Introduction
Ischemic stroke caused by cerebral artery embolism
is characterized by high morbidity (158/100,000 each year)
(1), mortality (11.6% of total
deaths) (2), and it accounts for
~80% of all strokes (3). At
present, restoring blood flow and oxygen in the ischemic area is
the primary clinical treatment for patients with ischemic stroke
(4). However, the process of
restoring blood flow can lead to severe secondary damage or
cerebral ischemia-reperfusion (I/R) injury (CIRI) (5), including aggravating a series of
pathological reactions, such as oxidative stress, neuroinflammation
and apoptosis, which inhibits treatment of ischemic stroke
(6). Therefore, improving CIRI is
a feasible way to treat ischemic stroke. It is reported that the
death of neurons is predominantly caused by hypoxia and
insufficient supply of glucose, which is closely related to
mitochondrial damage (7). Several
studies have shown that protecting mitochondrial function can
improve disorders in energy metabolism in vitro and inhibit
apoptosis, which serves an essential role in the treatment of
oxygen-/glucose-deprivation/reoxygenation in cell models and in
middle cerebral artery occlusion and reperfusion (MCAO/R) model
rats in vivo (8–10). The results suggest that maintaining
the health of mitochondria can improve CIRI and is of great
significance for treating ischemic stroke.
In recent years, it has been discovered that during
the treatment of cerebral ischemia, damaged mitochondria can be
eliminated by promotion of mitophagy, thus maintaining a healthy
mitochondrial network (11).
Mitochondrial autophagy is an early defense and protection process;
promoting mitochondrial autophagy causes dysfunctional or damaged
mitochondria to recruit mitochondrial autophagy proteins to the
mitochondrial membrane (12).
PTEN-induced kinase 1 (PINK1)/Parkin is a classical mitochondrial
autophagy pathway. Activation of the PINK1/Parkin pathway can
promote autophagy vesicle formation, degrade damaged mitochondria,
maintain intracellular homeostasis, improve mitochondrial function
and protect neurons from CIRI (13). Recent studies have shown that when
the PINK1/Parkin pathway is activated, it promotes mitochondrial
autophagy (14,15). However, knockout of the PINK1 or
Parkin gene inhibited the activation of the PINK1/Parkin pathway,
hindered mitochondrial clearance and weakened the neuroprotective
effect of drugs. Notably, the histone deacetylase sirtuin 1 (SIRT1)
is an essential regulator of mitochondrial autophagy; it performs a
vital role in the neuroprotection of cerebral ischemia (16). Previous studies have found that
activating SIRT1 expression can significantly improve hippocampal
neuronal damage, reduce hippocampal neuronal apoptosis and improve
cognitive impairment in MCAO/R mice (17,18).
However, the relationship between SIRT1 and mitochondrial autophagy
during cerebral I/R is unclear.
Gastrodia elata is an herbal medicine
commonly used in Asian countries to treat depression, epilepsy and
other neurological diseases (19).
Our previous study demonstrated that a component of G.
elata, p-hydroxybenzaldehyde, can inhibit apoptosis and improve
brain injury in MCAO/R rats (20).
In addition, another component, p-hydroxybenzyl alcohol (pHBA), was
found to have pharmacological effects such as anti-cerebral
ischemic injury, anti-platelet aggregation and anti-inflammation
(21). Previous studies found that
pHBA can reduce the permeability of the blood-brain barrier during
cerebral ischemia, inhibit the expression of aquaporins, reduce
brain edema induced by cerebral ischemia and reduce the
inflammatory response of acute cerebral ischemic injury in rats
(22,23). Other studies have found that pHBA
can improve cerebral ischemic injury by reducing apoptosis
(24), inhibiting oxidative stress
and excitotoxicity, and reducing neuronal death in the hippocampal
CA1 region (25). Therefore, pHBA
has a potential therapeutic effect on cerebral ischemia, but its
protective effect on CIRI through SIRT1-mediated mitophagy has yet
to be elucidated. In the present study, SIRT1 inhibitors were used
to investigate whether pHBA-mediated mitochondrial autophagy
inhibits neuronal apoptosis through SIRT1 to protect neurons in the
ischemic penumbra of MCAO/R rats.
Materials and methods
Animals
Male Sprague-Dawley rats (n=48; age, 5–8 weeks;
weight, 250–280 g) were purchased from the Hunan Slack Jingda
Experimental Animal Co., Ltd., China. All rats were raised in a
specific pathogen-free room, with a temperature of 23±2°C, relative
humidity of 40–60%, a 12-h light-dark cycle and free access to food
and water. The Animal Ethics Committee of Yunnan University of
Chinese Medicine (Kunming, China) approved the animal experiment
(approval no. R-062021088), and the experiments followed the
guidelines of the National Institutes of Health Care and Use of
Laboratory Animals (26). Before
the investigation, the animals were randomly divided into sham
operation group (Sham), MCAO/R model group (MCAO/R), pHBA group
(MCAO/R + pHBA) and EX527 group (MCAO/R + pHBA + EX527), with 12
rats in each group.
Drug administration
According to our previous studies (22,23),
the effective dose of pHBA (cat. no. AF21030253; Chengdu Alfa
Biotechnology Co., Ltd.) was determined to be 20 mg/kg. In the pHBA
and EX527 groups, pHBA was administered by gavage once a day, for 7
days. On the 7th day, the MCAO/R model was prepared after
intragastric administration of pHBA for 30 min. In the EX527 group,
during the intragastric administration of pHBA, 5 mg/kg of the
SIRT1 inhibitor EX527 (cat. no. 110221220509; Beyotime Biotech,
Inc.) was injected into the subarachnoid space, on four occasions,
once every other day. The control and MCAO groups were given equal
volumes of saline.
MCAO/R model and neurological
deficiency score
The rat MCAO/R model was established by operation
according to previous methods (27,28).
Briefly, rats were anesthetized with 5% isoflurane inhalation and
maintained with 3% isoflurane (29). The right common carotid artery,
external carotid artery (ECA) and internal carotid artery (ICA)
were exposed by an incision along the midline of the neck. A
0.36±0.02 mm diameter, rounded poly nylon monofilament (cat. no.
2636-50A4; Beijing Cinontech Co., Ltd.) was introduced into the ECA
and slowly inserted into the ICA to block the start of the middle
cerebral artery. After 2 h of occlusion, the nylon thread was
gently removed to restore blood flow. The body temperature of all
rats was maintained at 37°C throughout the operation. The rats were
evaluated by nerve score after 24 h of reperfusion. Only the
arteries were separated in the Sham operation group, and no thread
emboli were introduced. The neurological deficit score was given a
0–4 rating (Table I) (27).
 | Table I.Neurological deficit score in
rats. |
Table I.
Neurological deficit score in
rats.
| Score | Animal behavioral
characteristics |
|---|
| 0 | Regular activity;
no neurological impairment. |
| 1 | When lifting the
tail, the left forelimb was adducted and could not be completely
extended. |
| 2 | The body rotated to
the left when crawling. |
| 3 | The body tilted to
the left when crawling. |
| 4 | The levels of
consciousness decreased; unable to walk on their own. |
Measurement of cerebral infarct
area
The rats were anesthetized by inhalation of 5%
isoflurane and sacrificed by cervical dislocation. The whole brain
(n=3) was removed and frozen at −20°C for 20 min. Each brain was
cut into 5 slices, each ~2 mm thick, and stained with 2%
2,3,5-triphenyltetrazolium hydrochloride (TTC; cat. no. A610558;
Sangon Biotech) at 37°C for 20 min, before being fixed with 4%
paraformaldehyde at room temperature 24 h. Normal, healthy tissue
was stained red and ischemic tissue was stained white. The
experimental results were analyzed by ImageJ v1.52a (National
Institutes of Health).
Hematoxylin and eosin (H&E)
The ischemic hemisphere tissues were prepared for
H&E staining, and an H&E staining kit was used for
histomorphological analysis (cat. no. KGA224; Key GEN Biotech). The
brain tissue was fixed overnight at room temperature in 4%
paraformaldehyde, embedded in paraffin, and sectioned (5 µm).
Briefly, the prepared paraffin-embedded sections of brain tissue
(n=3) were baked at 60°C for 3 h before being stained with
hematoxylin for 2 min. Tissues were differentiated with 1%
hydrochloric acid alcohol for several seconds, rinsed for 5 min
with water and stained with eosin dye for 1 min before the residual
dye solution was washed off. The sections were dehydrated and dried
with an ascending ethanol gradient, then cleared with xylene to
make and sealed with neutral glue. The pathological changes of
neurons in the ischemic hemisphere were observed using an IXplore
inverted phase contrast microscope (Olympus Corporation; OM Digital
Solutions Corporation) at ×400 magnification, and the images were
analyzed by ImageJ.
Nissl staining
Brain paraffin sections (n=3) were routinely dewaxed
at room temperature with xylene I and xylene II, each 10 min, and
then gradient alcohol dehydration (100, 100, 95, 90, 80, 70 and
50%, each 5 min) and incubated in Nissl staining solution (cat. no.
G1430; Beijing Solarbio Science & Technology Co., Ltd.) for 40
min at 50–60°C. Sections were washed with deionized water,
differentiated with Nissl differentiation solution, dehydrated with
anhydrous ethanol, cleared with xylene and sealed with neutral gum.
The pathological changes of neurons in the ischemic hemisphere were
observed at ×400 magnification using an IX83 inverted phase
contrast microscope (Olympus Corporation; OM Digital Solutions
Corporation). Images were analyzed by ImageJ.
Transmission electron microscopy
(TEM)
At 24 h after reperfusion, the rats were euthanized
(n=3) and the ischemic brain tissue was isolated for TEM to observe
the changes in mitochondrial ultrastructure. The fresh tissues were
cut into small pieces of 1 mm3 and soaked in precooled
glutaraldehyde at 4°C for 4 h. Sections were then dehydrated with a
gradient of ethanol and acetone, as follows: 50% alcohol for 10
min; 70% alcohol for 10 min; 80% acetone for 10 min, twice; 90%
acetone for 10 min, twice; anhydrous acetone for 10 min, twice.
After dehydration, the slices were embedded in epoxy resin and
polymerized in an incubator at 60°C for 48 h. Ultra-thin sections
of 80 nm were prepared and stained with uranium acetate and lead
citrate (cat. nos. GZ02625 and GA1070, respectively; Beijing
Zhongjing Keyi Technology Co., Ltd.). Tissues were stained at room
temperature for 15 min. The images were observed and photographed
using a JEM-1400flash transmission electron microscope (JEOL,
Ltd.).
Detection of ATP levels, mitochondrial
permeability transition pores (mPTP) and mitochondrial membrane
potential (MMP)
The levels of ATP, mPTP and MMP were detected by
first grinding frozen brain tissue from the ischemic hemisphere
(n=6); 100 mg of tissue was used for each test. ATP levels in brain
tissue were detected using an ATP Detection kit (cat. no. A095-1-1;
Nanjing Jiancheng Biological Engineering Research Institute). A
Mitochondrial Extraction kit (cat. no. SM0020; Beijing Solarbio
Science & Technology Co., Ltd.) was used to isolate
mitochondria from brain tissue. BCA Protein Assay kits (cat. no.
PC0020; Beijing Solarbio Science & Technology Co., Ltd.) were
used to detect mitochondrial concentration. The mitochondrial
swelling method was used to determine the degree of opening of mPTP
(cat. no. GMS10101; Shanghai Genmed Gene Medicine Technology Co.,
Ltd.). Briefly, after the mitochondria were extracted, the
concentration of brain mitochondrial protein was adjusted to 10
mg/ml by the BCA method, and 20 µl mitochondrial samples were added
to a 96-well plate. A total of 170 µl GERMED buffer (Reagent A) was
added to each well, and the initial reading of 0 min was obtained.
After that, 10 µl GERMED expansion solution (Reagent B) was added
immediately at room temperature, and the changes of 10 min
absorbance were dynamically recorded by the VariosKan Flash
Multi-detection Microplate Reader (Thermo Fisher Scientific, Inc.).
The actual absorption readings were calculated as, optical density
(OD)0 min-OD10 min; if the actual absorption
reading is high, this indicates high mitochondrial expansion. The
VariosKan Flash Multi-detection Microplate Reader was used to
detect the level of MMP (cat. no. GMS10013.1; Shanghai Genmed Gene
Medicine Technology Co., Ltd.), according to the manufacturer's
protocol.
Immunofluorescence
Ischemic hemispheres of rats were selected for
immunofluorescence detection (n=3). The paraffin-embedded sections
(10 µm each) were blocked for 10 min at 37° containing 5% goat
serum (cat. no. C0265; Beyotime Institute of Biotechnology).
Primary antibodies anti-LC3 (1:200; cat. no. 4108; Cell Signaling
Technology, Inc.) and p62 (1:250; cat. no. 18420-1-AP; Proteintech
Group, Inc.) were incubated overnight at 4°C. Alexa Fluor
488-conjugated goat anti-rabbit IgG (1:1,000; cat. no. ab150077;
Abcam) and rabbit anti-mouse (1:1,000; cat. no. ab150125; Abcam)
secondary antibodies were incubated at 37°C for 1 h. The nuclei
were stained with DAPI for 5 min at room temperature in the dark,
and then the slides were sealed with 90% glycerol. The sections
were observed and photographed under ×400 magnification with a
Zeiss LSM laser-scanning confocal microscope (Carl Zeiss AG).
Western blotting
Frozen brain tissues (n=3) were lysed using ice-cold
RIPA buffer (PSMF:RIPA lysate, 1:100; cat. no. 051021210825;
Beyotime Institute of Biotechnology) for 20 min. The lysate was
centrifuged at 12,000 g for 5 min at 4°C. Total protein was
quantified using the BCA method. Proteins (80 µg) were separated by
SDS-PAGE on an 8% gel. The separated proteins were subsequently
transferred onto a PVDF membrane (Bio-Rad Laboratories, Ltd.) and
was blocked in 5% skim milk at room temperature for 2 h. The
membrane was incubated overnight with primary antibodies against
SIRT1 (1:1,000; cat. no. ab110304; Abcam), PINK1 (1:500; cat. no.
sc-517353; Santa Cruz Biotechnology), Parkin (1:1,000; cat. no.
4211; Cell Signaling Technology), LC3 A/B (1:1,000; cat. no. 4108;
Cell Signaling Technology), p62 (polyclonal; 1:500; cat. no.
18420-1-AP; Proteintech) and β-Actin (1:1,000; cat. no. ab8226;
Abcam) at 4°C. HRP-conjugated goat anti-rabbit IgG H&L
(1:10,000; cat. no. ab6721) and rabbit anti-mouse IgG H&L
(1:10,000; cat. no. ab6728; Abcam) secondary antibodies were added
and the membranes a were incubated for 1 h at room temperature. The
protein bands were visualized by ECL using SuperSignal™ West Atto
Ultimate Sensitivity Substrate (cat. no. A38555; Thermo Fisher
Scientific, Inc.), and images of the bands were captured using a
Tanon 6600 Luminescent Imaging Workstation (Tanon Science and
Technology Co., Ltd.) and analyzed using ImageJ software.
TUNEL assay
The ischemic hemispheres of rats (n=3) were stained
with TUNEL Detection Kit (n=3; cat. no. C1088; Beyotime Institute
of Biotechnology). The paraffin sections were routinely dewaxed, as
follows: After baking at 60° for 60 min, the sections were
incubated twice with xylene at room temperature for 10 min each
time, and rehydrated in a descending ethanol series (100, 95, 80
and 75%; 5 min each). Subsequently, 20 µg/ml protease K (without
DNase) was added, and the tissues were incubated at 37°C for 15–30
min. Sections were washed three times with PBS before 50 µl of
TUNEL detection solution was added to each sample and incubated at
37°C for 1 h. The sections were then washed three times with PBS,
and the nuclei were counterstained with DAPI for 5 min at room
temperature. The sections were observed and images captured using a
Zeiss LSM laser confocal microscope (Carl Zeiss AG) at ×400
magnification. ImageJ software was used to calculated the
fluorescence intensity of positive cells in three non-overlapping
visual fields.
Statistical analysis
GraphPad Prism software (version 9.0.0; Dotmatics)
was used for statistical analysis. For normal distribution, the
Kolmogorov-Smirnov test was used. If the data were normally
distributed, and if the variances were homogeneous, they were
compared using ANOVA followed by Bonferroni's multiple comparison
post hoc test. If the variances were unequal, Welch's ANOVA test
was used followed by Dunnett's T3 multiple comparison tests. All
values are presented as mean ± SEM. P<0.05 was used to indicate
a statistically significant difference.
Results
pHBA improves cerebral infarction
area, neurological function and pathological injury after MCAO/R
injury
An overview of the design of the present study is
shown in Fig. 1A. The chemical
structure of pHBA is a phenolic compound consisting of a benzene
ring and two hydroxyl groups (Fig.
1B). Our previous study examined a pHBA derivative,
p-hydroxybenzaldehyde, which comprised a benzene ring and an
aldehyde group with a hydroxyl group (20). Following drug administration, MCAO
modelling and reperfusion, the neurological deficit score of rats
was evaluated. It was found that the neurological deficit score of
the MCAO/R group was significantly higher compared with that of the
Sham group (Fig. 1C); neurological
deficit scores were decreased after pHBA treatment compared with
untreated MCAO/R. The rats were euthanized and the cerebral
infarction area was measured. The TTC staining results showed that
the area of cerebral infarction in the MCAO/R group was
significantly larger compared with the Sham group (Fig. 1D and E). Compared with MCAO/R
group, the cerebral infarction area in the pHBA group was
significantly smaller. In addition, H&E and Nissl staining
revealed that the neurons in the Sham operation group showed clear
nuclei and Nissl bodies (Fig. 1F and
G, respectively). H&E staining demonstrated that the nerve
cells in the Sham group were arranged neatly and the morphological
structure was normal, whereas the fiber arrangement in the MCAO/R
model group was disordered, and the nuclei were pyknotic. The
morphology and structure of nerve cells in the pHBA-treated group
were more neatly arranged compared with those in the model group,
and the cell bodies were transparent. In the EX527 co-treatment
group, fibers were displaced, cell distribution was disordered and
the cell body was blurred. Nissl staining showed that the structure
of Nissl corpuscles was clear and arranged neatly in the Sham
operation group. Cell damage in the MCAO/R group was characterized
by nuclear pyknosis and disordered arrangement of Nissl bodies.
Compared with the MCAO/R group, the injury of nerve cells in the
pHBA group was alleviated, and the shape and size of Nissl bodies
were uniform. Compared with the pHBA group, the arrangement of
Nissl bodies in the EX527 group was disordered, and some of the
nuclear pyknosis was deeply stained. These data suggested that pHBA
may improve infarct size, neurological function and neuronal damage
after CIRI, potentially through the SIRT1 pathway.
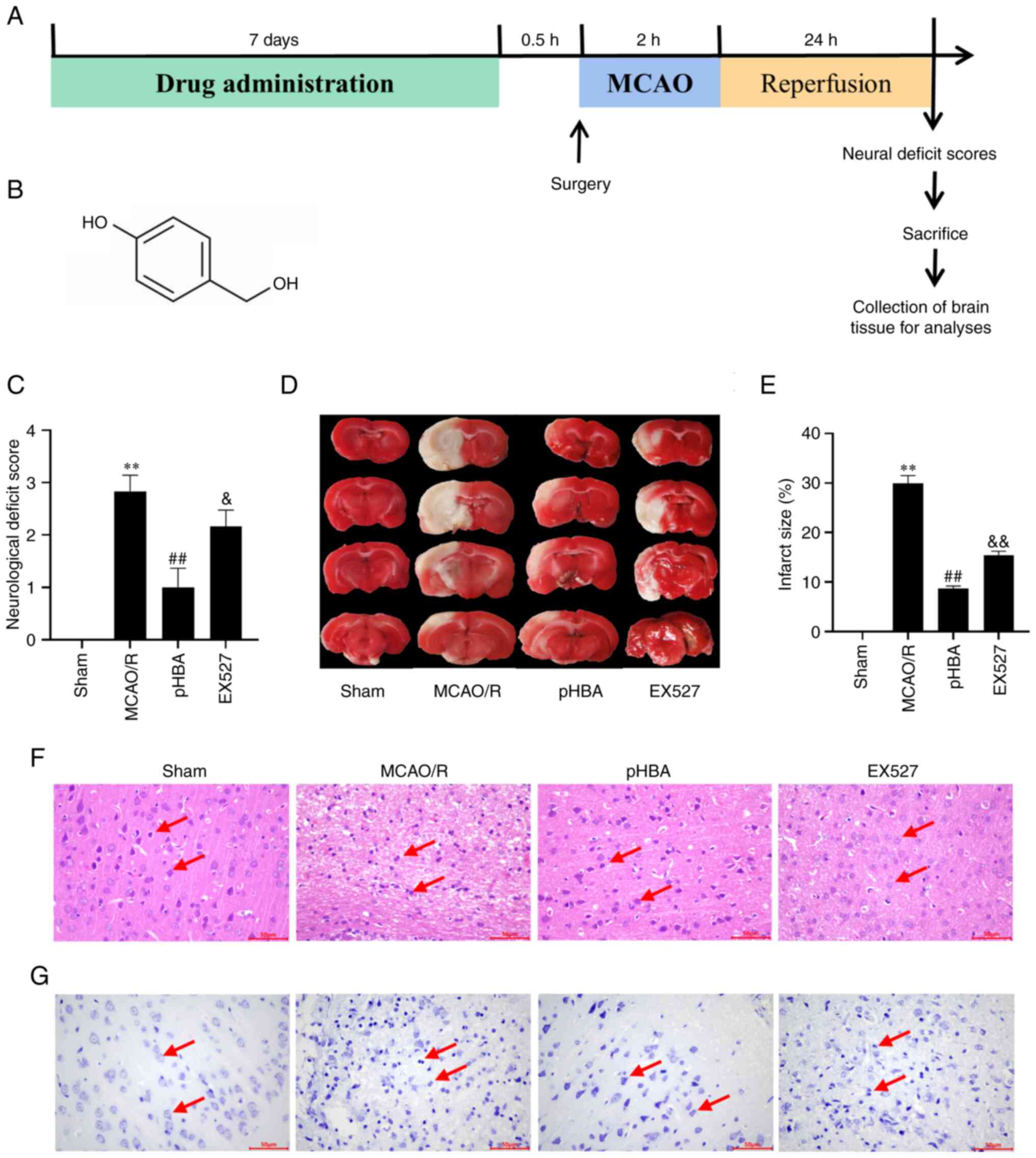 | Figure 1.pHBA treatment reduces infarct size,
neurological deficit and neuronal damage after cerebral I/R. (A)
Timeline schematic of MCAO/R surgery-induced cerebral I/R. Here,
0.5 h indicates that the model will be established following 0.5 h
of pHBA intragastric administration on the 7th day. (B) Chemical
structure of pHBA. (C) Neurological deficit score of each group
(n=12). At 24 h after reperfusion, (D) brain sections from rats in
each group (n=3) were stained with TTC, and (E) quantitative
analysis was conducted to determine the infarct size. (F)
Representative H&E stained tissue sections from each group
(n=6); arrows indicate nerve cells. Scale bar, 50 µm. (G)
Representative Nissl stained tissue sections from each group (n=6);
the arrows indicate Nissl bodies. Scale bar, 50 µm. All data are
presented as the mean ± SEM. **P<0.01 vs. Sham;
##P<0.01 vs. MCAO/R; &P<0.05,
&&P<0.01 vs. pHBA. EX527, sirtuin inhibitor;
I/R, ischemia-reperfusion; MCAO/R, middle cerebral artery occlusion
and reperfusion; pHBA, p-hydroxybenzyl alcohol; TTC,
2,3,5-triphenyltetrazolium hydrochloride. |
pHBA ameliorates mitochondrial
microstructure damage and dysfunction after MCAO/R injury
To determine whether pHBA can improve the
mitochondrial dysfunction caused by MCAO/R injury, the
microstructure of mitochondria in the ischemic penumbra was
observed by TEM. The results showed that the mitochondria swelled
and the cristae disappeared in the MCAO/R group compared with the
Sham group (Fig. 2A). Compared
with the MCAO/R group, pHBA could reduce mitochondrial swelling and
cristae disappearance (Fig. 2A).
Compared with the pHBA group, EX527 aggravated mitochondrial
swelling and cristae disappearance (Fig. 2A). In addition, the brain tissues
of ischemic hemispheres were isolated, and the levels of
mitochondrial ATP, mPTP and MMP were analyzed. The data showed
that, compared with the Sham group (ATP, 40.64±2.28 µm/g protein;
mPTP, 0.016±0.001 OD; MMP, 5.58±0.36 relative fluorescence)
(Fig. 2B-D, respectively), the
level of ATP (14.84±1.29 µm/g protein) and MMP (1.74±0.18 relative
fluorescence) in the MCAO/R group decreased significantly, whereas
the openness of mPTP (0.033±0.002 OD) increased. Treatment with
pHBA reversed the effect of MCAO/R on ATP (29.67±1.14 µm/g
protein), mPTP (0.018±0.001 OD) and MMP (4.08±0.40 relative
fluorescence). However, these improvements were inhibited by EX527
co-treatment. These results suggested that part of the protective
effect of pHBA on MCAO/R damage may be to improve mitochondrial
function through the SIRT1 pathway.
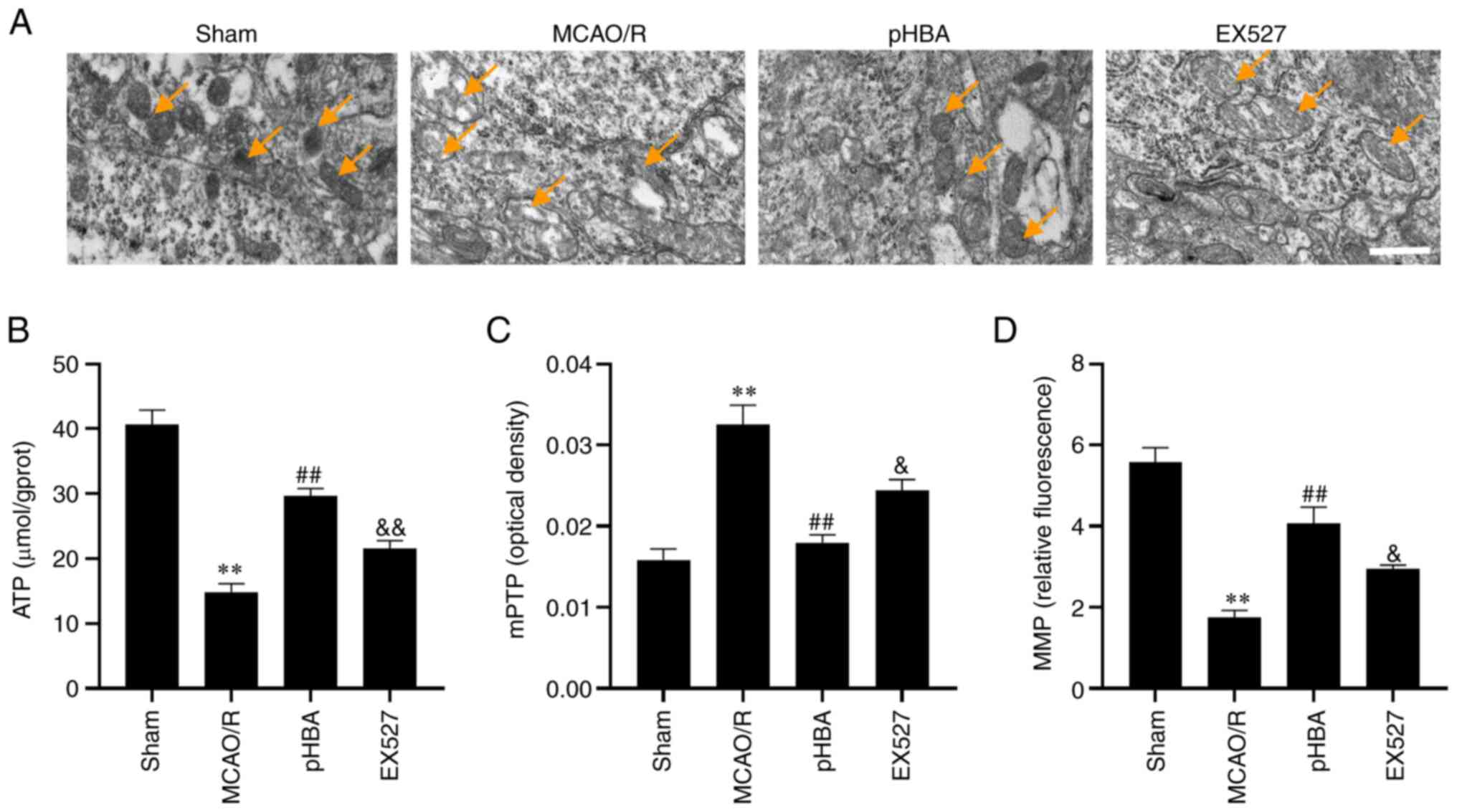 | Figure 2.pHBA improves microstructure damage
and dysfunction of mitochondria following cerebral
ischemia-reperfusion. (A) Representative transmission electron
microscopy images of tissue sections from rats each group (n=3)
showing mitochondria (arrows) in the ischemic penumbra affected
tissues. In the MCAO/R group, the shape of the mitochondria
(arrows) not clear, swelling occurred and the cristae disappeared.
In the pHBA group, the morphology of the mitochondria indicated by
the arrow was clear, and the structure was complete. In the EX527
group, the mitochondria indicated by the arrow were swollen, and
the cristae were broken. Scale bar, 500 nm. (B) ATP level, (C)
degree of opening of mPTP and (D) MMP in rats from each group
(n=6). All data are presented as the mean ± SEM. **P<0.01 vs.
Sham; ##P<0.01 vs. MCAO/R; &P<0.05,
&&P<0.01 vs. pHBA. EX527, sirtuin inhibitor;
MCAO/R, middle cerebral artery occlusion and reperfusion; MMP,
mitochondrial membrane potential; mPTP, mitochondrial permeability
transition pore; pHBA, p-hydroxybenzyl alcohol. |
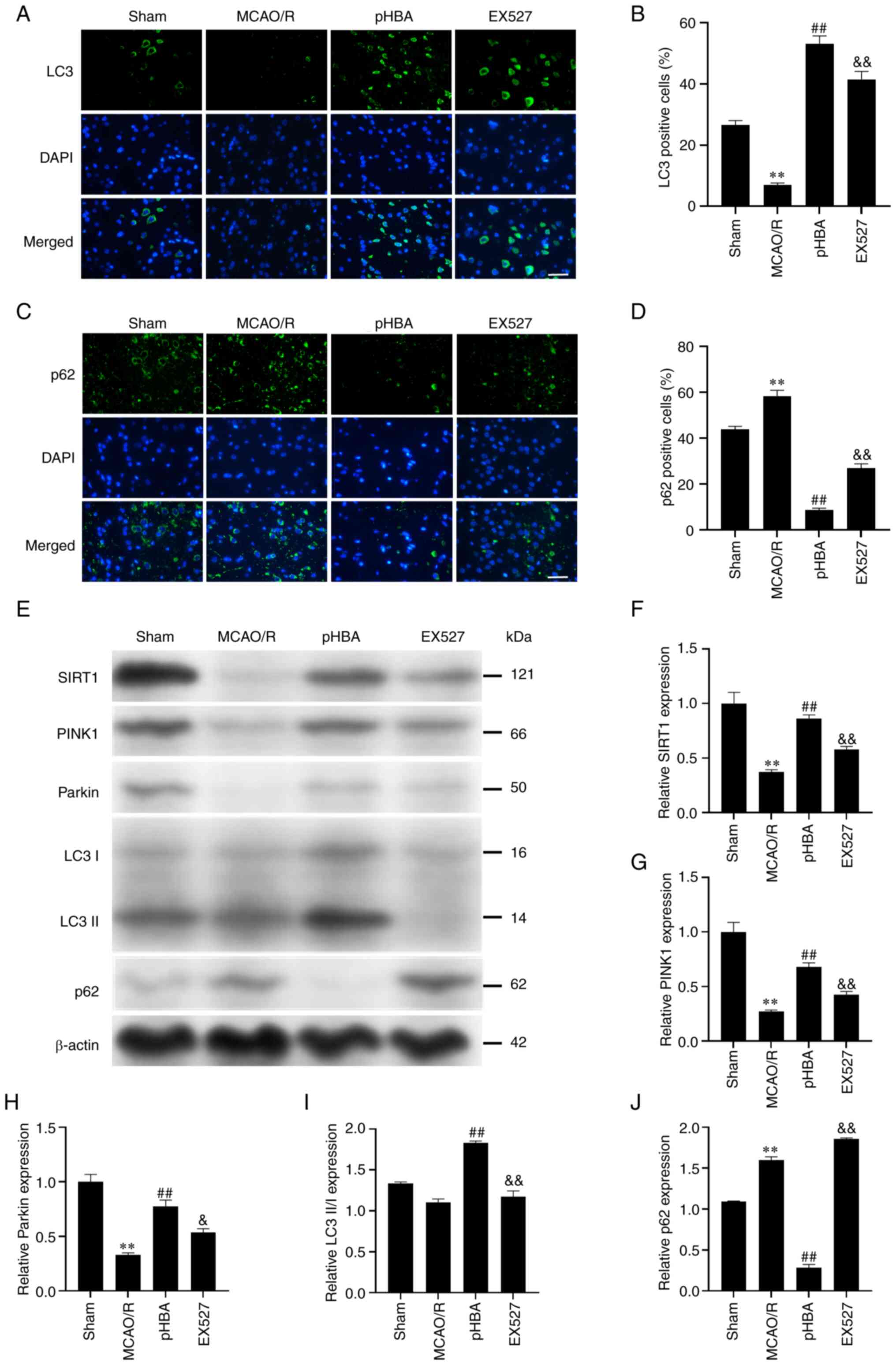
pHBA activates mitochondrial autophagy
in neurons through SIRT1
To verify whether pHBA activates mitochondrial
autophagy through the SIRT1 pathway, immunofluorescence staining
was performed. The data showed that the percentage of LC3-positive
cells in the MCAO/R group was significantly lower (6.93±0.70%)
compared with that in the Sham group (26.50±1.52%) (Fig. 3A-D), whereas the rate of
p62-positive cells was significantly higher in the MCAO/R group
(43.81±1.32 vs. 58.37±2.44%). Compared with the MCAO/R group, pHBA
significantly increased the percentage of LC3-positive cells
(53.16±2.50%) and decreased p62-positive cells (8.65±0.76%);
however, inhibiting the expression of SIRT1 reversed these effects.
Western blotting results showed that pHBA significantly increased
the relative protein expression levels of SIRT1, PINK1, Parkin and
LC3 II/I, and significantly decreased the relative expression of
p62 (Fig. 3E-J); these effects
were inhibited after treatment with the EX527 SIRT1 inhibitor.
These results suggested that the regulation of mitochondrial
autophagy by pHBA may be related to SIRT1.
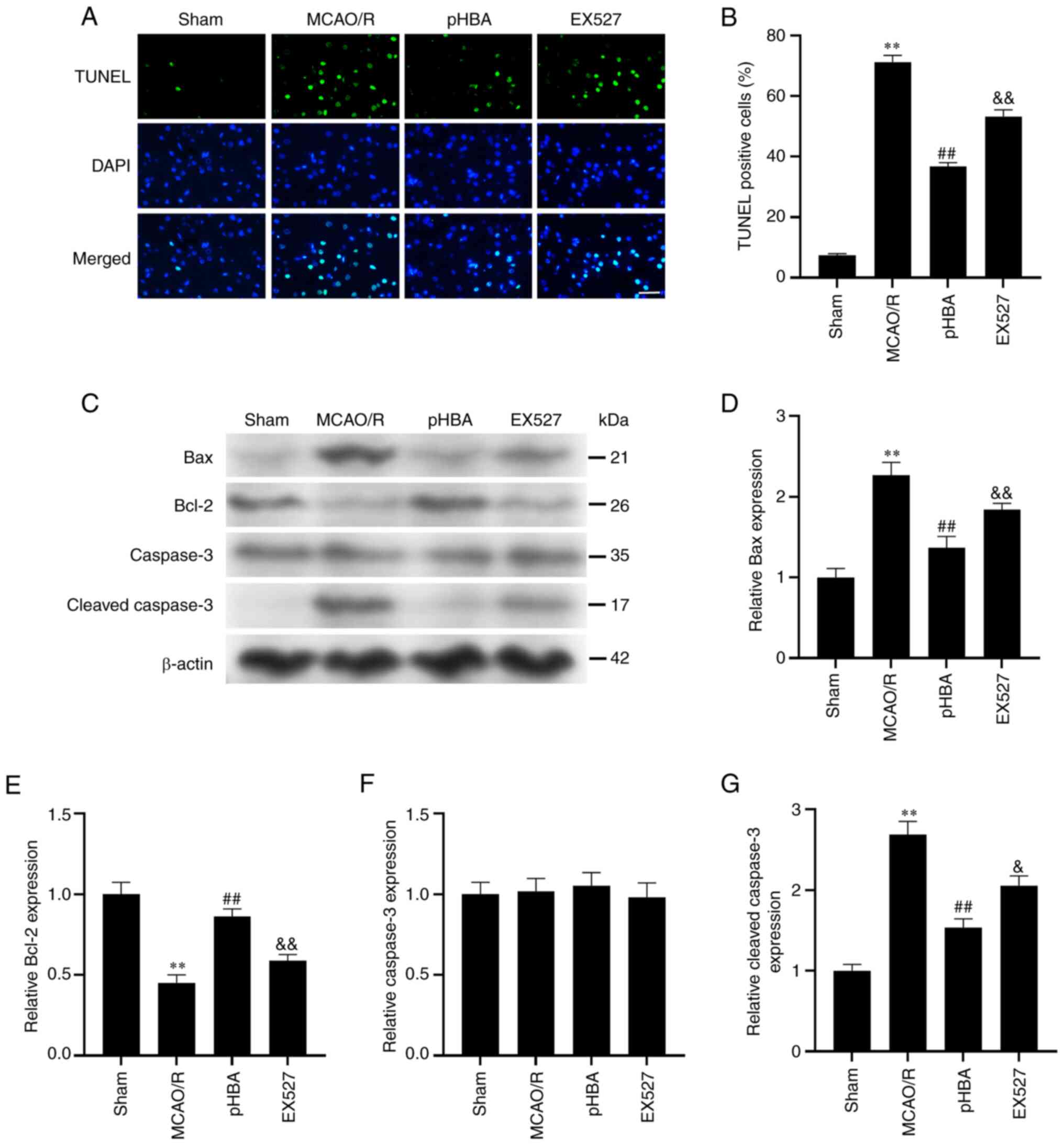
pHBA inhibits neuronal apoptosis
induced by MCAO/R injury by activating SIRT1
The effect of SIRT1 on neuronal apoptosis was
examined by TUNEL. The results showed that the number of
TUNEL-positive cells increased significantly in the MCAO/R group,
and this was decreased significantly after administration of pHBA
(Fig. 4A and B); however the
ameliorative effect of pHBA was reversed after co-treatment with
EX527. In addition, western blotting results showed that the
protein expression of Bax and cleaved caspase-3 protein in the
MCAO/R group was higher compared with that in the Sham group,
whereas the expression level of Bcl-2 was lower in the MCAO/R group
(Fig. 4C-G). Compared with the
MCAO/R group, pHBA treatment could inhibit the expression of Bax
and cleaved caspase-3 protein in MCAO/R injury and significantly
increase the expression level of Bcl-2. Notably, these effects were
blocked by the SIRT1 inhibitor. These results suggested that pHBA
may inhibit apoptosis through SIRT1.
Discussion
Our previous study showed that
p-hydroxybenzaldehyde, a component of G. elata, can protect
MCAO/R rats from oxidation and apoptosis (20). Compared with the MCAO/R group,
p-hydroxybenzaldehyde treatment reduced mitochondrial ROS and
malondialdehyde contents, reduced mPTP opening, and decreased Bax
and caspase-3 protein expression levels. In addition, ATP content,
cytochrome c oxidase and total superoxide dismutase
activities, as well as protein expression of Bcl2 all increased
(20). Similarly, results from the
present study demonstrated that pHBA treatment increased ATP level,
mPTP opening and Bcl2 expression, whereas mPTP opening and Bax and
cleaved caspase-3 expression levels were decreased compared with
the untreated MCAO/R model group. In addition, this study found
that pHBA may protect CIRI in MCAO/R model rats by autophagy and
apoptosis induced by mitochondrial pathway. The results suggested
that pHBA treatment may significantly reduce the neurological
deficit score and cerebral infarction and significantly improve
cell damage, such as nuclear condensation and reduction of Nissl
bodies caused by MCAO/R injury. It was also discovered that pHBA
may activate mitochondrial autophagy following brain injury. In
addition, pHBA treatment promoted the expression of SIRT1 after
MCAO/R, which was inhibited after EX527 treatment. Notably, pHBA
increased the expression of the anti-apoptotic protein Bcl-2 and
decreased the expression of the pro-apoptotic protein Bax in MCAO/R
model rats, suggesting that pHBA may inhibit neuronal apoptosis.
Therefore, these results indicate that pHBA may have a
neuroprotective effect on MCAO/R rats by activating the
SIRT1/mitochondrial autophagy pathway and inhibiting neuronal
apoptosis.
Mitochondrial injury is a marker of ischemic stroke.
During cerebral I/R, the structure and function of mitochondria are
damaged, which can aggravate pathological processes, such as
oxidative stress, calcium overload and inflammatory (27). However, these pathological
processes also aggravate mitochondrial damage, forming a negative
chain of events (14).
Mitochondrial damage is mainly characterized by a decrease of MMP
and an impairment of the ability to produce ATP. At the same time,
these pathological factors (ATP and MMP) will also open mPTP and
release apoptosis factors and accelerate nerve cell death (30). Huang et al (31) found that brain microvascular
endothelial cells injury induced by hypoxia-glucose deprivation and
reoxygenation (OGD/R) led to the opening of mPTP. Nonetheless,
hydroxysafflor yellow A treatment not only inhibited the opening of
mPTP, but also reduced the release of apoptotic factors. Wang et
al (32) found that influenza
A virus PB1-F2 protein treatment could increase levels of MMP and
ATP, and prevent damage to mitochondrial function caused by CIRI.
Consistent with these data, the present study found that pHBA
treatment improved the ultrastructure and morphology of
mitochondria in the ischemic penumbra of MCAO/R rats. It also
reduced the opening of mitochondrial mPTP in ischemic hemispheres
and increased the levels of MMP and ATP, reversing damage to
mitochondrial function. It is suggested that pHBA may improve
mitochondrial dysfunction after CIRI.
Mitochondrial autophagy is the primary way to
identify and remove damaged mitochondria, to maintain the
mitochondrial network's stability (4), and to recycle decomposed cellular
components to maintain cellular homeostasis (14). When mitochondrial autophagy occurs,
PINK1 in the cytoplasm accumulates on the outer membrane of damaged
mitochondria and recruits Parkin to initiate mitochondrial
autophagy (32,33). Subsequently, it promotes the
binding of p62 and LC3 to activate autophagosomes, resulting in the
degradation of damaged mitochondria by lysosomes, which supports
the use of p62 and LC3 proteins as indicators to verify autophagy
activation. In the present study, it was found that p62 decreased
and LC3 increased after pHBA treatment. Similarly, western blotting
showed that pHBA treatment promoted the expression of mitochondrial
autophagy-related proteins PINK1, Parkin and LC3 II/I, and
decreased the expression of p62. It is suggested that pHBA may
activate mitochondrial autophagy after cerebral I/R.
Neuronal apoptosis is the main cause of brain injury
after intracerebral hemorrhage, which can lead to brain infarction
and loss of nerve function (34).
Mitochondrial-mediated apoptosis is one of the main pathways of
apoptosis. When cells are injured, the expression levels of
anti-apoptotic protein Bcl-2 decreases, and the expression of the
pro-apoptotic protein Bax increases; Bax accumulates on the
mitochondrial membrane and reduces MMP (35). Additionally, Bax opens mPTP,
releases apoptotic signals and activates caspase-3 to trigger
apoptosis (36). It is suggested
that the function of mitochondria can affect apoptosis, and it has
been reported that improving mitochondrial function can reduce
apoptosis (7,37). In the present study, mitochondrial
autophagy was shown to maintain a healthy mitochondrial network,
which can enhance the level of mitochondrial MMP and ATP, and
reduce the opening of mPTP, indicating that mitochondrial autophagy
can affect apoptosis. Several studies have found that promoting
mitochondrial autophagy can reduce mitochondrial-mediated neuronal
apoptosis (13,38,39).
Similarly, in the present study, pHBA treatment significantly
reduced the number of TUNEL-positive cells and the protein
expression of Bax and cleaved caspase-3, and upregulated the
expression of Bcl2. These findings suggested that the protective
effect of pHBA on CIRI is related to the inhibition of
mitochondrial-mediated apoptosis.
SIRT1 is a nicotinamide adenine
dinucleotide-dependent deacetylase involved in various biological
processes, including metabolism, apoptosis, aging, oxidative
stress, energy metabolism and inflammation (40,41).
Studies have shown that SIRT1 is involved in the regulation of
energy metabolism, autophagy, apoptosis and oxidation following
ischemic stroke (42–44). In addition, SIRT1 also serves a
crucial role in eliminating damaged mitochondria through
mitochondrial autophagy. A number of studies have found that the
PINK1/Parkin signaling pathway can be activated by SIRT1 (45). For example, Huang et al
(46) found that SIRT1 can induce
mitochondrial autophagy by activating the PINK1/Parkin signaling
pathway, reducing brain CIRI damage and OGD/R human neuroblastoma
cell damage. In addition, Shao et al (47) found that SIRT1-mediated
PINK1/Parkin-dependent mitochondrial autophagy participates in the
neuroprotective effect of mouse hippocampal HT22 OGD/R model cells
on mitochondrial dysfunction. In the present study, pHBA treatment
upregulated the expression of SIRT1, and SIRT1 may be an upstream
autophagy protein. The results also showed that the use of SIRT1
inhibitor, EX527, repressed the improvement effects of pHBA and
decreased the expression of SIRT1 protein. Inhibition of
mitochondrial autophagy aggravated mitochondrial dysfunction,
reduced the expression of anti-apoptotic factors and increased the
expression of pro-apoptotic factors. In addition, EX527
co-treatment negatively affected neurological function and cerebral
infarction area, and inhibited the improvement of organelles such
as neuronal nuclei and Nissl bodies. Therefore, the present study
concluded that the protective effect of pHBA on CIRI may be related
to the promotion of mitochondrial autophagy and the inhibition of
mitochondrial-mediated apoptosis by the SIRT1 pathway.
In conclusion, the present study found that pHBA has
a neuroprotective effect against CIRI, potentially elicited by
activation of the SIRT1/mitochondrial autophagy pathway and the
inhibition of apoptosis in ischemic penumbra, as illustrated in
Fig. 5. Currently, pHBA can be
artificially synthesized (48),
and using commercial pHBA in the experiment will not harm G.
elata species. These findings suggested that the natural
component pHBA may be a promising option for treating CIRI, and the
SIRT1/mitophagy pathway may be a target for future treatment.
However, there are some limitations in the present study, including
the lack of detection of additional autophagy markers (ATG5, ATG7
and ATG12) and the lack of relevant cell experiments to analyze the
protein or mRNA expression levels of other apoptosis-related
proteins, such as cytochrome c or cleaved caspase-9. In
addition, the absence of observation of autophagosome and
autolysosome formation using electron microscopy is also a
limitation of this study and will form part of our next research
direction.
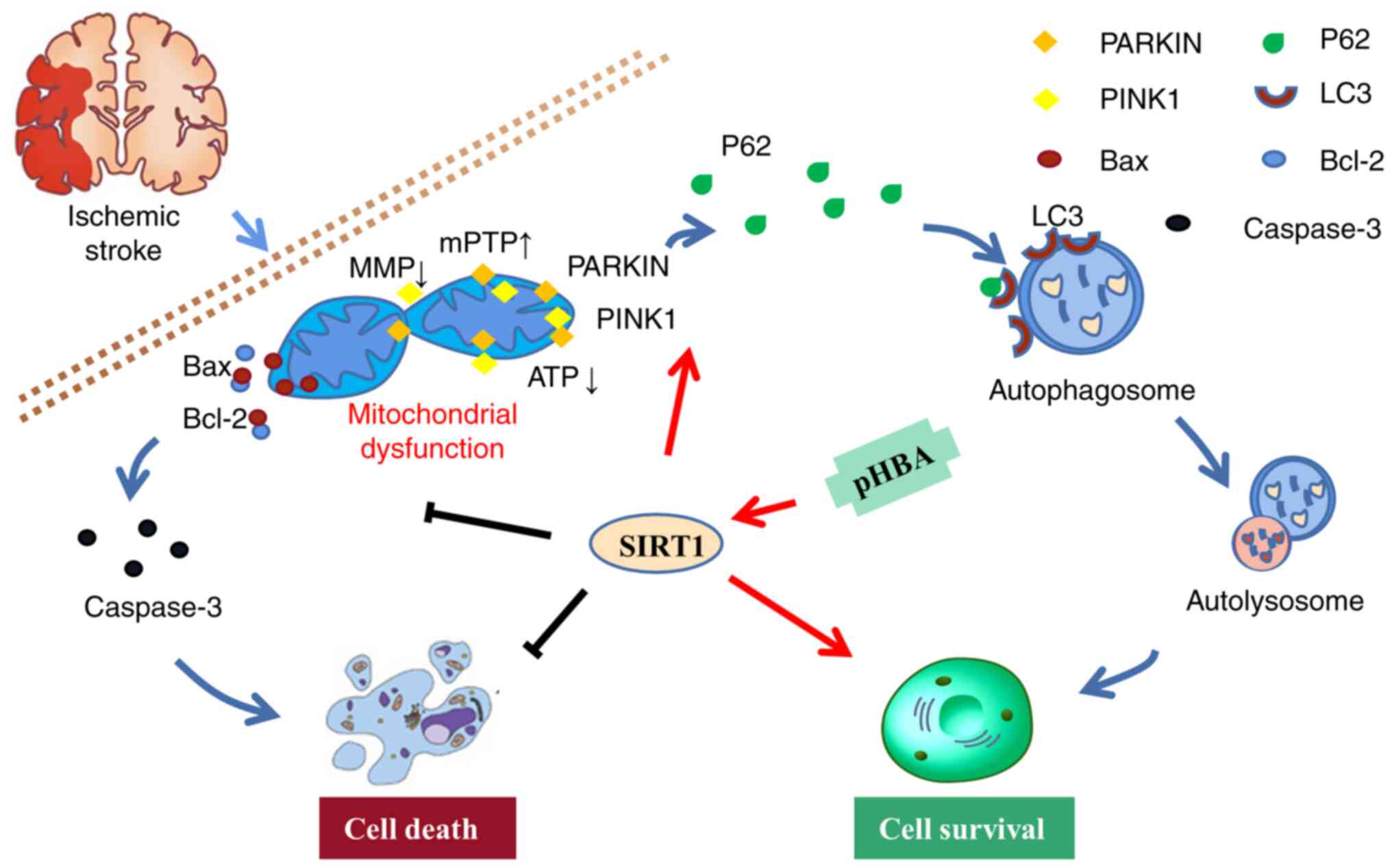 | Figure 5.Illustration of the ameliorating
effects of pHBA on MCAO/R-induced brain injury by activating
mitophagy and inhibiting mitochondria-mediated apoptosis through
the SIRT1 pathway. Mitochondrial damage was induced in rats
following MCAO/R surgery. SIRT1 can promote the mitochondrial
recruitment of PINK1 and Parkin, further promote the binding of p62
and LC3, contribute to the formation of autophagosomes, participate
in mitochondrial autophagy, clear the damaged mitochondria induced
by MCAO/R, and improve Cell survival. In addition, damaged
mitochondria cause the pro-apoptotic protein Bax to move to the
mitochondrial membrane, which triggers the activation of caspase-3
and promotes apoptosis. pHBA may serve a neuroprotective role by
activating SIRT1 to promote mitochondrial autophagy and inhibit
apoptosis. The black line represents inhibition, and the red arrow
represents promotion. MCAO/R, middle cerebral artery occlusion and
reperfusion; MMP, mitochondrial membrane potential; mPTP,
mitochondrial permeability transition pore; pHBA, p-hydroxybenzyl
alcohol; PINK1, PTEN-induced kinase 1; SIRT1, sirtuin 1. |
Acknowledgements
Not applicable.
Funding
This study was supported by The National Natural Science
Foundation of China (grant no. 81960733) and the Xingdian Talent
Support Program - Special for Young Talent (2022).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author upon reasonable
request.
Authors' contributions
XD, PC and XY designed the experiments. YL, LY and
XY performed the experiments. The results were analyzed by XY. LY
wrote the manuscript, and XD edited the manuscript. XD, PC and LY
confirm the authenticity of all the raw data. All the authors
reviewed and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Animal
Ethics Committee of Yunnan University of Chinese Medicine (Kunming,
China; approval no. R-062021088).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Feigin VL, Brainin M, Norrving B, Martins
S, Sacco RL, Hacke W, Fisher M, Pandian J and Lindsay P: World
Stroke Organization (WSO): Global stroke fact sheet 2022. Int J
Stroke. 17:18–29. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Collaborators GBDS, . Global, regional,
and national burden of stroke and its risk factors, 1990–2019: A
systematic analysis for the Global Burden of Disease Study 2019.
Lancet Neurol. 20:795–820. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liu D, Wang H, Zhang Y and Zhang Z:
Protective effects of chlorogenic acid on cerebral
ischemia/reperfusion injury rats by regulating oxidative
stress-related Nrf2 pathway. Drug Des Devel Ther. 14:51–60. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shen L, Gan Q, Yang Y, Reis C, Zhang Z, Xu
S, Zhang T and Sun C: Mitophagy in cerebral ischemia and
ischemia/reperfusion injury. Front Aging Neurosci. 13:6872462021.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Eryildiz ES and Ozdemir AO: Factors
associated with early recovery after intravenous thrombolytic
therapy in acute ischemic stroke. Noro Psikiyatr Ars. 55:80–83.
2018.PubMed/NCBI
|
|
6
|
Yuan Q, Yuan Y, Zheng Y, Sheng R, Liu L,
Xie F and Tan J: Anti-cerebral ischemia reperfusion injury of
polysaccharides: A review of the mechanisms. Biomed Pharmacother.
137:1113032021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Carinci M, Vezzani B, Patergnani S,
Ludewig P, Lessmann K, Magnus T, Casetta I, Pugliatti M, Pinton P
and Giorgi C: Different roles of mitochondria in cell death and
inflammation: Focusing on mitochondrial quality control in ischemic
stroke and reperfusion. Biomedicines. 9:1692021. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lai Y, Lin P, Chen M, Zhang Y, Chen J,
Zheng M, Liu J, Du H, Chen R, Pan X, et al: Restoration of L-OPA1
alleviates acute ischemic stroke injury in rats via inhibiting
neuronal apoptosis and preserving mitochondrial function. Redox
Biol. 34:1015032020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nakamura Y, Lo EH and Hayakawa K:
Placental mitochondria therapy for cerebral ischemia-reperfusion
injury in mice. Stroke. 51:3142–3146. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang SD, Fu YY, Han XY, Yong ZJ, Li Q, Hu
Z and Liu ZG: Hyperbaric oxygen preconditioning protects against
cerebral ischemia/reperfusion injury by inhibiting mitochondrial
apoptosis and energy metabolism disturbance. Neurochem Res.
46:866–877. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lei L, Yang S, Lu X, Zhang Y and Li T:
Research progress on the mechanism of mitochondrial autophagy in
cerebral stroke. Front Aging Neurosci. 13:6986012021. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Anzell AR, Maizy R, Przyklenk K and
Sanderson TH: Mitochondrial quality control and disease: Insights
into ischemia-reperfusion injury. Mol Neurobiol. 55:2547–2564.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mao Z, Tian L, Liu J, Wu Q, Wang N, Wang
G, Wang Y and Seto S: Ligustilide ameliorates hippocampal neuronal
injury after cerebral ischemia reperfusion through activating
PINK1/Parkin-dependent mitophagy. Phytomedicine. 101:1541112022.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wen H, Li L, Zhan L, Zuo Y, Li K, Qiu M,
Li H, Sun W and Xu E: Hypoxic postconditioning promotes mitophagy
against transient global cerebral ischemia via PINK1/Parkin-induced
mitochondrial ubiquitination in adult rats. Cell Death Dis.
12:6302021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wu M, Lu G, Lao YZ, Zhang H, Zheng D,
Zheng ZQ, Yi J, Xiang Q, Wang LM, Tan HS, et al:
Garciesculenxanthone B induces PINK1-Parkin-mediated mitophagy and
prevents ischemia-reperfusion brain injury in mice. Acta Pharmacol
Sin. 42:199–208. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tang Q, Len Q, Liu Z and Wang W:
Overexpression of miR-22 attenuates oxidative stress injury in
diabetic cardiomyopathy via Sirt 1. Cardiovasc Ther. 36:2018.
View Article : Google Scholar
|
|
17
|
Xian W, Li T, Li L, Hu L and Cao J:
Maresin 1 attenuates the inflammatory response and mitochondrial
damage in mice with cerebral ischemia/reperfusion in a
SIRT1-dependent manner. Brain Res. 1711:83–90. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen L, Cao J, Cao D, Wang M, Xiang H,
Yang Y, Ying T and Cong H: Protective effect of dexmedetomidine
against diabetic hyperglycemia-exacerbated cerebral
ischemia/reperfusion injury: An in vivo and in vitro study. Life
Sci. 235:1165532019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ojemann LM, Nelson WL, Shin DS, Rowe AO
and Buchanan RA: Tian ma, an ancient Chinese herb, offers new
options for the treatment of epilepsy and other conditions.
Epilepsy Behav. 8:376–383. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xiao T, Yang L, Chen P and Duan X:
Para-hydroxybenzaldehyde against transient focal cerebral ischemia
in rats via mitochondrial preservation. Exp Ther Med. 24:7162022.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang ZL, Gao YG, Zang P, Gu PP, Zhao Y,
He ZM and Zhu HY: Research progress on mechanism of gastrodin and
p-hydroxybenzyl alcohol on central nervous system. Zhongguo Zhong
Yao Za Zhi. 45:312–320. 2020.(In Chinese). PubMed/NCBI
|
|
22
|
Yan HW, Jia YY, Yang Y, Song XL, He FY and
Lin Q: The protective effect of 4-hydroxybenzyl alcohol on blood
brain barrier in MCAO/R model rats. Pharmacology and Clinics of
Chinese Materia. 32:22–25. 2016.
|
|
23
|
Jiang S, Liu SS, Xiang B, Lin Q and Li XF:
Effect of 4-hydroxybenzyl alcohol on inflammatory cytokines in rats
with acute cerebral ischemic injury. Chinese Traditional Patent
Medicine. 37:2132–2135. 2015.
|
|
24
|
Yu SS, Zhao J, Zheng WP and Zhao Y:
Neuroprotective effect of 4-hydroxybenzyl alcohol against transient
focal cerebral ischemia via anti-apoptosis in rats. Brain Res.
1308:167–175. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim HJ, Hwang IK and Won MH: Vanillin,
4-hydroxybenzyl aldehyde and 4-hydroxybenzyl alcohol prevent
hippocampal CA1 cell death following global ischemia. Brain Res.
1181:130–141. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Guide for the Care and Use of Laboratory
Animals. National Academies Press; Washington, DC: 1996
|
|
27
|
Longa EZ, Weinstein PR, Carlson S and
Cummins R: Reversible middle cerebral artery occlusion without
craniectomy in rats. Stroke. 20:84–91. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li H, Peng D, Zhang SJ, Zhang Y, Wang Q
and Guan L: Buyang Huanwu Decoction promotes neurogenesis via
sirtuin 1/autophagy pathway in a cerebral ischemia model. Mol Med
Rep. 24:7912021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lee J, Kang CG, Park CR, Hong IK and Kim
DY: The neuroprotective effects of pregabalin after cerebral
ischemia by occlusion of the middle cerebral artery in rats. Exp
Ther Med. 21:1652021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
He Z, Ning N, Zhou Q, Khoshnam SE and
Farzaneh M: Mitochondria as a therapeutic target for ischemic
stroke. Free Radic Biol Med. 146:45–58. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Huang P, Wu SP, Wang N, Seto S and Chang
D: Hydroxysafflor yellow A alleviates cerebral ischemia reperfusion
injury by suppressing apoptosis via mitochondrial permeability
transition pore. Phytomedicine. 85:1535322021. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang R, Zhu Y, Ren C, Yang S, Tian S, Chen
H, Jin M and Zhou H: Influenza A virus protein PB1-F2 impairs
innate immunity by inducing mitophagy. Autophagy. 17:496–511. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lyu W, Li Q, Wang Y, Du C, Feng F, Chi H,
Li Y, Liu W and Sun H: Computational design of binder as the
LC3-p62 protein-protein interaction. Bioorg Chem. 115:1052412021.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Deng S, Liu S, Jin P, Feng S, Tian M, Wei
P, Zhu H, Tan J, Zhao F and Gong Y: Albumin reduces oxidative
stress and neuronal apoptosis via the ERK/Nrf2/HO-1 pathway after
intracerebral hemorrhage in rats. Oxid Med Cell Longev.
2021:88913732021. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cai Y, Yang E, Yao X, Zhang X, Wang Q,
Wang Y, Liu J, Fan W, Yi K, Kang C and Wu J: FUNDC1-dependent
mitophagy induced by tPA protects neurons against cerebral
ischemia-reperfusion injury. Redox Biol. 38:1017922021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yang L, Tao Y, Luo L, Zhang Y, Wang X and
Meng X: Dengzhan Xixin injection derived from a traditional Chinese
herb Erigeron breviscapus ameliorates cerebral ischemia/reperfusion
injury in rats via modulation of mitophagy and mitochondrial
apoptosis. J Ethnopharmacol. 288:1149882022. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Duan C, Wang L, Zhang J, Xiang X, Wu Y,
Zhang Z, Li Q, Tian K, Xue M, Liu L and Li T: Mdivi-1 attenuates
oxidative stress and exerts vascular protection in ischemic/hypoxic
injury by a mechanism independent of Drp1 GTPase activity. Redox
Biol. 37:1017062020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gu C, Li L, Huang Y, Qian D, Liu W, Zhang
C, Luo Y, Zhou Z, Kong F, Zhao X, et al: Salidroside ameliorates
mitochondria-dependent neuronal apoptosis after spinal cord
ischemia-reperfusion injury partially through inhibiting oxidative
stress and promoting mitophagy. Oxid Med Cell Longev.
2020:35497042020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Song M, Zhou Y and Fan X: Mitochondrial
quality and quantity control: mitophagy is a potential therapeutic
target for ischemic stroke. Mol Neurobiol. 59:3110–3123. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
He Q, Li Z, Wang Y, Hou Y, Li L and Zhao
J: Resveratrol alleviates cerebral ischemia/reperfusion injury in
rats by inhibiting NLRP3 inflammasome activation through
Sirt1-dependent autophagy induction. Int Immunopharmacol.
50:208–215. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Han B, Li S, Lv Y, Yang D, Li J, Yang Q,
Wu P, Lv Z and Zhang Z: Dietary melatonin attenuates
chromium-induced lung injury via activating the
Sirt1/Pgc-1alpha/Nrf2 pathway. Food Funct. 10:5555–5565. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Li M, Li SC, Dou BK, Zou YX, Han HZ, Liu
DX, Ke ZJ and Wang ZF: Cycloastragenol upregulates SIRT1
expression, attenuates apoptosis and suppresses neuroinflammation
after brain ischemia. Acta Pharmacol Sin. 41:1025–1032. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Song W, Liu ML, Zhao ZJ, Huang CQ, Xu JW,
Wang AQ, Li P and Fan YB: SIRT1 inhibits high shear stress-induced
apoptosis in rat cortical neurons. Cell Mol Bioeng. 13:621–631.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Duan J, Cui J, Zheng H, Xi M, Guo C, Weng
Y, Yin Y, Wei G, Cao J, Wang Y, et al: Aralia taibaiensis protects
against I/R-induced brain cell injury through the Akt/SIRT1/FOXO3a
pathway. Oxid Med Cell Longev. 2019:76097652019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Tang BL: Sirt1 and the mitochondria. Mol
Cells. 39:87–95. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Huang S, Hong Z, Zhang L, Guo J, Li Y and
Li K: CERKL alleviates ischemia reperfusion-induced nervous system
injury through modulating the SIRT1/PINK1/Parkin pathway and
mitophagy induction. Biol Chem. 403:691–701. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Shao Z, Dou S, Zhu J, Wang H, Xu D, Wang
C, Cheng B and Bai B: Apelin-36 protects HT22 cells against
oxygen-glucose deprivation/reperfusion-induced oxidative stress and
mitochondrial dysfunction by promoting SIRT1-mediated
PINK1/Parkin-dependent mitophagy. Neurotox Res. 39:740–753. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Xu DH, Bao XQ, Wu XW, Xing Y and Tan CY:
Metabolic engineering study on biosynthesis of 4-hydroxybenzyl
alcohol from L-tyrosine in Escherichia coli. Zhongguo Zhong Yao Za
Zhi. 47:906–912. 2022.(In Chinese). PubMed/NCBI
|