Introduction
Inflammatory bowel disease (IBD), including mainly
Crohn's disease (CD) and ulcerative colitis (UC), is an
immune-mediated chronic and relapsing inflammatory disorder
resulting from dysregulation of the mucosal immune response in the
gastrointestinal track (1). The
etiologies of IBD remain elusive as they involve complex
interactions between genetic, environmental, and immunoregulatory
factors (2). Furthermore,
patients with long-standing UC are prone to colorectal cancer
(CRC). Although colitis-associated CRC accounts for only 1–2% of
all CRC cases, IBD with colon involvement is among the top three
risk factors for CRC (3).
Therefore, researchers are interested in identifying target genes
for IBD to reduce the overall risk of colitis-associated CRC.
Macrophages have a central role in intestinal
homeostasis, being strategically positioned in the subepithelial
lamina propria to clear microbes that breach the epithelial
barrier. Activated macrophages prolifically secrete inflammatory
cytokines in the gut, and the imbalance of cytokines is an
important part of the pathogenesis of IBD (4). Lipopolysaccharide (LPS), a vital
biological ingredient of the Gram-negative bacterial cell wall, is
well known to trigger inflammation due to its ability to activate
macrophages (5). One major target
of LPS is toll-like receptor 4, which induces the activation of
nuclear factor-kappa B (NF-κB) (6). NF-κB activation promotes the
phosphorylation of IκB-α, which results in translocation of the
NF-κB subunit into the nucleus. Finally, the subunit binds to the
promoter regions of target genes and induces the transcription of
pro-inflammatory cytokines, including tumor necrosis factor-alpha
(TNF-α), interleukin-1β (IL-1β), and IL-6 (7).
Lipocalins (LCNs) are a family of proteins with
various functions, including regulation of immune responses, cell
growth, metabolism, iron transport, and prostaglandin synthesis
(8). LCN2, also known as oncogene
24p3 neutrophil gelatinase-associated lipocalin, is a 25-kDa
secreted glycoprotein that was initially purified from neutrophil
granules (9). LCN2 can bind
hydrophobic molecules, including retinoids, fatty acids, and
various steroids (10). The LCN2
promoter region contains the binding sites of several transcription
factors, including NF-κB, STAT1/3, CREB, and C/EBP-β, indicating a
potential role of LCN2 in regulating inflammation and metabolism
(11–13).
Emerging evidence has shown that LCN2 is highly
associated with inflammatory conditions, and that it plays a role
in modulating macrophage activation in the adipose tissue, lungs,
liver, and the brain (14–18).
Moreover, the upregulation of LCN2 in IBD patients with UC has
recently been reported, suggesting that LCN2 can be a sensitive
biomarker for intestinal inflammation (19,20). In a previous study, we suggested
that the pro-inflammatory cytokine IL-6 induces LCN2 promoter
activity via NF-κB/STAT3 activation, and that increased LCN2 may
serve as an indicator of CRC development in a chronic inflammation
setting (21). Considering that
LCN2 is associated with intestinal inflammation, IBD as well as the
CRC supportive microenvironment may rely, at least in part, on the
regulation of LCN2 in activated macrophages. However, the
underlying mechanism of LCN2 in activated macrophages as a target
for IBD has not been established. A new perspective of LCN2 under
inflammatory conditions would provide greater insight into its role
in the tumor microenvironment. The aim of this study was to
investigate the role and molecular mechanism of LCN2 in regulating
inflammation, especially inflammasome assembly, using the murine
macrophage cell line RAW264.7.
Materials and methods
Chemicals and reagents
LPS and parthenolide were purchased from
Sigma-Aldrich (St. Louis, MO). Bay 11-7085 was obtained from
Calbiochem (St. Louis, MO). LPS was dissolved in 1×
phosphate-buffered saline (PBS) to a concentration of 1 mg/ml and
stored at −20°C. Parthenolide and BAY 11-7082 were dissolved in
dimethylsulfoxide (Sigma-Aldrich) and stored at −20°C in the dark.
Dextran sulfate sodium (DSS) was purchased from MP Biochemicals
(Irvine, CA, USA).
IBD murine model
Forty specific-pathogen-free, female, six-week-old
BALB/c mice were purchased from Orient Bio, Inc. (Seongnam, Korea).
Mice were given ad libitum access to water and standard
rodent food until they reached the desired weight (18–20 g). They
were maintained on a 12/12 hour light/dark cycle under
specific-pathogen-free conditions. All procedures were reviewed and
approved by Jeonbuk National University Animal Care and Use
Committee (Approval no: CBNU 2018-001). Following the guideline of
Institutional Animal Care and Use Committee (IACUC), over 20% of
body weight loss of animal model in oncology studies is defined as
severe suffering for humane endpoint decisions, although other
signs were not detected. Death of mice was verified through
cessation of breathing and the loss of heartbeats.
Mice were randomly assigned to either the control
(n=10) or the IBD group (n=12) after they were weighed. The
drinking water of the IBD group was laced with 3% DSS for 7 days.
Following the guideline of EU, over 20% of body weight loss of
animal model in oncology studies is defined as severe suffering for
humane endpoint decisions, although other signs were not detected.
Following sacrifice of animals from both groups by cervical
dislocation, their body weights were measured. All colons were
removed from the cecum to the anus and their lengths were measured.
The colons were cut open longitudinally along the main axis and
washed with 1X PBS (pH 7.4). After gross examination, half of each
groups' specimens were fixed in 10% neutral-buffered formalin for
histological and immunohistochemical analyses. The remaining colons
were used for enzyme-linked immunosorbent assay (ELISA) and western
blot analyses.
Histological staining and
immunohistochemistry
Paraffin-embedded samples were cut into 5-µm
sections and stained with hematoxylin and eosin for light
microscopic examination. The sections were immunostained with
anti-LCN2/NGAL antibody (Abcam, Cambridge, UK, catalog number:
ab216462, dilution: 5 µg/ml), visualized by appropriate
biotin-conjugated secondary antibodies, followed by
immunoperoxidase detection with HRP/DAB (ABC) Detection IHC Kit
(Abcam, catalog number: ab64261). The colon sections were captured
using a Leica DM750 (Wetzlar, Germany) photomicroscope.
LCN2 ELISA
Mouse LCN2 ELISA Kits (R&D Systems) were used to
measure LCN2 levels in mouse colon tissue lysates, per the
manufacturer's instructions. All samples were assayed in duplicate
and compared with a standard curve to quantitate the expression
level.
Cell culture
The murine macrophage cell line RAW264.7 was
purchased from the American Type Culture Collection (Manassas, VA,
USA). Cells were cultured in Dulbecco's modified Eagle's medium
(Gibco®, Carlsbad, CA, USA), supplemented with 10% fetal
bovine serum (Gibco®), 100 units penicillin (Invitrogen,
Carlsbad, CA, USA), and 100 units streptomycin (Invitrogen,
Carlsbad, CA, USA), in a humidified 5% CO2 environment
at 37°C.
RNA isolation and reverse
transcription-PCR (RT-PCR)
Total RNA was isolated from cultured cells using
TRIzol reagent (Invitrogen, Eugene, OR) and cDNA was synthesized
using SuperScript II Reverse Transcriptase (Invitrogen), according
to the manufacturer's protocol. The expression of
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as a
loading control. The following primer sequences were used: mouse
LCN2, 5′-GGACCAGGGCTGTCGCTACT-3′ (forward) and
5′-GGTGGCCACTTGCACATTGT-3′ (reverse), GAPDH,
5′-ACCACAGTCCATGCCATCAC-3′ (forward) 5′-TCCACCACCCTGTTGCTGTA-3′
(reverse), generating a 120- and 452-bp product, respectively.
After initial denaturation at 95°C for 1 min, PCR was performed for
various cycles (30 sec at 94°C, 1 min at annealing temperature, and
2 min at 72°C) using Taq polymerase. Reaction products (10 µl) were
separated on 2% agarose gels and stained with RedSafe™ (Intron,
Daejeon, Korea). DNA band intensity was analyzed by densitometry
using an NαBI imager (Neogene Science, Suwon, Korea).
Small interfering RNA (siRNA) for
inhibiting specific gene expression
siRNA, used for targeted silencing of the mouse LCN2
(NCBI Ref Seq NM_008491) gene, was purchased from Santa Cruz
Biotechnology (catalog number: sc-60044, CA, USA). Specific siRNAs
and scrambled siRNA (for negative controls, catalog number: AM4637,
Invitrogen, TX, USA) were transfected into RAW264.7 cells using
TransIT-X2® transfection reagent (Mirus Bio, WI, USA).
Briefly, Opti-MEM I Reduced-Serum Medium, transfection reagent and
30 nM of siRNA were gently mixed and incubated at room temperature
for 30 min for complexes to form. The complexes were added
drop-wise to the cell culture plates and the cells were incubated
in 37°C incubator for 72 h. Then the cells were used further
experiments.
Protein extraction
Cells were harvested and resolved in a lysis buffer
(20 mM Tris-HCl (pH 7.5), 137 mM NaCl, 10% glycerol (v/v), 1%
Triton X-100 (v/v), 1 mM Na3VO4, 1 mM
phenylmethylsulfonyl fluoride, and protease inhibitor cocktail).
After centrifugation at 16,000 × g for 15 min, the supernatants
were used as cytoplasmic extracts. To extract the nuclear fraction,
cells were resuspended in 150 µl of buffer A [10 mM HEPES (pH 7.9),
1.5 mM MgCl2, 10 mM KCl, 0.5 mM dithiothreitol, 0.5 mM
phenylmethylsulfonyl fluoride, 0.4% Nonidet P-40 (v/v), and
protease inhibitor cocktail], incubated for 20 min on ice, and
centrifuged at 2,300 × g for 5 min. The resulting pellets were
resolved in 100 µl of buffer C [20 mM
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (pH 7.9), 420 mM
NaCl, 1.5 mM MgCl2, 0.2 mM ethylenediamine tetraacetic
acid, 0.5 mM dithiothreitol, 0.5 mM phenylmethylsulfonyl fluoride,
and protease inhibitor cocktail], incubated for 30 min on ice, and
centrifuged at 16,000 × g for 15 min. The resulting supernatants
were used as nuclear extracts.
For whole cell extracts, the cells were resolved by
RIPA buffer (50 mM Tris-HCl, 150 mM NaCl, 1% Triton X-100, 1%
sodium deoxycholate, 0.1% sodium dodecyl sulfate, and protease
inhibitors) and centrifuged at 13,200 rpm at 4°C for 30 min. The
resulting supernatants were used as whole cell extracts.
Western blotting
After extraction, the protein concentration was
measured using a Bradford Reagent (Sigma-Aldrich). Conditioned
media (100 µl) was collected and concentrated using a speed vacuum
concentrator (Thermo Scientific™, MA, USA). Fifty microgram of cell
protein and concentrated media were loaded onto 10~13.5% of sodium
dodecyl sulfate-polyacrylamide gels. After transferring and
blocking, each polyvinylidene difluoride membrane (PVDF, 0.2 µm,
Bio-rad, USA) was probed with various antibodies [anti-LCN2
(R&D systems, catalog number: AF1757, 1:2,000) anti-p-IκB-α
(Santa Cruz Biotechnology, catalog number: SC-8404, 1:1,000),
anti-p65 (Santa Cruz Biotechnology, catalog number: SC-8008,
1:1,000), anti-NLRP3 (Adipoge, catalog number; AG-20B-0014,
1:2,000), anti-ASC (GeneTex, catalog number: GTX105780, 1:2,000),
anti-caspase-1 p20 (Adepogen, catalog number: AG-20B-0042,
1:2,000), anti-IL-1β (R&D systems, catalog number: AF-401-NA,
1:2,000), anti-GAPDH (GeneTex, catalog number: GTX100118, 1:2,000)
and anti-Lamin B (Santa Cruz Biotechnology, catalog number:
SC-6216, 1:1,000)]. Antibody-antigen binding was detected using
enhanced ECL prime (GE Healthcare, NJ, USA), captured by FUSION FX
Image Analyzer (VILBER Lourmat, France) and analyzed by
Evolution-Capt software (VILBER Lourmat).
Inflammatory cytokine detection
RAW264.7 cells were stimulated with LPS and then
transfected with mouse LCN2 siRNA or scrambled siRNA. After the
indicated time, concentrations of the inflammatory cytokines TNF-α
(catalog number: 88-7324), IL-18 (catalog number: BMS618-3), and
IL-1β (catalog number 88-7013) in the conditioned media were
determined using corresponding commercial ELISA kits
(Invitrogen).
Statistical analyses
The data are presented as the mean ± standard error
of at least three independent experiments performed in duplicate.
Microsoft Excel 5.0, and Graphpad Prism 5.0 was used to perform
two-tailed t-tests for in vivo experiments, and analysis of
variance (ANOVA) for in vitro ELISA assay. The differences
between groups were compared by the analysis of variance, followed
by post hoc Tukey's test to correct for multiple comparisons. A
P-value <0.05 was considered statistically significant.
Results
LCN2 levels are highly elevated in
DSS-induced colitis models
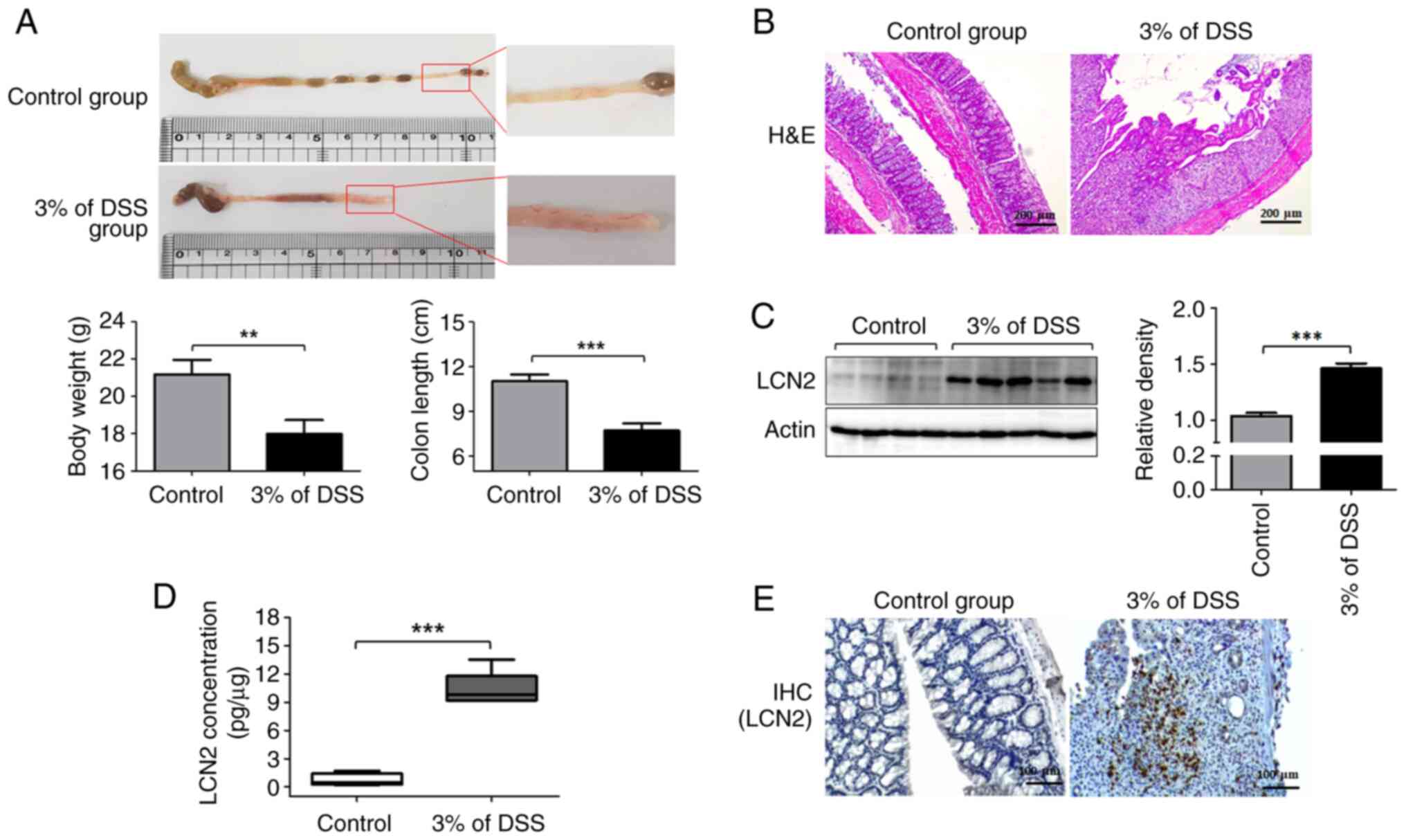
To assess whether the expression of LCN2 in the
colon differs between the control and the IBD groups, we analyzed
their pathological symptoms and colonic LCN2 levels. The severity
of colonic inflammation is strongly correlated with body weight
loss. Compared with the control group, mice with DSS-induced
colitis exhibited significant weight loss by ~15.05±3.49% (Fig. 1A left graph). Next, the entire
colons were collected from these mice to examine the gross
morphological damage. The colonic mucosal lesions in the colitis
group were obvious, with a wide range of hyperemia, edema, and
bleeding, compared with those in the control group (Fig. 1A). The colons in the colitis group
were considerably shorter than those in the control group (Fig. 1A right graph). Furthermore,
histological evaluation showed that colonic tissues from the
colitis group were characterized by epithelial cell destruction,
crypt deformation, ulceration, and inflammatory cells infiltrating
the lamina propria and submucosa (Fig. 1B).
Next, we examined the LCN2 protein levels in colon
tissues using western blotting (Fig.
1C). LCN2 levels were higher in the colon tissues from the
colitis group than in those from the control group (Fig. 1C). The ELISA results also
indicated that LCN2 levels in mice with colitis (10.35±1.821 pg/µg
of protein) were 12-fold higher than in healthy mice (0.699±0.673
pg/µg of protein; Fig. 1D).
Finally, immunohistochemistry also revealed more LCN2-positive
cells in the colitis group than in the control group, in line with
the western blotting and ELISA results (Fig. 1E).
LCN2 is specifically stimulated by LPS
in murine macrophages
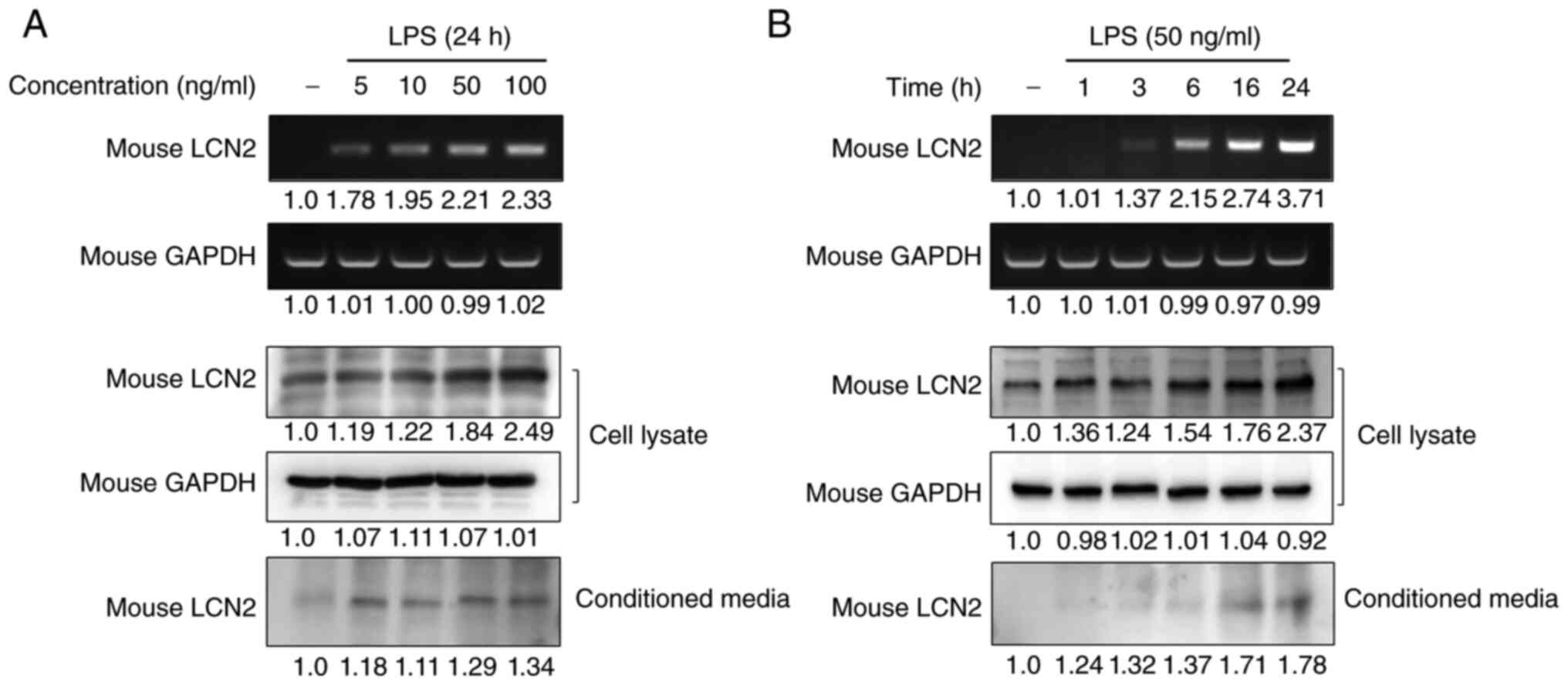
To investigate whether macrophages are one of the
main cellular targets for LCN2 regulation in IBD, we analyzed the
gene and protein expression of LCN2 in RAW264.7 macrophages after
stimulating them with LPS. LPS strongly induced LCN2 mRNA and
protein levels in a dose-dependent manner in RAW264.7 cells
(Fig. 2A). Moreover, stimulation
for 24 h with the lowest concentration of LPS led to increased LCN2
release in the conditioned media of RAW264.7 cells.
To confirm the time-dependency of this effect,
RAW264.7 cells were stimulated with LPS for the indicated time
periods and the expression and secretion of LCN2 were measured
(Fig. 2B). We found that LPS
induced the intracellular expression and secretion of LCN2 in a
time-dependent manner. These results indicate that the inflammatory
mediator LPS strongly induces LCN2 gene transcription, translation,
and secretion in macrophages.
NF-ĸB activation is required for
LPS-induced LCN2 expression
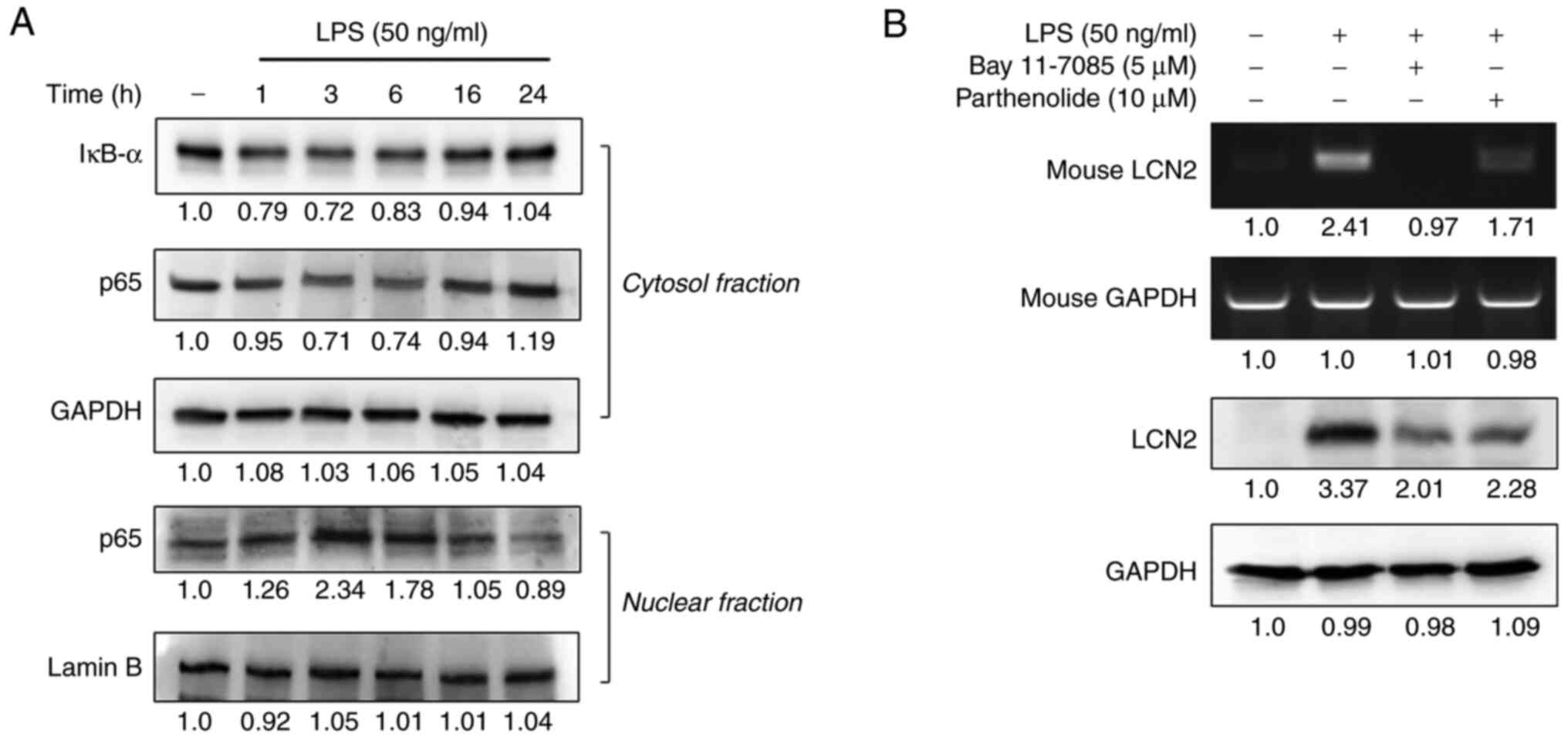
Exposing macrophages to LPS activates the
transcription factor NF-κB, which regulates the transcription of
various inflammation-responsive genes (22–24). After determining that 50 ng/ml of
LPS maximally induced LCN2 in macrophages, we first assessed the
degradation of IĸB-α and the nuclear translocation of NF-ĸB in
RAW264.7 cells in response to LPS stimulation for 1–24 h.
LPS-induced LCN2 decreased the expression of IĸB-α and p65 in the
cytosolic fraction between 1~6 h (Fig. 3A). In contrast, the expression of
p65 in the nuclear fraction was the greatest at the 3 h time point
and decreased slightly thereafter.
Next, we examined whether the expression of LCN2
depended upon NF-κB activation. Pretreatment with the
NF-ĸB-specific inhibitors BAY 11-7082 and parthenolide suppressed
the expression of LCN2 in LPS-stimulated RAW264.7 cells (Fig. 3B), suggesting that NF-ĸB
activation is required for LCN2 expression in LPS-stimulated
macrophages.
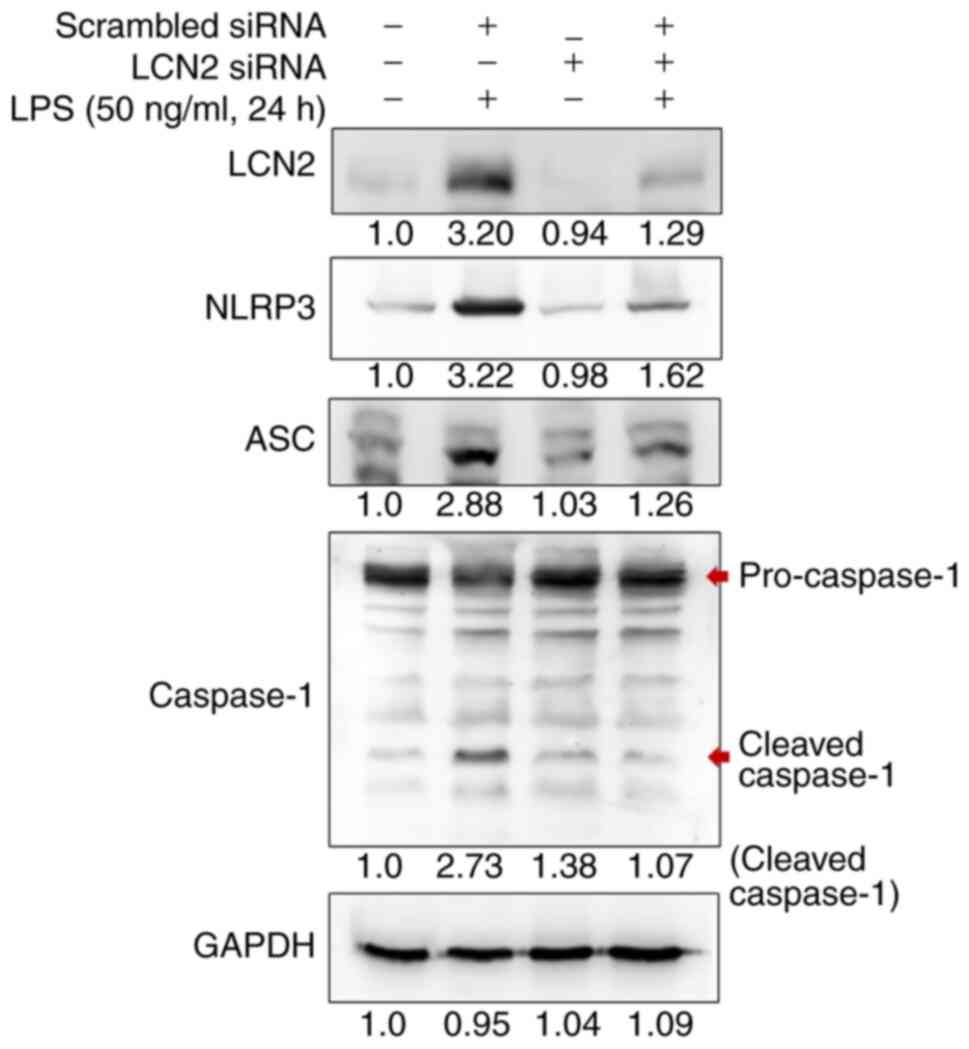
LCN2 regulates activation of the NLRP3
inflammasome in LPS-stimulated murine macrophages
The well-characterized NLRP3 (NOD-, LRR-, and pyrin
domain-containing protein 3) inflammasome is a multiprotein complex
that orchestrates an inflammatory response, which contributes to
IBD through increased epithelial permeability and a detrimental
immune response against invading commensal bacteria in the gut
(25). NF-κB is a central
mediator of the priming signal that activates the NLRP3
inflammasome (26). Thus, we
investigated whether the NLRP3 inflammasome was influenced by LCN2
in LPS-stimulated RAW264.7 cells (Fig. 4). After scrambled siRNA
transfection and LPS stimulation, the protein levels of NLRP3, ASC,
and cleaved Caspase 1 were strikingly augmented. Interestingly, the
expression of these inflammasome-related proteins was dramatically
attenuated when siRNA targeting LCN2 was used instead of the
scrambled siRNA, suggesting that LCN2 regulates the activation of
the NLRP3 inflammasome in LPS-stimulated macrophages.
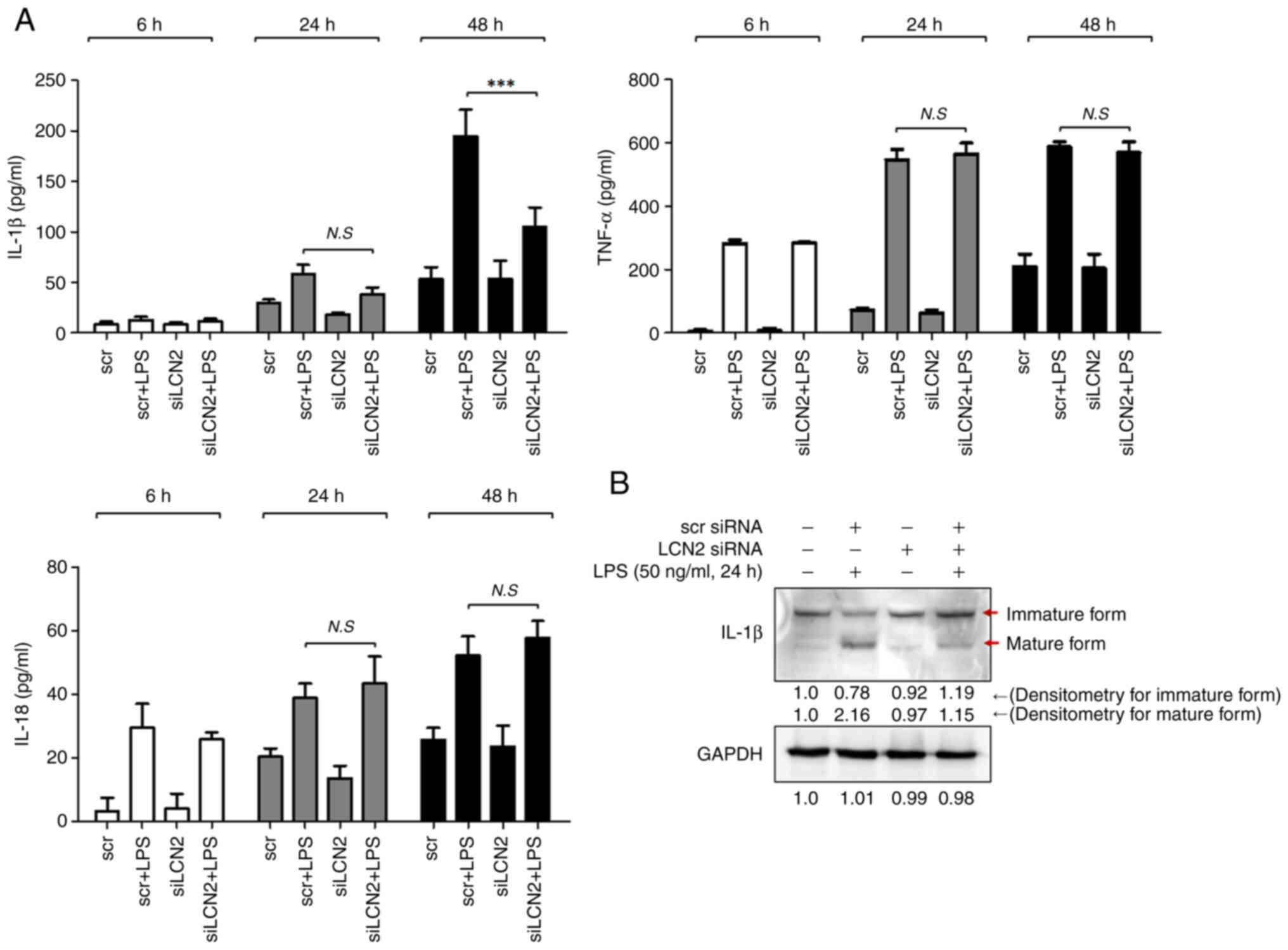
LCN2 is responsible for the production
of IL-1β in LPS-stimulated murine macrophages
The activated NLRP3 inflammasome serves as a
platform for the activation of caspase-1, which induces maturation
of the inflammatory cytokines IL-1β and IL-18 (27). Thus, we first measured the
concentrations of these two cytokines in media from RAW264.7 cell
cultures stimulated with LPS for 6, 24, and 48 h. After 48 h of LPS
stimulation, the concentration of IL-1β was markedly increased in
RAW264.7 cells, which was prominently suppressed by LCN2 silencing
(Fig. 5A). However, LCN2
silencing did not affect the LPS-induced IL-18 levels.
We confirmed the above observation by measuring the
intracellular IL-1β protein levels by western blotting. LPS
dramatically induced the matured form of the IL-1β protein, which
was diminished when LCN2 was silenced in RAW264.7 cells (Fig. 5B). These findings indicate that
LCN2 is involved in the maturation and secretion of the
pro-inflammatory cytokine IL-1β in LPS-stimulated macrophages.
Discussion
The pathogenesis of IBD is unknown, but the
uncontrolled immune response of genetically predisposed individuals
to environmental factors is considered to be the cause (28). In the present study, we
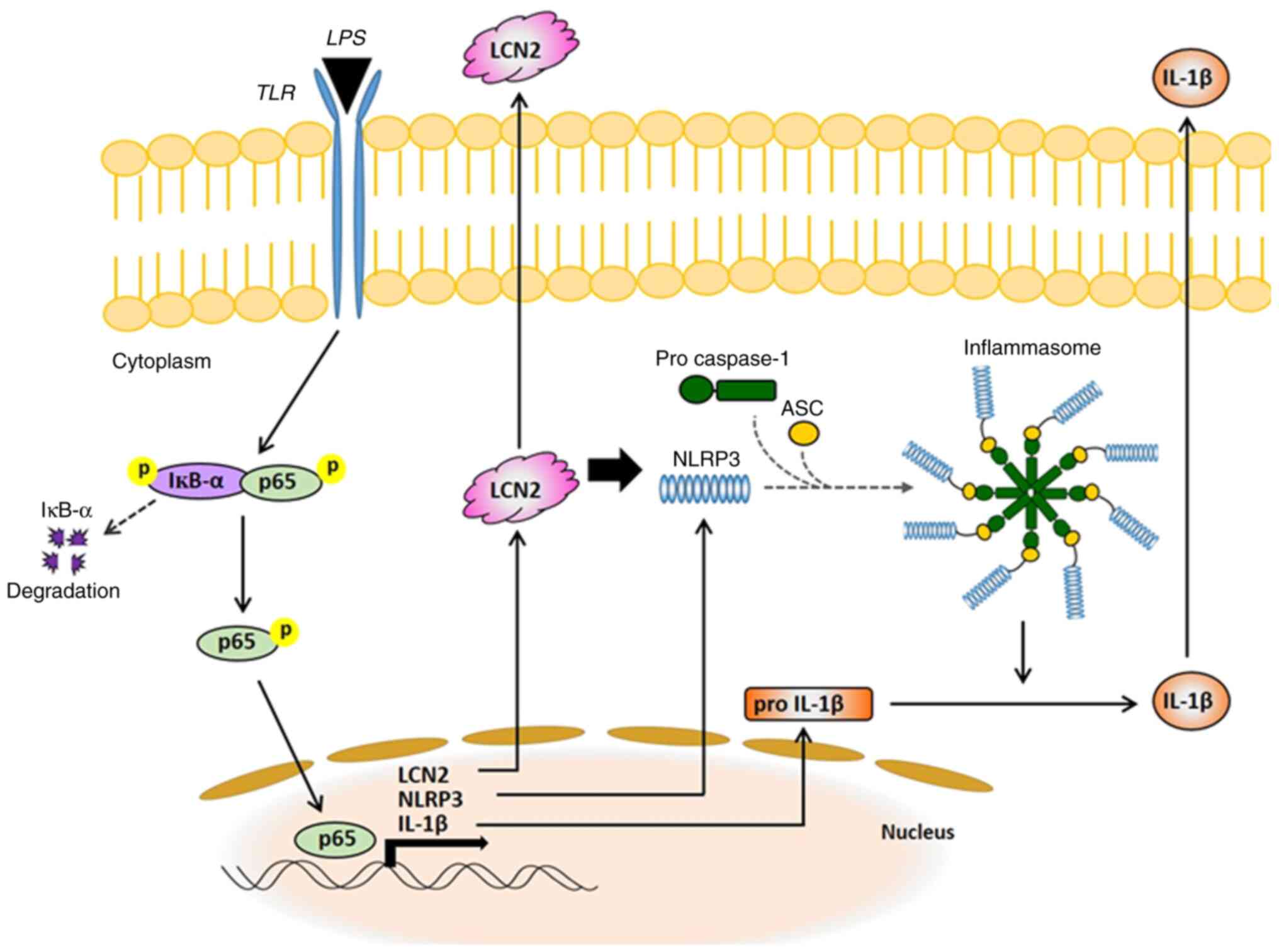
demonstrated that LCN2 in activated macrophages is a powerful
mediator of the NLRP3 inflammasome via NF-ĸB signaling and is
eventually responsible for secretion of the pro-inflammatory
cytokine IL-1β. Based on our results, we propose a new molecular
mechanism explaining the pro-inflammatory role of LCN2 in activated
macrophages (Fig. 6), thereby,
increasing our understanding of IBD.
Interestingly, LCN2 in IBD may serve as a biomarker
to distinguish active from inactive inflammation. In 2011,
Oikonomou et al suggested that serum LCN2 levels are
particularly elevated in patients with active IBD, correlating with
established markers of inflammation and disease activity (29). Moreover, Østvik et al
showed that LCN2 staining of colonic biopsies was significantly
increased in cases of active UC and CD compared with inactive UC
and CD (30). Our in vivo
experiments also demonstrate that LCN2 levels were augmented in
colonic tissues from DSS-induced animal models. These findings
suggest that LCN2 is involved in the pathophysiology of IBD, and is
associated with activity and severity of the disease.
When the intestine is invaded by pathogens that can
cross the damaged epithelial cell barrier, the intrinsic defense
cells in the epithelium, especially macrophages, produce and
secrete pro-inflammatory cytokines (31–33). LPS, a component of the microbial
pathogens, can directly activate macrophages and trigger
inflammation (34). Hence,
attenuating the LPS-induced inflammatory response in macrophages is
considered an attractive therapeutic strategy for numerous acute
and chronic inflammatory diseases. Emerging evidence has shown that
LCN2 plays a role in regulating LPS-stimulated macrophages. Guo
et al and Du et al showed that LCN2 was an essential
factor in the inflammatory response of macrophages to LPS (35,36). Likewise, our results (Fig. 2) confirm that LPS strongly induces
LCN2 expression in macrophages.
The transcription factor NF-ĸB, induced in
macrophages by LPS, controls the expression of numerous target
genes, such as inducible nitric oxide synthase, cyclooxygenase-2,
TNF-α, IL-1β, IL-18, and IL-6, all of which play important roles in
immune and inflammatory responses (37). Activated NF-κB has been detected
in lamina propria macrophages, epithelial cells from biopsy
specimens, and cultured cells from IBD patients (38). Therefore, NF-κB is a potential
target for IBD therapy, and nonsteroidal anti-inflammatory drugs,
such as 5-amino salicylic acid, inhibit the NF-ĸB signaling pathway
(39). Although substantial
evidence exists for the involvement of NF-κB in IBD, further
investigation is needed to elucidate the underlying mechanisms and
the precise role of NF-κB-related molecules in IBD pathogenesis. In
the present study, we suggest that LPS-induced upregulation of LCN2
strongly depends upon NF-κB activation, and LCN2 mediates the
NF-κB-induced NLRP3 inflammasome complex.
LCN2 is widely known to play various biological
roles; however, its specific role in different inflammatory
conditions is inconsistent. For example, LCN2 was reported to play
a pro-inflammatory role in autoimmune encephalomyelitis models of
neuroinflammation (40).
Moreover, it was shown to exert both pro-inflammatory and
iron-sequestering effects along the respiratory mucosa in response
to bacterial enterobactin (41).
In contrast, Du et al used LCN2 knockout mice to show that
it alleviates LPS-induced gut inflammation (36). Guo Guo et al also reported
that LCN2 plays an anti-inflammatory role in macrophage
polarization, potentially by modulating the activation of the
NF-κB-STAT3 loop (35). Our
results indicate that LPS-induced LCN2 in macrophages drives severe
inflammation by enhancing inflammasome assembly and IL-1β
secretion. Through our present study, we demonstrate the critical
role of LCN2 in IBD pathogenesis.
The NLRP3 inflammasome, a cytosolic multiprotein
complex, is well known to be involved in the development and
progression of numerous inflammatory disorders (42). Once activated by stimuli such as
LPS, NLRP3 recruits the adaptor protein ASC to promote recruitment
and activation of caspase-1, resulting in the production and
maturation of inflammatory cytokines (43). NF-κB is a central mediator of the
priming signal of NLRP3 inflammasome activation and induces the
transcriptional expression of NLRP3 (44). The function of LCN2 in
NF-κB-mediated activation of the NLRP3 inflammasome was
investigated in experimental models of heart failure in 2017. Song
E. and colleagues reported that LCN2 activates the NLRP3
inflammasome by inducing the release of the high-mobility group box
1 protein in primary cardiac fibroblasts from neonatal rats
(45). Consistent with these
findings, we found that LCN2 regulates LPS-induced NLRP3
inflammasome assembly in macrophages, which contributes to IBD
pathogenesis.
The NLRP3 inflammasome is critical for the
processing and maturation of IL-1β and is rapidly emerging as a
crucial regulator of intestinal homeostasis (46,47). Activated NF-κB transcriptionally
induces the expression of the 35-kDa pro-IL-1β, but the maturation
and secretion of 17-kDa IL-1β requires the activation of the NLRP3
inflammasome (48,49). Elevated levels of IL-1β were found
in colonic biopsies and isolated myeloid cells, correlating with
the severity of IBD (50). In the
present study, we observed that LCN2 silencing strongly repressed
IL-1β maturation and secretion in LPS-stimulated macrophages,
indicating that LCN2 regulates IL-1β production and maturation via
NF-κB/NLRP3 inflammasome signaling in macrophages.
In addition, IL-1β, produced mainly by the activated
NLRP3 inflammasome, can promote tumorigenesis in cancer cells
(51). Several publications have
demonstrated that IL-1β-specific upregulation of LCN2 is controlled
by the activation of NF-κB in cancer cells. Cowland et al
reported that stimulating lung cancer cells with IL-1β induced
NF-κB to bind to the LCN2 promoter and upregulate its expression
(52). In 2006, they also
reported that specific induction of LCN2 by IL-1β in lung
adenocarcinoma cells critically depends on activation of the NF-κB
cofactor IκB-ζ (53).
Collectively, this evidence suggests that upregulation of LCN2 in
macrophages results in the production and secretion of IL-1β to the
outer membrane of macrophages, where it subsequently induces
NF-κB-mediated LCN2 expression in cancer cells, leading to cancer
development and carcinogenesis. Therefore, the positive feedback
between LCN2 and IL-1β most likely contributes to the pathogenesis
of inflammation-driven cancer.
IL-18, an IL-1 family member, has pleiotropic
functions and is also chronically elevated in several inflammatory
diseases (54). Like IL-1β, the
production and release of IL-18 also depends on inflammasome
activation. Interestingly, IL-1β is only induced in response to
inflammatory stimuli, but IL-18 is constitutively expressed. In
other words, a cellular repository of IL-18 already exists even
before an inflammatory stimulus, and is ready to be activated and
released by the inflammasome (55,56). However, how different inflammatory
signals regulate the expression of IL-18 and IL-1β remains poorly
studied. In 2017, Zhu et al reported that type I interferon
(IFN) signaling is required for the induction of IL-18, but not
IL-1β, which points to a critical and differential role for type I
IFN in regulating IL-18 signaling (57). A correlation between LCN2 and
IL-18 has not directly been demonstrated; however, Østvik et
al showed that LCN2 in intestinal epithelial cells is not
affected by IFN-γ stimulation (30). In the present study, we observed
that LCN2 modulated the levels of IL-1β but did not affect
LPS-induced IL-18. Collectively, LCN2 is mostly irrelevant to
IFN-dependent IL-18 in macrophages. However, further study is
necessary to understand the interactions between LCN2 and
IFN-dependent IL-18 in macrophages and IBD pathogenesis.
In conclusion, our results show that LCN2 directly
upregulates the NLRP3 inflammasome complex by NF-κB activation in
response to stimulation of macrophages by LPS, consequently leading
to the production and maturation of IL-1β. We have presented novel
findings on the role and mechanism of LCN2 in IBD pathophysiology.
Moreover, this study clearly highlights the role of LCN2 as a
potential target for the NLRP3 inflammasome in the field of IBD
therapy.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Research Foundation of
Korea (NRF) grant funded by the Korea government (MSIT; grant no.
2020R1C1C1012108) and the Fund of the Biomedical Research Institute
of Jeonbuk National University Hospital (grant no. CUH2021-0041) in
Korea.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
SWK participated in the design of the study. SLK and
MWS carried out the in vitro and in vivo experiments.
SWK and SLK confirm the authenticity of all the raw data. SLK wrote
the draft and performed the statistical analysis. SWK edited the
draft and supervised all experimental procedures. All authors have
read and approved the final manuscript.
Ethics approval and consent to
participate
All procedures of animal experiments were reviewed
and approved by Jeonbuk National University Animal Care and Use
Committee (approval no. CBNU 2018-001). Animal experiments were
performed in strict compliance with European guidelines and
regulations on protection of animals used for scientific purposes
(EC Directive 2010/63/EU).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Khor B, Gardet A and Xavier RJ: Genetics
and pathogenesis of inflammatory bowel disease. Nature.
474:307–317. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sartor RB: Mechanisms of disease:
Pathogenesis of Crohn's disease and ulcerative colitis. Nat Clin
Pract Gastroenterol Hepatol. 3:390–407. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lennard-Jones JE, Morson BC, Ritchie JK
and Williams CB: Cancer surveillance in ulcerative colitis.
Experience over 15 years. Lancet. 2:149–152. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sanchez-Munoz F, Dominguez-Lopez A and
Yamamoto-Furusho JK: Role of cytokines in inflammatory bowel
disease. World J Gastroenterol. 14:4280–4288. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gioannini TL and Weiss JP: Regulation of
interactions of Gram-negative bacterial endotoxins with mammalian
cells. Immunol Res. 39:249–260. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Krappmann D, Wegener E, Sunami Y, Esen M,
Thiel A, Mordmuller B and Scheidereit C: The IkappaB kinase complex
and NF-kappaB act as master regulators of
lipopolysaccharide-induced gene expression and control subordinate
activation of AP-1. Mol Cell Biol. 24:6488–6500. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kueanjinda P, Roytrakul S and Palaga T: A
novel role of numb as a regulator of pro-inflammatory cytokine
production in macrophages in response to toll-like receptor 4 (vol
5, 12784, 2015). Sci Rep. 5:127842015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tong ZM, Wu XL, Ovcharenko D, Zhu JX, Chen
CS and Kehrer JP: Neutrophil gelatinase-associated lipocalin as a
survival factor. Biochem J. 391:441–448. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kjeldsen L, Johnsen AH, Sengelov H and
Borregaard N: Isolation and primary structure of ngal, a novel
protein associated with human neutrophil gelatinase. J Biol Chem.
268:10425–10432. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Akerstrom B, Flower DR and Salier JP:
Lipocalins: Unity in diversity. Biochim Biophys Acta. 1482:1–8.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shen F, Hu Z, Goswami J and Gaffen SL:
Identification of common transcriptional regulatory elements in
interleukin-17 target genes. J Biol Chem. 281:24138–24148. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lawrence T and Natoli G: Transcriptional
regulation of macrophage polarization: Enabling diversity with
identity. Nat Rev Immunol. 11:750–761. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao P and Stephens JM: STAT1, NF-κB and
ERKs play a role in the induction of lipocalin-2 expression in
adipocytes. Mol Metab. 2:161–170. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Guo H, Jin D and Chen X: Lipocalin 2 is a
regulator of macrophage polarization and NF-κB/STAT3 pathway
activation. Mol Endocrinol. 28:1616–1628. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Warszawska JM, Gawish R, Sharif O, Sigel
S, Doninger B, Lakovits K, Mesteri I, Nairz M, Boon L, Spiel A, et
al: Lipocalin 2 deactivates macrophages and worsens pneumococcal
pneumonia outcomes. J Clin Invest. 123:3363–3372. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Borkham-Kamphorst E, de Leur EV,
Zimmermann HW, Karlmark KR, Tihaa L, Haas U, Tacke F, Berger T, Mak
TW and Weiskirchen R: Protective effects of lipocalin-2 (LCN2) in
acute liver injury suggest a novel function in liver homeostasis.
Biochim Biophys Acta. 1832:660–673. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Srinivasan G, Aitken JD, Zhang BY,
Carvalho FA, Chassaing B, Shashidharamurthy R, Borregaard N, Jones
DP, Gewirtz AT and Vijay-Kumar M: Lipocalin 2 deficiency
dysregulates iron homeostasis and exacerbates endotoxin-induced
sepsis. J Immunol. 189:1911–1919. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang YY, Foncea R, Deis JA, Guo H,
Bernlohr DA and Chen XL: Lipocalin 2 expression and secretion is
highly regulated by metabolic stress, cytokines, and nutrients in
adipocytes. Plos One. 9:e969972014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chassaing B, Srinivasan G, Delgado MA,
Young AN, Gewirtz AT and Vijay-Kumar M: Fecal lipocalin 2, a
sensitive and broadly dynamic non-invasive biomarker for intestinal
inflammation. Plos One. 7:e443282012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nielsen OH, Gionchetti P, Ainsworth M,
Vainer B, Campieri M, Borregaard N and Kjeldsen L: Rectal dialysate
and fecal concentrations of neutrophil gelatinase-associated
lipocalin, interleukin-8, and tumor necrosis factor-alpha in
ulcerative colitis. Am J Gastroenterol. 94:2923–2928. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim SL, Shin MW, Seo SY and Kim SW:
Lipocalin 2 potentially contributes to tumorigenesis from colitis
via IL-6/STAT3/NF-κB signaling pathway. Biosci Rep.
42:BSR202124182022. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lawrence T: The nuclear factor NF-kappaB
pathway in inflammation. Cold Spring Harb Perspect Biol.
1:a0016512009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
O'Dea E and Hoffmann A: NF-κB signaling.
Wiley Interdiscip Rev Syst Biol Med. 1:107–115. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sharif O, Bolshakov VN, Raines S, Newham P
and Perkins ND: Transcriptional profiling of the LPS induced
NF-kappaB response in macrophages. BMC Immunol. 8:12007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Allen IC, TeKippe EM, Woodford RM, Uronis
JM, Holl EK, Rogers AB, Herfarth HH, Jobin C and Ting JPY: The
NLRP3 inflammasome functions as a negative regulator of
tumorigenesis during colitis-associated cancer. J Exp Med.
207:1045–1056. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu T, Zhang L, Joo D and Sun SC: NF-κB
signaling in inflammation. Signal Transduct Target Ther.
2:170232017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Swanson KV, Deng M and Ting JP: The NLRP3
inflammasome: Molecular activation and regulation to therapeutics.
Nat Rev Immunol. 19:477–489. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Han X, Ding S, Jiang H and Liu G: Roles of
macrophages in the development and treatment of gut inflammation.
Front Cell Dev Biol. 9:6254232021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Oikonomou KA, Kapsoritakis AN, Theodoridou
C, Karangelis D, Germenis A, Stefanidis I and Potamianos SP:
Neutrophil gelatinase-associated lipocalin (NGAL) in inflammatory
bowel disease: Association with pathophysiology of inflammation,
established markers, and disease activity. J Gastroenterol.
47:519–530. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ostvik AE, Granlund AV, Torp SH, Flatberg
A, Beisvag V, Waldum HL, Flo TH, Espevik T, Damås JK and Sandvik
AK: Expression of toll-like receptor-3 is enhanced in active
inflammatory bowel disease and mediates the excessive release of
lipocalin 2. Clin Exp Immunol. 173:502–511. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Baumgart DC and Carding SR: Inflammatory
bowel disease: Cause and immunobiology. Lancet. 369:1627–1640.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Geremia A, Biancheri P, Allan P, Corazza
GR and Di Sabatino A: Innate and adaptive immunity in inflammatory
bowel disease. Autoimmun Rev. 13:3–10. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Geremia A and Arancibia-Carcamo CV: Innate
lymphoid cells in intestinal inflammation. Front Immunol.
8:12962017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
West MA, Seatter SC, Bellingham J and
Clair L: Mechanisms of reprogrammed macrophage endotoxin signal
transduction after lipopolysaccharide pretreatment. Surgery.
118:220–228. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Guo H, Jin D and Chen X: Lipocalin 2 is a
regulator of macrophage polarization and NF-κB/STAT3 pathway
activation. Mol Endocrinol. 28:1616–1628. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Du H, Liang L, Li J, Xiong Q, Yu X and Yu
H: Lipocalin-2 Alleviates LPS-induced inflammation through
alteration of macrophage properties. J Inflamm Res. 14:4189–4203.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li Q and Verma IM: NF-kappaB regulation in
the immune system. Nat Rev Immunol. 2:725–734. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Rogler G, Brand K, Vogl D, Page S,
Hofmeister R, Andus T, Knuechel R, Baeuerle PA, Schölmerich J and
Gross V: Nuclear factor kappaB is activated in macrophages and
epithelial cells of inflamed intestinal mucosa. Gastroenterology.
115:357–369. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang Z, Liu N and Chen X, Zhang F, Kong
T, Tang X, Yang Q, Chen W, Xiong X and Chen X: UCHL1 regulates
inflammation via MAPK and NF-κB pathways in LPS-activated
macrophages. Cell Biol Int. 45:2107–2117. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nam Y, Kim JH, Seo M, Kim JH, Jin M, Jeon
S, Seo JW, Lee WH, Bing SJ, Jee Y, et al: Lipocalin-2 protein
deficiency ameliorates experimental autoimmune encephalomyelitis:
The pathogenic role of lipocalin-2 in the central nervous system
and peripheral lymphoid tissues. J Biol Chem. 289:16773–16789.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bachman MA, Miller VL and Weiser JN:
Mucosal lipocalin 2 has pro-inflammatory and iron-sequestering
effects in response to bacterial enterobactin. PLoS Pathog.
5:e10006222009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Coll RC, Robertson AA, Chae JJ, Higgins
SC, Munoz-Planillo R, Inserra MC, Vetter I, Dungan LS, Monks BG,
Stutz AJ, et al: A small-molecule inhibitor of the NLRP3
inflammasome for the treatment of inflammatory diseases. Nat Med.
21:248–255. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Abderrazak A, Syrovets T, Couchie D, El
Hadri K, Friguet B, Simmet T and Rouis M: NLRP3 inflammasome: From
a danger signal sensor to a regulatory node of oxidative stress and
inflammatory diseases. Redox Biol. 4:296–307. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sutterwala FS, Haasken S and Cassel SL:
Mechanism of NLRP3 inflammasome activation. Ann N Y Acad Sci.
1319:82–95. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Song E, Jahng JW, Chong LP, Sung HK, Han
M, Luo C, Wu D, Boo S, Hinz B, Cooper MA, et al: Lipocalin-2
induces NLRP3 inflammasome activation via HMGB1 induced TLR4
signaling in heart tissue of mice under pressure overload
challenge. Am J Transl Res. 9:2723–2735. 2017.PubMed/NCBI
|
|
46
|
Zaki MH, Lamkanfi M and Kanneganti TD: The
Nlrp3 inflammasome: Contributions to intestinal homeostasis. Trends
Immunol. 32:171–179. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Haneklaus M, Gerlic M, O'Neill LA and
Masters SL: MiR-223: Infection, inflammation and cancer. J Intern
Med. 274:215–226. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Mariathasan S and Monack DM: Inflammasome
adaptors and sensors: Intracellular regulators of infection and
inflammation. Nat Rev Immunol. 7:31–40. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Martinon F, Burns K and Tschopp J: The
inflammasome: A molecular platform triggering activation of
inflammatory caspases and processing of proIL-beta. Mol Cell.
10:417–426. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Reinecker HC, Steffen M, Witthoeft T,
Pflueger I, Schreiber S, MacDermott RP and Raedler A: Enhanced
secretion of tumour necrosis factor-alpha, IL-6, and IL-1 beta by
isolated lamina propria mononuclear cells from patients with
ulcerative colitis and Crohn's disease. Clin Exp Immunol.
94:174–181. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Klampfer L: Cytokines, inflammation and
colon cancer. Curr Cancer Drug Targets. 11:451–464. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Cowland JB, Sorensen OE, Sehested M and
Borregaard N: Neutrophil gelatinase-associated lipocalin is
up-regulated in human epithelial cells by IL-1 beta, but not by
TNF-alpha. J Immunol. 171:6630–6639. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Cowland JB, Muta T and Borregaard N:
IL-1beta-specific up-regulation of neutrophil gelatinase-associated
lipocalin is controlled by IkappaB-zeta. J Immunol. 176:5559–5566.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Canna SW, Girard C, Malle L, de Jesus A,
Romberg N, Kelsen J, Surrey LF, Russo P, Sleight A, Schiffrin E, et
al: Life-threatening NLRC4-associated hyperinflammation
successfully treated with IL-18 inhibition. J Allergy Clin Immunol.
139:1698–16701. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Garlanda C, Dinarello CA and Mantovani A:
The interleukin-1 family: Back to the future. Immunity.
39:1003–1018. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Sims JE and Smith DE: The IL-1 family:
Regulators of immunity. Nat Rev Immunol. 10:89–102. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhu Q and Kanneganti TD: Cutting edge:
Distinct regulatory mechanisms control proinflammatory cytokines
IL-18 and IL-1β. J Immunol. 198:4210–4215. 2017. View Article : Google Scholar : PubMed/NCBI
|