Introduction
Non-alcoholic fatty liver disease (NAFLD) is a
chronic liver disease characterized by the accumulation of
excessive fat in the liver, which is not attributed to alcohol
consumption and is a major global health burden (1,2).
Recently, after a Delphi consensus process involving global
experts, the novel term metabolic dysfunction-associated steatotic
liver disease (MASLD) was proposed to replace NAFLD (3–5).
MASLD includes a range of liver conditions, from mild steatosis to
more severe non-alcoholic steatohepatitis (NASH), which is
characterized by inflammation and damage to liver cells (6). The prevalence of MASLD is increasing
globally, and it is predicted that by 2030, ~300 million Chinese,
100 million Americans and 15–20 million Europeans will be affected
(7). Currently, there is no
standard treatment for NASH except improvements in diet and
lifestyle (8). The pathogenesis of
MASLD is complex and lipid metabolism disorders play a key role in
it (9). Excess fatty acid
accumulation in the liver due to disorders of lipid metabolism is
the most important pathogenetic cause of MASLD (10,11).
These lipotoxic metabolites lead to hepatocyte stress, injury and
death, leading to the progression of MASLD (12). Prior research has demonstrated the
widespread use of sorafenib, a multi-kinase inhibitor, for the
treatment of liver cancer (13).
Low doses of sorafenib, a mitochondrial uncoupling agent, inhibits
NASH progression in mice by activating the AMPK pathway (14). Donafenib, a novel oral multi-kinase
inhibitor, has been developed to replace the hydrogen atom in the
methyl group of sorafenib with three deuterium atoms, and has
similar therapeutic effects as sorafenib (15). Similar to sorafenib, donafenib may
be a potential therapeutic agent for treating MASLD. Statins are
frequently prescribed to lower blood lipid levels and have been
demonstrated to play a crucial role in fat metabolism (16) and can act on the AMPK signaling
pathway through various mechanisms (17). Therefore, the present study
examined the potential effects of low-dose donafenib combined with
atorvastatin on MASLD and explored the underlying mechanisms with
the aim to provide a promising treatment strategy for MASLD and
contribute to an improved understanding of its pathogenesis.
Materials and methods
Cell culture
The HepG2 cell line is derived from human liver
cancer and was used as an in vitro model to study hepatocyte
metabolism because of its homologous biotransforming metabolic
enzymes to normal human hepatocytes (18,19).
The HepG2 cells (Procell Life Science & Technology Co., Ltd)
were cultured in 10% fetal bovine serum and 1%
penicillin/streptomycin in addition to Dulbecco's modified eagle
medium (Thermo Fisher Scientific, Inc.). Before the experiments,
the cells were assessed for mycoplasma contamination and the assay
was negative. Mycoplasma contamination was detected by
chemiluminescence for 15 min at room temperature using a Mycoplasma
test kit (Wuhan Servicebio Technology Co., Ltd.). The cell line was
authenticated by the manufacturer of the cells using Short Tandem
Repeat profiling to ensure that the HepG2 cells were not
misidentified and cross-contaminated. The cells were cultured at
37°C in a humidified incubator with 5% CO2 to ensure
proper cell growth and maintenance.
Cell viability assay
For the Cell Counting Kit-8 (CCK-8) assay, a CCK-8
kit (Wuhan Servicebio Technology Co., Ltd.) was used. Cells were
collected after centrifugation at 500 × g) for 5 min at 4°C,
resuspended in fresh medium and seeded at a density of five million
cells/well in 96-well plates containing 200 µl of medium per well.
Atorvastatin (Shanghai Aladdin Biochemical Technology Co., Ltd.) at
5, 10, 20, 40, 80, 160 µM and donafenib (Suzhou Zelgen
Biopharmaceuticals Co., Ltd.) at 1, 2, 4, 8, 16, 32 µM were added
to the wells, and the plates were incubated for 24 h at 37°C. After
incubation, the medium was aspirated, and 10 µl of medium
containing CCK-8 solution was added to each well. Then, the plates
were incubated for an additional hour, and a microplate reader
(BioTek; Agilent Technologies, Inc.) was used to measure the
absorbance at 450 nm.
Cell modeling of MASLD
To prepare the free fatty acid (FFA) mixture, fatty
acid-free bovine serum albumin, sodium oleate and sodium palmitate
(2:1, sodium oleate to sodium palmitate) were used (20). Next, this FFA mixture was used to
construct a cellular model of MASLD. HepG2 cells were in the
logarithmic growth phase, the cells were divided into five groups:
i) CTRL group (without treatment); ii) FFA group (treated with the
FFA mixture); iii) FFA-ATO group (treated with the FFA mixture and
atorvastatin); iv) FFA-DON group (treated with the FFA mixture and
donafenib); and v) FFA-DON + ATO group (treated with FFA mixture,
donafenib and atorvastatin). Following serum starvation, the FFA
and other treatment groups were exposed to high-fat medium
containing 1 mM FFA for 24 h at 37°C, whereas the CTRL group
remained untreated. The FFA-ATO, FFA-DON and FFA-DON + ATO groups
were also treated with atorvastatin, donafenib or both,
respectively, for 24 h at 37°C. The next day, the cells were
collected and analyzed.
AMPK inhibitor
Compound C has been described to have effects on
other kinases but is currently widely used as an AMPK inhibitor
(21,22). In the present study, Compound C
(Shanghai Aladdin Biochemical Technology Co., Ltd.) was used to
inhibit AMPK activity. In vitro, compound C (5 µM) was added
to the FFA group, FFA-ATO group and FFA-ATO + DON group. After 24 h
of incubation at 37°C, expression of AMPK and p-AMPK were analyzed
in each group and the accumulation of fatty acids in the cells of
each group was observed using Oil Red O staining.
Animals
A total of male 40 Sprague-Dawley rats weighing
180–200 g (6–8 weeks) were obtained from The Laboratory Animal
Center at Tongji Medical College (Huazhong University of Science
and Technology; Wuhan, China). Rats were maintained under specific
pathogen-free conditions at 23±2°C, 55±5% relative humidity, and a
12-h light/dark cycles and had free access to food and water. All
experimental procedures were approved by The Ethics Committee of
Tongji Medical College, Huazhong University of Science and
Technology (Wuhan, China; approval no. 3462) and adhered to the
ARRIVE guidelines (arriveguidelines.org/).
Treatment regimens with atorvastatin
and donafenib
The rats were randomly divided into five groups: i)
CTRL (n=8); ii) high-fat diet (HFD; n=8); iii) HFD-ATO (n=8); iv)
HFD-DON (n=8); and v) HFD-DON + ATO (n=8). Rats in the CTRL group
were fed a regular diet (Table
SI) throughout the experimental period, whereas those in the
other four groups were fed a HFD (Table SII). A previous study has shown
that a 12-week HFD can successfully establish a MASLD model in rats
(23). During the first 12 weeks,
no intervention was administered to any rats. At the beginning of
week 13, the rats in the HFD-ATO, HFD-DON and HFD-DON + ATO groups
were treated with drug (0.5 ml) daily by gavage for 4 weeks. Rats
in the CTRL and HFD groups were administered dimethyl sulfoxide
(0.5 ml) by gavage. Body weight was measured during the experiment,
and the drug amount was adjusted according to the evolution of the
weight of the rats during the study period. Atorvastatin and
donafenib were dissolved in a dimethyl sulfoxide solution, and the
dose of atorvastatin was 10 mg/kg/day, based on previous studies
(24,25). Owing to possible liver and kidney
toxicity, a low dose of 1 mg/kg/day donafenib was used (14).
Tissue collection
The rats were fasted for 24 h at the end of the
16-week experiment. Sodium pentobarbital 150 mg/kg was injected
intraperitoneally, and the rats were executed by overdose of
anesthesia. Blood was drawn from the inferior vena cava after
opening the abdominal cavity. After centrifugation at 1,500 × g)
for 10 min at 4 °C, the serum was collected. The liver, heart,
spleen, lung and kidneys were then collected and weighed, and the
liver index was calculated as the ratio of liver weight to body
weight. The serum and liver tissue samples were stored in a
refrigerator at −80°C for subsequent testing.
Biochemical analyses
The corresponding diagnostic kits (Thermo Fisher
Scientific, Inc.) were used to measure the serum concentrations of
FFA, total triglycerides (TG), total cholesterol (TC), high-density
lipoprotein cholesterol (HDL-C), low-density lipoprotein
cholesterol (LDL-C), very-low-density lipoprotein cholesterol
(VLDL-C), creatinine (CRE) and blood urea nitrogen (BUN), as well
as the levels of alanine aminotransferase (ALT) and aspartate
aminotransferase (AST). Part of the liver tissue was prepared as a
liver homogenate, and the lipids were extracted according to
previously described methods (26). The hepatic lipid profiles were
analyzed using the same method as that used for serum lipid profile
assays.
H&E staining and Oil red O
staining
The rat liver samples were fixed in 4% (w/v)
paraformaldehyde at 4°C for 6 h. After dehydration with graded
ethanol solutions, tissues were embedded in paraffin wax. A serial
frontal section was cut at intervals of 5 µm and stained with
H&E staining at room temperature (hematoxylin staining for 3
min and eosin staining for 15 sec) for histopathology. The sections
were visualized under a light microscope.
For HepG2 cells, after the intervention, they were
fixed with 4% paraformaldehyde for 25–30 min at room temperature.
Fresh liver tissue measuring 1.0×1.0 cm was cut and embedded in a
frozen section embedding agent and frozen at −20°C for section
processing. The cells and frozen liver tissue were then stained
with Oil Red O dye for 10 min at room temperature, and the
morphological changes in the cells or tissues and deposition of
lipid droplets were observed under a fluorescence microscope
(Olympus Corporation).
Western blotting analysis
Protein samples were prepared in lysis buffer
(Thermo Fisher Scientific, Inc.; 25 mmol/l HEPES, Kac 150 mmol/l,
EDTA pH 8.0 2 mmol/l, NP-40 0.1%, NaF 10 mmol/l, PMSF 50 mmol/l,
aprotinin 1 µg/µl, pepstatin 1 µg/µl, leupeptin 1 µg/µl, DTT 1
mmol/l). The protein concentrations were quantified by BCA protein
assay (Thermo Fisher Scientific, Inc.). Gel electrophoresis was
performed with 10–20 µg protein using 4–15% gels (Thermo Fisher
Scientific, Inc.), followed by transblotting to 0.2 µm
nitrocellulose membrane. Subsequently, the membranes were blocked
using 5% skimmed milk at room temperature for 2 h. Next, the
membranes were incubated at 4°C overnight with the primary
antibodies against p-AMPK (1:1,000; Abcam; cat. no. ab133448), AMPK
(1:1,000; Abcam; cat. no. ab32047), sterol regulatory
element-binding protein-1 (SREBP-1; 1:1,000; Abcam; cat. no.
ab28481), 3-hydroxy-3-methylglutaryl-CoA reductase (HMGCR; 1:1,000;
Abcam; cat. no. ab242315), acyl-CoA oxidase 1 (ACOX1; 1:1,000;
Abcam; cat. no. ab184032), carnitine palmitoyl-transferase 1C
(CPT1C; 1:1,000; Abcam; cat. no. ab189182) and β-actin (1:1,000;
Abcam; cat. no. ab8226) in all blocking membranes, and then
immersed in a goat anti-rabbit horseradish peroxidase-conjugated
IgG secondary antibody (1:3,000; Abcam; cat. no. ab6721) or a goat
anti-mouse horseradish peroxidase-conjugated IgG secondary antibody
(1:3,000; Abcam; cat. no. ab150113) at room temperature for 2 h.
Finally, the blots were visualized by enhanced chemiluminescence
(Thermo Fisher Scientific, Inc.), and the results were analyzed
using ImageJ software (version 1.0; National Institutes of Health)
with β-actin as the loading control.
Immunohistochemical staining
Immunohistochemical (IHC) staining was performed on
formalin-fixed, paraffin-embedded samples as previously described
(27). Briefly, the slides were
de-paraffinized at room temperature, washed with xylene and
rehydrated in descending ethanol series. The liver sections were
subjected to antigen retrieval with 0.01 M sodium
citrate-hydrochloric acid buffer solution and their endogenous
peroxidase activity was blocked for 30 min in 1% hydrogen
peroxide/phosphate-buffered saline (H2O2/PBS)
solution. 3% BSA (Wuhan Servicebio Technology Co., Ltd.) was used
for blocking for 30 min at room temperature. Subsequently, the
sections were incubated with the primary antibody against fatty
acid synthase (FAS; 1:1,000; Abcam; cat. no. ab 128870) overnight
at 4°C. Then the sections were incubated with a goat anti-rabbit
horseradish peroxidase-conjugated IgG secondary antibody (1:3,000;
Abcam; cat. no. ab6721) at room temperature for 2 h, and the
sections were washed three times with PBS, followed by
immunostaining with 3,3′-diaminobenzidine for 5 min at room
temperature and counterstaining of the nuclei with hematoxylin for
3 min at room temperature. Tissue staining was visualized under a
fluorescence microscope (Olympus) after dehydration and sealing of
the slides. Images were acquired and analyzed using a Nikon DS-U3
microscope (Nikon Corporation). Fatty acid synthase (FAS)
antibodies (1:800; Abcam) were used for immunohistochemical
analysis of FAS expression. IHC staining results were analyzed
using ImageJ software (version 1.0; National Institutes of
Health).
ROS content in liver tissue
A dihydroethidium (DHE) Assay Kit (Abcam; cat. no.
ab236206) was used to measure the ROS content. DHE was incubated
with the cells at 37°C for 30 min protected from light, and then
the ROS content was observed under a fluorescence microscope. DHE
freely enters the cell through the living cell membrane and
exhibits blue fluorescence in the cytoplasm. After oxidation by
intracellular ROS, DHE forms ethidium oxide, which is incorporated
into the chromosomal DNA to produce bright red fluorescence in the
nucleus. Eventually, DNA in the nucleus and cytoplasm appears red.
The amount and change in cellular ROS content was measured
according to the amount of red fluorescence.
Statistical analysis
Data are presented as mean ± standard deviation.
GraphPad Prism software (version 8.0; Dotmatics) was used for data
visualization and SPSS software (version 26.0; IBM Corp.) was used
for statistical analysis. Unpaired Student's t test was employed to
analyze differences between two groups. One-way analysis of
variance was used to analyze the differences between groups when
data was normally distributed. For multiple comparisons between
groups, the least significant difference test was used to compare
the three groups, Dunnett's test was used for the comparison
between different experimental groups to a single control and Tukey
test was performed for multiple comparisons among ≥3 groups. For
datasets with a skewed distribution, the Kruskal-Wallis test was
used for multiple comparisons followed by the post hoc Dunn's test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Donafenib and atorvastatin reduce
lipid accumulation in HepG2 cells treated with FFA
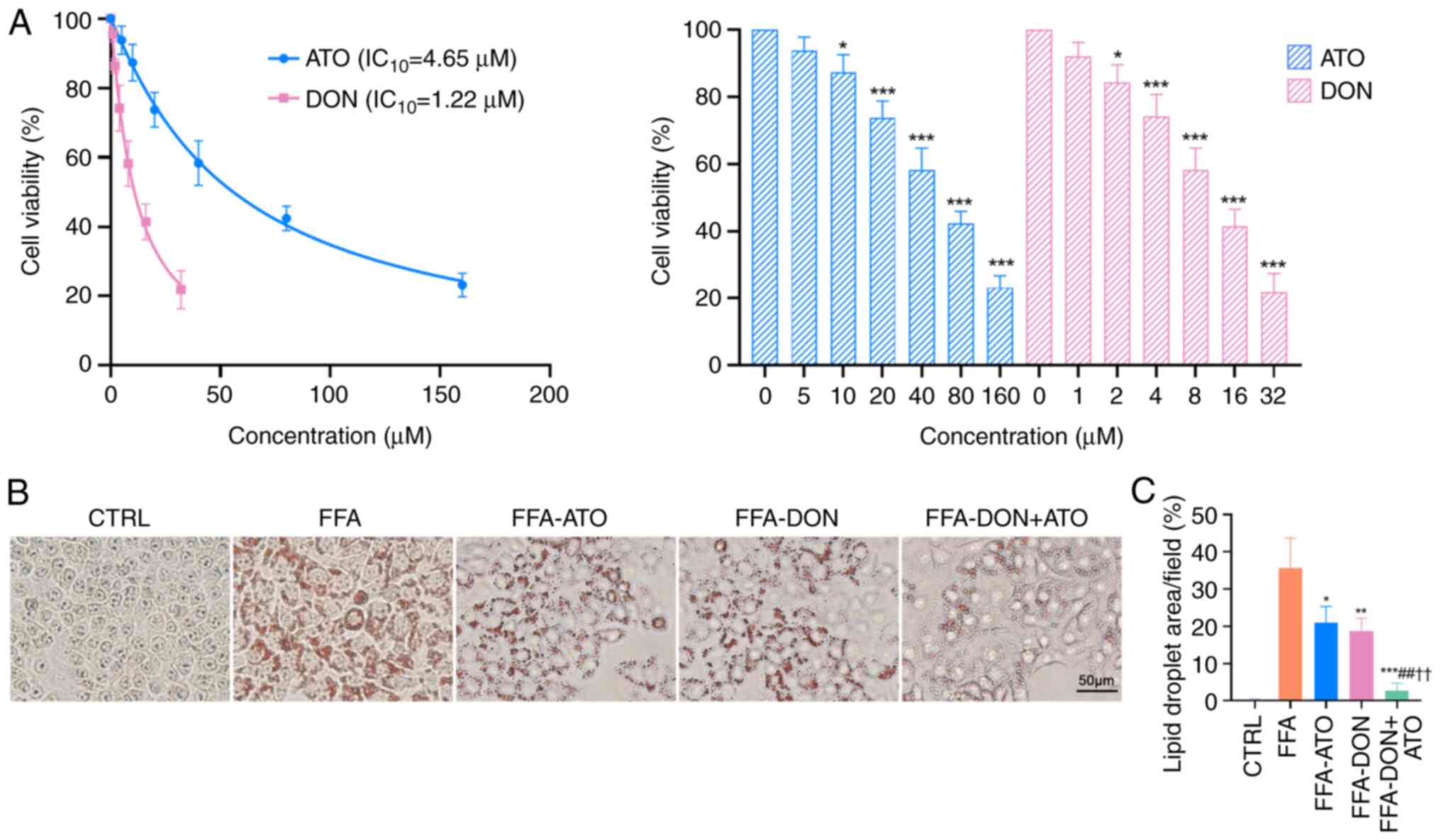
Treatment of HepG2 cells with different
concentrations of donafenib and atorvastatin decreased cell
viability (Fig. 1A). Specifically,
1.22 µM of donafenib achieved a 10% inhibition rate of HepG2 cells,
whereas treatment with 4.65 µM of atorvastatin achieved a similar
inhibition rate. On the basis of these results, HepG2 cells were
treated with 1.22 µM of donafenib and 4.65 µM of atorvastatin. Oil
Red O staining of HepG2 cells (Fig.
1B) revealed significant lipid accumulation in the FFA group,
whereas lipid accumulation in the FFA-ATO, FFA-DON and FFA-DON +
ATO groups was reduced to varying degrees, with the most
significant reduction observed in the FFA-DON + ATO group. The
effects of high concentrations of donafenib and atorvastatin on
fatty acid metabolism were also analyzed. Higher concentrations of
donafenib and atorvastatin inhibited fatty acid accumulation more
significantly than lower concentrations; simultaneously, cell
proliferation was significantly inhibited (Fig. S1).
Donafenib and atorvastatin improves
MASLD in rats
A 12-week HFD successfully induced MASLD in rats
(Fig. 2A). H&E staining
revealed that the livers of rats in the HFD group exhibited
steatosis, hepatocyte ballooning and extensive lobular
inflammation, however, the three treatment groups showed varying
degrees of improvement. According to the NAFLD Activity Score (NAS)
(24) of the liver histological
results (Fig. 2B), the HFD-DON +
ATO group had the lowest mean NAS, indicating that the combination
treatment effectively improved hepatocyte steatosis and
inflammation. The liver tissue was weighed and the liver index of
rats in the HFD group was higher than that in the other groups
(Fig. 2C). Similarly, Oil Red O
staining of liver tissue revealed significant liver fat
accumulation in the HFD group, whereas that in the three treatment
groups showed varying degrees of improvement in fat accumulation
(Fig. 2D). The most significant
decrease in fat accumulation was observed in the HFD-DON + ATO
group; however, it did not reach the level observed in the CTRL
group.
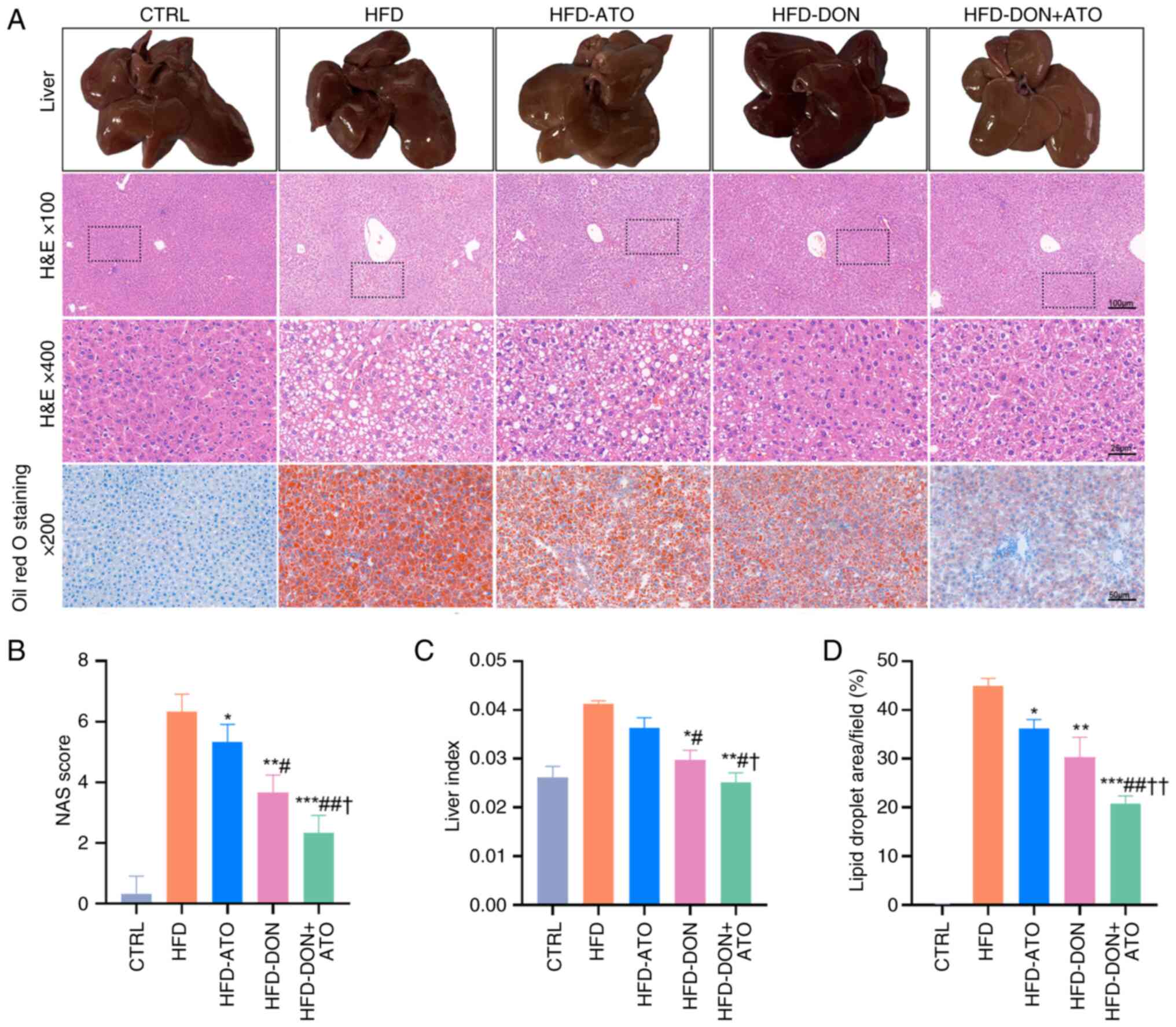 | Figure 2.Effects of donafenib and atorvastatin
on the histology of rats with MASLD. (A) Histological sections of
rats in each group were shown (n=8). (B) NAS, which is the
unweighted sum of semi-quantitative scores for steatosis, lobular
inflammation and hepatocyte ballooning, was calculated. (C) Liver
index of rats (liver wet weight/rat body weight) was measured. (D)
Oil Red O staining was used to determine lipid droplet deposition
in hepatocytes. Data are presented as the means ± SD. A
Kruskal-Wallis test was used for statistical analysis of the data
in (B). One-way ANOVA (Tukey was used for post hoc multiple
comparisons) was used for (C) and (D). *P<0.05, **P<0.01,
***P<0.001 vs. HFD group; #P<0.05,
##P<0.01 vs. HFD-ATO; †P<0.05,
††P<0.01 vs. HFD-DON group. NAS, NAFLD activity
score; ATO, atorvastatin; DON, donafenib; MASLD, metabolic
dysfunction-associated steatotic liver disease; HFD, high-fat
diet. |
Donafenib and atorvastatin regulate
the activation of AMPK
The AMPK pathway plays a crucial role in fat
metabolism, and the expression levels of AMPK in cells and liver
tissues were analyzed using western blotting. In vitro, the
expression of p-AMPK in the FFA group decreased, and the
p-AMPK/AMPK ratio was significantly lower than that in the CTRL
group (Fig. 3A and B). In
vivo, similar results were observed, with the p-AMPK/AMPK ratio
in the livers of rats in the HFD group being significantly lower
than that in the livers of rats in the CTRL group (Fig. 4A and B). The p-AMPK/AMPK ratios in
the three treatment groups were higher than that in the HFD group,
with the HFD-DON + ATO group showing the most significant increase.
When Compound C was added, p-AMPK expression was suppressed in the
CTRL group (Fig. 5A and B). There
was no significant difference in p-AMPK expression in the FFA-ATO
and FFA-DON groups compared to the FFA group (Fig. 5C and D) and the results of Oil Red
O staining showed that the FFA-ATO and FFA-DON groups were not
significantly different from the FFA group (Fig. 5E and F) when compound C was added.
This indicated that the addition of Compound C caused atorvastatin
and donafenib to lose their pharmacological effect of improving
MASLD and proved that donafenib and atorvastatin improved MASLD by
activating AMPK.
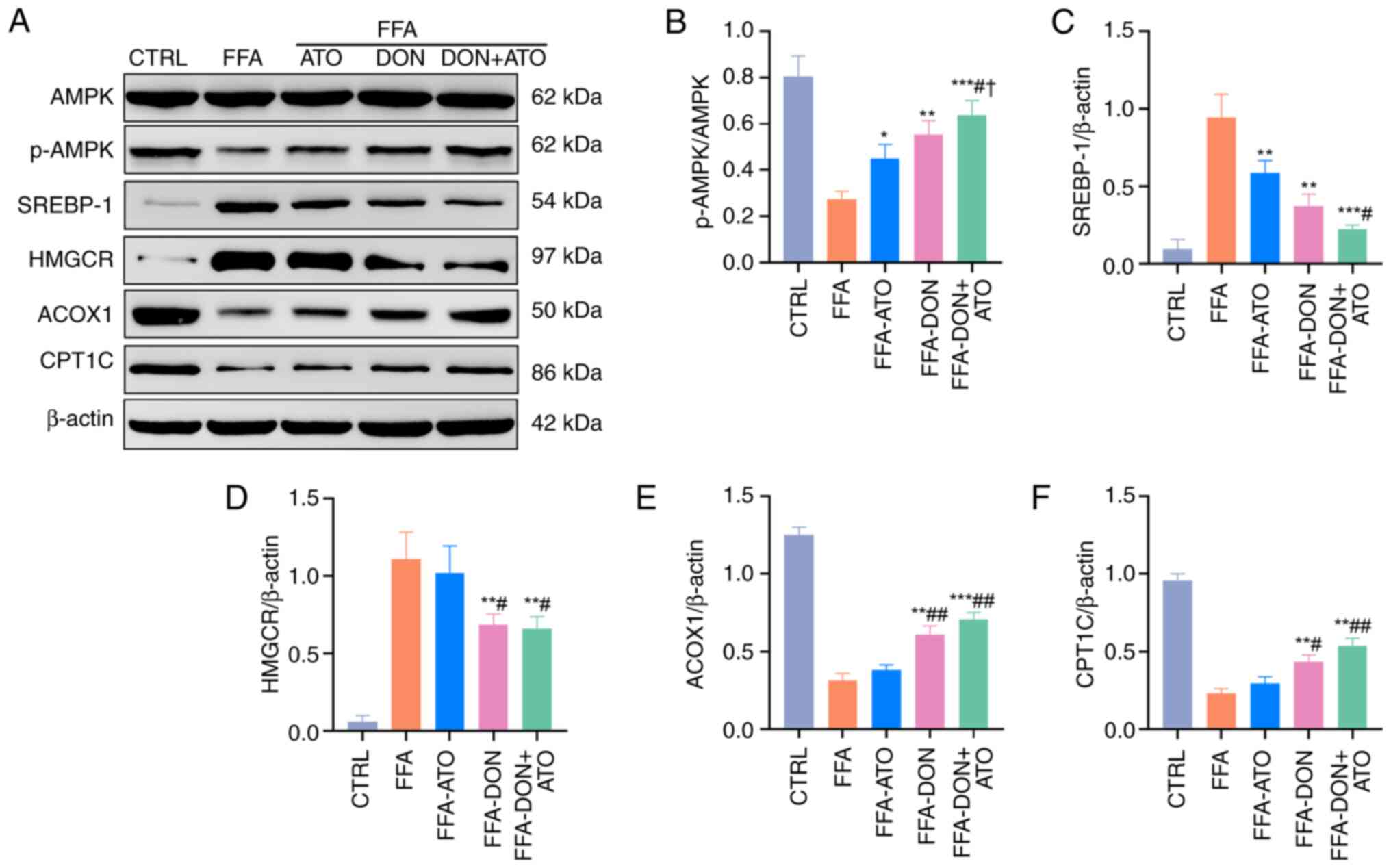 | Figure 3.Effects of donafenib and atorvastatin
on lipid metabolism of HepG2 cells. (A) Representative blots for
proteins involved in lipid metabolism were analyzed. (B-F)
Quantitative analysis of p-AMPK/AMPK, SREBP-1, HMGCR, ACOX1 and
CPT1C were shown. Data were presented as the means ± SD and
analyzed by one-way ANOVA (Tukey was used for post hoc multiple
comparisons). *P<0.05, **P<0.01, ***P<0.001 vs. FFA group;
#P<0.05, ##P<0.01 vs. FFA-ATO group;
†P<0.05 vs. FFA-DON group. FFA, free fatty acid; ATO,
atorvastatin; DON, donafenib; SREBP-1, sterol regulatory
element-binding protein-1; CPT1C, carnitine palmitoyl-transferase
1C; HMGCR, 3-hydroxy-3-methylglutaryl-CoA reductase; ACOX1,
acyl-CoA oxidase 1. |
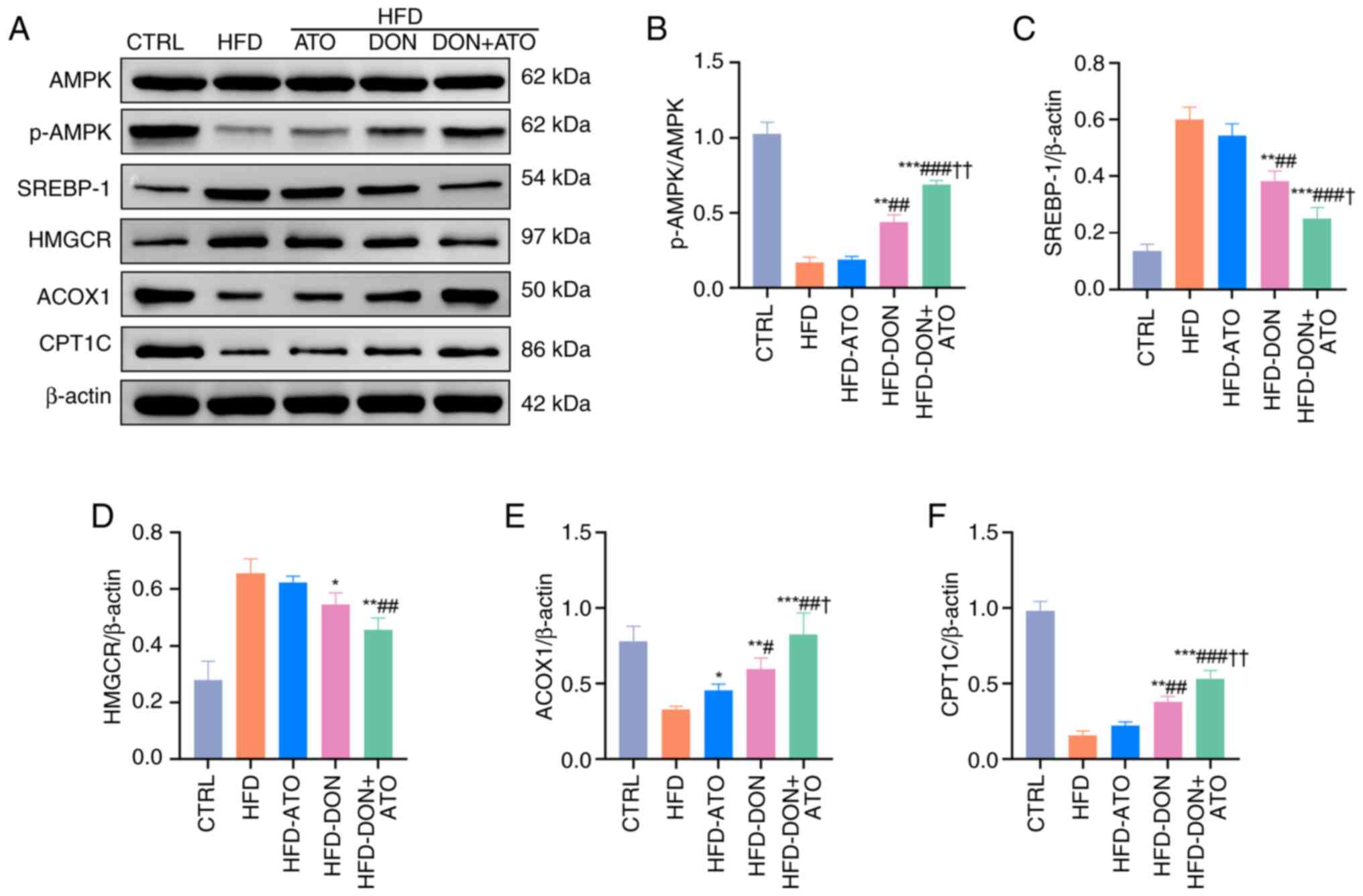 | Figure 4.Effects of donafenib and atorvastatin
on lipid metabolism in the liver of rats with MASLD. (A)
Representative blots for proteins involved in lipid metabolism were
analyzed. (B) Quantitative analysis of (B) p-AMPK/AMPK. (C)
Quantitative analysis of SREBP-1. (D) HMGCR. (E) Quantitative
analysis of ACOX1. (F) Quantitative analysis of CPT1C. Data were
presented as the means ± SD and analyzed by one-way ANOVA (Tukey
was used for post hoc multiple comparisons). *P<0.05,
**P<0.01, ***P<0.001 vs. HFD group; #P<0.05,
##P<0.01, ###P<0.001 vs. HFD-ATO group;
†P<0.05, ††P<0.01 vs. HFD-DON group.
HFD, high-fat diet; ATO, atorvastatin; DON, donafenib; MASLD,
metabolic dysfunction-associated steatotic liver disease; SREBP-1,
sterol regulatory element-binding protein-1; CPT1C, carnitine
palmitoyl-transferase 1C; HMGCR, 3-hydroxy-3-methylglutaryl-CoA
reductase; ACOX1, acyl-CoA oxidase 1. |
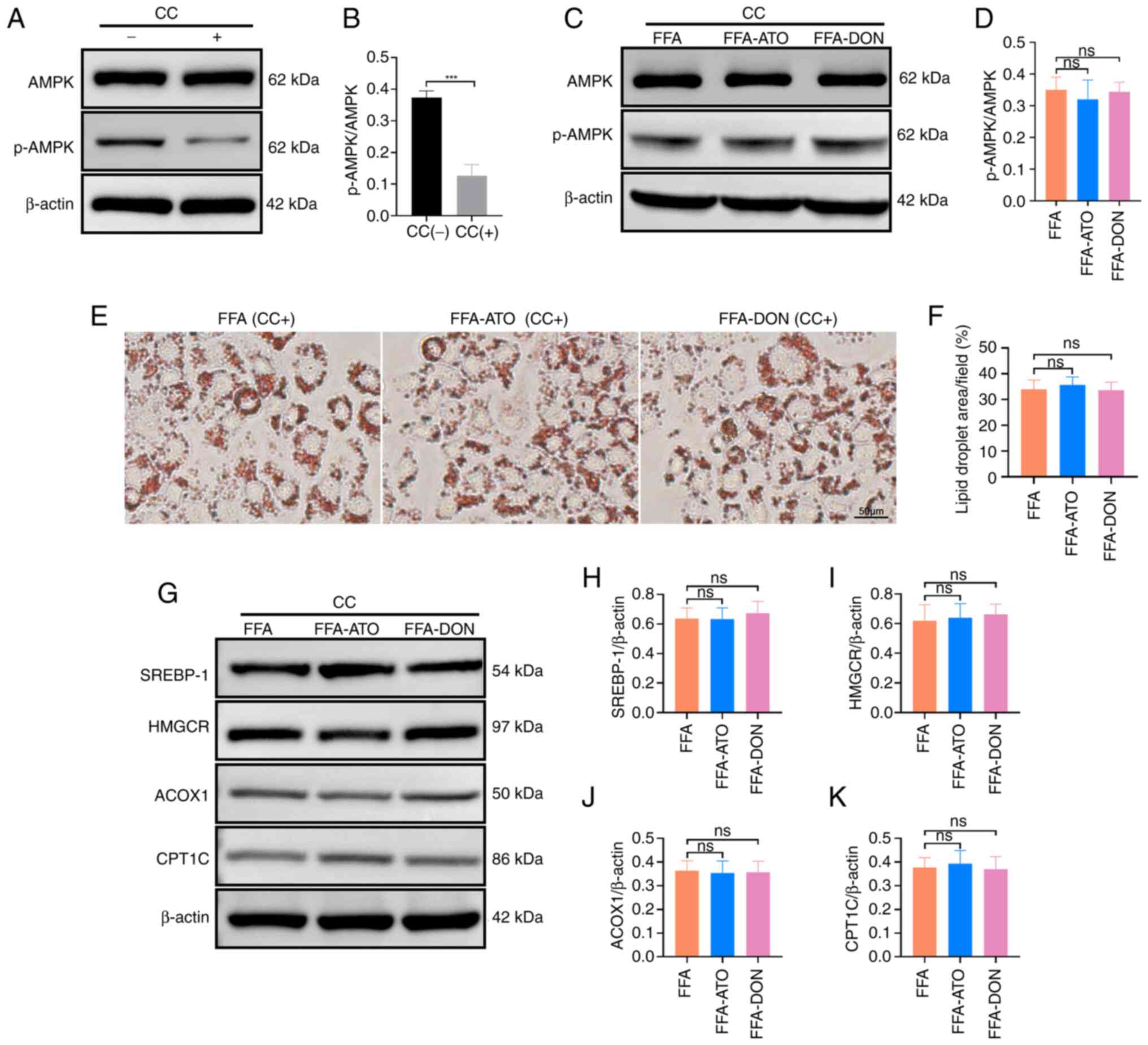 | Figure 5.Addition of AMPK inhibitor, CC,
prevented donafenib and atorvastatin from affecting fatty acid
metabolism. (A) Representative blots of AMPK and p-AMPK with or
without the addition of CC in the CTRL group. (B) Quantitative
analysis of the expressions of AMPK and p-AMPK with or without the
addition of CC in the CTRL group. (C) Representative blots of AMPK
and p-AMPK expressions with the addition of CC in the FFA, FFA-ATO
and FFA-DON groups. (D) Quantitative analysis of the expressions of
AMPK and p-AMPK in the FFA, FFA-ATO and FFA-DON groups with the
addition of CC. (E) Effects of donafenib and atorvastatin on fatty
acid accumulation in the FFA, FFA-ATO and FFA-DON groups with the
addition of CC were analyzed by Oil Red O staining. (F)
Quantitative analysis of Oil Red O staining in the FFA, FFA-ATO and
FFA-DON groups after adding CC. (G) Representative blots of lipid
metabolism-related proteins in the FFA, FFA-ATO and FFA-DON groups
with the addition of CC. (H-K) Quantitative analysis of the
expressions of lipid metabolism-related proteins in the FFA,
FFA-ATO and FFA-DON groups with the addition of CC. The data were
presented as the means ± SD. For statistical analysis, Student's t
test was used in (B). One-way ANOVA (LSD was used for post hoc
multiple comparisons) was used for the other data. ns, no
significance; CC, Compound C; FFA, free fatty acid; ATO,
atorvastatin; DON, donafenib; SREBP-1, sterol regulatory
element-binding protein-1; CPT1C, carnitine palmitoyl-transferase
1C; HMGCR, 3-hydroxy-3-methylglutaryl-CoA reductase; ACOX1,
acyl-CoA oxidase 1. |
Donafenib and atorvastatin regulate
fatty acid synthesis
Hepatic fatty acids are derived from the hepatic
synthesis of fatty acids and the transport of extrahepatic FFAs to
the liver. SREBP-1, HMGCR and FAS play important roles in de
novo fatty acid synthesis. The expression levels of SREBP-1 and
HMGCR in both cells and liver tissues were analyzed using western
blotting. In vitro, the expressions of SREBP-1 and HMGCR in
the FFA group were significantly higher than those in the CTRL
group, whereas the expressions of SREBP-1 and HMGCR decreased in
the FFA-ATO, FFA-DON and FFA-DON + ATO groups (Fig. 3C and D). In vivo, the
expression levels of SREBP-1 and HMGCR in the three treatment
groups were lower than those in the HFD group, with the HFD-DON +
ATO group exhibiting the most significant decrease (Fig. 4C and D). Immunohistochemistry was
used to detect FAS expression (Fig.
S2). The results showed a significant decrease in FAS
expression in all three treatment groups compared with that in the
HFD group, with the combined treatment group showing the most
significant decrease. When Compound C was added, there was no
significant difference in the expression of SREBP-1 and HMGCR in
the FFA-ATO and FFA-DON groups compared with the FFA group
(Fig. 5G-I).
Donafenib and atorvastatin regulate
fatty acid β-oxidation
Fatty acid β-oxidation is an important location of
fatty acid metabolism in the liver, with mitochondria and
peroxisomes being the sites of β-oxidation. The crucial enzymes for
fatty acid β-oxidation in mitochondria and peroxisomes are CPT1C
and ACOX1, respectively. In vitro, the expression levels of
ACOX1 and CPT1C in the FFA group were significantly lower than
those in the CTRL group, whereas those of ACOX1 and CPT1C in the
three treatment groups were increased (Fig. 3E and F). In vivo, the
expression levels of ACOX1 and CPT1C in the liver of the HFD group
were significantly lower than those of the CTRL group. The levels
of ACOX1 and CPT1C in the three treatment groups were higher than
those in the HFD group, and the increase in the HFD-DON + ATO group
was the most pronounced (Fig. 4E and
F). When Compound C was added, there was no significant
difference in the expression of CPT1C and ACOX1 in the FFA-ATO and
FFA-DON groups compared with the FFA group (Fig. 5J and K).
Donafenib and atorvastatin affect
lipid metabolism parameters
Lipid metabolism parameters in the serum and liver
tissues were analyzed (Fig. 6). In
both the serum and liver tissues, the FFA, TG and TC levels of rats
in the HFD group were significantly higher than those of rats in
the CTRL group. Compared with the HFD group, the aforementioned
parameters of rats in the three treatment groups were lower, which
was consistent with the results of the liver histological analysis.
A decrease in the serum LDL-C levels in the HFD-ATO and HFD-DON +
ATO groups was observed; however, no significant changes were
observed in the HFD-DON group. Additionally, the serum VLDL-C and
HDL-C levels in the three treatment groups were not significantly
different from those in the HFD group.
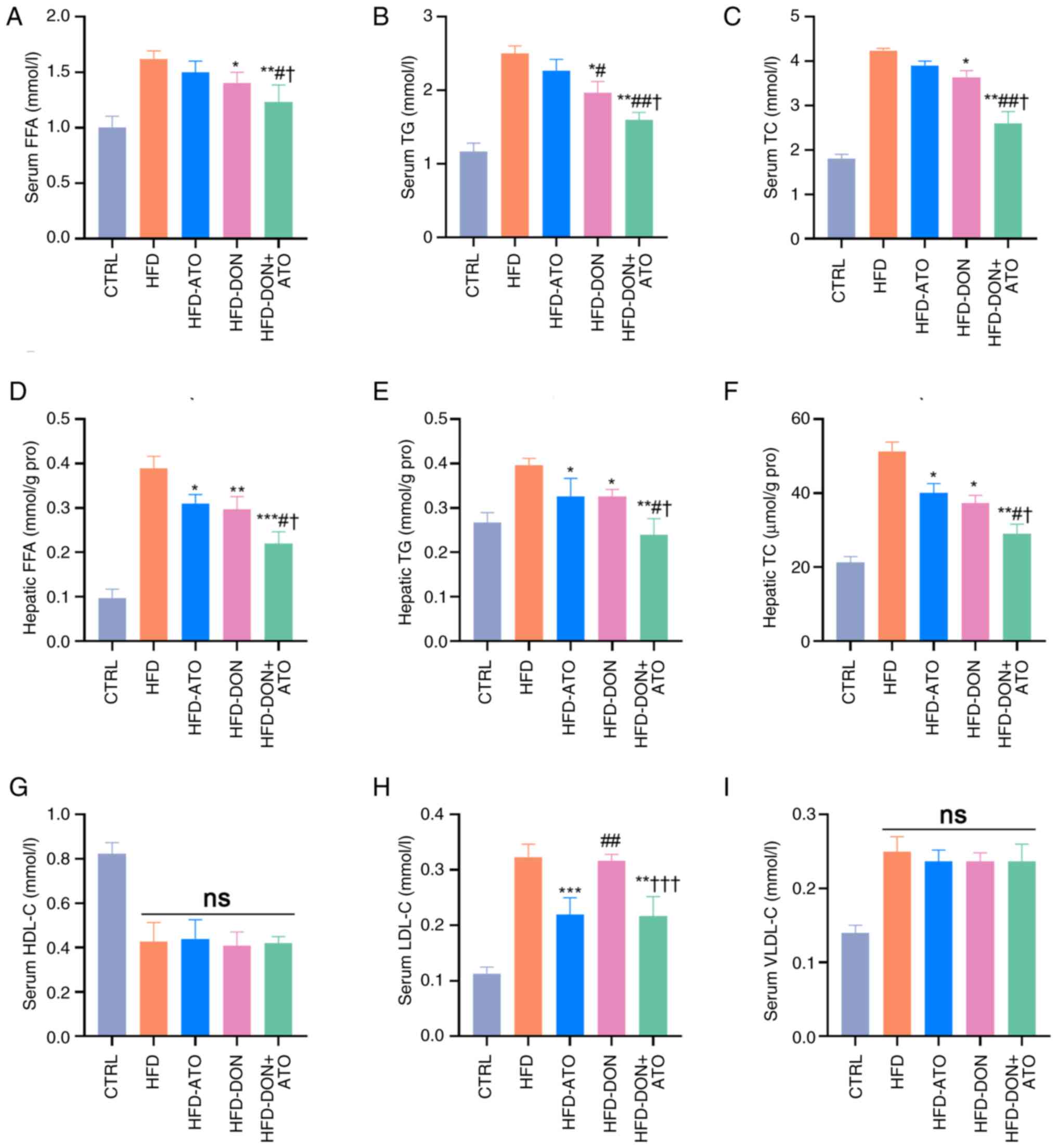 | Figure 6.Effects of donafenib and atorvastatin
on lipid metabolism parameters in the serum and liver. (A) Serum
(A) FFA was measured. (B) Serum TG was measured. (C) Serum TC was
measured. (D) Hepatic FFA was measured. (E) Hepatic TG was
measured. (F) Hepatic TC was measured. (G) Serum HDL-C was
measured. (H) Serum LDL-C was measured. (I) Serum (VLDL-C) was
measured. Data were presented as the means ± SD and analyzed by
one-way ANOVA (Tukey was used for post hoc multiple comparisons).
*P<0.05, **P<0.01, ***P<0.001 vs. HFD;
#P<0.05, ##P<0.01 vs. HFD-ATO group;
†P<0.05, †††P<0.001 vs. HFD-DON group.
ns, no significance. FFA, free fatty acid; TG, triglyceride; TC,
total cholesterol; HDL-C, high-density lipoprotein cholesterol;
LCL-C, low-density lipoprotein cholesterol; VLDL-C,
very-low-density lipoprotein cholesterol; HFD, high-fat diet; ATO,
atorvastatin; DON, donafenib. |
Donafenib and atorvastatin improve
oxidative stress
Oxidative stress plays an important role in the
progression of MASLD, and the level of oxidative stress in liver
tissues was examined. Both atorvastatin and donafenib reduced the
ROS content in the liver tissue (Fig.
7A and B). Donafenib and atorvastatin exhibited different
degrees of antioxidant effects and the ability of donafenib to
reduce ROS was stronger than atorvastatin. In the HFD-DON + ATO
group, ROS levels in the liver tissue decreased significantly and
were only slightly higher than those in the CTRL group. At the same
time, malondialdehyde content in HFD-DON + ATO group was also
significantly lower than that in HFD group (Fig. 7C). In contrast, superoxide
dismutase and glutathione peroxidase levels increased in each
treatment group (Fig. 7D and
E).
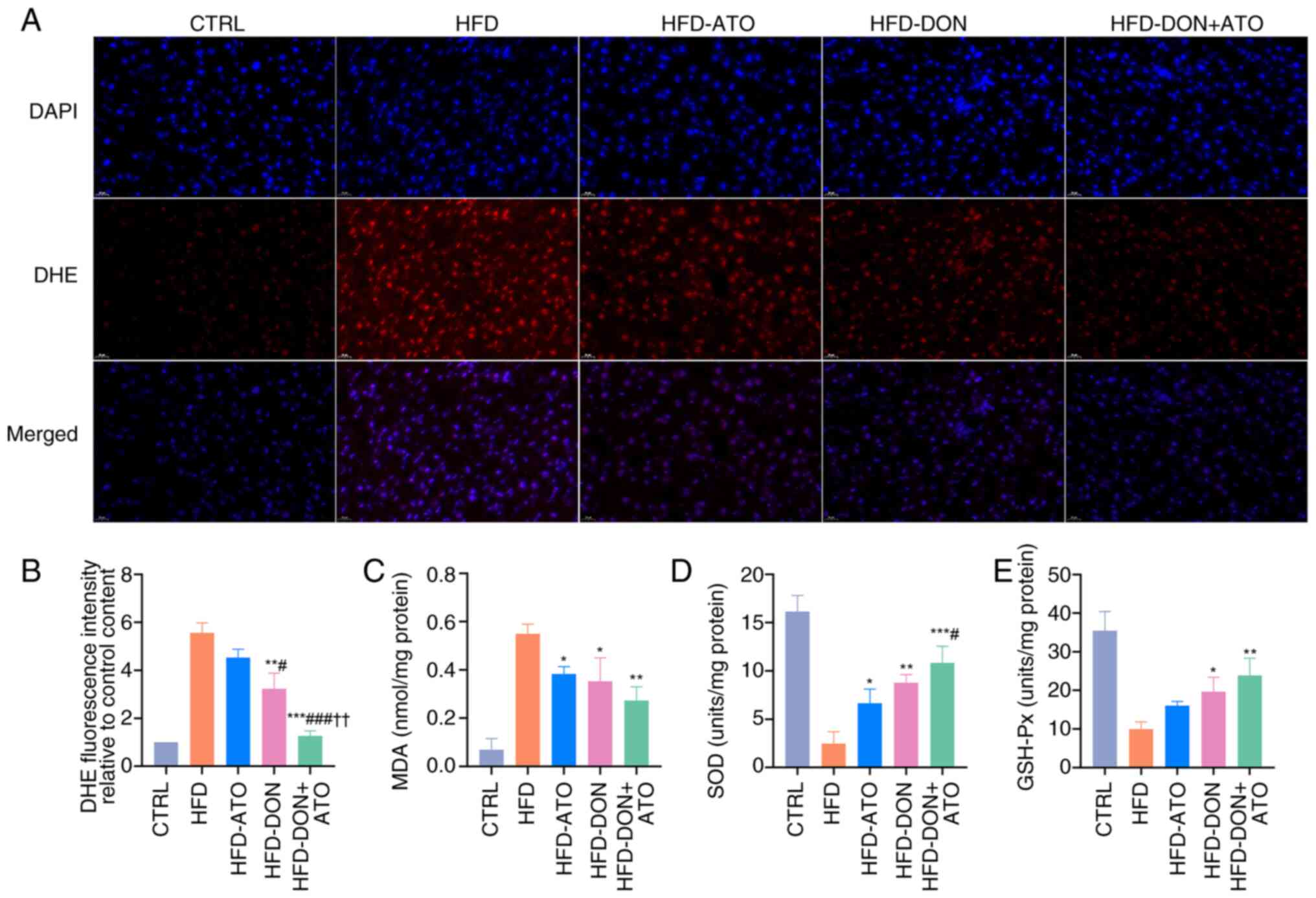 | Figure 7.Effects of donafenib and atorvastatin
on oxidative stress. (A) Representative images of ROS
immunofluorescence. (B) Quantitative analysis of ROS content in
liver tissue. (C) MDA content in liver tissue. (D) SOD content in
liver tissue. (E) GSH-Px content in liver tissue. Data were
presented as the means ± SD and analyzed by one-way ANOVA (Tukey
was used for post hoc multiple comparisons). *P<0.05,
**P<0.01, ***P<0.001 vs. HFD group; #P<0.05,
###P<0.001 vs. HFD-ATO group; ††P<0.01
vs. HFD-DON group. ROS, reactive oxygen species; MDA,
malondialdehyde; SOD, superoxide dismutase; GSH-Px; glutathione
peroxidase; HFD, high-fat diet; ATO, atorvastatin; DON, donafenib;
DHE, dihydroethidium. |
Safety of donafenib and atorvastatin
in improving MASLD of rats
To assess the safety of the administered drug doses,
H&E staining of the heart, spleen, lung and kidney tissues in
all rat groups was performed (Fig.
S3). No significant differences were observed between the
groups, suggesting that the doses used did not cause toxicity to
other organs. Notably, no adverse reactions related to donafenib
use in rats, such as diarrhea or rash were observed. Serum analyses
of liver and kidney function were performed by measuring the levels
of ALT, AST, BUN and CRE in each group. AST and ALT levels
decreased in the HFD-DON and HFD-ATO + DON groups, but no
statistically significant differences were found. Furthermore, no
significant differences in ALT and AST levels were observed between
the HFD-ATO and HFD groups.
Discussion
MASLD is an increasingly significant public health
issue on a global scale, causing significant liver-related and
extrahepatic morbidity and mortality (28). One major feature of MASLD is the
presence of lipid metabolism disorder. In vitro, the HepG2
cell line was used to study fat metabolism. This cell line is
highly differentiated, has an intact biotransformation profile of
metabolic enzymes in the cells, does not require the incorporation
of an exogenous activation system and contains biotransformation
metabolic enzymes homologous to those of normal human liver
parenchymal cells (29,30). As a result, the HepG2 cell line is
widely utilized in MASLD research (18,19,31,32).
Similar to the methods used by Iturrospe et al (33), a drug concentration of
IC10 was used. The aim was not to inhibit cell
proliferation but to regulate cell metabolism, so low drug
concentrations were used. The present study did not focus on the
ameliorative effect of varying drug concentrations on MASLD.
Instead, it demonstrated, to the best of our knowledge, the first
time that donafenib modulated lipid metabolism, which was the
primary focus and innovation of the present research. To simulate
the dietary structure and pathogenesis in modern humans, an MASLD
model in rats using a HFD was successfully established (34).
The combination of donafenib and atorvastatin
improved HFD-induced MASLD in rats and FFA-stimulated cells.
Additionally, the present analysis of related proteins showed that
combination therapy activated the AMPK pathway, down-regulated the
expressions of SREBP-1, HMGCR and FAS, and up-regulated the
expressions of ACOX1 and CPT1C, thereby inhibiting the synthesis of
fatty acids and promoting β-oxidation of fatty acids. Serum and
liver lipid metabolism parameters were also measured, and it was
found that FFAs, triglycerides and cholesterol levels in the serum
and liver decreased after treatment. Serum VLDL-C levels remained
high, indicating that the extrahepatic transport of fatty acids
remained active. Notably, donafenib and atorvastatin reduced
oxidative stress in the liver tissue and decreased ROS levels.
Combined therapy can reduce fatty acid production, promote
β-oxidation and transport of fatty acids and reduce liver fat
content while improving oxidative stress and ultimately improving
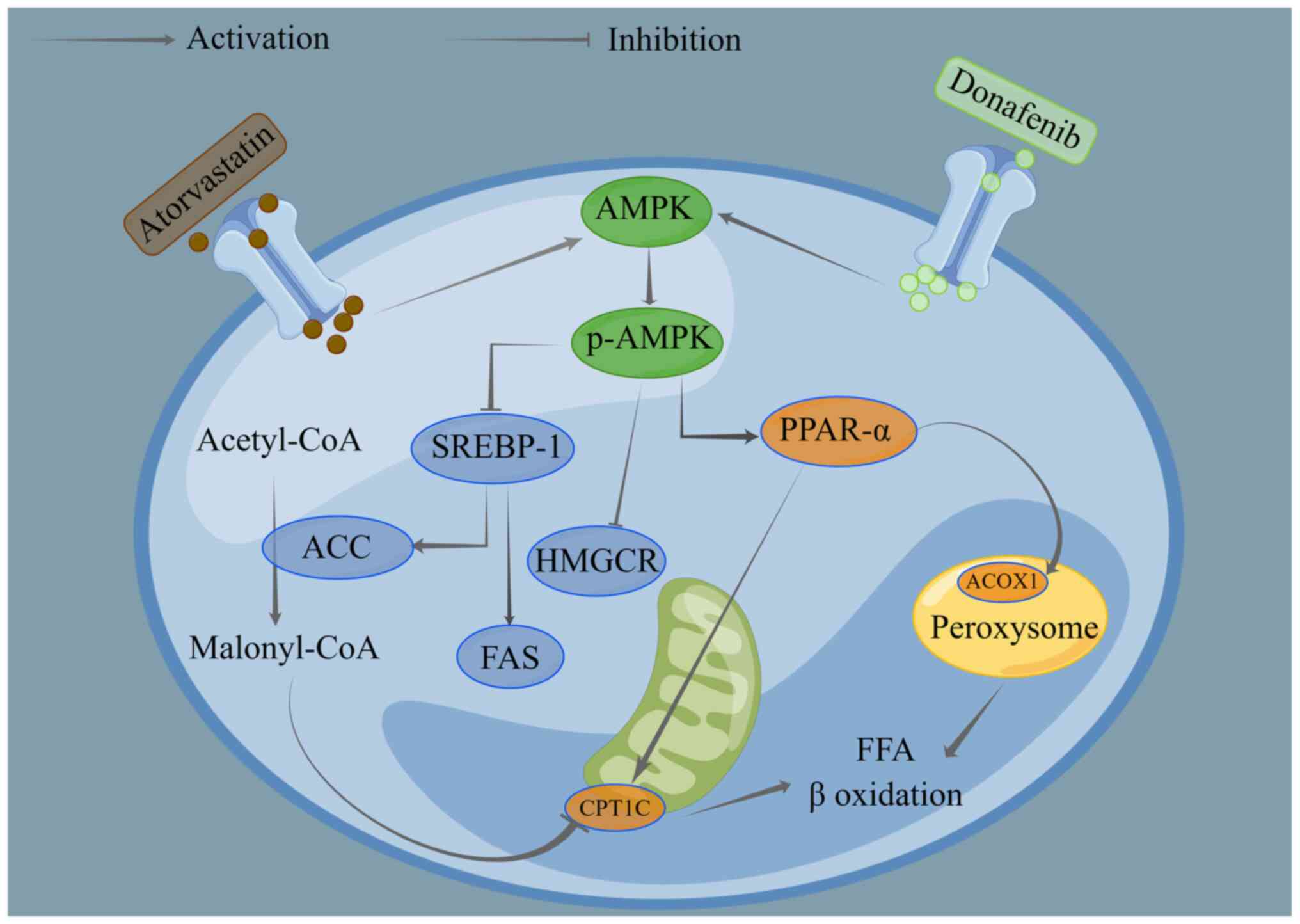
MASLD (Fig. 8). The combination of
the two drugs provided an improved therapeutic effect, and
therefore it was hypothesized that there was an additive effect of
the two drugs; however, it is unclear whether there was a
synergistic effect of the combined drugs. As to whether they have
synergistic effects in addition to additive effects, this will be
the focus of future studies.
Currently, there is no clear conclusion regarding
the pathogenic mechanism of MASLD; however, it is generally
hypothesized that the liver is overloaded with processing energy
metabolism substrates (sugars and fatty acids), leading to
excessive fatty acid deposition in the liver (6). Fatty acids and their lipotoxic
metabolites can trigger endoplasmic reticulum stress, oxidative
stress and the activation of inflammatory agents, leading to the
progression of NASH (10–12). There are four main reasons for
increased liver lipid levels (35). Firstly, the conversion of
carbohydrates, such as fructose into fatty acids by hepatocytes, is
one of the main sources of fatty acids in the liver. Secondly, FFAs
released by the breakdown of triglycerides in adipose tissue are
transported to the liver via the bloodstream, leading to an
increase in liver fatty acids. Thirdly, reduced β-oxidation of
fatty acids contributes to the accumulation of fatty acids in the
liver. Lastly, impaired synthesis of triglycerides from fatty acids
also leads to an increase in fatty acids within the liver (6,35).
Therefore, by acting on these processes, liver lipids can be
reduced, and MASLD can be improved. Recently, the therapeutic role
of the AMPK signaling pathway in metabolic diseases such as MASLD
has been widely investigated (36,37).
p-AMPK directly phosphorylates acetyl CoA carboxylase, thereby
inhibiting fatty acid synthesis (38). AMPK also reduces chronic
inflammation (39) and directly
phosphorylates caspase-6, thereby inhibiting lipotoxic
metabolite-induced hepatocyte apoptosis in NASH (40). A low dose of sorafenib, a
mitochondrial uncoupling agent, activates the AMPK pathway and
considerably improves MASLD in rats and monkeys (14). Moreover, statins have been widely
used in experimental studies of MASLD to regulate lipid metabolism
by acting on proteins such as HMG-CoA reductase (41). A recent study showed that diabetic
mice receiving long-term statins expressed more renal SREBP-1 and
showed increased renal fatty acid synthesis (42). The present study found that
donafenib and atorvastatin upregulated p-AMPK expression. This
effect disappeared when the AMPK inhibitor was added, while there
was no significant difference in fat accumulation and expressions
of lipid metabolism-related proteins in the FFA-ATO and FFA-DON
groups compared with the FFA group. This suggested that donafenib
and atorvastatin exert their effects on improving MASLD by
activating the AMPK pathway. Additionally, future endeavors involve
conducting more in-depth studies in the future to explore the
mechanisms by which donafenib and atorvastatin improve MASLD.
Given that both donafenib and atorvastatin affect
lipid metabolism through the AMPK pathway, the effects of donafenib
and atorvastatin in treatment of MASLD were explored and the
underlying mechanisms investigated. In the present study, donafenib
and atorvastatin inhibited the synthesis of fatty acids, increased
the β-oxidation of fatty acids and ultimately reduced the
accumulation of fatty acids and their metabolites such as
triglycerides and cholesterol in the liver, thereby improving the
fatty liver. Owing to the targeted therapeutic mechanisms of
donafenib, it was speculated that for patients with liver cancer
who progress from NAFLD, the etiology can be improved by treating
liver cancer. Further studies are needed to determine whether
donafenib produces improved therapeutic effects in these patients.
In contrast, lipid metabolism plays an important role in tumor
proliferation (43). In addition
to its anti-angiogenic effect, the anti-tumor effect of donafenib
may be manifested in the regulation of lipid metabolism. To the
best of our knowledge, the present study is the first study to
demonstrate that donafenib improves MASLD in rats.
Additionally, oxidative stress is indeed a key
pathogenic mechanism in MASLD, and studies have shown that
addressing oxidative stress can improve MASLD (44–46).
Furthermore, the American Association for the Study of Liver
Diseases) has recommended vitamin E supplementation, a common
antioxidant, for biopsy-proven NAFLD in nondiabetic patients to
improve NAFLD (47). In the
present study, it was found that donafenib and atorvastatin reduced
ROS levels in liver tissue, suggesting their ability to ameliorate
oxidative stress, leading to an improvement in MASLD. In the
future, more in-depth studies will be conducted on the mechanisms
by which donafenib and atorvastatin improve MASLD by ameliorating
oxidative stress. Meanwhile, the therapeutic effects of other
antioxidants on MASLD should be more thoroughly investigated.
Donafenib is clinically used to treat malignant
tumors and has not been applied to MASLD (15), therefore special attention was paid
to the dose safety of donafenib and atorvastatin. The dose of
donafenib used was 1 mg/kg/day, which is ~4.5 mg/day for a 60-kg
person according to the body surface area normalization method
(48), which is much lower than
the clinical dose (400 mg/day). Based on good therapeutic effects,
no similar clinical adverse reactions, such as diarrhea, skin
erythema and rash, were observed. There were no significant
differences in serum ALT, AST, BUN and CRE levels between the
treatment and the HFD groups, and no abnormalities were observed
upon histological examination of other organs. Therefore, both the
clinical safety data and our results suggest that the donafenib
dose used was safe. Additionally, statins may increase
transaminases levels (49,50), thereby failing to ameliorate the
decline in liver function caused by NASH. The present study
detected serum ALT and AST in rats and found that atorvastatin did
not lead to an abnormal elevation of transaminase levels, and no
significant difference was observed in transaminase levels between
the HFD-ATO and HFD groups.
The present study had several limitations. First,
only one cell model and one animal model were used and whether the
drug would have similar therapeutic effects in other models was not
explored. In addition, the drug dose used in rats was based on
previous literature, and a drug dose gradient was not set in the
present study. Finally, although the dose used was safe for rats,
it is important to note that the dose calculated by body surface
area normalization cannot be directly applied in clinical settings.
The efficacy and safety of donafenib combined with atorvastatin in
the treatment of patients with MASLD still requires further
study.
In conclusion, low-dose donafenib combined with
atorvastatin improved MASLD by regulating fatty acid metabolism and
reducing oxidative stress through activation of the AMPK signaling
pathway.
Supplementary Material
Supporting Data
Supporting Data
Acknowledgements
The authors would like to thank Professor Xiaoming
Liu (Union Hospital, Tongji Medical College, Huazhong University of
Science and Technology, Wuhan, China) for his help in animal
husbandry.
Funding
This work was funded by The National Natural Science Foundation
of China (grant no. 81873917).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author
Authors' contributions
YB, KC, JL, CZho and WY collected the data. YB, BX
and CZhe conceptualized and designed the study; BX and CZhe
acquired the funding; YB, KC, JL, YW, CW and SJ performed the
investigation; YB, KC, JL, YW, CW, SJ, CZho and WY performed the
study methodology; YB, KC, JL, BX and CZhe performed the project
administration; BX and CZhe supervised the project; YB, KC and JL
wrote the original draft of the manuscript; and BX and CZhe
reviewed and edited the manuscript. All authors read and approved
the final version of the manuscript. YB and CZhe confirm the
authenticity of all the raw data.
Ethics approval and consent to
participate
The study protocol conformed to the ARRIVE
guidelines and was approved by The Ethics Committee of Tongji
Medical College, Huazhong University of Science and Technology
(Wuhan, China; approval no. 3462).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
MASLD
|
metabolic dysfunction-associated
steatotic liver disease
|
|
SREBP-1
|
sterol regulatory element-binding
protein-1
|
|
CPT1C
|
carnitine palmitoyl-transferase 1C
|
|
HMGCR
|
3-hydroxy-3-methylglutaryl-CoA
reductase
|
|
ACOX1
|
acyl-CoA oxidase 1
|
|
ROS
|
reactive oxygen species
|
|
FFA
|
free fatty acid
|
|
LDL-C
|
low-density lipoprotein
cholesterol
|
|
HDL-C
|
high-density lipoprotein
cholesterol
|
|
VLDL-C
|
very low-density lipoprotein
cholesterol
|
|
FAS
|
fatty acid synthase
|
|
TG
|
total triglycerides
|
|
TC
|
total cholesterol
|
|
AST
|
aspartate aminotransferase
|
|
ALT
|
Alanine aminotransferase
|
|
CRE
|
creatinine
|
|
BUN
|
blood urea nitrogen
|
|
NAS
|
non-alcoholic fatty liver disease
activity score
|
References
|
1
|
Younossi Z, Anstee QM, Marietti M, Hardy
T, Henry L, Eslam M, George J and Bugianesi E: Global burden of
NAFLD and NASH: Trends, predictions, risk factors and prevention.
Nat Rev Gastroenterol Hepatol. 15:11–20. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Greenhill C: Phase IIa results for
potential NAFLD therapy. Nat Rev Endocrinol. 18:22022. View Article : Google Scholar
|
|
3
|
Rinella ME, Lazarus JV, Ratziu V, Francque
SM, Sanyal AJ, Kanwal F, Romero D, Abdelmalek MF, Anstee QM, Arab
JP, et al: A multi-society Delphi consensus statement on new fatty
liver disease nomenclature. Hepatology. 78:1966–1986. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Song SJ, Lai JCT, Wong GLH, Wong GLH, Wong
VWS and Yip TCF: Can we use old NAFLD data under the new MASLD
definition? J Hepatol. 2:S0168–S8278. 2023.
|
|
5
|
De A, Bhagat N, Mehta M, Taneja S and
Duseja A: Metabolic dysfunction-associated steatotic liver disease
(MASLD) definition is better than MAFLD criteria for lean patients
with non-alcoholic fatty liver disease (NAFLD). J Hepatol.
7:S0168–S0827. 2023.
|
|
6
|
Friedman SL, Neuschwander-Tetri BA,
Rinella M and Sanyal AJ: Mechanisms of NAFLD development and
therapeutic strategies. Nat Med. 24:908–922. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Estes C, Anstee QM, Arias-Loste MT, Bantel
H, Bellentani S, Caballeria J, Colombo M, Craxi A, Crespo J, Day
CP, et al: Modeling NAFLD disease burden in China, France, Germany,
Italy, Japan, Spain, United Kingdom, and United States for the
period 2016–2030. J Hepatol. 69:896–904. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Régnier M, Carbinatti T, Parlati L,
Benhamed F and Postic C: The role of ChREBP in carbohydrate sensing
and NAFLD development. Nat Rev Endocrinol. 19:336–349. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Loneker AE, Alisafaei F, Kant A, Li D,
Janmey PA, Shenoy VB and Wells RG: Lipid droplets are intracellular
mechanical stressors that impair hepatocyte function. Proc Natl
Acad Sci USA. 120:e22168111202023. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mazilescu LI, Selzner M and Selzner N:
Defatting strategies in the current era of liver steatosis. JHEP
Rep. 3:1002652021. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Parlati L, Régnier M, Guillou H and Postic
C: New targets for NAFLD. JHEP Rep. 3:1003462021. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Inci MK, Park SH, Helsley RN, Attia SL and
Softic S: Fructose impairs fat oxidation: Implications for the
mechanism of western diet-induced NAFLD. J Nutr Biochem.
114:1092242023. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yau T, Park JW, Finn RS, Cheng AL,
Mathurin P, Edeline J, Kudo M, Harding JJ, Merle P, Rosmorduc O, et
al: Nivolumab versus sorafenib in advanced hepatocellular carcinoma
(CheckMate 459): A randomised, multicentre, open-label, phase 3
trial. Lancet Oncol. 23:77–90. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jian C, Fu J, Cheng X, Shen LJ, Ji YX,
Wang X, Pan S, Tian H, Tian S, Liao R, et al: Low-Dose Sorafenib
acts as a mitochondrial uncoupler and Ameliorates Nonalcoholic
Steatohepatitis. Cell Metab. 31:12062020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Qin S, Bi F, Gu S, Bai Y, Chen Z, Wang Z,
Ying J, Lu Y, Meng Z, Pan H, et al: Donafenib Versus Sorafenib in
first-line treatment of unresectable or metastatic hepatocellular
carcinoma: A randomized, open-label, parallel-controlled phase
II–III trial. J Clin Oncol. 39:3002–3011. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dongiovanni P, Petta S, Mannisto V,
Mancina RM, Pipitone R, Karja V, Maggioni M, Kakela P, Wiklund O,
Mozzi E, et al: Statin use and non-alcoholic steatohepatitis in at
risk individuals. J Hepatol. 63:705–712. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dehnavi S, Kiani A, Sadeghi M, Biregani
AF, Banach M, Atkin SL, Jamialahmadi T and Sahebkar A: Targeting
AMPK by Statins: A potential therapeutic approach. Drugs.
81:923–933. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tanaka S, Hikita H, Tatsumi T, Sakamori R,
Nozaki Y, Sakane S, Shiode Y, Nakabori T, Saito Y, Hiramatsu N, et
al: Rubicon inhibits autophagy and accelerates hepatocyte apoptosis
and lipid accumulation in nonalcoholic fatty liver disease in mice.
Hepatology. 64:1994–2014. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang P, Ge Z, Wang H, Feng W, Sun X, Chu
X, Jiang C, Wang Y, Zhu D and Bi Y: Prolactin improves hepatic
steatosis via CD36 pathway. J Hepatol. 68:1247–1255. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jiang JJ, Zhang GF, Zheng JY, Sun JH and
Ding SB: Targeting mitochondrial ROS-mediated ferroptosis by
quercetin alleviates high-fat diet-induced hepatic lipotoxicity.
Front Pharmacol. 13:8765502022. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vucicevic L, Misirkic M, Janjetovic K,
Vilimanovich U, Sudar E, Isenovic E, Prica M, Harhaji-Trajkovic L,
Kravic-Stevovic T, Bumbasirevic V and Trajkovic V: Compound C
induces protective autophagy in cancer cells through AMPK
inhibition-independent blockade of Akt/mTOR pathway. Autophagy.
7:40–50. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wu Y, Yan B, Xu W, Guo L, Wang Z, Li G,
Hou N, Zhang J and Ling R: Compound C enhances the anticancer
effect of aspirin in HER-2-positive breast cancer by regulating
lipid metabolism in an AMPK-independent pathway. Int J Biol Sci.
16:583–597. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu B, Xu J, Lu L, Gao L, Zhu S, Sui Y,
Cao T and Yang T: Metformin induces pyroptosis in leptin
receptor-defective hepatocytes via overactivation of the AMPK axis.
Cell Death Dis. 14:822023. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bravo M, Raurell I, Barberá A, Hide D, Gil
M, Estrella F, Salcedo MT, Augustin S, Genescà J and Martell M:
Synergic effect of atorvastatin and ambrisentan on sinusoidal and
hemodynamic alterations in a rat model of NASH. Dis Model Mech.
14:dmm0488842021. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Matafome P, Louro T, Rodrigues L,
Crisóstomo J, Nunes E, Amaral C, Monteiro P, Cipriano A and Seiça
R: Metformin and atorvastatin combination further protect the liver
in type 2 diabetes with hyperlipidaemia. Diabetes Metab Res Rev.
27:54–62. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li HY, Huang SY, Zhou DD, Xiong RG, Luo M,
Saimaiti A, Han MK, Gan RY, Zhu HL and Li HB: Theabrownin inhibits
obesity and non-alcoholic fatty liver disease in mice via
serotonin-related signaling pathways and gut-liver axis. J Adv Res.
52:59–72. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kebbel A and Röcken C: Immunohistochemical
classification of amyloid in surgical pathology revisited. Am J
Surg Pathol. 30:673–683. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Devarbhavi H, Asrani SK, Arab JP, Nartey
YA, Pose E and Kamath PS: Global burden of liver disease: 2023
Update. J Hepatol. 79:516–537. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Amorim R, Simões ICM, Veloso C, Carvalho
A, Simões RF, Pereira FB, Thiel T, Normann A, Morais C, Jurado AS,
et al: Exploratory data analysis of cell and mitochondrial
high-fat, high-sugar toxicity on human HepG2 cells. Nutrients.
13:17232021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Green CJ, Johnson D, Amin HD, Sivathondan
P, Silva MA, Wang LM, Stevanato L, McNeil CA, Miljan EA, Sinden JD,
et al: Characterization of lipid metabolism in a novel immortalized
human hepatocyte cell line. Am J Physiol Endocrinol Metab.
309:E511–E522. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Besse-Patin A, Léveillé M, Oropeza D,
Nguyen BN, Prat A and Estall JL: Estrogen signals through
peroxisome proliferator-activated receptor-γ coactivator 1α to
reduce oxidative damage associated with diet-induced fatty liver
disease. Gastroenterology. 152:243–256. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dongiovanni P, Crudele A, Panera N, Romito
I, Meroni M, De Stefanis C, Palma A, Comparcola D, Fracanzani AL,
Miele L, et al: β-Klotho gene variation is associated with liver
damage in children with NAFLD. J Hepatol. 72:411–419. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Iturrospe E, Robeyns R, da Silva KM, van
de Lavoi M, Boeckmans J, Vanhaeck T, van Nuij ALN and Covaci A:
Metabolic signature of HepaRG cells exposed to ethanol and tumor
necrosis factor alpha to study alcoholic steatohepatitis by
LC-MS-based untargeted metabolomics. Arch Toxicol. 97:1335–1353.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kakimoto PA and Kowaltowski AJ: Effects of
high fat diets on rodent liver bioenergetics and oxidative
imbalance. Redox Biol. 8:216–225. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen H, Tan H, Wan J, Zeng Y, Wang J, Wang
H and Lu X: PPAR-γ signaling in nonalcoholic fatty liver disease:
Pathogenesis and therapeutic targets. Pharmacol Ther.
245:1083912023. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Fang C, Pan J, Qu N, Lei Y, Han J, Zhang J
and Han D: The AMPK pathway in fatty liver disease. Front Physiol.
13:9702922022. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Day EA, Ford RJ and Steinberg GR: AMPK as
a therapeutic target for treating metabolic diseases. Trends
Endocrinol Metab. 28:545–560. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wakil SJ and Abu-Elheiga LA: Fatty acid
metabolism: Target for metabolic syndrome. J Lipid Res. 50
(Suppl):S138–S143. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kelly B and O'Neill LAJ: Metabolic
reprogramming in macrophages and dendritic cells in innate
immunity. Cell Res. 25:771–784. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhao P and Saltiel AR: From overnutrition
to liver injury: AMP-activated protein kinase in nonalcoholic fatty
liver diseases. J Biol Chem. 295:12279–12289. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tziomalos K, Athyros VG, Paschos P and
Karagiannis A: Nonalcoholic fatty liver disease and statins.
Metabolism. 64:1215–1223. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Huang TS, Wu T, Wu YD, Li XH, Tan J, Shen
CH, Xiong SJ, Feng ZQ, Gao SF, Li H and Cai WB: Long-term statins
administration exacerbates diabetic nephropathy via ectopic fat
deposition in diabetic mice. Nat Commun. 14:3902023. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lally JSV, Ghoshal S, DePeralta DK, Moaven
O, Wei L, Masia R, Erstad DJ, Fujiwara N, Leong V, Houde VP, et al:
Inhibition of Acetyl-CoA carboxylase by phosphorylation or the
inhibitor ND-654 suppresses lipogenesis and hepatocellular
carcinoma. Cell Metab. 29:174–182. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lee SM, Koh DH, Jun DW, Roh YJ, Kang HT,
Oh JH and Kim HS: Auranofin attenuates hepatic steatosis and
fibrosis in nonalcoholic fatty liver disease via NRF2 and NF-κB
signaling pathways. Clin Mol Hepatol. 28:827–840. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yu H, Yan S, Jin M, Wei Y, Zhao L, Cheng
J, Ding L and Feng H: Aescin can alleviate NAFLD through Keap1-Nrf2
by activating antioxidant and autophagy. Phytomedicine.
113:1547462023. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chen X, Xue H, Fang W, Chen K, Chen S,
Yang W, Shen T, Chen X, Zhang P and Ling W: Adropin protects
against liver injury in nonalcoholic steatohepatitis via the Nrf2
mediated antioxidant capacity. Redox Biol. 21:1010682019.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chalasani N, Younossi Z, Lavine JE,
Charlton M, Cusi K, Rinella M, Harrison SA, Brunt EM and Sanyal AJ:
The diagnosis and management of nonalcoholic fatty liver disease:
Practice guidance from the american association for the study of
liver diseases. Hepatology. 67:328–357. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Reagan-Shaw S, Nihal M and Ahmad N: Dose
translation from animal to human studies revisited. FASEB J.
22:659–661. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Davidson MH and Robinson JG: Safety of
aggressive lipid management. J Am Coll Cardiol. 49:1753–1762. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Stone NJ, Robinson JG, Lichtenstein AH,
Merz CN, Blum CB, Eckel RH, Goldberg AC, Gordon D, Levy D,
Lloyd-Jones DM, et al: 2013 ACC/AHA guideline on the treatment of
blood cholesterol to reduce atherosclerotic cardiovascular risk in
adults: A report of the American college of Cardiology/American
heart association task force on practice guidelines. Circulation.
129:S1–45. 2014. View Article : Google Scholar : PubMed/NCBI
|