Mihály J, Liebisch G, Marosvölgyi
T, Schmitz G, Decsi T and Rühl R: Effect of synthetic ligands of
PPAR α, β/δ, γ, RAR, RXR and LXR on the fatty acid composition of
phospholipids in mice. Lipids. 46:1013–1020. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Rosell M, Hondares E, Iwamoto S, Gonzalez
FJ, Wabitsch M, Staels B, Olmos Y, Monsalve M, Giralt M, Iglesias R
and Villarroya F: Peroxisome proliferator-activated receptors-α and
-γ, and cAMP-mediated pathways, control retinol-binding protein-4
gene expression in brown adipose tissue. Endocrinology.
153:1162–1173. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Moraes-Vieira PM, Yore MM,
Sontheimer-Phelps A, Castoldi A, Norseen J, Aryal P, Simonyté
Sjödin K and Kahn BB: Retinol binding protein 4 primes the NLRP3
inflammasome by signaling through Toll-like receptors 2 and 4. Proc
Natl Acad Sci USA. 117:31309–31318. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yesudhas D, Gosu V, Anwar MA and Choi S:
Multiple roles of toll-like receptor 4 in colorectal cancer. Front
Immunol. 5:3342014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kilicarslan M, de Weijer BA, Simonyté
Sjödin K, Aryal P, Ter Horst KW, Cakir H, Romijn JA, Ackermans MT,
Janssen IM, Berends FJ, et al: RBP4 increases lipolysis in human
adipocytes and is associated with increased lipolysis and hepatic
insulin resistance in obese women. FASEB J. 34:6099–6110. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Grant RW and Stephens JM: Fat in flames:
Influence of cytokines and pattern recognition receptors on
adipocyte lipolysis. Am J Physiol Endocrinol Metab. 309:E205–E213.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Morigny P, Houssier M, Mouisel E and
Langin D: Adipocyte lipolysis and insulin resistance. Biochimie.
125:259–266. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Farjo KM, Farjo RA, Halsey S, Moiseyev G
and Ma JX: Retinol-binding protein 4 induces inflammation in human
endothelial cells by an NADPH oxidase- and nuclear factor kappa
B-dependent and retinol-independent mechanism. Mol Cell Biol.
32:5103–5115. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Norseen J, Hosooka T, Hammarstedt A, Yore
MM, Kant S, Aryal P, Kiernan UA, Phillips DA, Maruyama H, Kraus BJ,
et al: Retinol-binding protein 4 inhibits insulin signaling in
adipocytes by inducing proinflammatory cytokines in macrophages
through a c-Jun N-terminal kinase- and toll-like receptor
4-dependent and retinol-independent mechanism. Mol Cell Biol.
32:2010–2019. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Pandey GK, Balasubramanyam J, Balakumar M,
Deepa M, Anjana RM, Abhijit S, Kaviya A, Velmurugan K, Miranda P,
Balasubramanyam M, et al: Altered circulating levels of retinol
binding protein 4 and transthyretin in relation to insulin
resistance, obesity, and glucose intolerance in Asian Indians.
Endocr Pract. 21:861–869. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Grundy SM: Hypertriglyceridemia, insulin
resistance, and the metabolic syndrome. Am J Cardiol. 83:25F–29F.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yousefi MR and TaheriChadorneshin H: The
effect of moderate endurance training on gastrocnemius
retinol-binding protein 4 and insulin resistance in
streptozotocin-induced diabetic rats. Interv Med Appl Sci.
10:59–63. 2018.PubMed/NCBI
|
|
55
|
Marschner RA, Pinto G, Borges J, Markoski
MM, Schaan BD and Lehnen AM: Short-term detraining does not change
insulin sensitivity and RBP4 in rodents previously submitted to
aerobic exercise. Horm Metab Res. 49:58–63. 2017.PubMed/NCBI
|
|
56
|
Sapra A and Bhandari P: Diabetes mellitus.
StatPearls. StatPearls Publishing; Treasure Island, FL: 2022
|
|
57
|
Huang R, Bai X, Li X, Wang X and Zhao L:
Retinol-binding protein 4 activates STRA6, provoking pancreatic
β-cell dysfunction in type 2 diabetes. Diabetes. 70:449–463. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Ediger BN, Du A, Liu J, Hunter CS, Walp
ER, Schug J, Kaestner KH, Stein R, Stoffers DA and May CL: Islet-1
Is essential for pancreatic β-cell function. Diabetes.
63:4206–4217. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Li JY, Chen XX, Lu XH, Zhang CB, Shi QP
and Feng L: Elevated RBP4 plasma levels were associated with
diabetic retinopathy in type 2 diabetes. Biosci Rep.
38:BSR201811002018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Han W, Wei H, Kong W, Wang J, Yang L and
Wu H: Association between retinol binding protein 4 and diabetic
retinopathy among type 2 diabetic patients: A meta-analysis. Acta
Diabetol. 57:1203–1218. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Behl T, Kaur I and Kotwani A: Implication
of oxidative stress in progression of diabetic retinopathy. Surv
Ophthalmol. 61:187–196. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Zhang L, Cheng YL, Xue S and Xu ZG: The
role of circulating RBP4 in the type 2 diabetes patients with
kidney diseases: A systematic review and meta-analysis. Dis
Markers. 2020:88304712020. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Mahfouz MH, Assiri AM and Mukhtar MH:
Assessment of neutrophil gelatinase-associated lipocalin (NGAL) and
retinol-binding protein 4 (RBP4) in type 2 diabetic patients with
nephropathy. Biomark Insights. 11:31–40. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Zachariah JP, Hwang S, Hamburg NM,
Benjamin EJ, Larson MG, Levy D, Vita JA, Sullivan LM, Mitchell GF
and Vasan RS: Circulating adipokines and vascular function:
Cross-sectional associations in a community-based cohort.
Hypertension. 67:294–300. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Zhang JX, Zhu GP, Zhang BL and Cheng YY:
Elevated serum retinol-binding protein 4 levels are correlated with
blood pressure in prehypertensive Chinese. J Hum Hypertens.
31:611–615. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Liu G, Ding M, Chiuve SE, Rimm EB, Franks
PW, Meigs JB, Hu FB and Sun Q: Plasma levels of fatty acid-binding
protein 4, retinol-binding protein 4, high-molecular-weight
adiponectin, and cardiovascular mortality among men with type 2
diabetes: A 22-year prospective study. Arterioscler Thromb Vasc
Biol. 36:2259–2267. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Liu Y, Zhong Y, Chen H, Wang D, Wang M, Ou
JS and Xia M: Retinol-binding protein-dependent cholesterol uptake
regulates macrophage foam cell formation and promotes
atherosclerosis. Circulation. 135:1339–1354. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Rychter AM, Skrzypczak-Zielińska M,
Zielińska A, Eder P, Souto EB, Zawada A, Ratajczak AE, Dobrowolska
A and Krela-Kaźmierczak I: Is the retinol-binding protein 4 a
possible risk factor for cardiovascular diseases in obesity? Int J
Mol Sci. 21:52292020. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Yaribeygi H, Sathyapalan T, Atkin SL and
Sahebkar A: Molecular mechanisms linking oxidative stress and
diabetes mellitus. Oxid Med Cell Longev. 2020:86092132020.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Papachristoforou E, Lambadiari V, Maratou
E and Makrilakis K: Association of glycemic indices (hyperglycemia,
glucose variability, and hypoglycemia) with oxidative stress and
diabetic complications. J Diabetes Res. 2020:74897952020.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Daneshzad E, Farsad-Naeimi A, Heshmati J,
Mirzaei K, Maghbooli Z and Keshavarz SA: The association between
dietary antioxidants and adipokines level among obese women.
Diabetes Metab Syndr. 13:1369–1373. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Liu Y, Wang D, Li D, Sun R and Xia M:
Associations of retinol-binding protein 4 with oxidative stress,
inflammatory markers, and metabolic syndrome in a middle-aged and
elderly Chinese population. Diabetol Metab Syndr. 6:252014.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Wang J, Chen H, Liu Y, Zhou W, Sun R and
Xia M: Retinol binding protein 4 induces mitochondrial dysfunction
and vascular oxidative damage. Atherosclerosis. 240:335–344. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Umeno A, Sakashita M, Sugino S, Murotomi
K, Okuzawa T, Morita N, Tomii K, Tsuchiya Y, Yamasaki K, Horie M,
et al: Comprehensive analysis of PPARγ agonist activities of
stereo-, regio-, and enantio-isomers of hydroxyoctadecadienoic
acids. Biosci Rep. 40:BSR201937672020. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Makino S, Fujiwara M, Suzukawa K, Handa H,
Fujie T, Ohtaka Y, Komatsu Y, Aoki Y, Maruyama H, Terada Y, et al:
Visceral obesity is associated with the metabolic syndrome and
elevated plasma retinol binding protein-4 level in obstructive
sleep apnea syndrome. Horm Metab Res. 41:221–226. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Nena E, Steiropoulos P, Tzouvelekis A,
Tsara V, Hatzizisi O, Kyriazis G, Froudarakis M, Trakada G, Papanas
N and Bouros D: Reduction of serum retinol-binding protein-4 levels
in nondiabetic obstructive sleep apnea patients under continuous
positive airway pressure treatment. Respiration. 80:517–523. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Karamfilova V, Gateva A, Alexiev A,
Zheleva N, Velikova T, Ivanova-Boyanova R, Ivanova R, Cherkezov N,
Kamenov Z and Mateva L: The association between retinol-binding
protein 4 and prediabetes in obese patients with nonalcoholic fatty
liver disease. Arch Physiol Biochem. 128:217–222. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Saeed A, Bartuzi P, Heegsma J, Dekker D,
Kloosterhuis N, de Bruin A, Jonker JW, van de Sluis B and Faber KN:
Impaired hepatic vitamin A metabolism in NAFLD mice leading to
vitamin A accumulation in hepatocytes. Cell Mol Gastroenterol
Hepatol. 11:309–325.e3. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Wang X, Chen X, Zhang H, Pang J, Lin J, Xu
X, Yang L, Ma J, Ling W and Chen Y: Circulating retinol-binding
protein 4 is associated with the development and regression of
non-alcoholic fatty liver disease. Diabetes Metab. 46:119–128.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Tsai YL, Liu CW, Huang SF, Yang YY, Lin
MW, Huang CC, Li TH, Huang YH, Hou MC and Lin HC: Urinary fatty
acid and retinol binding protein-4 predict CKD progression in
severe NAFLD patients with hypertension: 4-Year study with clinical
and experimental approaches. Medicine (Baltimore). 99:e186262020.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Toussirot E, Aubin F, Desmarets M,
Wendling D, Augé B, Gillard J, Messica O, Guillot X, Laheurte C,
Monnet E and Dumoulin G: Visceral adiposity in patients with
psoriatic arthritis and psoriasis alone and its relationship with
metabolic and cardiovascular risk. Rheumatology (Oxford).
60:2816–2825. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Scotece M, Koskinen-Kolasa A, Pemmari A,
Leppänen T, Hämäläinen M, Moilanen T, Moilanen E and Vuolteenaho K:
Novel adipokine associated with OA: Retinol binding protein 4
(RBP4) is produced by cartilage and is correlated with MMPs in
osteoarthritis patients. Inflamm Res. 69:415–421. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Fan J, Zhu J, Sun L and Li Y, Wang T and
Li Y: Causal association of adipokines with osteoarthritis: A
Mendelian randomization study. Rheumatology (Oxford). 60:2808–2815.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Li M, Wang Z, Zhu L, Shui Y, Zhang S and
Guo W: Down-regulation of RBP4 indicates a poor prognosis and
correlates with immune cell infiltration in hepatocellular
carcinoma. Biosci Rep. 41:BSR202103282021. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Papiernik D, Urbaniak A, Kłopotowska D,
Nasulewicz-Goldeman A, Ekiert M, Nowak M, Jarosz J, Cuprych M,
Strzykalska A, Ugorski M, et al: Retinol-binding protein 4
accelerates metastatic spread and increases impairment of blood
flow in mouse mammary gland tumors. Cancers (Basel). 12:6232020.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Wu M, Li Q and Wang H: Identification of
novel biomarkers associated with the prognosis and potential
pathogenesis of breast cancer via integrated bioinformatics
analysis. Technol Cancer Res Treat. 20:15330338219920812021.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Tsakogiannis D, Kalogera E, Zagouri F,
Zografos E, Balalis D and Bletsa G: Determination of FABP4, RBP4
and the MMP-9/NGAL complex in the serum of women with breast
cancer. Oncol Lett. 21:852021. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Hu X, Huang W, Wang F, Dai Y, Hu X, Yue D
and Wang S: Serum levels of retinol-binding protein 4 and the risk
of non-small cell lung cancer: A case-control study. Medicine
(Baltimore). 99:e212542020. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Komor MA, Bosch LJ, Coupé VM, Rausch C,
Pham TV, Piersma SR, Mongera S, Mulder CJ, Dekker E, Kuipers EJ, et
al: Proteins in stool as biomarkers for non-invasive detection of
colorectal adenomas with high risk of progression. J Pathol.
250:288–298. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Wang Y, Wang Y and Zhang Z: Adipokine RBP4
drives ovarian cancer cell migration. J Ovarian Res. 11:292018.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Preitner F, Mody N, Graham TE, Peroni OD
and Kahn BB: Long-term Fenretinide treatment prevents high-fat
diet-induced obesity, insulin resistance, and hepatic steatosis. Am
J Physiol Endocrinol Metab. 297:E1420–E1429. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Cioffi CL, Racz B, Varadi A, Freeman EE,
Conlon MP, Chen P, Zhu L, Kitchen DB, Barnes KD, Martin WH, et al:
Design, synthesis, and preclinical efficacy of novel nonretinoid
antagonists of retinol-binding protein 4 in the mouse model of
hepatic steatosis. J Med Chem. 62:5470–5500. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Kim N and Priefer R: Retinol binding
protein 4 antagonists and protein synthesis inhibitors: Potential
for therapeutic development. Eur J Med Chem. 226:1138562021.
View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Zhu C, Xiao Y, Liu X, Han J, Zhang J, Wei
L and Jia W: Pioglitazone lowers serum retinol binding protein 4 by
suppressing its expression in adipose tissue of obese rats. Cell
Physiol Biochem. 35:778–788. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Hu H, Xu M, Qi R, Wang Y, Wang C, Liu J,
Luo L, Xia L and Fang Z: Sitagliptin downregulates retinol-binding
protein 4 and upregulates glucose transporter type 4 expression in
a type 2 diabetes mellitus rat model. Int J Clin Exp Med.
8:17902–17911. 2015.PubMed/NCBI
|
|
96
|
Miller AP, Black M and Amengual J:
Fenretinide inhibits vitamin A formation from β-carotene and
regulates carotenoid levels in mice. Biochim Biophys Acta Mol Cell
Biol Lipids. 1867:1590702022. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Blaner WS: Vitamin A signaling and
homeostasis in obesity, diabetes, and metabolic disorders.
Pharmacol Ther. 197:153–178. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Racz B, Varadi A, Kong J, Allikmets R,
Pearson PG, Johnson G, Cioffi CL and Petrukhin K: A non-retinoid
antagonist of retinol-binding protein 4 rescues phenotype in a
model of Stargardt disease without inhibiting the visual cycle. J
Biol Chem. 293(29): 11574–11588. 2018. View Article : Google Scholar : PubMed/NCBI
|