Introduction
Atherosclerosis (AS) is one of the most common
vascular lesions and is also considered the main pathological basis
of cardiovascular disease (1).
Several cell types and molecules are involved in the pathogenesis
of AS, mainly including endothelial cells, smooth muscle cells and
inflammatory cells (2).
Furthermore, it has been reported that the pathophysiology of AS
originates from endothelial cell injury; however, the specific
pathogenesis has not yet been clarified (3).
As a class of endogenous non-coding RNA, circulating
RNAs (circRNAs) do not have a 5′ cap and a 3′ poly tail, and exist
in their characteristic covalent closed-loop form (4). Previous studies have indicated that
circRNAs are widely present in vivo, are stably expressed in
plasma, serum and tissues, and are involved in regulating gene
expression in eukaryotes (5–7).
Notably, circRNAs can adsorb microRNAs (miRs/miRNAs) by acting as
molecular sponges, thus removing the negative regulatory effect of
miRNAs on their downstream target genes, so as to achieve their own
biological function (8). In
addition, circRNAs have been shown to be involved in the
development and progression of various cardiovascular diseases
(9). For example, circ_0010283 has
been reported to be highly expressed in oxidized low-density
lipoprotein (ox-LDL)-induced vascular smooth muscle cells, and to
promote cell proliferation and migration by regulating the
miR-370-3P/HMGB1 signaling axis (10). Another study indicated that
circ_0124644 was highly expressed in ox-LDL-induced human umbilical
vein cells, and PAAP-A expression was upregulated via sponge
adsorption of miR-149-5p, thus further worsening endothelial cell
injury (11).
High expression levels of circ_0005699 in
ox-LDL-stimulated foam cells have previously been reported
(12). Formation of macrophage foam
cells is known to be a major hallmark of the initiation of AS, and
uncontrolled uptake of ox-LDL has been confirmed as one of the
contributing factors leading to foam cell formation (13); however, the role of circ_0005699 in
cardiovascular disease is still unclear. Therefore, the present
study mainly investigated the role of circ_0005699 in regulating
endothelial cell function and further explored its possible
molecular mechanism. The online bioinformatics tool starBase was
used to predict the miRNA target of circ_0005699 and the target
gene of the identified miRNA, and the findings were confirmed by
molecular and cellular biological assays. Furthermore,
ApoE-deficient mice, which exhibit high ox-LDL levels, were used
for in vivo studies. ox-LDL was also applied to human
umbilical vein endothelial cells (HUVECs) in vitro to
investigate its effects on the regulation of cell survival and
circ_0005699 expression.
Materials and methods
Cell culture and treatment
The immortalized HUVEC line was obtained from
American Type Culture Collection (cat. no. CRL-1730). HUVECs were
cultured as previously described (14). The cells were cultured in a nutrient
solution containing F-12 K basic medium, 10% fetal bovine serum and
1% penicillin-streptomycin (all from Gibco; Thermo Fisher
Scientific, Inc.) at 37°C with 5% CO2. Ox-LDL can act on
endothelial cells and is the main risk factor of AS (15). Therefore, the present study treated
HUVECs with different concentrations (0, 25, 50 and 100 µg/ml) of
ox-LDL (Union-Biol) for different durations (0, 12, 24 and 48 h) at
37°C to establish the AS cell model. For experiments, the cells
used were between passages 3 and 6.
Cell transfection
pLKO.1-puro was used as the backbone for the shRNA
constructs. circ_0005699 short hairpin RNA
(sh-circ_0005699#1/sh-circ_0005699#2), negative control shRNA
(sh-NC), miR-450b-5p mimics and negative control (mimics-NC),
miR-450b-5p inhibitor and negative control (inhibitor-NC) and
pcDNA3.1-NFKB1 plasmid were constructed by Shanghai GenePharma Co.,
Ltd. Empty pcDNA3.1 was used as a control for pcDNA3-1-NFKB1. Cell
(4×105) transfection was performed using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) at 37°C for 48 h and cells were collected 72 h
after transfection for subsequent experimentation. The
concentration of shRNAs, mimics and inhibitors were 50
nM/1×105 cells; the concentration of overexpression
plasmid was 0.8 µg/1×105 cells. The sequences were as
follows: miR-450b-5p mimics, 5′-UUUUGCAAUAUGUUCCUGAAUA-5′; miR-NC,
5′-TTCTCCGAACGTGTCACGT-3′; miR-450b-5p inhibitor,
5′-UAUUCAGGAACAUAUUGCAAAA-3′; inhibitor NC:
5′-UUCUCCGAACGUGUCACGUTT-3′; sh-NC,
5′-GGCAACAAGATGAAGAGCACCAACTCGAGTTGGTGCTCTTCATCTTGTTGTTTTTG-3′;
sh-circ_0005699#1,
5′-CACCGCCACAAGGTCTATGGATTTCCGAAGAAATCCATAGACCTTGTGGC-3′; and
sh-circ_0005699#2,
5′-CACCGCTTGGAAAGTCATCACTAAGCGAACTTAGTGATGACTTTCCAAGC-3′.
Animal handling
Six male C57BL/6 wild-type (WT) mice and six
ApoE-knockout mice with a C57BL/6 background were obtained from
Beijing HFK Bioscience Co., Ltd. Mice were maintained under the
following standard laboratory conditions: Temperature, 22°C;
humidity, 55%; 12-h light/dark cycle; and ad libitum access
to food and water. Handling of mice was performed according to the
Institutional Animal Care of The Affiliated Ganzhou Hospital of
Nanchang University (Gangzhou, China) (16). After 12 weeks, the mice were
sacrificed under excessive anesthesia using sodium pentobarbital
(120 mg/kg; intraperitoneal injection). The research team and
veterinary staff monitored the mice twice daily. Health was
monitored by measuring their weight (twice weekly), assessing food
and water intake, and general assessment of animal activity,
panting and fur condition. Before the end of the experiment, the
mice were euthanized once they had fasted for 12 h. The mice were
euthanized by sodium pentobarbital. The drug was injected
intraperitoneally at a dose of 120 mg/kg, and death was confirmed
by the lack of a heartbeat. Serum samples were obtained via cardiac
puncture and centrifugation of blood samples for 10 min at 1,000 ×
g at 4°C, and were stored at −80°C until further use. All of the
procedures were reviewed and approved by the Ethical Committee for
Animal Experimentation, The Affiliated Ganzhou Hospital of Nanchang
University (Ganzhou, China; approval no. KY-E-2019-11-19).
ELISA
ELISA kits for the determination of serum and cell
concentrations of TNF-α (cat. no. ab208348), IL-6 (cat. no.
ab222503) and IL-1β (cat. no. ab197742) were purchased from Abcam.
Prior to ELISA, cells were treated with 100 µg/ml ox-LDL for 48 h.
The experiments were conducted according to the manufacturer's
protocols.
Annexin V/PI assay
The Annexin V/PI assay was used to determine the
apoptosis of HUVECs according to a previously described protocol
(17). After treatment (100 µg/ml
ox-LDL for 48 h), cells were harvested and washed three times with
PBS. Subsequently, cells (1×104) were incubated in the
dark at room temperature with 5 µl Annexin V-FITC and PI (cat. no.
331200; Thermo Fisher Scientific, Inc.) for 15 min. Cells
undergoing apoptosis (early + late) were detected by flow cytometry
(Accuri C6; BD Biosciences) and cell apoptosis was determined using
FlowJo 7.6.1 software (FlowJo LLC).
Cell counting kit-8 (CCK-8) assay
The CCK-8 assay was used to assess the proliferation
of cells. Briefly, HUVECs were seeded in 96-well plates at a
density of 1×103 cells/well, and 10 µl CCK-8 solution
(Dojindo Laboratories, Inc.) was added to each well after 0, 24, 48
and 72 h. Subsequently, the cells were cultured at 37°C for a
further 120 min, followed by the measurement of optical density at
450 nm using a plate reader (Tecan Group, Ltd.).
Western blot analysis
Briefly, cells were lysed using RIPA buffer (Wuhan
Sanying Biotechnology) containing protease inhibitors, and total
protein was extracted. The BCA method was used to determine protein
concentration in the samples. Subsequently, SDS-PAGE was used for
separation of proteins (40 µg) on 10% gels, which were transferred
onto polyvinylidene difluoride membranes (MilliporeSigma).
Membranes were then blocked with 4% BSA (Cell Signaling Technology,
Inc.) at 4°C for 2 h and incubated with the following primary
antibodies (Abcam): Anti-B-cell lymphoma-2 (Bcl-2; cat. no.
ab196495; 1:1,000), anti-Bax (cat. no. ab32503; 1:1,000),
anti-NFKB1 (cat. no. ab32360; 1:1,000) and anti-GAPDH (cat. no.
ab8245; 1:3,000; Abcam) at 4°C overnight; anti-GAPDH was used as
the internal reference. The primary antibodies were then removed
and the membranes were further incubated with HRP-conjugated
secondary antibodies (anti-mouse IgG(H+L); cat. no. ab205719;
anti-rabbit IgG (H+L); cat. no. ab205718; 1:5,000; Abcam) for 1 h
at room temperature. Protein bands were visualized using an ECL
assay kit (Bio-Rad Laboratories, Inc.).
Bioinformatics analysis
The online bioinformatics tool starBase database
(http://starbase.sysu.edu.cn/), which
predicts the target miRNA of the circRNA (18) was used in the present study. This
software was also used to predict the miRNA and target gene
interaction.
Dual-luciferase reporter gene
assay
WT and mutant (Mut) sequences of circ_0005699 and
NFKB1 were inserted into the BamHI and SalI sites
downstream of the luciferase gene in the pGL3-control vector
(Promega Corporation); the constructs were named circ_0005699
WT/circ_0005699 Mut and NFKB1 WT/NFKB1 Mut, respectively. The final
concentration of 20 nM for each plasmid was subsequently
co-transfected into HUVECs (lx106) along with miR-NC and
miR-450b-3p mimics using Lipofectamine 3000 for 48 h at 37°C
according to the manufacturer's protocol. Relative firefly
luciferase activity was normalized against Renilla
luciferase activity. The Dual-Glo Luciferase Reporter Assay System
(Promega Corporation) was used for measurement of luciferase
activity according to the manufacturer's protocol.
RNA pull-down assay and RNA
immunoprecipitation (RIP)
The RNA pull-down assay was performed according to a
previously published protocol (19,20).
This assay was performed to confirm the interaction between
circ_0005699 and miR-450b-3p. Biotin-labeled circ_0005699 probe and
Bio-Oligo (Oligo probe) was purchased from Sangon Biotech Co., Ltd.
HUVECs were trypsinized (Gibco; Thermo Fisher Scientific, Inc.)
followed by cell lysis using RIP lysis buffer (Thermo Fisher
Scientific, Inc.). The lysate alone served as the input control.
The 0.7 ml lysate was incubated at 4°C overnight with 50 µl
magnetic Dynabeads M-280 Streptavidin beads (Invitrogen; Thermo
Fisher Scientific, Inc.). RNA complexes were subjected to
centrifugation at 11,100 × g for 10 min at 37°C and then eluted by
denaturation in 1X protein loading buffer for 10 min at 100°C. The
enrichment of miR-450b-3p was detected using reverse
transcription-quantitative PCR (RT-qPCR). Furthermore, the Magna
RIP kit (cat. no. 17-704; MilliporeSigma) was used according to the
manufacturer's guidelines and a previously described protocol
(21). Briefly, cells were lysed
using RIP lysis buffer (MedChemExpress,). Subsequently, 5 µg
AGO2-specific antibody (cat. no. 2897; Cell Signaling Technology,
Inc.) and a normal IgG antibody (cat. no. 58802; Cell Signaling
Technology, Inc.) were conjugated to magnetic beads and mixed with
the lysate (40 µg) for 4 h at 4°C. The magnetic beads were then
harvested and incubated with 50 µl protein G at 37°C for 1 h.
Finally, RNA was extracted from the beads and relative enrichment
was analyzed using RT-qPCR.
RT-qPCR
RNAiso Plus reagent (Takara Bio, Inc.) was used to
extract total RNA from HUVECs and 1 µg total RNA was reverse
transcribed into cDNA using PrimeScript cDNA synthesis kit (Takara
Bio, Inc.) according to manufacturer's protocol. The resulting cDNA
was amplified using SYBR Premix Ex Taq™ II (Takara Bio,
Inc.) in a real-time PCR detection system (Applied Biosystems;
Thermo Fisher Scientific, Inc.). GAPDH was used as an internal
reference for circ_0005699 and NFKB1, and U6 served as an
endogenous control for miR-450b-5p. qPCR was performed as follows:
Pre-denaturation at 95°C for 1 min; followed by 35 cycles of 95°C
for 10 sec, 60°C for 20 sec and 72°C for 10 sec; and a final
extension step at 72°C for 2 min. Gene expression was analyzed
using the 2−ΔΔCq method (22). The primer sequences were as follows:
circ_0005699 forward (F), 5′-TCCCCTTGTACGAAATCATTCCA-3′ reverse
(R), 5′-ATTGAGACGTGTGAAGATGCCC-3′; miR-450b-5p F,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGTATTCAGG-3′
R,5′-ACACTCCAGCTGGGTTTTGCAATATGTTCC-3′; NFKB1 F,
5′-GCAGCACTACTTCTTGACCACC-3′ R, 5′-TCTGCTCCTGAGCATTGACGTC-3′. GAPDH
F, 5′-ACGGGAAGCTCACTGGCATGG-3′ R, 5′-GGTCCACCACCCTGTTGCTGTA-3′. U6
F, 5′-CTCGCTTCGGCAGCACA-3′ R, 5′-AACGCTTCACGAATTTGCGT-3′.
Statistical analysis
GraphPad Prism software version 6 (GraphPad
Software, Inc.) was used to statistically analyze the results. The
data are presented as the mean ± SD of at least three independent
experiments. Unpaired Student's t-test was used to compare two
groups, whereas the statistical analysis of more than two groups
was performed using one-way ANOVA with Bonferroni's correction.
P<0.05 was considered to indicate a statistically significant
difference.
Results
circ_0005699 is upregulated in HUVECs
induced by ox-LDL and in vivo
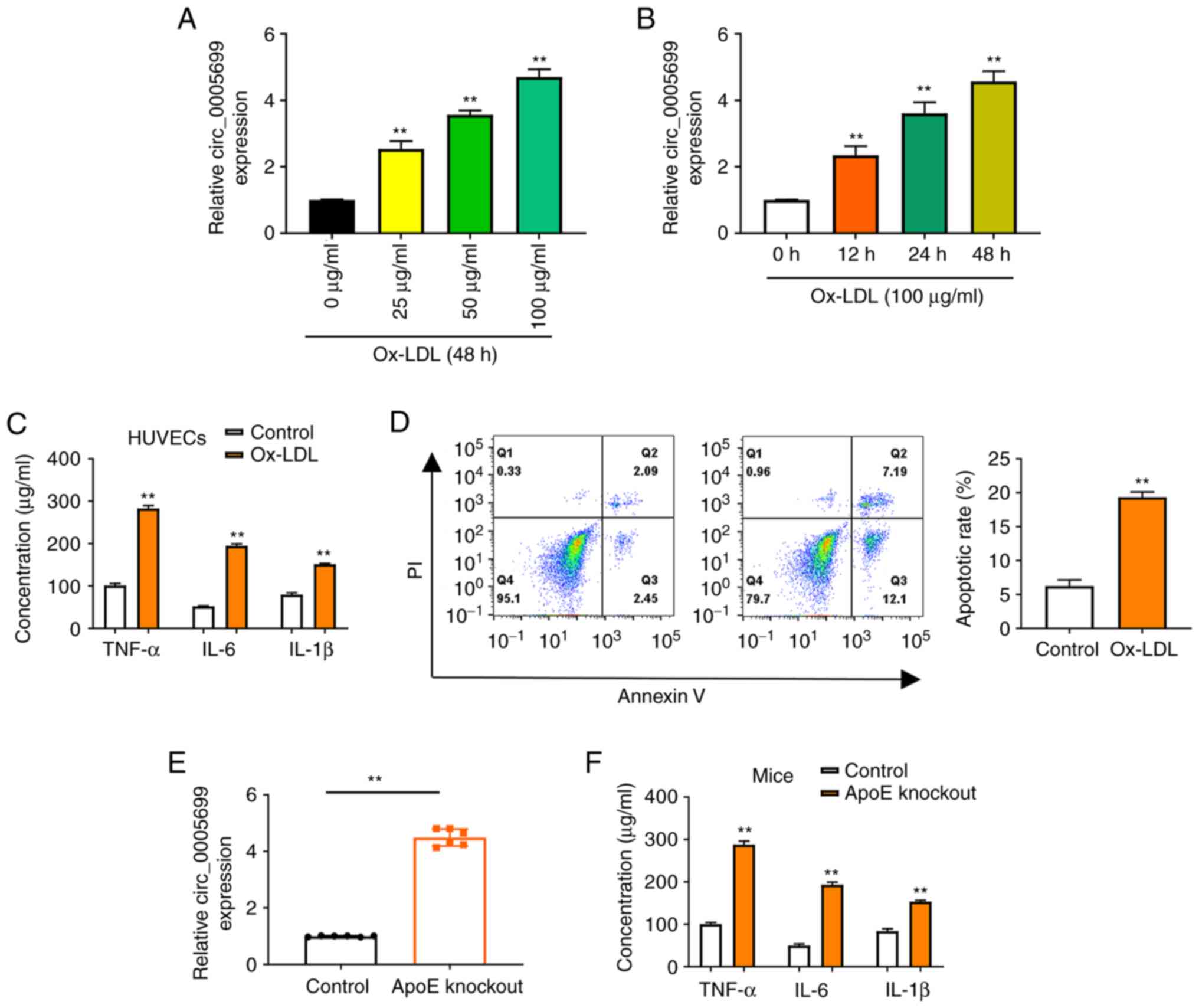
The present study observed that ox-LDL resulted in
increased expression of circ_0005699 in HUVECs in a dose-dependent
(Fig. 1A) and time-dependent manner
(Fig. 1B). Notably, ox-LDL
treatment was also associated with significantly increased
concentrations of TNF-α, IL-6 and IL-1β in HUVECs compared with
those in the control group (untreated cells) (Fig. 1C). Annexin V/PI double-staining and
flow cytometry was used to detect the apoptotic rate of
ox-LDL-treated HUVECs. It was observed that the apoptotic rate was
significantly increased in the ox-LDL group compared with that in
the control group (Fig. 1D).
Increased ox-LDL has previously been reported in ApoE-knockout mice
(23). In the present study,
increased expression of circ_0005699 (Fig. 1E), and significantly increased serum
concentrations of TNF-α, IL-6 and IL-1β were observed in
ApoE-knockout mice (Fig. 1F).
circ_0005699 knockdown decreases
ox-LDL-treated HUVEC apoptosis and inflammation
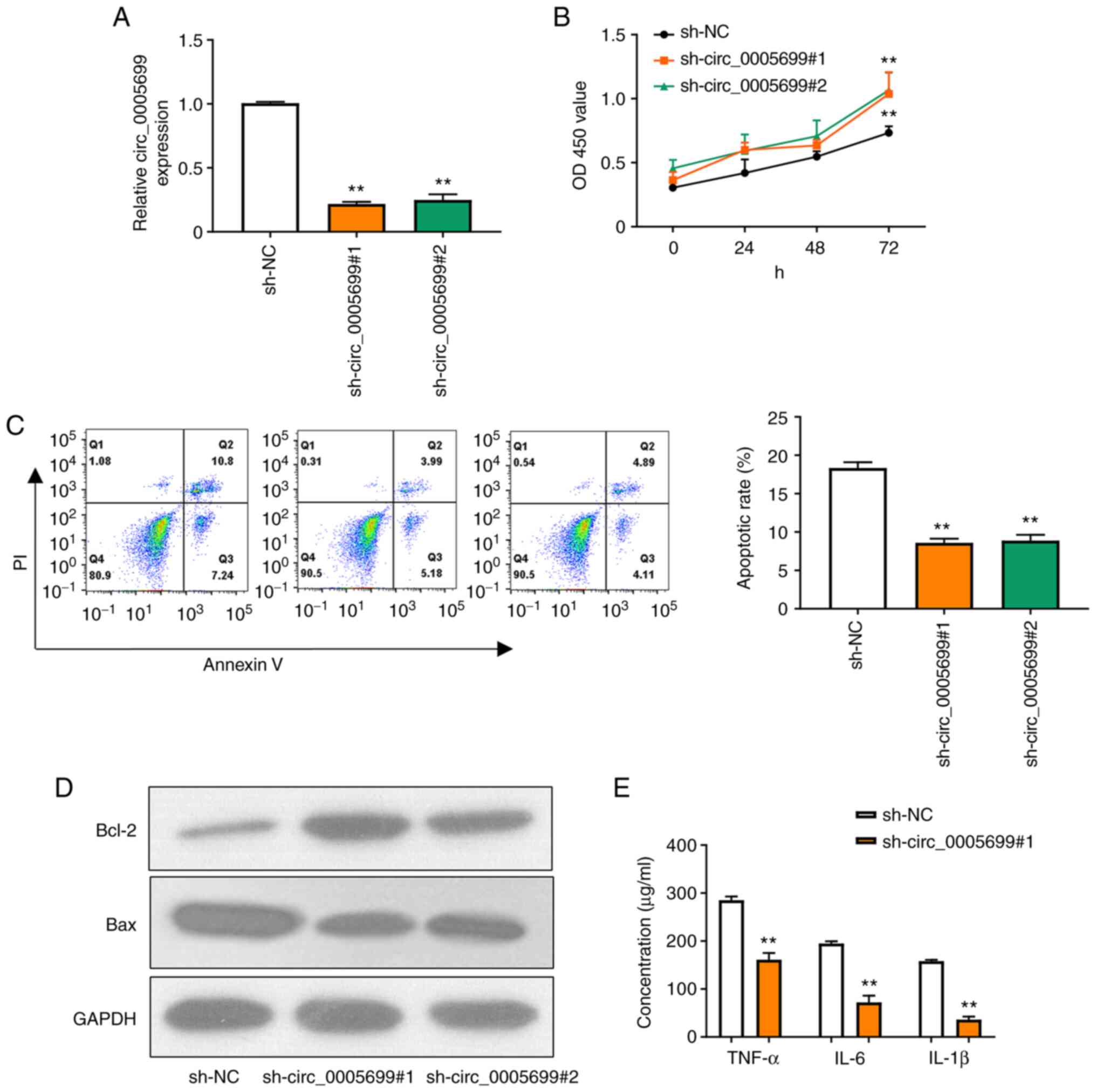
Two shRNAs were designed to target circ_0005699 and
the knockdown efficiency was confirmed by RT-qPCR (Fig. 2A); as illustrated, the expression of
circ_0005699 was significantly decreased following transfection
with the two knockdown shRNAs and there was no significant
difference between the two, thus one of them could be selected for
subsequent experiments. Notably, circ_0005699 knockdown was
associated with increased proliferation of ox-LDL-treated HUVECs
(Fig. 2B). In addition,
circ_0005699 knockdown significantly reduced the apoptotic rate in
these cells, as indicated by Annexin V/PI double-staining and flow
cytometry (Fig. 2C). The
apoptosis-related proteins Bcl-2 and Bax were detected by western
blotting. Compared with that in the control group, knockdown of
circ_0005699 markedly increased the protein expression levels of
Bcl-2, whereas Bax expression was decreased. Furthermore, knockdown
of circ_0005699 significantly reduced the levels of inflammatory
factors, including TNF-α, IL-6 and IL-1β in the supernatants of
ox-LDL-treated HUVECs (Fig.
2E).
circ_0005699 sponges miR-450b-5b
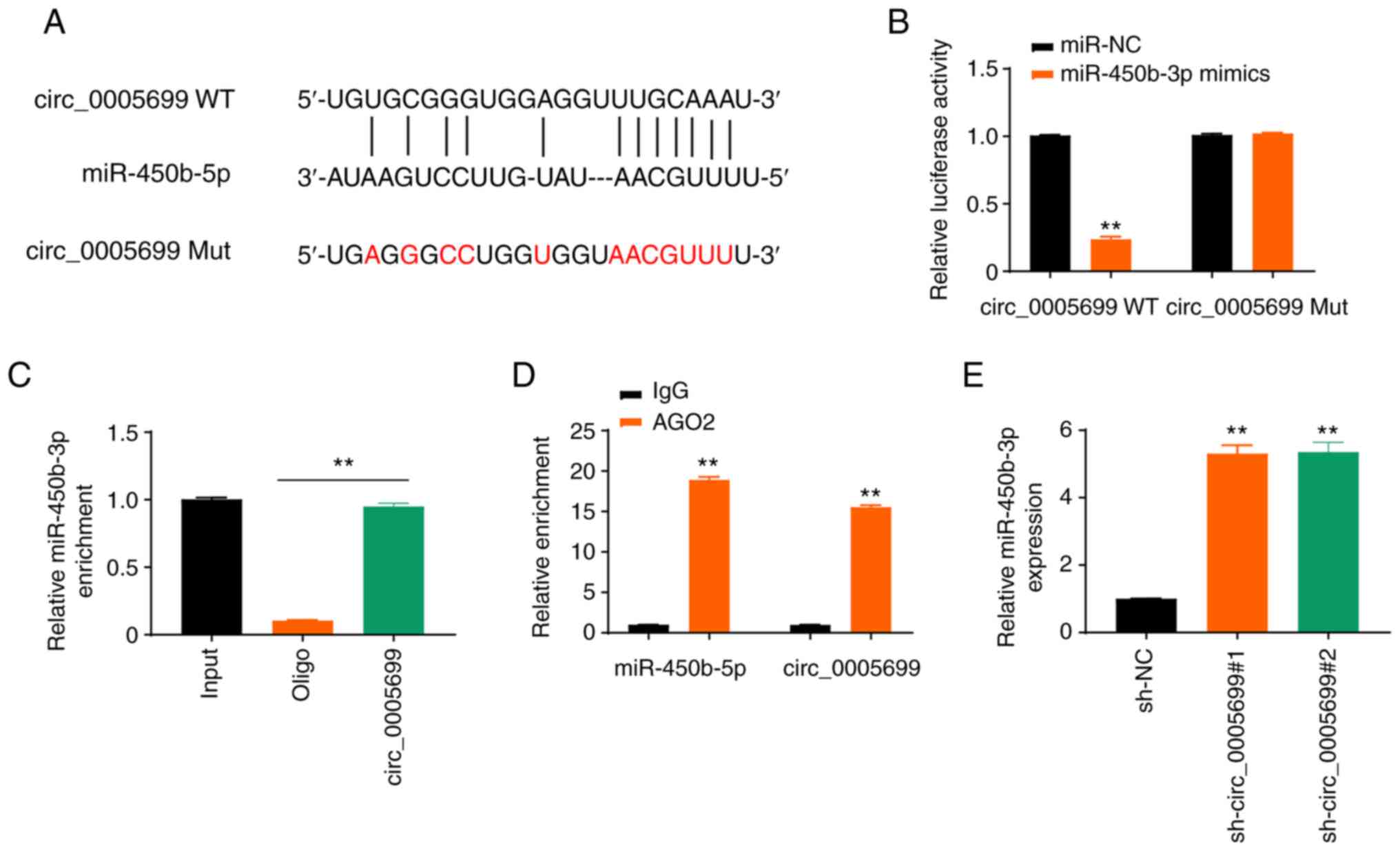
The starBase online database revealed that
circ_0005699 possessed a binding site for miR-450b-5p (Fig. 3A). The expression levels of
miR-450b-5p in ox-LDL-treated cells transfected with miR-450b-5p
mimics were detected and it was revealed that the expression levels
of miR-450b-5p were significantly increased (Fig. S1A). Notably, miR-450b-5p
overexpression resulted in decreased luciferase activity in
circ_0005699 WT ox-LDL-treated cells, but not in circ_0005699 Mut
cells (Fig. 3B). In addition, the
circ_0005699 probe pulled down more miR-450b-5p than the Oligo
probe (Fig. 3C), which verified
that circ_0005699 could bind to miR-450b-5p, whereas the beads
coupled with AGO2 pulled down significantly greater circ_0005699
and miR-450b-5p than the IgG control (Fig. 3D). Furthermore,
circ_0005699-knockdown via shRNA resulted in significantly
increased expression of miR-450b-3p compared with that in the
control group, as indicated by RT-qPCR analysis (Fig. 3E).
NFKB1 is the target of
miR-450b-5p
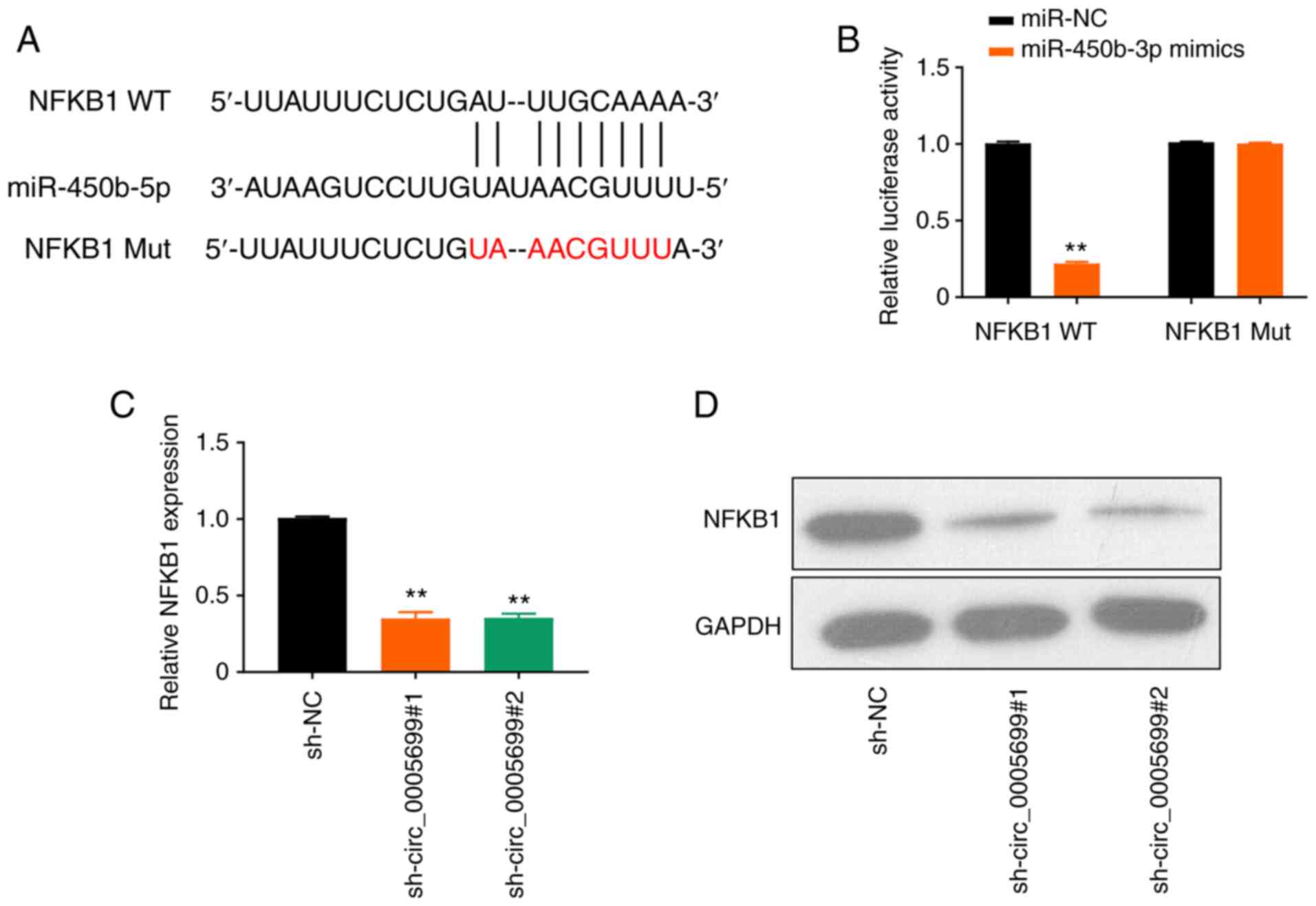
The online database starBase predicted that
miR-450b-5p may target NFKB1 (Fig.
4A). It was observed that the overexpression of miR-450b-5p
significantly decreased luciferase activity in NFKB1 WT
ox-LDL-treated cells compared with the control; however, this
effect was not observed following mutation in the binding site of
NFKB1 (Fig. 4B). Furthermore,
circ_0005699 silencing resulted in significantly decreased
expression levels of NFKB1 in ox-LDL-treated cells compared with
those in the control group, as determined using RT-qPCR (Fig. 4C) and western blot analysis
(Fig. 4D).
circ_0005699 attenuates ox-LDL-treated
HUVEC apoptosis and inflammation through the miR-450b-5p/NFKB1
axis
Western blotting and RT-qPCR were used to detect the
protein and mRNA expression levels of NFKB1 in different groups of
ox-LDL-treated HUVECs following different treatments. First,
miR-450b-5p expression was markedly decreased in ox-LDL-treated
HUVECs transfected with miR-450b-5p inhibitor (Fig. S1B). In addition, NFKB1 expression
was significantly increased in ox-LDL-treated HUVECs transfected
with NFKB1 plasmid (Fig. S1C).
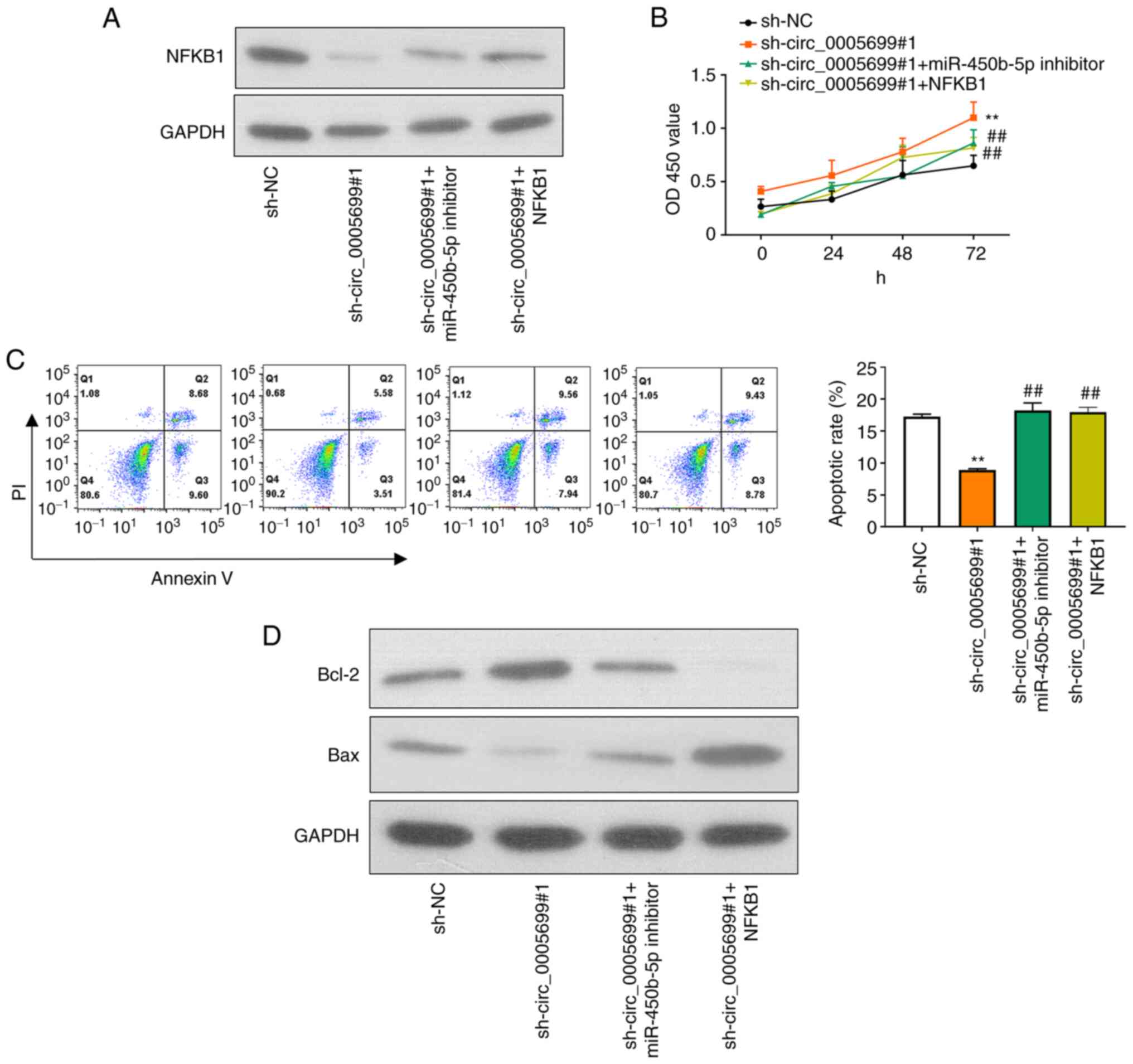
Compared with that in the control group, knockdown of circ_0005699
was associated with decreased NFKB1 protein expression (Fig. 5A). Notably, ox-LDL-treated HUVECs
co-transfected with sh-circ_0005699#1 and miR-450b-5p inhibitor or
NFKB1 overexpression plasmid exhibited partial restoration of NFKB1
expression compared with that in the circ_0005699 knockdown group
(Fig. 5A). Furthermore,
circ_0005699 knockdown was associated with increased cell
proliferation (Fig. 5B) and
significantly reduced apoptotic rate compared with sh-NC group
(Fig. 5C); these effects were
reversed when cells were co-transfected with sh-circ_0005699#1 and
miR-450b-5p inhibitor or NFKB1 overexpression plasmid (Fig. 5C). In addition, the protein
expression levels of Bcl-2 were markedly increased, whereas Bax
protein expression was inhibited following knockdown of
circ_0005699 (Fig. 5D). By
contrast, when cells were co-transfected with sh-circ_0005699#1 and
miR-450b-5p inhibitor or NFKB1 overexpression plasmid, Bcl-2
protein expression was reduced and Bax protein expression was
partially increased compared with that in cells transfected with
sh-circ_0005699#1 only (Fig.
5D).
Discussion
Cardiovascular disease caused by ASAS remains the
leading cause of mortality worldwide (24) and is considered a type of chronic
inflammatory disease (25). It has
previously been reported that ox-LDL may serve a critical role in
the development of atherosclerotic plaque formation through various
mechanisms. These mechanisms may include endothelial dysfunction
(26–28), foam cell formation (29–31),
smooth muscle cell migration and proliferation (32,33),
and the induction of platelet adhesion and aggregation (34,35).
Wang et al (12) previously
indicated that ox-LDL-induced foam cells (an in vitro AS
model) had increased expression of circ_0005699. To the best of our
knowledge, there is no previous study available that investigates
the role of circ_0005699 in the regulation of vascular endothelial
cells. Therefore, HUVECs, endothelial cells isolated from the human
umbilical cord vein, were used in the present study. These cells
have been widely used as a model for the study of function and
pathologies of endothelial cells, including AS and plaque formation
(36–40). In addition, ApoE-deficient mice were
used as an in vivo model of ASAS. These mice, first produced
by Plump et al (41) exhibit
atherosclerotic transformation even under a normal chow-diet
(42).
The results of the present study indicated that
ox-LDL resulted in increased expression of circ_0005699 in HUVECs
in a time- and dose-dependent manner, which is in corroboration
with a previous report (12).
Notably, circ_0005699 silencing was associated with increased
proliferation of HUVECs, along with a decreased rate of apoptosis
in the present study. Furthermore, circ_0005699 was shown to sponge
miR-450b-5b and NFKB1 was identified as a target gene of
miR-450b-5p; therefore, circ_0005699 may attenuate HUVEC apoptosis
and inflammation through the miR-450b-5p/NFKB1 axis.
The starBase database predicted that circ_0005699
could sponge miR-450b-5p. Furthermore, circ_0005699 silencing was
associated with increased miR-450b-5p expression, thus indicating
that miR-450b-5p was a target of circ_0005699. The role of
miR-450b-5p in the regulation of various genes has previously been
investigated, particularly in cancer. For example, miR-450b-5p
downregulation has been reported to be associated with the
progression of hepatocellular carcinoma (43), whereas miR-450b-5p expression was
revealed to be upregulated in colorectal cancer cells (44). Furthermore, another study has
indicated miR-450b-5p may be a potential biomarker of ischemic
injury (45).
The starBase database further predicted that NFKB1
was a target gene of miR-450b-5p and this was confirmed by
luciferase reporter gene assay. NFKB is a transcription factor that
has been shown to regulate innate and adaptive immune responses,
and can induce the transcription of various inflammatory mediators,
including TNF-α, IL-1β, IL-6, IL-12p40 and cyclooxygenase-2
(46). Therefore, NFKB1 has been
implicated in the pathogenesis of multiple inflammatory diseases,
including AS (47–49). A recent report by Huang et al
(50) revealed that miR-450b-5p
inhibition could reduce ischemic injury in hepatic cells through an
NFKB-dependent mechanism. Another report suggested that miR-148a-3p
reduced NFKB signaling and thus proinflammatory gene expression in
aortic valve cells (51), which
further corroborates the present findings, which indicated that
NFKB promoted the apoptosis of HUVECs and aggravated
inflammation.
In conclusion, the findings of the present study
indicated that circ_0005699 knockdown may have beneficial effects
on the survival and function of endothelial cells through the
regulation of miR-450b-5p/NFKB1.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
TC designed the project and collected data. BY and
WC analyzed the data and drafted the manuscript. GZ performed
almost all of the experiments. HX and YG performed some
experiments, and LL was involved in data collection and analysis.
TC and LL confirm the authenticity of all the raw data. All of the
authors revised and corrected the manuscript, and read and approved
the final manuscript.
Ethics approval and consent to
participate
All of the procedures were reviewed and approved by
the Ethical Committee for Animal Experimentation, The Affiliated
Ganzhou Hospital of Nanchang University (Ganzhou, China; approval
no. KY-E-2019-11-19).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Shah PK: Inflammation, infection and
atherosclerosis. Trends Cardiovasc Med. 29:468–472. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Katakami N: Mechanism of development of
atherosclerosis and cardiovascular disease in diabetes mellitus. J
Atheroscler Thromb. 25:27–39. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vidanapathirana AK, Psaltis PJ, Bursill
CA, Abell AD and Nicholls SJ: Cardiovascular bioimaging of nitric
oxide: Achievements, challenges, and the future. Med Res Rev.
41:435–463. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kristensen LS, Andersen MS, Stagsted LVW,
Ebbesen KK, Hansen TB and Kjems J: The biogenesis, biology and
characterization of circular RNAs. Nat Rev Genet. 20:675–691. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Holdt LM, Kohlmaier A and Teupser D:
Molecular roles and function of circular RNAs in eukaryotic cells.
Cell Mol Life Sci. 75:1071–1098. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhao L, Guo Y, Guo Y, Ji X, Fan D, Chen C,
Yuan W, Sun Z and Ji Z: Effect and mechanism of circRNAs in tumor
angiogenesis and clinical application. Int J Cancer. 150:1223–1232.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ye D, Gong M, Deng Y, Fang S, Cao Y, Xiang
Y and Shen Z: Roles and clinical application of exosomal circRNAs
in the diagnosis and treatment of malignant tumors. J Transl Med.
20:1612022. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kulcheski FR, Christoff AP and Margis R:
Circular RNAs are miRNA sponges and can be used as a new class of
biomarker. J Biotechnol. 238:42–51. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ryu J, Ahn Y, Kook H and Kim YK: The roles
of non-coding RNAs in vascular calcification and opportunities as
therapeutic targets. Pharmacol Ther. 218:1076752021. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ding P, Ding Y, Tian Y and Lei X: Circular
RNA circ_0010283 regulates the viability and migration of oxidized
low-density lipoprotein-induced vascular smooth muscle cells via an
miR-370-3p/HMGB1 axis in atherosclerosis. Int J Mol Med.
46:1399–1408. 2020.PubMed/NCBI
|
|
11
|
Wang G, Li Y, Liu Z, Ma X, Li M, Lu Q, Li
Y, Lu Z, Niu L, Fan Z and Lei Z: Circular RNA circ_0124644
exacerbates the ox-LDL-induced endothelial injury in human vascular
endothelial cells through regulating PAPP-A by acting as a sponge
of miR-149-5p. Mol Cell Biochem. 471:51–61. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang L, Zheng Z, Feng X, Zang X, Ding W,
Wu F and Zhao Q: circRNA/lncRNA-miRNA-mRNA network in oxidized,
low-density, lipoprotein-induced foam cells. DNA Cell Biol.
38:1499–1511. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yu XH, Fu YC, Zhang DW, Yin K and Tang CK:
Foam cells in atherosclerosis. Clin Chim Acta. 424:245–252. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Le Saux G, Plawinski L, Parrot C, Nlate S,
Servant L, Teichmann M, Buffeteau T and Durrieu MC: Surface bound
VEGF mimicking peptide maintains endothelial cell proliferation in
the absence of soluble VEGF in vitro. J Biomed Mater Res A.
104:1425–1436. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lu J, Mitra S, Wang X, Khaidakov M and
Mehta JL: Oxidative stress and lectin-like ox-LDL-receptor LOX-1 in
atherogenesis and tumorigenesis. Antioxid Redox Signal.
15:2301–2333. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huang F, Chen W, Peng J, Li Y, Zhuang Y,
Zhu Z, Shao C, Yang W, Yao H and Zhang S: LncRNA PVT1 triggers
Cyto-protective autophagy and promotes pancreatic ductal
adenocarcinoma development via the miR-20a-5p/ULK1 axis. Mol
Cancer. 17:982018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhu X, Li Z, Li T, Long F, Lv Y, Liu L,
Liu X and Zhan Q: Osthole inhibits the PI3K/AKT signaling pathway
via activation of PTEN and induces cell cycle arrest and apoptosis
in esophageal squamous cell carcinoma. Biomed Pharmacother.
102:502–509. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42:(Database Issue). D92–D97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cui J, Li W, Liu G, Chen X, Gao X, Lu H
and Lin D: A novel circular RNA, hsa_circ_0043278, acts as a
potential biomarker and promotes non-small cell lung cancer cell
proliferation and migration by regulating miR-520f. Artif Cells
Nanomed Biotechnol. 47:810–821. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhao W, Geng D, Li S, Chen Z and Sun M:
LncRNA HOTAIR influences cell growth, migration, invasion, and
apoptosis via the miR-20a-5p/HMGA2 axis in breast cancer. Cancer
Med. 7:842–855. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Peng L, Chen G, Zhu Z, Shen Z, Du C, Zang
R, Su Y, Xie H, Li H, Xu X, et al: Circular RNA ZNF609 functions as
a competitive endogenous RNA to regulate AKT3 expression by
sponging miR-150-5p in Hirschsprung's disease. Oncotarget.
8:808–818. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kato R, Mori C, Kitazato K, Arata S, Obama
T, Mori M, Takahashi K, Aiuchi T, Takano T and Itabe H: Transient
increase in plasma oxidized LDL during the progression of
atherosclerosis in apolipoprotein E knockout mice. Arterioscler
Thromb Vasc Biol. 29:33–39. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gisterå A and Hansson GK: The immunology
of atherosclerosis. Nat Rev Nephrol. 13:368–380. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kattoor AJ, Pothineni NVK, Palagiri D and
Mehta JL: Oxidative stress in atherosclerosis. Curr Atheroscler
Rep. 19:422017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Frostegård J, Haegerstrand A, Gidlund M
and Nilsson J: Biologically modified LDL increases the adhesive
properties of endothelial cells. Atherosclerosis. 90:119–126. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Catapano AL, Maggi FM and Tragni E: Low
density lipoprotein oxidation, antioxidants, and atherosclerosis.
Curr Opin Cardiol. 15:355–363. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Forstermann U and Sessa WC: Nitric oxide
synthases: Regulation and function. Eur Heart J. 33:829–837.
837a–837d. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Barbieri SS, Cavalca V, Eligini S,
Brambilla M, Caiani A, Tremoli E and Colli S: Apocynin prevents
cyclooxygenase 2 expression in human monocytes through NADPH
oxidase and glutathione redox-dependent mechanisms. Free Radic Biol
Med. 37:156–165. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hansson GK, Robertson AK and
Söderberg-Nauclér C: Inflammation and atherosclerosis. Annu Rev
Pathol. 1:297–329. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mietus-Snyder M, Friera A, Glass CK and
Pitas RE: Regulation of scavenger receptor expression in smooth
muscle cells by protein kinase C: A role for oxidative stress.
Arterioscler Thromb Vasc Biol. 17:969–978. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mitra S, Goyal T and Mehta JL: Oxidized
LDL, LOX-1 and atherosclerosis. Cardiovasc Drugs Ther. 25:419–429.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shen CM, Mao SJ, Huang GS, Yang PC and Chu
RM: Stimulation of smooth muscle cell proliferation by ox-LDL- and
acetyl LDL-induced macrophage-derived foam cells. Life Sci.
70:443–452. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Maiolino G, Rossitto G, Caielli P, Bisogni
V, Rossi GP and Calò LA: The role of oxidized low-density
lipoproteins in atherosclerosis: The myths and the facts. Mediators
Inflamm. 2013:7146532013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Podrez EA, Byzova TV, Febbraio M, Salomon
RG, Ma Y, Valiyaveettil M, Poliakov E, Sun M, Finton PJ, Curtis BR,
et al: Platelet CD36 links hyperlipidemia, oxidant stress and a
prothrombotic phenotype. Nat Med. 13:1086–1095. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Park HJ, Zhang Y, Georgescu SP, Johnson
KL, Kong D and Galper JB: Human umbilical vein endothelial cells
and human dermal microvascular endothelial cells offer new insights
into the relationship between lipid metabolism and angiogenesis.
Stem Cell Rev. 2:93–102. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yamada T, Fan J, Shimokama T, Tokunaga O
and Watanabe T: Induction of fatty streak-like lesions in vitro
using a culture model system simulating arterial intima. Am J
Pathol. 141:1435–1444. 1992.PubMed/NCBI
|
|
38
|
Burns MP and DePaola N: Flow-conditioned
HUVECs support clustered leukocyte adhesion by coexpressing ICAM-1
and E-selectin. Am J Physiol Heart Circ Physiol. 288:H194–H204.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kokura S, Wolf RE, Yoshikawa T, Granger DN
and Aw TY: Molecular mechanisms of neutrophil-endothelial cell
adhesion induced by redox imbalance. Circ Res. 84:516–524. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang W, DeMattia JA, Song H and Couldwell
WT: Communication between malignant glioma cells and vascular
endothelial cells through gap junctions. J Neurosurg. 98:846–853.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Plump AS, Smith JD, Hayek T, Aalto-Setälä
K, Walsh A, Verstuyft JG, Rubin EM and Breslow JL: Severe
hypercholesterolemia and atherosclerosis in apolipoprotein
E-deficient mice created by homologous recombination in ES cells.
Cell. 71:343–353. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Piedrahita JA, Zhang SH, Hagaman JR,
Oliver PM and Maeda N: Generation of mice carrying a mutant
apolipoprotein E gene inactivated by gene targeting in embryonic
stem cells. Proc Natl Acad Sci USA. 89:4471–4475. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Li H, Shen S, Chen X, Ren Z, Li Z and Yu
Z: miR-450b-5p loss mediated KIF26B activation promoted
hepatocellular carcinoma progression by activating PI3K/AKT
pathway. Cancer Cell Int. 19:2052019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ye YP, Wu P, Gu CC, Deng DL, Jiao HL, Li
TT, Wang SY, Wang YX, Xiao ZY, Wei WT, et al: miR-450b-5p induced
by oncogenic KRAS is required for colorectal cancer progression.
Oncotarget. 7:61312–61324. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Luo X, Wang W, Li D, Xu C, Liao B, Li F,
Zhou X, Qin W and Liu J: Plasma exosomal miR-450b-5p as a possible
biomarker and therapeutic target for transient ischaemic attacks in
rats. J Mol Neurosci. 69:516–526. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cartwright T, Perkins ND and L Wilson C:
NFKB1: A suppressor of inflammation, ageing and cancer. FEBS J.
283:1812–1822. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Fiordelisi A, Iaccarino G, Morisco C,
Coscioni E and Sorriento D: NFkappaB is a key player in the
crosstalk between inflammation and cardiovascular diseases. Int J
Mol Sci. 20:15992019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Hernández-Presa MA, Ortego M, Tuñón J,
Martín-Ventura JL, Mas S, Blanco-Colio LM, Aparicio C, Ortega L,
Gómez-Gerique J, Vivanco F and Egido J: Simvastatin reduces
NF-kappaB activity in peripheral mononuclear and in plaque cells of
rabbit atheroma more markedly than lipid lowering diet. Cardiovasc
Res. 57:168–177. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kumar A, Takada Y, Boriek AM and Aggarwal
BB: Nuclear factor-kappaB: Its role in health and disease. J Mol
Med (Berl). 82:434–448. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Huang Z, Mou T, Luo Y, Pu X, Pu J, Wan L,
Gong J, Yang H, Liu Y, Li Z, et al: Inhibition of miR-450b-5p
ameliorates hepatic ischemia/reperfusion injury via targeting
CRYAB. Cell Death Dis. 11:4552020. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Patel V, Carrion K, Hollands A, Hinton A,
Gallegos T, Dyo J, Sasik R, Leire E, Hardiman G, Mohamed SA, et al:
The stretch responsive microRNA miR-148a-3p is a novel repressor of
IKBKB, NF-κB signaling, and inflammatory gene expression in human
aortic valve cells. FASEB J. 29:1859–1868. 2015. View Article : Google Scholar : PubMed/NCBI
|