Introduction
Heat stroke (HS) is a life-threatening condition
clinically characterized by extreme hyperthermia (>40.5°C),
central nervous system dysfunction, systematic inflammatory
response syndrome and multiorgan dysfunction syndrome (1). According to its cause and degree of
susceptibility within the population, HS can be categorized as
either classic or exertional, although both are associated with
high morbidity and mortality rates, and warrant attention and
research (2). It has been reported
that episodes of extreme heat waves will increase in the future,
meaning that the incidence of HS is likely to increase further
(3). Furthermore, a previous study
from Kyoto indicated that the estimated number of patients with HS
transported by ambulance is predicted to be >3× greater among
people aged 0–64 years and 6× greater among people aged ≥65 years
(4). Therefore, it is necessary to
explore the medical intervention methods against HS.
The endothelium, a barrier formed by squamous
endothelial cells, is a highly dynamic layer that mainly lines the
blood vessels and is responsible for maintaining the stability of
microvascular permeability (5).
Accumulating evidence has suggested that the disruption of the
normal vascular endothelial barrier plays a key role during the
pathophysiological processes of HS (1,6). The
vascular endothelium exhibits discontinuity in HS, which manifests
as increased intercellular gaps and extensive endothelial cell
apoptosis, ultimately leading to compromised structural-barrier
integrity (6). The increase in
vascular endothelial permeability is the main mechanism underlying
capillary leakage, which results in severe coagulation problems
(7). Protection of the vascular
endothelial barrier has resulted in beneficial effects on HS such
as inhibition of coagulopathy and reduction of the risk of vascular
leakage, although the specific mechanisms underlying this have not
been revealed (7,8). The present study conducted
genome-wide CRISPR-Cas9 screening research (Fig. S1) to explore the specific genes
responsible for resisting HS and enhanced heat tolerance in the
early stage using human umbilical vein endothelial cells (HUVECs)
(9,10).
DNAJA1 is a protein-coding gene encoding DnaJ heat
shock protein (HSP)40 family member A1, which is often considered
to act as a HSP70 co-chaperone, and facilitate protein folding and
trafficking and prevent aggregation (11,12).
Traditionally, HSPs serve a crucial role in the cellular response
to heat stress by maintaining protein homeostasis and preventing
protein denaturation (13).
However, few studies have investigated the effects of DNAJA1-KO on
HS, and its protective effects on HS go against the conventional
theories surrounding HSPs. Of note, a previous study revealed that
HSP90 inhibitors prevent lipopolysaccharide (LPS)-induced
endothelial barrier dysfunction by suppressing RhoA signaling
(14). As HUVECs were used to
identify heat tolerance-related genes, the present study aimed to
investigate the protective effects and mechanisms of DNAJA1-KO on
HS-induced endothelial barrier disruption. Specifically, it was
demonstrated that DNAJA1 may be a novel potential therapeutic
target for regulating the damage of vascular endothelial barrier
during HS.
Materials and methods
Cell culture and HS protocol
Immortalized HUVECs (cat. no. 1101HUM-PUMC000437;
Shanghai Institute of Biochemistry and Cell Biology, Chinese
Academy of Sciences) were cultured in DMEM (Wuhan Servicebio
Technology Co., Ltd.) supplemented with 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin/streptomycin at 37°C in 5% CO2. The control
cells were incubated at 37°C, while HS was induced in the
experimental cells via incubation at 43°C for 4 h, before returning
to 37°C for further research (12). The cell culture was refreshed every
2 days to remove non-adherent cells. EDTA-trypsin (Gibco; Thermo
Fisher Scientific, Inc.) was used to passage the cells when the
cells reached 80% confluence. The HUVECs were divided into Control
(CON) and DNAJA1-KO groups according to whether or not they were
subjected to CRISPR/Cas9 gene KO; these two groups were further
divided into CON, CON + HS, DNAJA1-KO and KO + HS groups according
to whether they underwent HS treatment.
Construction of a stable cell
line
CRISPR/Cas9 technology was used to construct a gene
KO cell line. The single guide RNA (sgRNA; forward,
5′-ACCGCAGCTCGTTGAAGCACTGTG-3′) targeting the transcription start
site of the DNAJA1 gene exon 1 (Gene ID: NM_001314039;
5′-CAGCTCGTTGAAGCACTGTG-3′) and the control sgRNA
(5′-ACGGAGGCTAAGCGTCGCAA-3′) were designed using VectorNTI
(version, 11.5.1; Invitrogen; Thermo Fisher Scientific, Inc.) and
induced into the pLenti-U6-spgRNA
v2.0-CMV-Puro-P2A-3×FLAG-spCas9-WPRE plasmid vector [OBiO
Technology (Shanghai) Corp., Ltd.]. Subsequently, the two plasmids
were transduced into HUVECs according to the recommended protocol
of OBiO. The sgRNA for knockout of DNAJA1 was used in KO groups and
the control sg RNA was used in CON groups. The efficiency of DNAJA1
knockout was confirmed by western blotting.
Cell viability assays
To measure cell viability, HUVECs were analyzed
using a Cell Counting Kit-8 (CCK-8; Dojindo Laboratories, Inc.)
assay and a lactate dehydrogenase (LDH) Cytotoxicity Assay Kit-WST
(LDH; Dojindo Laboratories, Inc.). Cells were seeded in a 96-well
plate at a density of 1×105/ml and incubated at 37°C
overnight. Subsequently, cells were treated according to the group,
mixed with CCK-8 solution and incubated for 2 h. According to the
manufacturer's instructions, the absorbances of CCK-8 and LDH were
measured using a microplate reader (ND-171; Omega Bio-Tek, Inc.) at
wavelengths of 450 and 490 nm, respectively.
Flow cytometric analysis
Apoptosis was assessed using an Annexin V-FITC
Apoptosis Detection Kit [cat. no. AT101; Multi Sciences (Lianke)
Biotech Co., Ltd.] according to the manufacturer's instructions.
Briefly, HUVECs (~1×106) were collected, washed with
pre-cooled phosphate-buffered saline (PBS), and resuspended in
binding buffer containing 5 µl Annexin V-FITC and 10 µl PI. The
samples were then incubated at room temperature for 5–15 min in the
dark, before running on a flow cytometer (FACSCalibur; BD
Biosciences). The results were analyzed using Cell Quest Pro
software (version 6.0; BD Biosciences). The apoptosis rate was
calculated as: Apoptosis rate (%)=(apoptotic cell number/total cell
number) ×100.
Endothelial monolayer permeability
assays
Transendothelial electrical resistance (TEER) and
FITC-Dextran permeability assays were conducted to evaluate the
barrier function of HUVECs. Briefly, cells were seeded on Transwell
filters (0.4-µm-pore, 6.5-mm-diameter, 24-well plates; cat. no.
356008; Corning, Inc.) coated with fibronectin which had been
incubated for 24 h at 37°C in 5% CO2 at a density of
1×106 cells/ml and were incubated at 37°C in 5%
CO2. The cells seeded on Transwell filters were also
cultured in DMEM (Wuhan Servicebio Technology Co., Ltd.) with 10%
FBS (Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin/streptomycin. Each Transwell insert was placed in a well
with 200 µl medium (DMEM with 10% FBS) in the upper chamber and 1
ml medium (DMEM with 10% FBS) in the lower chamber. After reaching
100% confluence, an EVOM2 Epithelial Voltohmmeter (World Precision
Instruments) was used to detect the TEER values of each group. The
TEER (in Ω/cm2) was calculated using the following
equation: (resistance of sample well-resistance of corresponding
blank well) ×0.33, where 0.33 is the area (in cm2) of
the insert membrane. The TEER was monitored daily and was
considered to form the integrated endothelial monolayer when the
values remained stable after increasing. As aforementioned, the
integrated endothelial monolayer samples were cultured in an
incubator at 43°C for 4 h to simulate HS injury, while the
corresponding control samples were cultured in a normal incubator
at 37°C. After HS treatment, the TEER was detected in each group.
The measurements were recorded in triplicate and are reported as
the TEER values (mean + standard error) comparing four groups.
Furthermore, based on the integrated endothelial monolayer model
using Transwell filters, the cell medium was washed and refreshed
with HEPES medium (Beyotime Institute of Biotechnology), before
adding FITC-Dextran (Invitrogen; Thermo Fisher Scientific, Inc.) to
the upper chamber. The plates were incubated at 37°C in 5%
CO2 for 4 h and the samples were then aspirated from the
bottom chamber to a solid black plate and fluorescence intensity
(excitation/emission wavelength, 490/520 nm) was read using a
fluorometer (ND-171; Omega Bio-Tek, Inc.).
Western blot analysis
After corresponding treatments, HUVEC monolayers
grown in 6-well plates (Corning, Inc.) were lysed on ice using RIPA
lysis buffer (Beyotime Institute of Biotechnology) with a protease
inhibitor cocktail and phosphatase inhibitor cocktail (Roche
Diagnostics GmbH), before centrifuging cell lysates at 15,000 × g
at 4°C for 10 min. Subsequently, the protein concentration in the
supernatant was measured using the bicinchoninic acid (BCA) protein
assay kit (Beyotime Institute of Biotechnology), before detecting
the protein expression by western blotting. Protein samples were
mixed with loading buffer, denatured and separated on 4–12 and
4–20% SurePAGE™ gels (GenScript) with 30 µg protein per lane.
Proteins were transferred to PVDF membranes using an eBlot™ L1 Fast
Wet Transfer System (GenScript). The blotted wet membrane was
blocked for 2 h at room temperature with blocking buffer
(QuickBlock Western Blocking Buffer; cat. no. P0252; Beyotime
Institute of Biotechnology). Next, the membranes were incubated
overnight with primary antibodies at 4°C, followed by incubation
with secondary antibodies for 2 h at 37°C. The Omni-ECLTM Femto
Light Chemiluminescence Kit (cat. no. SQ201; Epizyme, Inc.; Ipsen
Biopharmaceuticals, Inc.) was used to detect luminescence signals
on the membrane, and the Amersham Imager 600 (Cytiva) was used to
image the blots. The protein expression was quantified using ImageJ
software (version 1.52e; National Institutes of Health). The
following primary antibodies were used overnight at 4°C: DNAJA1
(Proteintech Group, Inc.; cat. no. 11713-1-AP; 1:1,000; rabbit),
GAPDH (Beyotime Institute of Biotechnology; cat. no. AF0006;
1:1,000; mouse), cleaved-caspase-3 p17 (Cell Signaling Technology,
Inc.; cat. no. 9661; 1:1,000; rabbit), caspase-3 (ABclonal Biotech
Co., Ltd.; cat. no. A22869; 1:500; rabbit), Bcl-2 (Proteintech
Group, Inc.; cat. no. 68103-1-Ig; 1:8,000; mouse), Bax (Proteintech
Group, Inc.; cat. no. 60267-1-Ig; 1:8,000; mouse), zonula
occludens-1 (ZO-1; Cell Signaling Technology, Inc.; cat. no. 13663;
1:1,000, rabbit), junctional adhesion molecule A (JAM-A; Cell
Signaling Technology, Inc.; cat. no. 82196; 1:1,000; rabbit),
claudin-5 (Abcam; cat. no. ab131259; 1:1,000; rabbit), occludin
(Invitrogen; Thermo Fisher Scientific, Inc.; cat. no. 33-1500;
1:1,000; mouse), phosphorylated (p)-myosin light-chain kinase
(MLCK; Abcam; cat. no. ab200809; 1:1,000; rabbit), MLCK (Santa Cruz
Biotechnology, Inc.; cat. no. sc-365352; 1:1,000; mouse), p-MLC
(Cell Signaling Technology, Inc.; cat. no. 3675S; 1:1,000; mouse)
and MLC (Cell Signaling Technology, Inc.; cat. no. 3672S; 1:1,000;
rabbit). The following secondary antibodies were used: 1:8,000,
Goat Anti-Mouse IgG (H+L) HRP (1:8,000, cat. no. AB0102, Shanghai
Abways Biotechnology Co., Ltd.); and Goat Anti-Rabbit IgG (H+L) HRP
(1:8,000, cat. no. AB0101, Shanghai Abways Biotechnology Co.,
Ltd.).
Immunofluorescence analysis
Cells were washed three times with PBS and fixed in
4% paraformaldehyde and 2% Triton X-100 at room temperature for 10
min, before incubating with ZO-1 (Cell Signaling Technology, Inc.;
cat. no. 13663; 1:100, rabbit) and JAM-A (Cell Signaling
Technology, Inc.; cat. no. 82196; 1:200, rabbit) antibodies
overnight at 4°C. Following incubation, the samples were incubated
with Alexa Fluor 488-labeled Goat Anti-Rabbit IgG(H+L) secondary
antibodies (1:500, cat. no. A0423, Beyotime Institute of
Biotechnology) and TRITC phalloidin for labeling F-actin (Beijing
Solarbio Science & Technology Co., Ltd.) for 2 h at room
temperature. After washing, the cells were stained with DAPI for 5
min at room temperature (cat. no. C1006, Beyotime Institute of
Biotechnology) and visualized using an inverted fluorescence
microscope (IX71; Olympus Corporation).
Proteomic analysis
Cell samples were disrupted in lysis buffer (8M
urea, 1% SDS) and the concentrations were determined using a BCA
kit according to the manufacturer's protocol. Protein digestion was
performed according to the standard procedure. Subsequently, the
peptides were subjected to vacuum drying, followed by resuspension
with a solution containing 2% acetonitrile and 0.1% trifluoroacetic
acid. The samples were then desalted using Sep-Pak and underwent
vacuum drying. The peptide concentrations were determined using a
peptide quantification kit (cat. no. 23275; Thermo Fisher
Scientific, Inc.). Loading buffer was added to each tube to prepare
samples for mass spectrometry analysis and the concentration of
each sample was 0.5 µg/µl. To enhance the proteomic depth, the
samples were fractionated through high pH reversed-phase
separation. The peptides were reconstituted using a loading buffer
consisting of ammonium hydroxide solution containing 2%
acetonitrile and adjusted to pH 10. Separation of the peptides was
performed by Shanghai Majorbio Bio-pharm Technology Co., Ltd.,
using high pH reversed-phase liquid chromatography (LC) (RPLC;
Acquity Ultra Performance LC; Waters Corporation). The gradient
elution was performed on a high pH RPLC column (Acquity Ultra
Performance Liquid Chromatography Ethylene Bridged Hybrid C18
Column, 1.7 µm, 2.1×150 mm, Waters Corporation) at 200 µl/min with
the gradient increased for 66 min (phase B, ammonium hydroxide
solution containing 80% acetonitrile, pH 10; phase A: ammonium
hydroxide solution containing 2% acetonitrile, pH 10). A total of
10 fractions were obtained per sample by pooling 20 fractions from
each. LC-tandem mass spectrometry (MS/MS) (EASY-nLC 1200, Thermo
Fisher Scientific, Inc.) analysis was performed by Shanghai
Majorbio Bio-pharm Technology Co., Ltd. The sample was loaded onto
a C18-reversed phase column (75 µm × 25 cm; Thermo Fisher
Scientific, Inc.) in buffer A (2% acetonitrile and 0.1% formic
acid) and separated with a linear gradient of buffer B (80%
acetonitrile and 0.1% formic acid) at a flow rate of 300 nl/min. An
electrospray voltage of 1.8 kV against the inlet of the mass
spectrometer was used. The nitrogen temperature was 22°C (room
temperature in the mass spectrometry laboratory) and the positive
ionization mode was used. A Q Exactive mass spectrometer (Thermo
Fisher Scientific, Inc) was operated in the data-dependent mode, to
switch automatically between MS and MS/MS acquisition. Survey
full-scan MS spectra (m/z 350-1,300) were acquired with a mass
resolution of 70K, followed by twenty sequential high energy
collisional dissociation MS/MS scans with a resolution of 17.5K. In
all cases, one microscan was recorded using dynamic exclusion of 18
sec. MS/MS spectra were searched using Proteome Discoverer Software
2.2 software (Thermo Fisher Scientific, Inc.) against the Homo
sapiens database from UniProt website (https://www.uniprot.org/proteomes/UP000005640).
The identification of parent proteins was accomplished using the
peptide mass with the highest score, which corresponded to the
predicted value in the database. The parameters employed for
protein searching were established as follows: Tryptic digestion
with a maximum of two missed cleavages, fixed modification of
carbamidomethylation of cysteines, and variable modification of
oxidation of methionines and protein N-terminal acetylation.
Validation of peptide spectral matches was executed through the
assessment of q-values at a 1% false discovery rate. In the present
study, a total of 4,394 proteins expressed were identified as
belonging to the proteome of HUVECs. The thresholds of the fold
change (>1.2 or <0.83) and P<0.05 were used to identify
differentially expressed proteins (DEPs). Annotation of all
identified proteins was performed using Gene Ontology (GO) term
(http://www.blast2go.com/b2ghome;
http://geneontology.org/) and the Kyoto
Encyclopedia of Genes and Genomes (KEGG) pathway (http://www.genome.jp/kegg) enrichment analyses. The
data were analyzed using the free online Majorbio Choud platform
(cloud.majorbio.com) (15). Raw
proteomics data are available on ProteomeXchange (https://www.proteomexchange.org/) with accession
number PXD043512.
Animals
HS protocols and sample collection
Male C57BL/6J mice (age, 7–8 weeks; weight, 18–22 g)
were purchased from Shanghai SIPPR-Bk Laboratory Animal Co., Ltd.
Standard pellet rat chow and distilled tap water were provided
ad libitum. A total of 12 mice (CON (Wild type mice), n=3;
KO (DNAJA1−/− mice), n=3; CON (Wild type mice)CON + HS,
n=3; and KO + HS (DNAJA1−/− mice), n=3) were used for
the experiments. All mice were raised in the Specific Pathogen Free
Animal Experiment Center of the Naval Medical University (Shanghai,
China) and were acclimated to the animal facility conditions
(22±1°C; humidity, 50±5%; 12-h light/dark cycle) for 1 week prior
to the experiment. All experimental procedures were approved by The
Institutional Animal Ethics Committee of the Naval Medical
University (approval no. NMUMREC-2021-002) according to the Guide
for the Care and Use of Laboratory Animals of the National
Institutes of Health (16). All
mice in this experiment were anesthetized with isoflurane
(induction, 3–5%; maintenance, 1–2%) using a gas anesthesia machine
(KW-MZJ; Nanjing Calvin Biotechnology Co., Ltd.). Humane endpoints
of mice unable to live normally due to severe pain and discomfort
caused by the experimental procedures were included in the
experimental design, however no mice reached these endpoints in the
present study. The animals were placed in the chamber and perfused
with CO2 at a rate of 30% volume/min until the animals
were not moving, not breathing and their pupils were dilated.
Subsequently, the CO2 was turned off, and the animals
were observed for 5 min to confirm that the animals had been
sacrificed. DNAJA1-KO mice with C57BL/6 genetic background were
generated and purchased by Cyagen Biosciences, Inc. All mice were
genotyped by PCR analysis. In brief, the PCR genotyping
identification protocol for gene knockout mice involved tail
sampling (a 2 mm section of the end of the mouse tail was and
stored at −80°C), DNA extraction was performed using a Servicebio
Genomic DNA Kit (cat. no. G3633, Wuhan Servicebio Technology Co.,
Ltd.), PCR amplification was performed using 2× Fast Pfus PCR
Master Mix (cat. no. G3305, Wuhan Servicebio Technology Co., Ltd.),
gel electrophoresis was performed using a 2% agarose gel and
visualized using. SerRed nucleic acid dye (G3606, Wuhan Servicebio
Technology Co., Ltd.) and a Gel Imaging System (WD-9413B; Beijing
Liuyi Biotechnology Co., Ltd.). Briefly, PCR amplification was
performed as follows, pre-denaturation at 98°C for 2 min, then 30
cycles of denaturation at 98°C for 20 sec, annealing at 55°C for 20
sec and extension at 72°C for 10 sec before a final extension at
72°C for 5 min. Two primer sequences were designed to identify the
knockout mice. The primer sequences targeting DNAJA1−/−
mice genotyping (designed by GemPharmatech Co., Ltd.) were as
follows: Primer 1 forward (F), 5′-CGGATATTCAGAAAGGATTGGGAC-3′ and
reverse (R), 5′-ACCTTGAACTCACTGTGAAGCCAA-3′ (product for wild type,
7354 bp; product for KO, 325 bp). Primer 2 F,
5′-TCAACTTTCTGAGTACCAGGTCC-3′ and R, 5′-CAAAGCCTCATCCCTGAAGAGA-3′
(product for wild type, 349 bp; product for KO: 0 bp). The banding
patterns expected were as follows: Wild type with Primer 1, a
single wild type band; wild type with Primer 2, a single wild type
band; heterozygote with Primer 1, a wild type band and a KO band;
heterozygote with Primer 2, a wild type band; homozygote
(DNAJA1−/−): with Primer 1, a single KO band; and
homozygote (DNAJA1−/−) with Primer 2 reaction, no
product.
The improved HS protocol was based on our previously
reported study (17). Before HS
induction, a temperature-monitoring capsule (SV223 capsule
thermometer; Shenzhen Flamingo Technology Co., Ltd.) with real-time
temperature monitoring function was implanted in the experimental
mice to obtain the core body temperature (Tc) at 5-min intervals.
An artificial climate chamber (LTH-575N-01; Shanghai Drawell
Instrument Co., Ltd.), where ambient temperature was controlled at
40±1°C and relative humidity at 60±5% to create an environment with
high temperature and humidity, was used to induce HS onset. The
timepoint of stable hyperthermia occurrence (Tc >42.7°C) was
taken as the primary criterion of HS onset in mice. The use of
humane endpoints in animal research was mainly based on the
criteria for HS by monitoring the core body temperature using
temperature-monitoring capsules mentioned above (8). If the mice met the criteria of
hyperthermia occurrence (Tc >42.7°C), they were sampled and
euthanized 3 h after the onset of HS. Control mice not subjected to
heat stress were sampled and euthanized at the same time as
experimental mice (3 h after the onset of HS).
Blood sample examination and
histological examination
Blood samples were collected from the abdominal
aorta and separated by centrifugation at 1,000 × g for 10 min at
4°C, then sub-packed and stored at −80°C for later examination.
Serum biochemical indices of alanine aminotransferase (ALT),
aspartate aminotransferase (AST), blood urea (BU), creatine kinase
(CK) and creatinine (CREA) were examined using an automatic
biochemical analyzer (HITACHI 7080 automated analyzer; Hitachi,
Ltd.). Serum von Willebrand factor (VWF, cat. no. D721155, Sangon
Biotech Co., Ltd.), thrombomodulin (TM, cat. no. D721144, Sangon
Biotech Co., Ltd.) and endothelin-1 (ET-1, cat. no. D721180, Sangon
Biotech Co., Ltd.) were respectively detected using corresponding
ELISA kits according to the manufacturer's instructions. Lung
tissue samples were treated according to the protocol described in
our previous study (17). Briefly,
the organ samples were fixed in 4% paraformaldehyde at room
temperature for >24 h, dehydrated, embedded, sectioned to a
thickness of 4–6 µm using a rotary microtome and stained with
hematoxylin and eosin (H&E) for 3–5 min and 15 sec at room
temperature, respectively. All the H&E-stained slides were
examined using a light microscope (Leica DM2000; Leica Microsystems
GmbH) and scanned using a Panoramic MIDI Slide scanner (3DHISTECH,
Ltd.). Paraffin-embedded sections of the lung samples were also
used for immunofluorescence staining. Sections were incubated with
xylene at room temperature for 15 min, twice, and then rehydrated
in a descending alcohol series. The sections were then immersed in
sodium citrate antigen retrieval solution (pH 6.0) and heated to
99°C in the pressure cooker and held for 3 min before air cooling.
Sections were incubated with 3% bovine serum albumin for 30 min to
block non-specific binding at room temperature. Sections were
incubated with primary antibodies against VE-cadherin (1:500; cat.
no. AF8319; rabbit; Beyotime Institute of Biotechnology) overnight
at 4°C. Sections were rinsed with water and then incubated with
Cy3-conjugated Goat Anti-Rabbit IgG secondary antibodies (1:500;
cat. no. GB21301500; rabbit; Wuhan Servicebio Technology Co., Ltd.)
at room temperature for 50 min in dark condition. Sections were
washed with PBS and then counterstained with DAPI solution at room
temperature for 10 min, in the dark. Spontaneous fluorescence was
quenched by incubation with Antifluorescence Quenching Mounting
Media (G1401, Wuhan Servicebio Technology Co., Ltd.) for 5 min at
room temperature. Sections were imaged using a Nikon Eclipse TI-SR
fluorescent microscope (Nikon Corporation). CY3 glows red by
excitation wavelength 510–560 nm and emission wavelength 590 nm.
DAPI glows blue by UV excitation wavelength 330–380 nm and emission
wavelength 420 nm. The image acquisition system (Nikon DS-U3; Nikon
Corporation) and microscope settings were maintained throughout the
process.
Statistical analysis
Data are presented as the mean ± standard error of
mean. The sample size for the cell viability and endothelial
monolayer permeability assays was six replicates per group. The
sample size for the flow cytometric analysis, western blot
analysis, proteomic analysis and animal experiments was three
replicates per group. Unpaired Student's t-test was used to compare
two groups. One-way ANOVA with Bonferroni's post-hoc test was used
for comparisons among multiple groups. The statistical values were
calculated using GraphPad Prism (version 9.4.0, GraphPad Software;
Dotmatics) with P<0.05 considered to indicate a statistically
significant difference.
Results
DNAJA1-KO improves the thermal
tolerance of HUVECs following HS
The stable DNAJA1-KO cell line was constructed
successfully, according to the results of western blotting
(Fig. 1A). The CCK-8 assay
demonstrated that DNAJA1-KO had no significant effect on cell
viability, whereas it significantly improved viability compared
with the CON + HS cells incubated at 43°C for 4 h (Fig. 1B). A previous study reported that
HS exerts a cytotoxic effect on HUVECs (18,19).
DNAJA1-KO cells could markedly withstand HS-induced cytotoxic
effects, as indicated by the concentration of LDH detected by the
LDH release assay (Fig. 1C).
Furthermore, HS-induced endothelial apoptosis has been widely
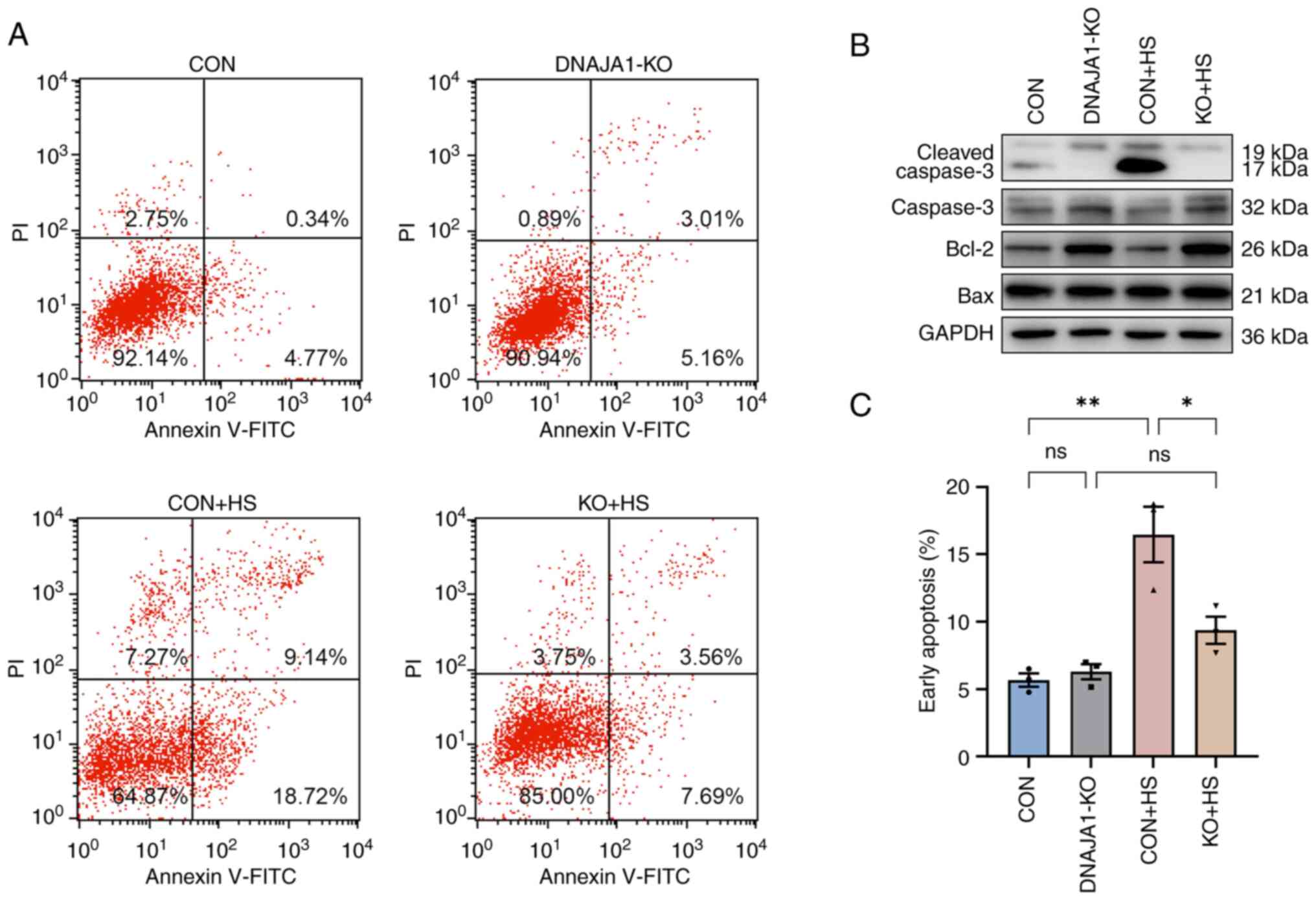
confirmed in the early stages of experimental heat stress (20). Next, the early apoptosis rate of
HS-treated cells was determined by flow cytometry. The results
demonstrated that the apoptosis rate of HS-treated CON cells was
significantly increased compared with that of untreated CON cells,
whereas only a few DNAJA1-KO cells were apoptotic (Fig. 2A and C). In parallel, western blot
analysis confirmed that DNAJA1-KO could alleviate HS-induced
apoptosis, as demonstrated by the apparent reduction of cleaved
caspase-3 by KO in HS cells (Fig.
2B). Notably, the classic apoptosis-inducing protein Bax showed
no difference in expression, whereas the classic anti-apoptotic
protein Bcl-2 was upregulated in DNAJA1-KO cells (21). Taken together, these data
demonstrated that DNAJA1-KO significantly improved the thermal
tolerance of HUVECs, as indicated by the results of cell viability
and apoptosis assays.
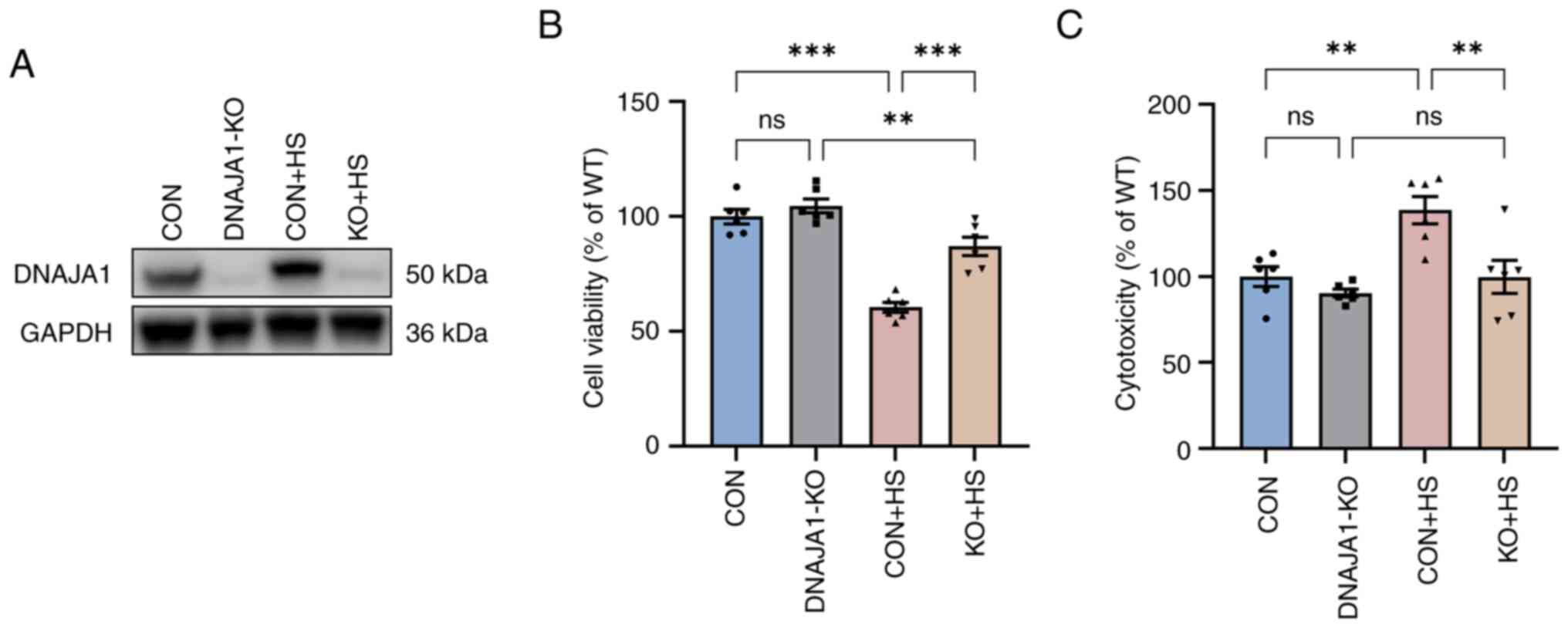 | Figure 1.DNAJA1-KO increases cell viability
and reduces cytotoxicity in HUVECs. Cell Counting Kit-8 and LDH
release assays were conducted immediately after incubation of
HUVECs for 4 h at 43°C. (A) Western blotting was used to evaluate
the efficiency of DNAJA1-KO, with GAPDH as the loading control. (B)
DNAJA1-KO significantly alleviated the HS-induced decrease in cell
viability. (C) DNAJA1-KO significantly mitigated HS-induced cell
damage, as indicated by lactate dehydrogenase release.
Representative values of cell viability and cytotoxicity, presented
as the mean percentage of the control ± SEM, are shown (n=6).
**P<0.01, ***P<0.001. ns, not significant; KO, knockout; CON,
control; HS, heat stroke; HUVECs, human umbilical vein endothelial
cells. |
DNAJA1-KO alleviates HS-induced
endothelial barrier dysfunction
After the cells were seeded on Transwell filters,
three wells were sampled from CON cell-derived monolayers and
DNAJA1-KO cell-derived monolayers to monitor the integrity of the
cells. Once the TEER values became stable, endothelial monolayer
permeability assays were conducted to determine the effects of
DNAJA1-KO. Based on the monitored TEER, the endothelial monolayer
met the experimental requirements and was generally >100
Ω/cm2 (Fig. 3A). The
integrated endothelial monolayer was incubated at 43°C for 4 h to
simulate HS injury, while the corresponding control filters were
cultured in a normal incubator. The results demonstrated that TEER
of the CON + HS group decreased, while more FITC-Dextran penetrated
the damaged monolayer to the bottom chamber of the Transwell. By
contrast, the KO + HS group maintained partial integrity of the
endothelial monolayer (Fig. 3B and
C). As endothelial barrier function could be protected by
DNAJA1-KO, investigation into the condition of the structure that
maintained the barrier was prompted. Numerous studies have proposed
that the endothelial barrier shares a number of features with tight
junctions (TJs) in epithelial cells (22,23).
Therefore, the expression of TJ proteins in each group was assessed
by western blotting to estimate the key structure of the barrier.
Under heat stress for 4 h, the expression of TJ proteins, including
ZO-1, occludin, JAM-A and claudin-5, decreased significantly in CON
+ HS cells compared with that in the CON cells (Fig. 3D). The expression of these
representative TJ proteins was normalized to GAPDH and statistical
analysis was performed. Although the expression of occludin, JAM-A
and claudin-5 in KO + HS cells was enhanced compared with that in
CON + HS cells, these differences were not statistically
significant. Of note, the expression of ZO-1 was effectively
upregulated in KO + HS with statistical significance, which
strongly demonstrated that the endothelial barrier of DNAJA1-KO
cells showed greater heat resistance ability against HS.
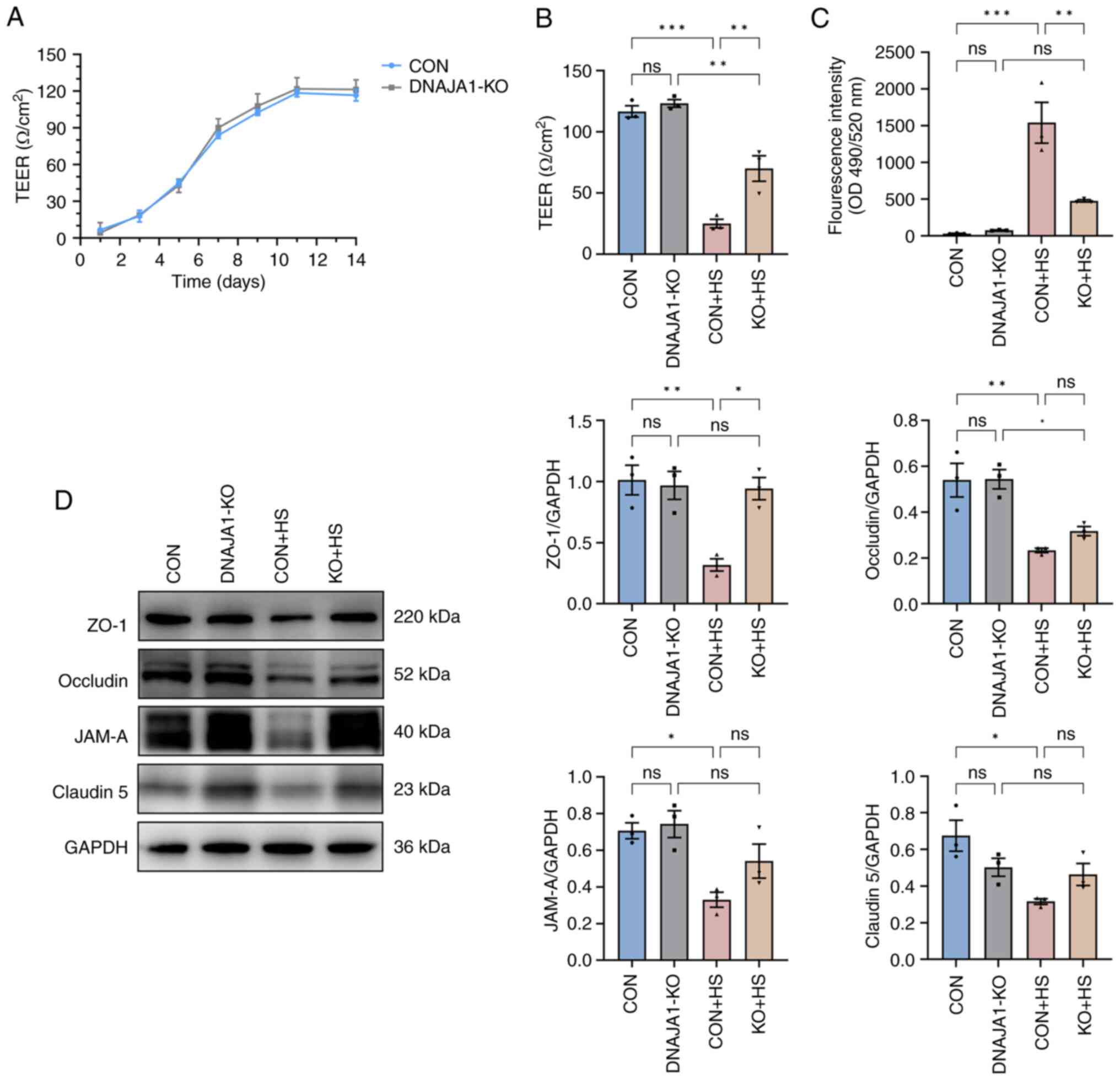 | Figure 3.DNAJA1-KO alleviates HS-induced
endothelial barrier dysfunction in the HUVEC monolayer. Endothelial
monolayer samples were cultured at 43°C for 4 h and experiments
were conducted on confluent HUVECs monolayers after a stable TEER
was recorded. (A) TEER was monitored at the indicated time points
and was considered to form the integrated endothelial monolayer
when the values remained stable after increasing. (B) TEER in each
group was detected and compared after 4 h of HS treatment. (C)
Concentrations of FITC-Dextran in the bottom chamber were measured
and compared after 4 h of HS treatment. (D) Expression levels of
ZO-1, JAM-A, Claudin-5 and occludin were analyzed by western
blotting, with GAPDH used as the loading control. Representative
data are presented as the mean ± SEM (n=3). *P<0.05,
**P<0.01, ***P<0.001. ns, not significant; KO, knockout;
TEER; transendothelial electrical resistance; HS, heat stroke; CON,
control; HUVECs, human umbilical vein endothelial cells; ZO-1,
zonula occludens-1; JAM-A, junctional adhesion molecule A. |
DNAJA1-KO protects against HS-induced
damage of the cytoskeleton and TJs
Although the expression of TJ proteins was
maintained in DNAJA1-KO cells, the arrangement of the TJ proteins
into a particular formation was required for the TJs to act as a
functional barrier (22). It has
also been reported that both the downregulation of TJ protein
expression and the alterations of subcellular localization of TJ
proteins contribute to barrier dysfunction (23). ZO-1 and JAM-A were examined by
immunofluorescence assay to evaluate the structural integrity of
the TJs. As shown in Fig. 4, in
the CON and DNAJA1-KO cell monolayers without HS, ZO-1 staining
exhibited an integrated and continuous structure with belt-like
lines. However, this continuous structure was difficult to observe
in CON + HS cells, and the overall fluorescence intensity also
decreased compared with that in untreated CON cell monolayers.
Correspondingly, a clear structure of ZO-1 staining could be
observed in KO + HS, with a slight decrease in fluorescence
intensity compared with KO (Fig.
4A). Similar results were obtained for JAM-A, which indicated
that the disrupted structure of JAM-A IN CON + HS was protected by
DNAJA1-KO (Fig. 4B). Meanwhile,
TRITC phalloidin was used to investigate the changes in F-actin in
HUVECs under HS. F-actin is a key cytoskeletal component, which is
dynamic in the context of cell-cell junctions and has been reported
to be rearranged under HS (8,24,25).
Similar to the findings of previous studies (8), the cortical actin rim became
incomplete, the inter-cell gap widened, and stress fibers thickened
and reorganized in CON + HS cells subjected to HS at 43°C, all of
which was alleviated in KO + HS cells (Fig. 4A and B).
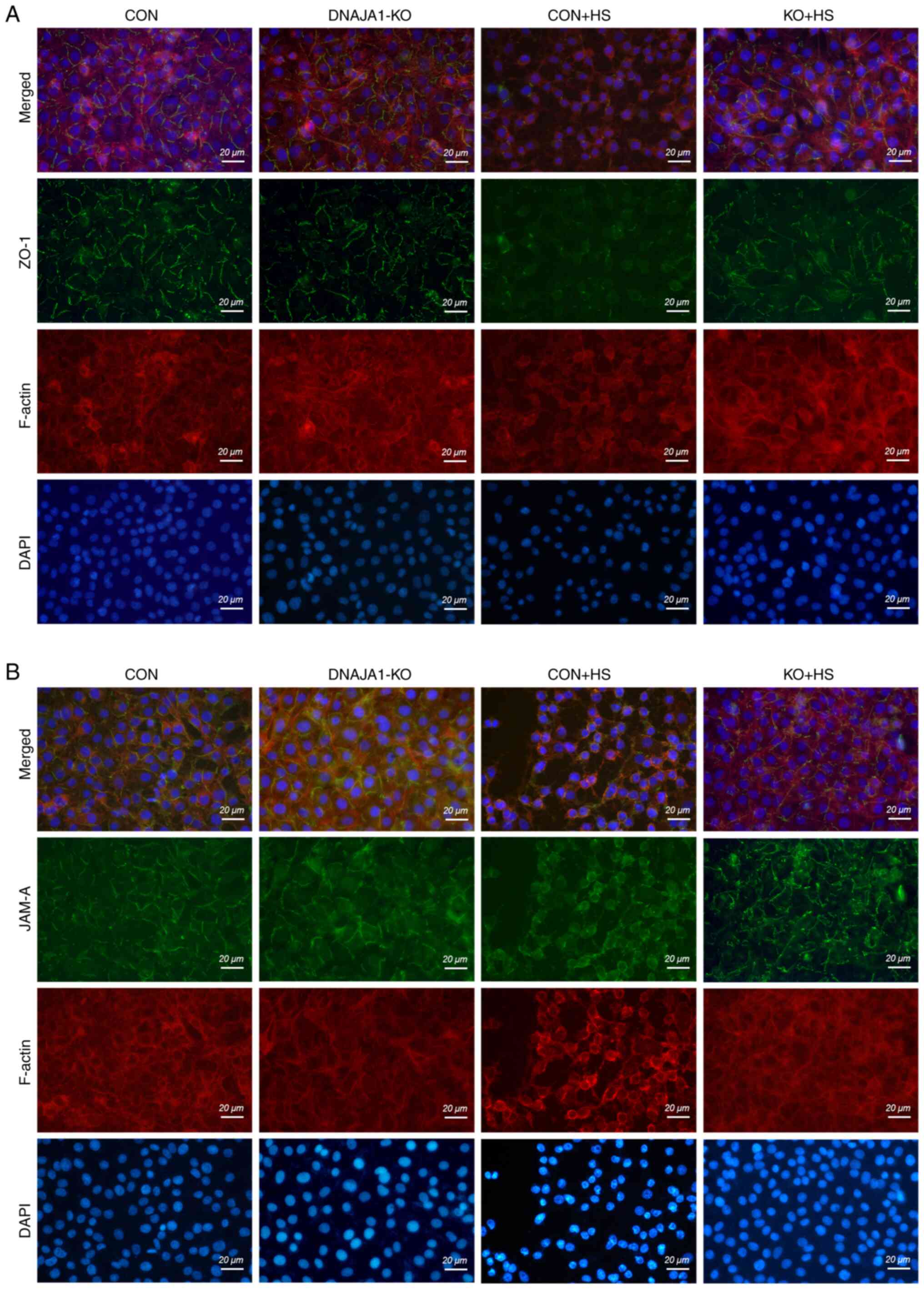 | Figure 4.DNAJA1-KO protects the HS-damaged
cytoskeleton and TJs in the HUVEC monolayer. Confluent HUVEC
monolayers were cultured at 43°C for 4 h and prepared for testing
immediately thereafter. The cytoskeleton and TJs were examined
using immunofluorescence analysis. (A) ZO-1 (green), phalloidin
(red) and DAPI (blue) staining were conducted to assess the
integrity of the cytoskeleton and TJs. (B) JAM-A (green),
phalloidin (red) and DAPI (blue) staining were conducted to assess
the integrity of the cytoskeleton and TJs (scale bar, 20 µm). KO,
knockout; HS, heat stroke; CON, control; HUVECs, human umbilical
vein endothelial cells; ZO-1, zonula occludens-1; JAM-A, junctional
adhesion molecule A; TJs, tight junctions. |
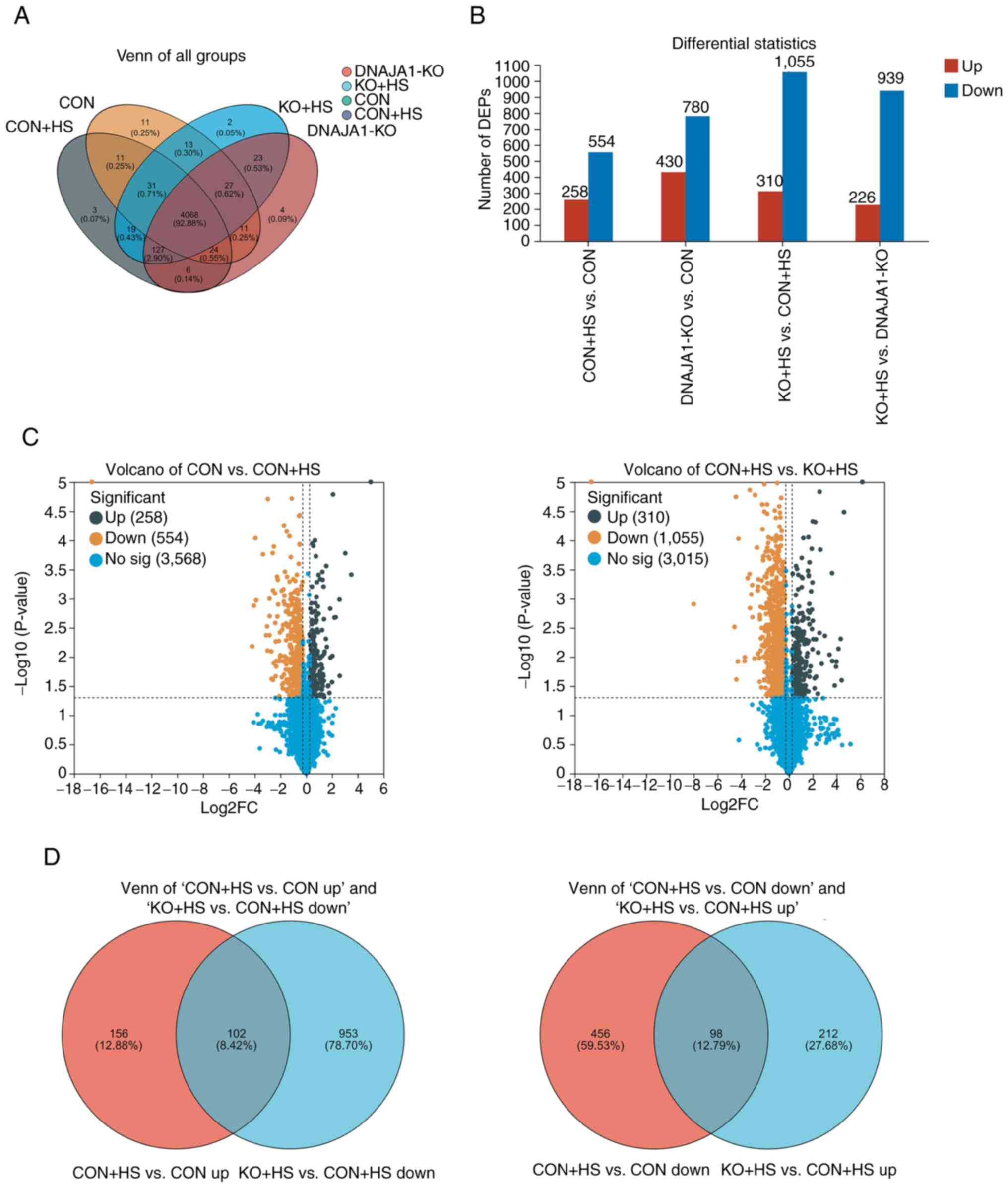
Proteomics analysis and identification
of DEPs in HUVECs
A proteomics experiment using a label-free technique
was performed to analyze and identify the DEPs of the four groups
of HUVECs. A total of 4,394 expressed proteins were identified as
belonging to the proteome of HUVECs in the present study. The
distribution of these proteins in the four groups is shown in a
Venn diagram (Fig. 5A). These
proteins were analyzed statistically with thresholds of fold change
(>1.2 or <0.83) and P<0.05. As a result, 2,499 proteins
were identified as DEPs and were plotted in statistical histograms
and volcano plots (Fig. 5B and C).
Furthermore, Venn diagrams (Fig.
5D) were employed to analyze the distribution of upregulated
proteins in CON + HS compared with CON cells (CON + HS vs. CON up),
downregulated proteins in KO + HS compared with CON + HS cells (KO
+ HS vs. CON + HS down), downregulated proteins in CON + HS
compared with CON cells (CON + HS vs. CON down) and upregulated
proteins in KO + HS compared with CON + HS cells (KO + HS vs. CON +
HS up). In the present study, the protein expression differences in
CON vs. CON + HS and KO + HS vs. CON + HS were mainly focused on.
As a result, 102 DEPs were found to be upregulated in CON + HS
compared with CON and downregulated in KO + HS compared with CON +
HS. These proteins, activated by HS and inhibited by KO, could be
described as ‘bad’ proteins. Correspondingly, 98 DEPs were
downregulated in CON + HS compared with CON, and upregulated in KO
+ HS compared with CON + HS. These proteins, activated by HS and
inhibited by KO, could be described as ‘good’ proteins. Analysis of
these screened proteins could help to clarify the mechanisms
underlying the protective effects of DNAJA1-KO.
Analysis of interaction networks for
DEPs
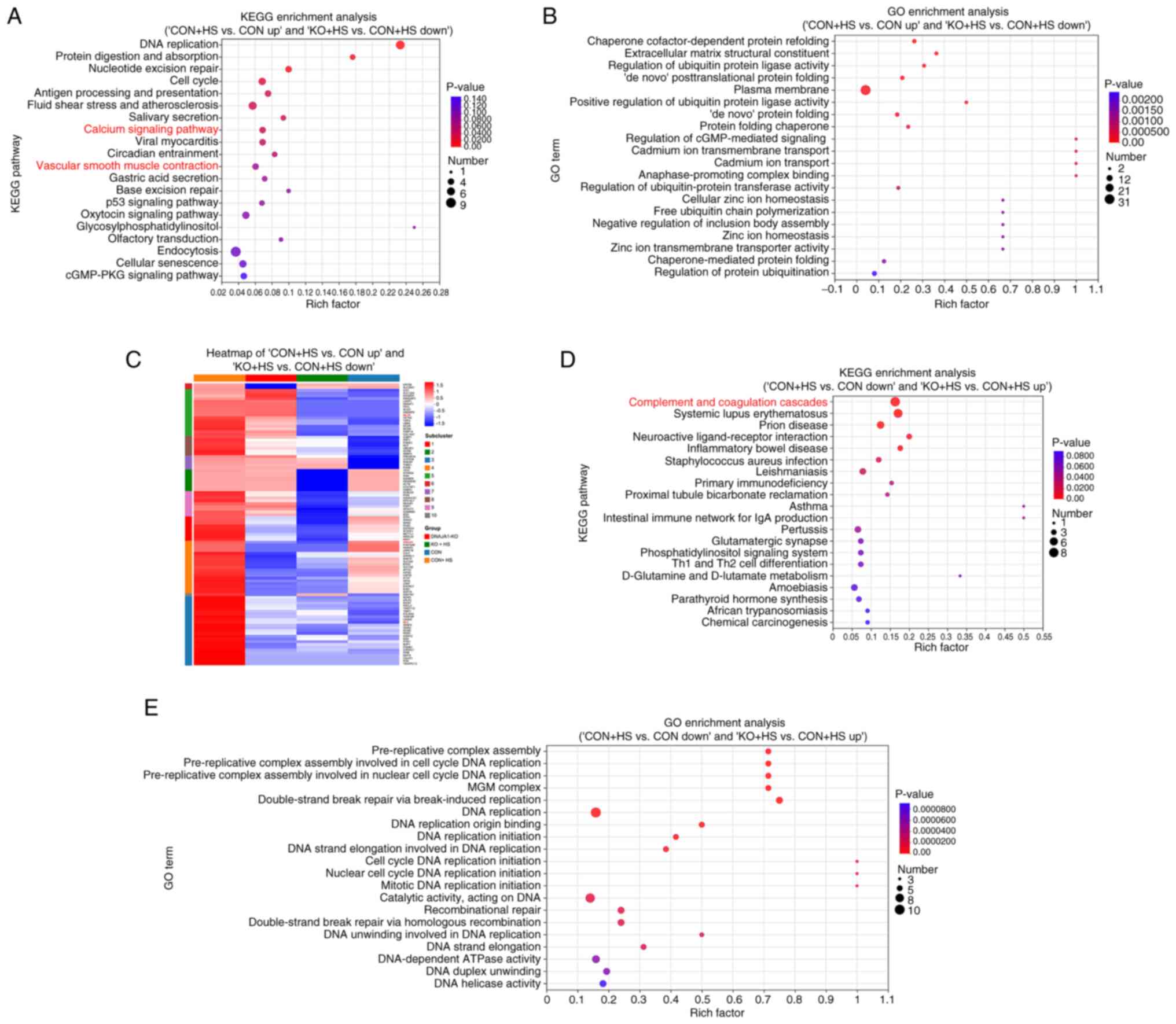
KEGG and GO enrichment analyses were performed to
improve the understanding of the functional basis and biological
pathways of the aforementioned screened DEPs. KEGG enrichment
analysis of the 102 DEPs revealed that 10 pathways were
significantly enriched (Fig. 6A),
among which, the ‘calcium signaling pathway’ and vascular smooth
muscle contraction were highly relevant to the present study.
Therefore, these two pathways were further analyzed and the
associated proteins framed in blue (Fig. S1B-E). Coincidently, the two
pathways were similar and shared various DEPs with parallel
regulation, including MLCK and calmodulin. GO enrichment analysis
of the 102 DEPs demonstrated that the enriched pathways were
associated with a number of protein-folding processes (Fig. 6B). KEGG enrichment analysis of the
98 DEPs revealed that 13 pathways were significantly enriched
(Fig. 6D). GO enrichment analysis
of the 98 DEPs revealed that the enriched pathways were associated
with numerous DNA replication-related processes (Fig. 6E). Enriched complement and
coagulation cascades may be related to endothelial injury-induced
coagulopathy in HS. As the 102 ‘bad’ DEPs could provide novel
potential pathways, a heat map was used to classify the DEP
expression pattern between ‘CON + HS vs. CON up’ and ‘KO + HS vs.
CON + HS down’ (Fig. 6C).
Hierarchical clustering of the 102 DEPs showed distinguished
expression patterns between the four groups. Particularly, proteins
framed in red echoed the results of the aforementioned results of
the present study and were demonstrated to be significant.
DNAJA1-KO on HS is regulated by the
MLCK-MLC signaling pathway
It is widely recognized that MLCK plays a key role
in the regulation of TJs, and the associated endothelial and
epithelial barrier (26).
Moreover, pathways that were found to be enriched by proteomic
analysis confirmed that MLCK had a negative or pro-destructive role
in the protection of DNAJA1-KO. To further investigate the
MLCK-related signaling pathway, MLCK, MLC, p-MLCK and p-MLC were
examined by western blotting and semi-quantified relative to GAPDH
(Fig. 7A). Consistent with the
proteomics results, MLCK expression was significantly elevated in
CON + HS cells and inhibited by DNAJA1-KO in KO + HS cells. The
expression levels of p-MLC were accordingly increased in CON + HS
cells and reduced in KO + HS cells, whereas no statistical
difference was found in p-MLCK and MLC between the groups (Fig. 7D). The ratio of p-MLCK to total
MLCK (Fig. 7B) and that of p-MLC
to total MLC (Fig. 7C) were
semi-quantified to further verify the mechanism. The ratio of
p-MLCK to total MLCK was not statistically significant, whereas the
ratio of p-MLC to total MLC was significantly higher in CON + HS
cells than that in both CON and KO + HS groups, indicating that the
activation of p-MLC may be associated with MLCK but not with
unchanged MLC. These results suggested that DNAJA1-KO could
alleviate HS-induced endothelial barrier disruption by
downregulating the elevated MLCK expression and reducing p-MLC
expression.
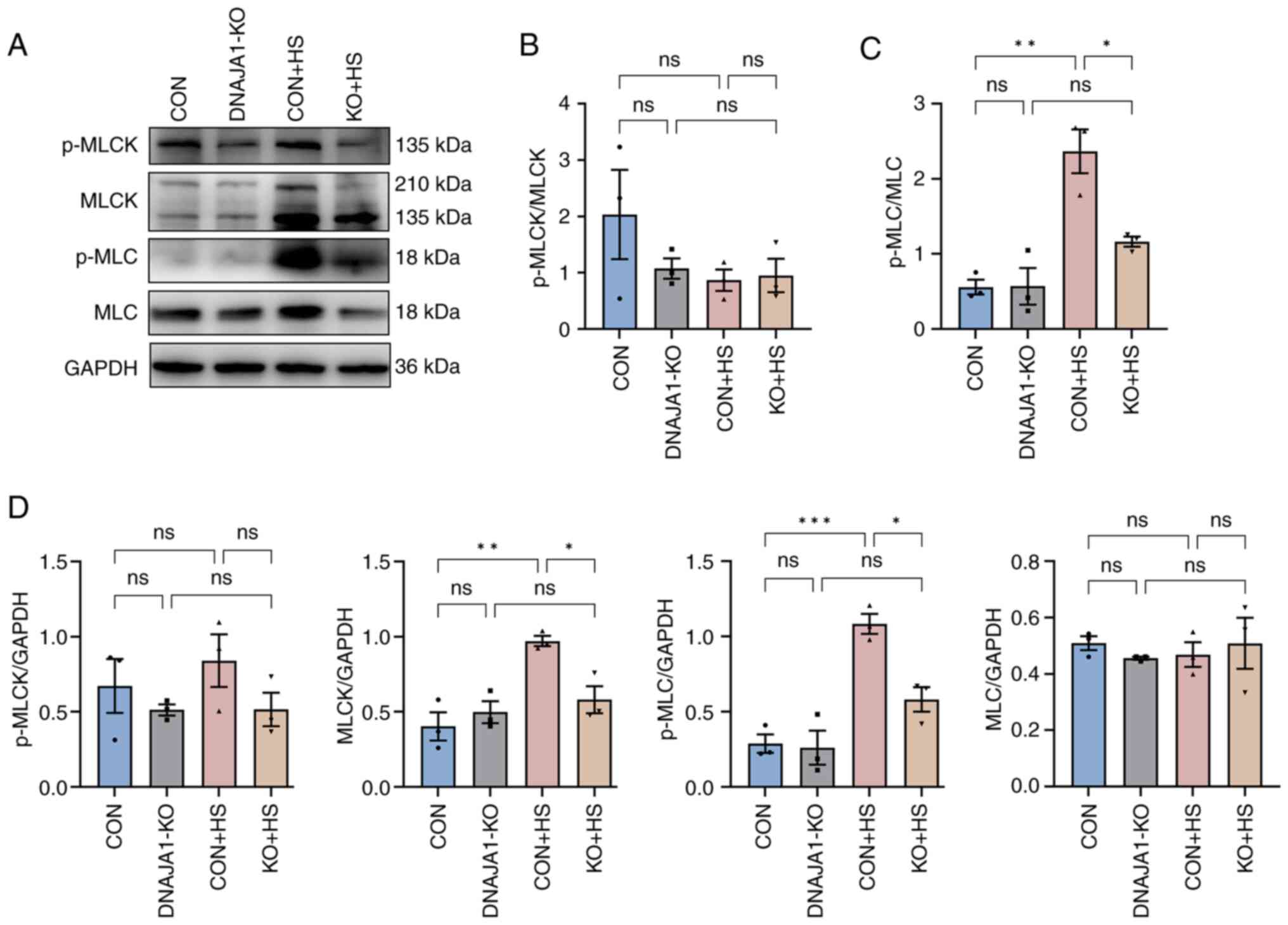 | Figure 7.DNAJA1-KO inhibits HS-induced
activation of MLCK, MLC, p-MLCK and p-MLC. (A) Expression levels of
p-MLCK, MLCK, p-MLC and MLC were measured by western blotting. (B)
Ratio of p-MLCK to total MLCK was semi-quantified. (C) Ratio of
p-MLC to total MLC was semi-quantified. (D) Expression levels of
p-MLCK, MLCK, p-MLC and MLC were semi-quantified relative to GAPDH
(loading control). Representative data are presented as the mean ±
SEM (n=3). *P<0.05, **P<0.01, ***P<0.001. ns, not
significant; KO, knockout; HS, heat stroke; p-, phosphorylated;
MLC, myosin light-chain; MLCK, MLC kinase. |
DNAJA1-KO attenuates multiple-organ
injury and endothelial barrier dysfunction in HS mice
All KO mice were genotyped by PCR analysis (Fig. S1F). Multiple-organ injury in mice
of each group was evaluated by detecting biochemical markers that
represent the related function of each organ. Serum biochemical
markers of liver function (ALT and AST), kidney function (BU and
CREA), and skeletal and/or cardiac muscle function (CK) were
determined to evaluate organ injury. As shown in Fig. 8A-E, the marked organ injury of CON
+ HS mice was markedly inhibited by DNAJA1-KO. Among the
biochemical markers, although BU and CK were not significantly
decreased, a downward trend was apparent. In addition, VWF, TM and
ET-1 were selected to evaluate the degree of vascular endothelial
injury (Fig. 8F-H). Significant
high levels of serum vascular endothelial injury biomarkers were
detected in CON + HS, while the levels of these three biomarkers
were relatively low in KO + HS. Consistent with the serum
biomarkers, lung, as an organ rich in vascular endothelial cells,
showed alveolar wall thickening, narrowed alveolae, numerous
monocytes scattered with granulocytes and severe congestion in
vessels from CON + HS mice (Fig.
8K). Typical lung injury was attenuated by DNAJA1-KO as shown
in Fig. 8L. VE-cadherin, which
represents vascular endothelial function, had a markedly lowered
expression in the lungs of mice subjected to HS induction (Fig. 8O). The expression of VE-cadherin
was notably higher in the KO + HS group (Fig. 8P) compared with the CON + HS group,
which indicated that DNAJA1-KO could counteract the HS-induced lung
vascular endothelial barrier dysfunction.
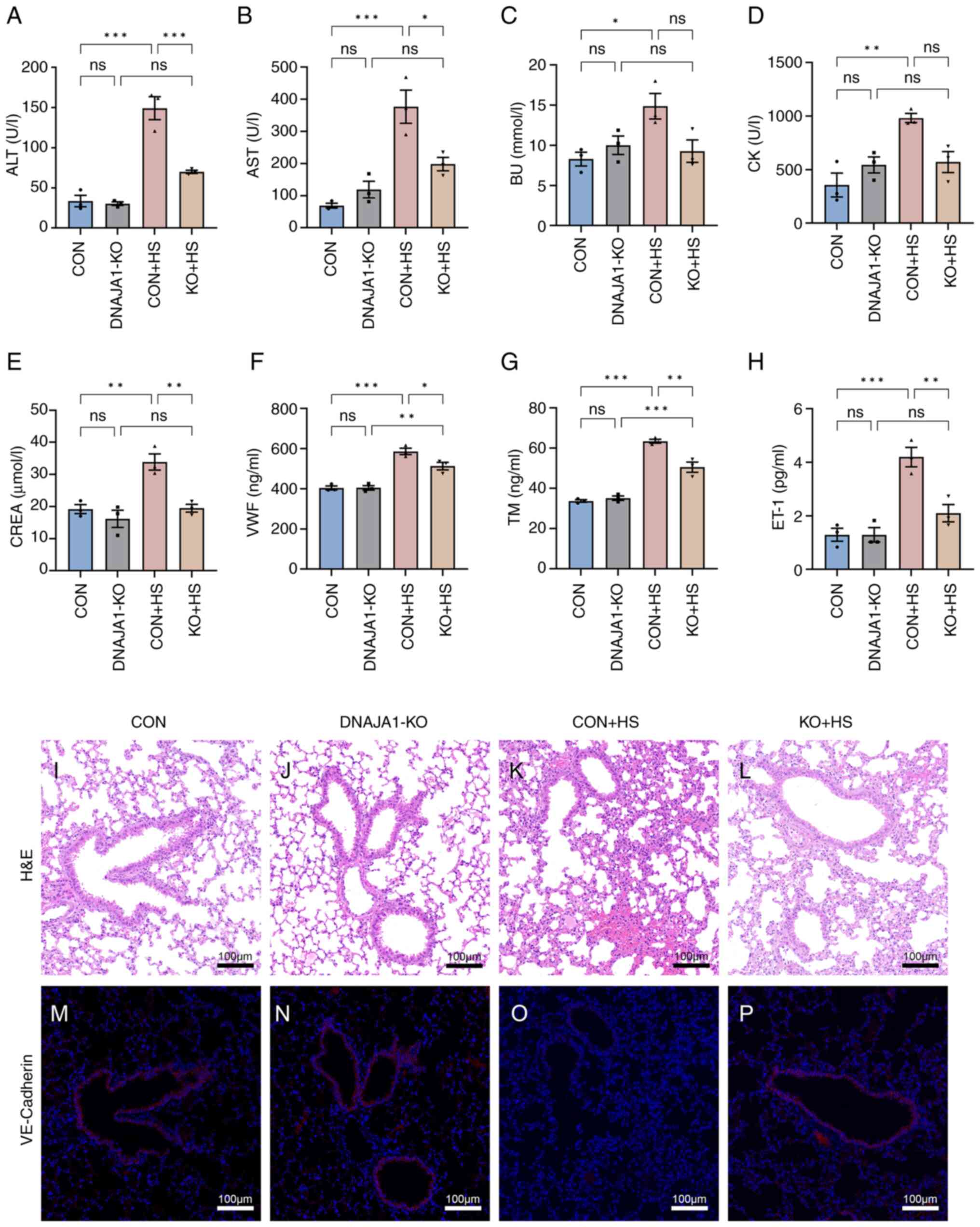 | Figure 8.DNAJA1-KO alleviates multiple-organ
injury and lung endothelial barrier dysfunction in HS mice. Blood
samples of mice in each group were collected 3 h after HS
induction. Serum biochemical indices of (A) ALT, (B) AST, (C) BU,
(D) CK and (E) CREA were examined by automatic biochemical
analyzer. Serum (F) VWF, (G) TM and (H) ET-1 were detected by ELISA
kits. Representative histopathological images are shown of lungs
from (I) CON, (J) DNAJA1-KO, (K) CON + HS and (L) KO + HS mice
(magnification, ×100; scale bar, 100 µm). (M-P) VE-cadherin was
stained with the corresponding antibody (red) and cell nuclei were
stained using DAPI (blue; scale bar, 100 µm) in (M) CON, (N)
DNAJA1-KO, (O) CON + HS and (P) KO + HS (magnification, ×100; scale
bar, 100 µm). Representative data are presented as the mean ± SEM
(n=3). *P<0.05, **P<0.01, ***P<0.001. ns, not significant;
KO, knock out; HS, heat stroke; ALT, alanine aminotransferase; AST,
aspartate aminotransferase; BU, blood urea; CK, creatine kinase;
CREA, creatinine; VWF, von Willebrand factor; TM, thrombomodulin;
ET-1, endothelin-1; CON, control; H&E, hematoxylin and
eosin. |
Discussion
The endothelium, formed by squamous endothelial
cells, is a highly dynamic layer that lines the heart, blood
vessels, lymph vessels and serous cavities of the body (26). It is involved in numerous
physiological functions, including filtrating fluid, forming a
barrier, maintaining vascular tissues, regulating immune function
and other organ-specific functions (27). Endothelial responses are damaged by
exposure to heat stress and play a key role in the pathophysiology
of HS (7). Heat stress not only
damages the endothelium directly in the acute phase, but also
indirectly disrupts endothelial structures and functions through
the inflammatory immune response (28). Numerous studies have found that
endothelial cells can be damaged by HS a high temperature of 43°C
(29–31). To identify the specific gene
associated with thermal tolerance, genome-wide CRISPR-Cas9 screen
research was conducted on HUVECs (10). The increased cell viability and
decreased concentration of LDH release preliminarily proved that
DNAJA1-KO improved the thermal tolerance of HUVECs. A number of
researchers consider that cells become apoptotic immediately after
HS (18,19,29,32).
The present study showed that the rate of early apoptotic cells was
significantly decreased in KO + HS cells. At this point, it was
proposed that the ameliorative effects of DNAJA1-KO on cell damage
or death could be interpreted as an improvement in thermal
tolerance. As aforementioned, DNAJA1 belongs to the HSP40 family
and is essential for the heat shock response. A previous study
found that HSP90 protein inhibitors may help to protect and restore
endothelial barrier function following LPS treatment via regulating
myosin phosphatase target subunit 1 (14,33).
Notably, another study suggested that the HSP70 protein can play a
protective role in sepsis by maintaining endothelial permeability
via p38 signaling (34). Studies
have indicated that HSP proteins have a huge number of families and
are associated with various biological effects (13). To the best of our knowledge,
DNAJA1, as a member of the HSP40 family, has not been previously
investigated in terms of its effects on the endothelial barrier.
The notable protective effects of DNAJA1-KO on HS led to further
investigations into whether it could help sustain endothelial
barrier integrity. Therefore, a confluent monolayer of HUVECs was
successfully constructed by monitoring the TEER according to
previously reported methods (35,36).
It has been reported that a decrease in TEER inversely reflects a
change in endothelial monolayer permeability (37). Endothelial monolayer permeability
assays showed that the barrier function disruption of HS could be
inhibited by DNAJA1-KO. The maintenance of endothelial junction
integrity is governed by proteins that establish adhesion and TJs,
thereby facilitating intercellular communication in a dynamic
manner. Endothelial cell TJs share numerous features with TJs in
epithelial cells and have been explored widely. Several
representative TJ proteins were downregulated by HS and upregulated
by DNAJA1-KO. Staining of ZO-1 and JAM-A further confirmed these
barrier protective effects. Moreover, the endothelial cytoskeleton
plays a critical role in barrier function and regulation. The
hyperpermeability of the endothelial monolayer is attributed to the
destabilization of actin microfilaments or disassembly of
microtubules (38). Growing
evidence has indicated that HSP proteins participate in the
organization of the cytoskeleton in various cell lines such as the
HaCaT and HeLa cell lines (39–42)
It was also found that the structure of F-actin was damaged by HS
and mitigated by DNAJA1-KO, which could sustain the cytoskeleton
structure. To date, the protective effects of DNAJA1-KO on
HS-induced endothelial barrier disruption have only been classified
at a basic level.
Proteomic analysis was conducted to determine the
potential pathways responsible for the specific underlying
mechanism of DNAJA1-KO in HS. It was hypothesized that proteins
activated by HS and inhibited by DNAJA1-KO could play a pro-damage
role, while those inhibited by HS and activated by DNAJA1-KO could
play an anti-damage role. As a result, 102 ‘pro-damage’ proteins
and 98 ‘anti-damage’ proteins were identified, while the subsequent
enrichment analysis provided potential pathways to verify these
results. Among these enriched pathways, the calcium signaling
pathway and vascular smooth muscle contraction pathway were of note
as they are often reported to be related to regulating the
endothelial barrier. Both signaling pathways were shown to be
associated with the MLCK-MLC pathway, and the trend shown by
proteomics was consistent with relevant literature reports
(43–45). In endothelial cells, the calcium
signaling pathway is crucial for modulating barrier function and
promoting inflammation. It could cause cell-cell junction
disassembly and cytoskeletal rearrangement to facilitate
endothelial cell retraction and increase permeability (22,23).
Moreover, the associated proteins were activated by HS and
inhibited by DNAJA1-KO, which is consistent with the aforementioned
hypothesis. At the same time, it was found that the MLCK-MLC
pathway was highly related to the regulation of vascular
endothelial barrier under heat stress through sufficient literature
research (46–48). The pivotal protein, MLCK, a key
effector of barrier dysfunction, was verified by western blotting.
As aforementioned, the elevated expression of MLCK activated MLC
and increased the expression of p-MLC in CON + HS cells, which may
cause remodeling of the actin cytoskeleton, leading to endothelial
hyperpermeability. The present study demonstrated that DNAJA1-KO
could inhibit HS-induced MLCK expression and block the subsequent
MLC phosphorylation. Taken together, these data confirmed that
HS-induced endothelial barrier disruption was mitigated in
DNAJA1-KO cells, as indicated by the results of functional tests,
and TJ and actin cytoskeleton observations. Moreover, DNAJA1-KO
could improve the thermal tolerance of HUVECs, as indicated by cell
viability and apoptosis assays. Regarding the mechanism underlying
the effects of DNAJA1-KO on the HS-damaged barrier, involvement of
the MLCK-MLC signaling pathway was demonstrated from the
perspective of endothelial barrier regulation, but could not
explain the complete regulatory pathways of DNAJA1-KO-mediated
thermal tolerance effects. Although, it was successfully confirmed
that DNAJA1KO has a significant protective effect against heat
stress injury and HS in vivo and in vitro, DNAJA1
overexpression cell lines must be constructed in future work to
fully clarify the mechanisms of the gene action in the future.
In conclusion, DNAJA1-KO could effectively protect
the endothelial barrier against HS via improving thermal tolerance
and suppressing the MLCK-MLC signaling pathway. These findings
emphasize the distinct effects of HSPs on HS and provide a novel
insight into the classic heat shock response, which could assist
with the identification of therapeutic targets against HS.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This research was funded by the National Key Basic Research
(grant no. 2022-JCJQ-ZD-097-11) and the Shanghai Military Civilian
Integration Industry Development Project (grant no.
2019-jmrh1-kj52).
Availability of data and materials
The raw data of proteomic analysis generated in the
present study may be found in the ProteomeXchange under accession
number PXD043512 or at the following URL (https://www.ebi.ac.uk/pride/archive/projects/PXD043512).
The remaining data generated in the present study may be requested
from the corresponding author.
Authors' contributions
JKC, SGX and JBZ designed the study and, analyzed
and interpreted data. JKC and SGX confirm the authenticity of all
the raw data. LL, YWW, XC, JLC, MW, JQZ, JFL, LJR, XYD, LY, QS and
XCF conducted the experiments and analyzed the data. LL, YWW and XC
wrote the article. JKC, SGX, QS and LL revised the article. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Committee on
Ethic of Medical Research (The Institutional Animal Ethics
Committee), Naval Medical University (Shanghai, China; approval no.
NMUMREC-2021-002).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bouchama A, Abuyassin B, Lehe C, Laitano
O, Jay O, O'Connor FG and Leon LR: Classic and exertional
heatstroke. Nat Rev Dis Primers. 3:82022. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Epstein Y and Yanovich R: Heatstroke. N
Engl J Med. 380:2449–2459. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Raymond C, Matthews T and Horton RM: The
emergence of heat and humidity too severe for human tolerance. Sci
Adv. 6:eaaw18382020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fujimoto M and Nishiura H: Baseline
scenarios of heat-related ambulance transportations under climate
change in Tokyo, Japan. PeerJ. 10:e138382022. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Aird WC: Endothelium as an organ system.
Crit Care Med. 32:S271–S279. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cheng JL and MacDonald MJ: Effect of heat
stress on vascular outcomes in humans. J Appl Physiol (1985).
126:771–781. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Iba T, Connors JM, Levi M and Levy JH:
Heatstroke-induced coagulopathy: Biomarkers, mechanistic insights,
and patient management. EClinicalMedicine. 44:1012762022.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xu Q, Liu J, Wang Z, Guo X, Zhou G, Liu Y,
Huang Q and Su L: Heat stress-induced disruption of endothelial
barrier function Is via PAR1 signaling and suppressed by Xuebijing
injection. PLoS One. 10:e01180572015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shi J, Zhao Y, Wang K, Shi X, Wang Y,
Huang H, Zhuang Y, Cai T, Wang F and Shao F: Cleavage of GSDMD by
inflammatory caspases determines pyroptotic cell death. Nature.
526:660–665. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang S: Genome-wide CRISPR/Cas9 library
screening identified DNAJA1 regulating protein homeostasis in heat
stress injury, master's dissertation. Naval medical university;
2023
|
|
11
|
Ergulen E, Becsi B, Csomos I, Fésüs L and
Kanchan K: Identification of DNAJA1 as a novel interacting partner
and a substrate of human transglutaminase 2. Biochem J.
473:3889–3901. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Qiu XB, Shao YM, Miao S and Wang L: The
diversity of the DnaJ/Hsp40 family, the crucial partners for Hsp70
chaperones. Cell Mol Life Sci. 63:2560–2570. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kurop MK, Huyen CM, Kelly JH and Blagg
BSJ: The heat shock response and small molecule regulators. Eur J
Med Chem. 226:1138462021. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Joshi AD, Dimitropoulou C, Thangjam G,
Snead C, Feldman S, Barabutis N, Fulton D, Hou Y, Kumar S, Patel V,
et al: Heat shock protein 90 inhibitors prevent LPS-induced
endothelial barrier dysfunction by disrupting RhoA signaling. Am J
Respir Cell Mol Biol. 50:170–179. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ren Y, Yu G, Shi C, Liu L, Guo Q, Han C,
Zhang D, Zhang L, Liu B, Gao H, et al: Majorbio cloud: A one-stop,
comprehensive bioinformatic platform for multiomics analyses.
iMeta. 1:e122022. View Article : Google Scholar
|
|
16
|
Health NIO, . Guide for the care and use
of laboratory animals. National Academies; 1985
|
|
17
|
Li L, Man W, Chen J, Xu Z, Wang S, Xia X,
Liu D, Wang S, Xie C, Wu J, et al: Preventive effects of bacillus
licheniformis on heat stroke in rats by sustaining intestinal
barrier function and modulating gut microbiota. Front Microbiol.
12:5482021.
|
|
18
|
Gu ZT, Li L, WU F, Zhao P, Yang H, Liu YS,
Geng Y, Zhao M and Su L: Heat stress induced apoptosis is triggered
by transcription-independent p53, Ca2+ dyshomeostasis and the
subsequent Bax mitochondrial translocation. Sci Rep. 5:114972015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gu ZT, Wang H, Li L, Liu YS, Deng XB, Huo
SF, Yuan FF, Liu ZF, Tong HS and Su L: Heat stress induces
apoptosis through transcription-independent p53-mediated
mitochondrial pathways in human umbilical vein endothelial cell.
Sci Rep. 4:44692015. View Article : Google Scholar
|
|
20
|
Zhang S, Liu Y, Wang Z, Liu J, Gu Z, Xu Q
and Su L: PAR1-mediated c-Jun activation promotes heat
stress-induced early stage apoptosis of human umbilical vein
endothelial cells. Mol Med Rep. 15:2595–2603. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang Y, Yang C, Elsheikh N, Elsheikh NAH,
Li C, Yang F, Wang G and Li L: HO-1 reduces heat stress-induced
apoptosis in bovine granulosa cells by suppressing oxidative
stress. Aging (Albany NY). 11:5535–5547. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dalal PJ, Muller WA and Sullivan DP:
Endothelial cell calcium signaling during barrier function and
inflammation. Am J Pathol. 190:535–542. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Claesson-Welsh L, Dejana E and McDonald
DM: Permeability of the endothelial barrier: Identifying and
reconciling controversies. Trends Mol Med. 27:314–331. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Clarke JP and Mearow KM: Cell stress
promotes the association of phosphorylated HspB1 with F-actin. PLoS
One. 8:e689782013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yin B, Tang S, Xu J, Sun J, Zhang X, Li Y
and Bao E: CRYAB protects cardiomyocytes against heat stress by
preventing caspase-mediated apoptosis and reducing F-actin
aggregation. Cell Stress Chaperones. 24:59–68. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dzwinel W, Boryczko K and Yuen DA: A
discrete-particle model of blood dynamics in capillary vessels. J
Colloid Interface Sci. 258:163–173. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Joffre J and Hellman J: Oxidative Stress
and endothelial dysfunction in sepsis and acute inflammation.
Antioxid Redox Signal. 35:1291–1307. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Iba T, Helms J, Levi M and Levy JH:
Thromboinflammation in acute injury: Infections, heatstroke, and
trauma. J Thromb Haemost. 22:7–22. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu Z, Zhong T, Zheng D, Cepinskas I, Peng
T and Su L: Heat stress pretreatment decreases
lipopolysaccharide-induced apoptosis via the p38 signaling pathway
in human umbilical vein endothelial cells. Mol Med Rep.
14:1007–1013. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Huang W, Mao L, Xie W, Cai S, Huang Q, Liu
Y and Chen Z: Impact of UCP2 depletion on heat stroke-induced
mitochondrial function in human umbilical vein endothelial cells.
Int J Hyperthermia. 39:287–296. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wu F, Dong XJ, Zhang HQ, Li L, Xu QL, Liu
ZF, Gu ZT and Su L: Role of MnSOD in propofol protection of human
umbilical vein endothelial cells injured by heat stress. J Anesth.
30:410–419. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu Y, Zhou G, Wang Z, Guo X, Xu Q, Huang
Q and Su L: NF-κB signaling is essential for resistance to heat
stress-induced early stage apoptosis in human umbilical vein
endothelial cells. Sci Rep. 5:135472015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Antonov A, Snead C, Gorshkov B, Antonova
GN, Verin AD and Catravas JD: Heat shock protein 90 inhibitors
protect and restore pulmonary endothelial barrier function. Am J
Respir Cell Mol Biol. 39:551–559. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yuan X, Chen Y, Chen G, Liu G, Hang M,
Wang P, Luo Y, Guo D and Xu L: The heat shock protein 70 plays a
protective role in sepsis by maintenance of the endothelial
permeability. Biomed Res Int. 2020:21940902020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bi J, Zhang J, Ren Y, Du Z, Zhang Y, Liu
C, Wang Y, Zhang L, Shi Z, Wu Z, et al: Exercise hormone irisin
mitigates endothelial barrier dysfunction and microvascular
leakage-related diseases. JCI Insight. 5:e1362772020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang Z, Lu YL, Chen M, Xu HF and Zheng LR:
Piceatannol alleviates glucolipotoxicity induced vascular barrier
injury through inhibition of the ROS/NF-kappa B signaling pathway.
Am J Transl Res. 14:120–134. 2022.PubMed/NCBI
|
|
37
|
Du L, Dong F, Guo L, Hou Y, Yi F, Liu J
and Xu D: Interleukin-1beta increases permeability and upregulates
the expression of vascular endothelial-cadherin in human renal
glomerular endothelial cells. Mol Med Rep. 11:3708–3714. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hohmann T and Dehghani F: The
cytoskeleton-A complex interacting meshwork. Cells. 8:3622019.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu R, Niu X, Yuan J, Chen H, Gao X and Qi
R: DnaJA4 is involved in responses to hyperthermia by regulating
the expression of F-actin in HaCaT cells. Chin Med J (Engl).
134:456–462. 2021. View Article : Google Scholar
|
|
40
|
Wettstein G, Bellaye PS, Micheau O and
Bonniaud P: Small heat shock proteins and the cytoskeleton: An
essential interplay for cell integrity? Int J Biochem Cell Biol.
44:1680–1686. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Almeida-Souza L, Asselbergh B, D'Ydewalle
C, Moonens K, Goethals S, de Winter V, Azmi A, Irobi J, Timmermans
JP, Gevaert K, et al: Small heat-shock protein HSPB1 mutants
stabilize microtubules in charcot-marie-tooth neuropathy. J
Neurosci. 31:15320–15328. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kayser J, Haslbeck M, Dempfle L, Krause M,
Grashoff C, Buchner J, Herrmann H and Bausch AR: The small heat
shock protein Hsp27 Affects assembly dynamics and structure of
keratin intermediate filament networks. Biophys J. 105:1778–1785.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Du L, Zhu L, Lu X, Yu Y, Liu P and Pan J:
Inhibition of the MLCK/MLC2 pathway protects against intestinal
heat stroke-induced injury in rats. J Therm Biol. 116:1036552023.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yang PC, He SH and Zheng PY: Investigation
into the signal transduction pathway via which heat stress impairs
intestinal epithelial barrier function. J Gastroenterol Hepatol.
22:1823–1831. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li H, Chen H, Zhang S, Wang S, Zhang L, Li
J, Gao S and Qi Z: Taurine alleviates heat stress-induced mammary
inflammation and impairment of mammary epithelial integrity via the
ERK1/2-MLCK signaling pathway. J Therm Biol. 116:1035872023.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cheng M, Yang Z, Qiao L, Yang Y, Deng Y,
Zhang C and Mi T: AGEs induce endothelial cells senescence and
endothelial barrier dysfunction via miR-1-3p/MLCK signaling
pathways. Gene. 851:1470302023. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhong J, Yu R, Zhou Q, Liu P, Liu Z and
Bian Y: Naringenin prevents TNF-alpha-induced gut-vascular barrier
disruption associated with inhibiting the NF-kappaB-mediated
MLCK/p-MLC and NLRP3 pathways. Food Funct. 12:2715–2725. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Huang Y, Luo X, Li X, Song X, Wei L, Li Z,
You Q, Guo Q and Lu N: Wogonin inhibits LPS-induced vascular
permeability via suppressing MLCK/MLC pathway. Vascul Pharmacol.
72:43–52. 2015. View Article : Google Scholar : PubMed/NCBI
|