Introduction
Cancer is an ongoing public health challenge
globally and currently ranks as the second most common cause of
mortality worldwide (1–3). Owing to the heterogeneity,
uncertainty and imperceptibility of cancer, coupled with the lack
of experimental or clinical research on treatment targets, the
mortality rate of cancer remains high. In addition, cancer has
become a significant barrier to increasing life expectancy in the
21st century (4). Therefore, genes
associated with cancer have become the focus of intense
research.
SPARC/osteonectin, CWCV and Kazal-like domain
proteoglycan (SPOCK) is a chimeric proteoglycan that is highly
conserved. Its modular architecture consists of a core protein,
which consists of SPARC/osteonectin domains, CWCV and Kazal-like
domains. In total, three members of the SPOCK subfamily, namely
SPOCK1, SPOCK2 and SPOCK3, which are members of the
BM-40/SPARC/osteonectin protein family, have follistatin-like and
extracellular calcium-binding domains. SPOCK was initially detected
in the seminal human plasma (5),
but further study has revealed that SPOCK can be found in certain
parts of the nervous system and brain, such as the pyramidal cells
of the hippocampus (6). In
addition, chondrocytes and endothelial cells have been found to
express the SPOCK protein (7).
SPOCK has been previously documented to serve an
important role in controlling the physiology of tumor cells
(8). Therefore, it may be of
benefit to further explore the role of SPOCK proteins in regulating
the degradation or repair of the extracellular matrix (ECM) and
physiology of tumor cells. In the present review, the most recent
research on the mechanism of various isoforms of SPOCK in cancer
was summarized to provide a theoretical basis and research
direction for the development of diagnostic markers or gene therapy
targets.
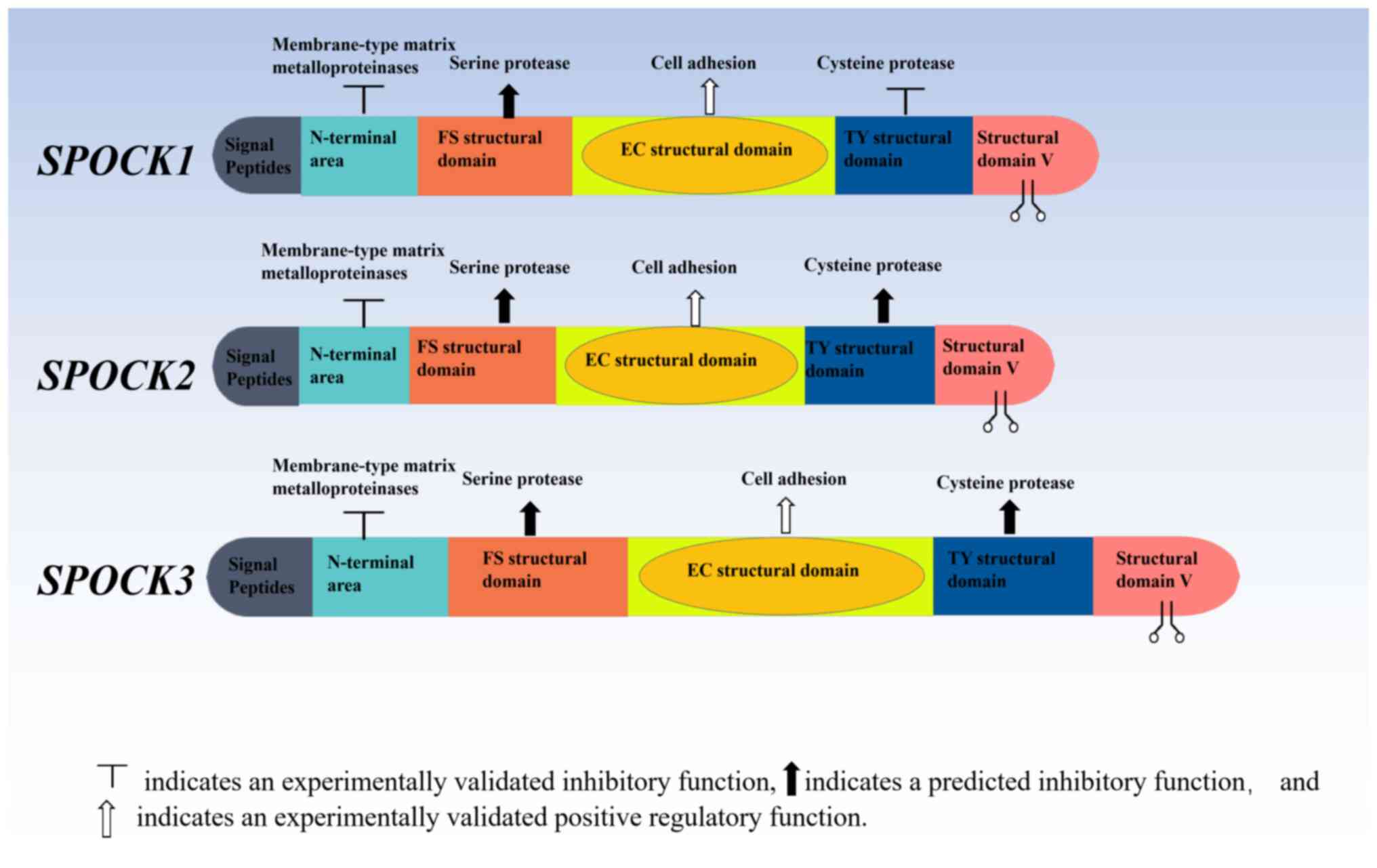
Overview of SPOCK
Structural overview
The SPOCK family consists of cysteine-rich acidic
secretory proteins that represent a group of cysteine-rich acidic
secretory proteins that are integral components of the ECM
(6). The nomenclature of the SPOCK
proteins mirrors their modular architecture. SPOCK has been found
to interact with both cell surface and ECM molecules (6). In addition, each SPOCK family member
has a follicle-like calcium-binding domain. By contrast, the
thyroglobulin domains of SPOCK family members can bind to growth
factors (9) or interact with the
cell matrix (10).
Gene structure and chromosomal
localization
Gene structure of SPOCK
The SPOCK1 gene is the most extensively
characterized member of the SPOCK family. This gene is
located in the long arm of chromosome 5 (5q31), which contains the
chondroitin sulphate chain and heparan sulphate chain, between the
interleukin-9 and early growth response 1 gene segments. It
comprises 11 exons, possessing large introns in the 5′ regions
(11,12). Based on the size of the
non-overlapping exonic genomic clones, exon 1 does not have a
transcriptional function, whereas exon 2 begins transcription at
the ATG initiation site. There is an overlap among exons 2, 3 and 4
and the first two structural domains of osteonectin. By contrast,
exons 5 and 6 encode the third osteonectin- and Kazal-like fields,
respectively. Exons 8 and 10 encode the fourth osteonectin
structural domain and the CWCV structural domains. The last 42
amino acids and the 3′ untranslated region that follows are
surrounded by the final exon. Exons 2 and 4 are 46 bp apart,
supporting the hypothesis of a single exon. All inline-exon
junctions follow the GT/AG rule. This crucial feature in precursor
RNA splicing encompasses two conserved sequences at the intron-exon
junction: GT at the 5′ end and AG at the 3′ end. This rule
predominantly governs eukaryotic gene splicing sites and
underscores a shared mechanism for intron excision. It is important
to note that this conservation does not extend to
post-transcriptional processing of mitochondrial, chloroplast, and
yeast tRNA genes. Analyzing exon 1, which includes the 5′
untranslated region, revealed that the sequence covering the first
270 bp of the cDNA appears to be associated with a chimeric gene
rather than the SPOCK gene. Therefore, the possibility of a
small number of spliced exons in this region must be partially
ruled out. Because not all introns are at the same stage, it is
possible that the alternative splicing of any one of the
aforementioned domains can significantly affect reading frame
interpretation (13).
The SPOCK2 gene is located in the long arm of
chromosome 10 (10q22) and its corresponding protein binds to
glycosaminoglycans (GAGs) to form part of the ECM (14). This gene encodes a protein
precursor consisting of 424 amino acids, containing a
follistatin-like structural domain, a Ca2+-binding
domain, a thyroglobulin type-1 structural domain and two possible
glycosaminoglycan-binding sites at the C-terminus. However, the
N-terminus shows no homology with other known proteins, except for
SPOCK1. The cysteine-rich domain (FS domain) is similar to
that of follistatin-like proteins. The third domain is homologous
to the EC domain of the BM-40 signaling peptide (15). This domain contains two EF-hand
motifs, EF-1 and EF-2, which can bind to Ca2+. All
essential features exhibited by the EC domains are conserved in
SPOCK2. Specifically, the EF-hand binding site in the
binding domain and the recombinant fragment consisting of a
thyroglobulin-like domain can undergo reversible,
Ca2+-dependent conformational changes. Following the EC
domain is the thyroglobulin type-1 domain (TY domain). The TY
domain is stabilized by three conserved disulfide bonds and
contains a characteristic CWCV tetrapeptide sequence (16,17).
The C-terminal domain of SPOCK2 is unique to the SPOCK
subfamily and has two possible GAG attachment sites.
SPOCK3 has an indistinct characteristic for a
SPOCK gene. It is encoded by the human chromosome region
4q32.3, which is similar to SPOCK1 in terms of its structure
and function. It has been previously observed to encode multiple
isoforms of protein caused by alternative splicing transcript
variants (18) (Fig. 1).
Distribution and physiological
function of the SPOCKs
SPOCKs are widely expressed in brain tissue.
SPOCK1 and SPOCK2 have been documented to regulate
the development and repair of the nervous system, providing ideas
for future studies into neurodevelopmental disorders. In terms of
tumorigenesis, SPOCK1 and SPOCK3 have been reported
to mediate oncogenic roles, whereas SPOCK2 tended to serve
as a tumor suppressor (11,13,19,20).
Epigenetic regulation and
expression
Sustained hypermethylation commonly occurs at the
locus of the gene encoding SPOCK1 across diverse
malignancies leading to transcriptional repression (21). Histone deacetylation along with
specific microRNAs (miRs) contribute significantly towards this
gene's silencing within neoplastic lesions (11,22).
SPOCK2 is subject to modulation by distinct histone
methylation patterns specifically within neuronal tissue. MiRs
directed against SPOCK2 exert profound effects on processes
related to neuronal differentiation as well as pathogenesis
underlying conditions like autism spectrum disorder and
intellectual disability (23–25).
Conversely, hypomethylation at the SPOCK3 promoter region
closely associates with heightened expression levels observed
across various carcinomas. Epigenetic control mechanisms involving
histone acetylation alongside long non-coding RNAs demonstrate
pivotal roles during developmental stages within cerebral tissue
(26). The clinical relevance
pertaining to genetic alterations affecting SPOCK family members
remains extensively documented.
Specifically, SPOCK1 exhibits frequent
dysregulation, which is particularly evident within glioblastoma
multiforme (27), as well as
breast carcinoma (28), whereas
SPOCK3 aberrations predominantly manifest within colorectal
cancer (29) and prostate cancer
(30). Mutational events impacting
SPOCK2 associate primarily with neurodevelopmental anomalies
characterized by perturbed histone modification profiles coupled
with miR-mediated regulatory networks (31–33).
Recent findings further indicate the involvement of SPOCK
gene family members in cardiovascular pathology mediated through
epigenetic reprogramming occurring within cardiac tissues (34). A comprehensive understanding
regarding epigenetically governed control mechanisms dictating
expression patterns exhibited by SPOCK genes holds paramount
significance towards unraveling their intricate involvements and
spanning physiological homeostasis pathological state. While
significant progress has been made, further research is required to
fully explore the therapeutic potential of genetically modifiable
loci located on SPOCK.
Functional insights
The human brain exhibits the highest expression
levels of SPOCK1 (35–37),
particularly in the thalamus. Previous cell culture studies have
shown that SPOCK1 can suppress adhesion and facilitates
axonal growth (38). It is also
associated with axon regeneration following injury. In addition,
SPOCK1 has been reported to affect neuronal system
development and interconnectivity (39), whilst being crucial for synaptic
plasticity (40,41). Elsewhere, SPOCK1 is highly
expressed in the adrenal gland, endometrium, prostate, heart,
kidney, gallbladder and testis, with much lower expression levels
in the bone marrow, colon, duodenum, liver, lung, lymph nodes,
ovary, pancreas, salivary glands, skin, small intestine, stomach
and thyroid tissues. In developing mice, SPOCK1 is mainly
expressed in the central and peripheral nervous systems. By
contrast, in adult mice, SPOCK1 is only detectable in the
brain, particularly in the hippocampal neurons.
Human tissues express SPOCK2 widely, with
particularly high expression observed in lung tissues. The brain,
adrenal gland, appendix, bone marrow, kidney, lymph nodes, spleen
and testes also readily express SPOCK2 (20,31).
It exhibits low expression in the colon, duodenum, endometrium,
esophagus, fat, heart, liver, ovary, pancreas, placenta, prostate,
salivary glands and skin. Functionally, SPOCK2 has been
documented to regulate various stages of neurogenesis. This
suggests that SPOCK1 and SPOCK2 form a new family of
calcium-binding proteoglycans that participate in various steps of
neurogenesis (25). According to
its expression pattern, SPOCK2 may be involved in alveolar
formation, regulating the equilibrium between type 1 and 2
epithelial alveolar cells in the lungs. Furthermore, SPOCK2
may also serve as a susceptibility gene for bronchopulmonary
dysplasia (42).
Human tissues express SPOCK3 at higher levels
in the adrenal gland, brain, prostate and ovaries, whilst at lower
levels in the bladder, endometrium, appendix, small intestine and
colon. Previous studies on SPOCK3 were mainly performed on
animal models. During embryonic development in mice, the
SPOCK3 gene has been observed in the neurons of the vascular
system, liver, inner ear and central nervous system (CNS) (43). However, in adulthood, SPOCK3
is only expressed in the brain, suggesting that it may serve a
critical role in the CNS. According to immunohistochemistry
analysis, SPOCK3 expression was found to be the most
abundant in adult mouse brain regions, particularly the olfactory
bulb, cortex, thalamus, hippocampus and striatum (44). Notably, the thalamus, hippocampus
and striatum are regions that have been repeatedly reported to be
associated with psychiatric disorders (45–47).
Therefore, previous studies have explored the association between
SPOCK3 and defective hyperactivity disorder, which
identified single nucleotide polymorphisms in tSPOCK3 gene
loci (48). Mechanistically,
SPOCK1 has been reported to function as a potent competitive
inhibitor of cathepsin L (CTSL). Its inhibitory activity on CTSL is
independent of its chondroitin sulphate GAG and heparan sulphate
GAG, which is optimal at pH 5.5 and 7.2. By contrast, CTSL
inhibition by SPOCK1 is dependent on its TY homology domain
(49). Given that SPOCK2
and SPOCK3 also have TY domains similar to SPOCK1, it
remains possible that SPOCK2 and SPOCK3 can also
function as CTSL inhibitors.
Regulatory effects of SPOCKs in tumor
development
Tumor metastasis is the malignant tumor growth
process at other sites secondary to its primary site and is one of
the main causes of tumor-related mortality (50,51).
The ECM serves as the first barrier to tissue tumor metastasis
(52,53). As an ECM glycoprotein, SPOCK
has been garnering attention because of its possible role in the
pathogenesis of malignancies. As the number of in-depth studies on
the SPOCK family increased, it became evident that this protein
family is associated with the occurrence and progression of solid
tumors.
Roles of SPOCK1 in tumor
development
SPOCK1 is the most extensively studied gene
in the SPOCK family. SPOCK1 has frequently been reported to
function as an oncogene. A multitude of miRNA-targeted genes have
been observed to regulate the occurrence and development of cancer,
leading to successful tumor cell metastasis by inhibiting the
formation of MMP2. In addition, SPOCK1 has been previously
found to promote epithelial-mesenchymal transition (EMT) in tumor
cells to render them more invasive. It can also enhance the
malignancy and invasiveness of tumor cells by participating in the
Wnt and PI3K signaling pathways. These observations suggest that
SPOCK1 is a regulatory factor in numerous malignant
tumors.
Activation of EMT
EMT is the cell transformation from an epithelial to
a mesenchymal cell phenotype (54,55).
It is involved in various physiological and pathological processes,
such as embryogenesis, tissue healing and fibrosis. In particular,
EMT leads to the weakening of cellular junctions, which enhances
cell motility and invasiveness, promoting cancer development.
EMT is regulated by a number of EMT-inducible
transcription factors, such as members of the Zinc finger
E-box-binding homeobox, Snail and Slug families. Various other
signaling pathways can also regulate the EMT process, where Wnt,
TGF-β and Notch are prominent promoters of EMT. In addition, E- and
N-cadherins (epithelial markers) and vimentin (mesenchymal markers)
are important EMT markers. SPOCK1 has been reported to
promote EMT progression in a variety of cancers by upregulating the
expression of N-cadherin, Snail, vimentin and Slug, whilst
downregulating that of E-cadherin (21). However, to the best of our
knowledge, studies on the relationship between other members of the
SPOCK family and EMT remain elusive, which warrants further
study.
SPOCK1 in gastric cancer (GC)
Tumor cell metastasis is a major obstacle to the
treatment of various cancers (56). SPOCK1 has been previously
shown to facilitate cancer metastasis in various cancer types. Chen
et al (57) revealed an
association between elevated SPOCK1 expression and that of
EMT-related markers in GC tissues, metastasis and poor GC
prognosis. In addition, downregulating SPOCK1 expression was
found to significantly inhibit GC-cell invasion and metastasis,
whereas upregulating SPOCK1 expression resulted in the
opposite effect. An investigation into the mechanism of
SPOCK1-induced aggressive metastasis of GC cells
demonstrated that SPOCK1-induced EMT could promote
aggressive metastasis in GC cells, suggesting SPOCK1 to be a
novel therapeutic target for GC.
SPOCK1 in prostate cancer
Prostate cancer is a representative non-skin cancer
and the second major cause of cancer-related mortality among men in
the US. Metastasis is a major cause of mortality (58). Several studies (22,59–61)
have previously evaluated the role of SPOCK1 in prostate
cancer progression. Prostate cancer tissues express substantially
higher quantities of SPOCK1 compared with those in
non-cancerous tissues. In addition, metastatic cells showed
significantly higher SPOCK1 expression compared with that in
non-metastatic cells. SPOCK1 knockdown resulted in cell
cycle arrest at the G0/G1 phase in prostate
cancer cells. Conversely, overexpression of SPOCK1 resulted
in cell cycle arrest in the S phase of human prostate epithelial
cells (RWPE-1). This suggests that SPOCK1 may be involved in
the aberrant division of prostate cancer cells, providing a novel
target for the treatment of prostate cancer and further study in
prostate cancer stem cells. In addition, apoptosis was previously
found to be increased after SPOCK1 expression was
downregulated in PC3 cells, whilst cell migration and invasion were
increased in vitro when SPOCK1 was overexpressed.
Furthermore, metastatic lung nodules in mice were significantly
reduced when SPOCK1-null PC3 cells were injected. These
findings suggest that SPOCK1 is a mediator of prostate
cancer metastasis and malignant growth. Ma et al (62) previously found that the mRNA
expression of SPOCK1 is significantly higher in cancer
tissue compared with that in adjacent lesions. In addition, a
positive association was found between SPOCK1 and Cyclin D1,
c-Myc, vimentin, MMP2 cycle and proliferation indicators. This
indicates that the high expression of SPOCK1 in cancer is
associated with changes in proliferation and the expression of
genes linked to invasion.
SPOCK1 in breast cancer
Fan et al (63) previously revealed that immortalized
breast cancer cells exhibit enhanced invasive abilities after
overexpressing SPOCK1. In addition, another previous study
also assessed the association between SPOCK1 expression and
the clinicopathological features of invasive ductal carcinoma
(64). High SPOCK1
expression was found to be associated with the pathological tumor
size (50). Therefore,
SPOCK1 expression may serve as an independent predictive
marker of poor survival outcomes. The study of SPOCK1
expression in non-mammary invasive ductal carcinoma subtypes has
shown that SPOCK1 expression is abundant in intraductal
carcinoma in situ in the breast and is associated with EMT.
In summary, SPOCK1-targeted therapy is promising and offers
novel ideas for clinical application in patients with breast
cancer.
Effect of SPOCK1 on MMPs
The ECM is a non-cellular matrix that incorporates
various cellular components, including elastin, collagen,
non-collagen, proteoglycans and aminoglycans. Previously, the ECM
was considered to be a static scaffold for cells and tissues.
However, in the proceeding decades, the ECM has been gradually
found to also regulate cell survival, proliferation, polarity,
shape, migration and metabolism, making it an important component
in the tumorigenesis of cancers (65,66).
Matricellular proteins are non-structural ECM molecules that mainly
regulate the interaction between the ECM and cells. They typically
contain different domains that can interact with the ECM, cell
surface receptors, growth factors and cytokines to regulate the
transmission of molecular signals and communication among cells. In
addition, matricellular proteins can also bind a number of
intrinsic enzymes, thereby modulating ECM component synthesis and
degradation. During tissue damage, changes in the matrix
composition can affect tissue regeneration. Therefore, the
alteration of MCPs can influence the occurrence and development of
specific diseases.
MMPs form a family of endopeptidases that degrade
the ECM. They are known to regulate a myriad of physiological
activities and the progression of different diseases, such as
cancer and inflammation, rendering them potential targets for
disease therapy. Previous studies have demonstrated that the
overexpression of SPOCK1 can increase MMP2 and MMP9
expression and activity, which in turn promotes ECM degradation
(67,68).
Activation of the PI3K/Akt signaling
pathway
The PI3K/Akt signaling pathway is of significant
importance in cancer research. This pathway has been demonstrated
to be abnormally activated in various tumors, thereby promoting
occurrence and progression. In addition, PI3K/Akt has been shown to
serve a key role in the regulation of a diverse array of
intracellular processes, including cell cycle regulation, cell
proliferation, apoptosis and migration. In addition, it has been
implicated in the maintenance of tumor stem cells and the
development of drug resistance. In recent years, numerous
inhibitors targeting this pathway have been developed, which
revealed considerable anti-tumor activity in clinical trials. In
particular, the SPOCK1 gene has been demonstrated to induce
tumorigenesis in a variety of cancers by activating the PI3K/Akt
signaling pathway.
SPOCK1 in colorectal cancer
Zhang et al (69) previously discovered that colorectal
cancer tissues exhibit elevated SPOCK1 expression compared
with that in adjacent normal tissues. In addition, high expression
levels of SPOCK1 in patients with colorectal cancer were
found to be associated with tumor size and lymphatic system
metastasis (55). Furthermore,
knocking down SPOCK1 expression was found to restrict cell
proliferation, decrease tumorigenicity and increase cell apoptosis.
SPOCK1 can also promote malignant proliferation by
regulating the PI3K/Akt signaling pathway, suggesting that
SPOCK1 may serve as a potential therapeutic and preventive
target for colorectal cancer.
SPOCK1 in glioblastoma multiforme
Glioblastoma multiforme is one of the most
aggressive forms of human brain malignancies with a dismal
prognosis, owing to its invasive nature and high recurrence rate.
Treatment of recurrent glioblastoma multiforme (RGS) is
particularly challenging due to its resistance to chemotherapy.
SPOCK1 expression was observed to be significantly
upregulated in RGSs, where it can regulate the migration, invasion
and EMT processes. In addition, downregulation of SPOCK1
significantly sensitized these cells to temozolomide, resulting in
a substantial reduction in their aggressiveness and malignancy.
SPOCK1 was also observed to mediate drug resistance in
glioblastoma multiforme by regulating the PI3K/Akt signaling
pathway (27), contributing to a
novel direction for genetic testing for the treatment of advanced
glioblastoma.
SPOCK1 in gallbladder cancer
(GBC)
GBC is one of the leading causes of
cancer-associated mortality worldwide, with a poor prognosis and a
5-year overall survival (OS) rate of only 5% (70). SPOCK1 expression has been
reported to be increased in GBC. Shu et al (71) previously investigated the impact of
SPOCK1 on the progression and prognosis of patients with GBC
and found that SPOCK1 activated the PI3K/Akt signaling
pathway, which inhibited cell apoptosis whilst promoting
proliferation and metastasis. Therefore, patients with GBC may
benefit from targeting SPOCK1 as a prognostic or therapeutic
marker (27).
SPOCK1 in pancreatic cancer
Pancreatic cancer is well-known for its high
mortality rate (72,73). The most significant characteristic
of pancreatic ductal adenocarcinoma is the presence of massive
stromal deposits. Given the complexity of tumor-stromal
interactions in pancreatic ductal adenocarcinoma, identifying
stromal proteins that can promote tumors is essential.
SPOCK1 was discovered as a possible candidate protein. In
addition, SPOCK1 expression is predominantly stromal, where
its high expression can result in poor disease outcomes. Functional
assessment in co-culture assays revealed that SPOCK1 can
potently influence extracellular collagen matrix composition and
promote pancreatic ductal adenocarcinoma proliferation (64,74).
This suggests that elevated SPOCK1 expression is a
contributing factor in pancreatic cancer progression, providing
novel ideas for future drug research and development.
SPOCK1 in osteosarcoma
In osteosarcoma (75,76),
high levels of SPOCK1 expression have been associated with
tumor size, metastasis, staging and pathology, where its
downregulation was found to inhibit osteosarcoma cell proliferation
in vitro and weaken tumorigenicity in nude mice. In
addition, SPOCK1 may promote osteosarcoma cell development
through the mTOR/S6 kinase signaling pathway, suggesting a novel
therapeutic strategy for treating osteosarcoma.
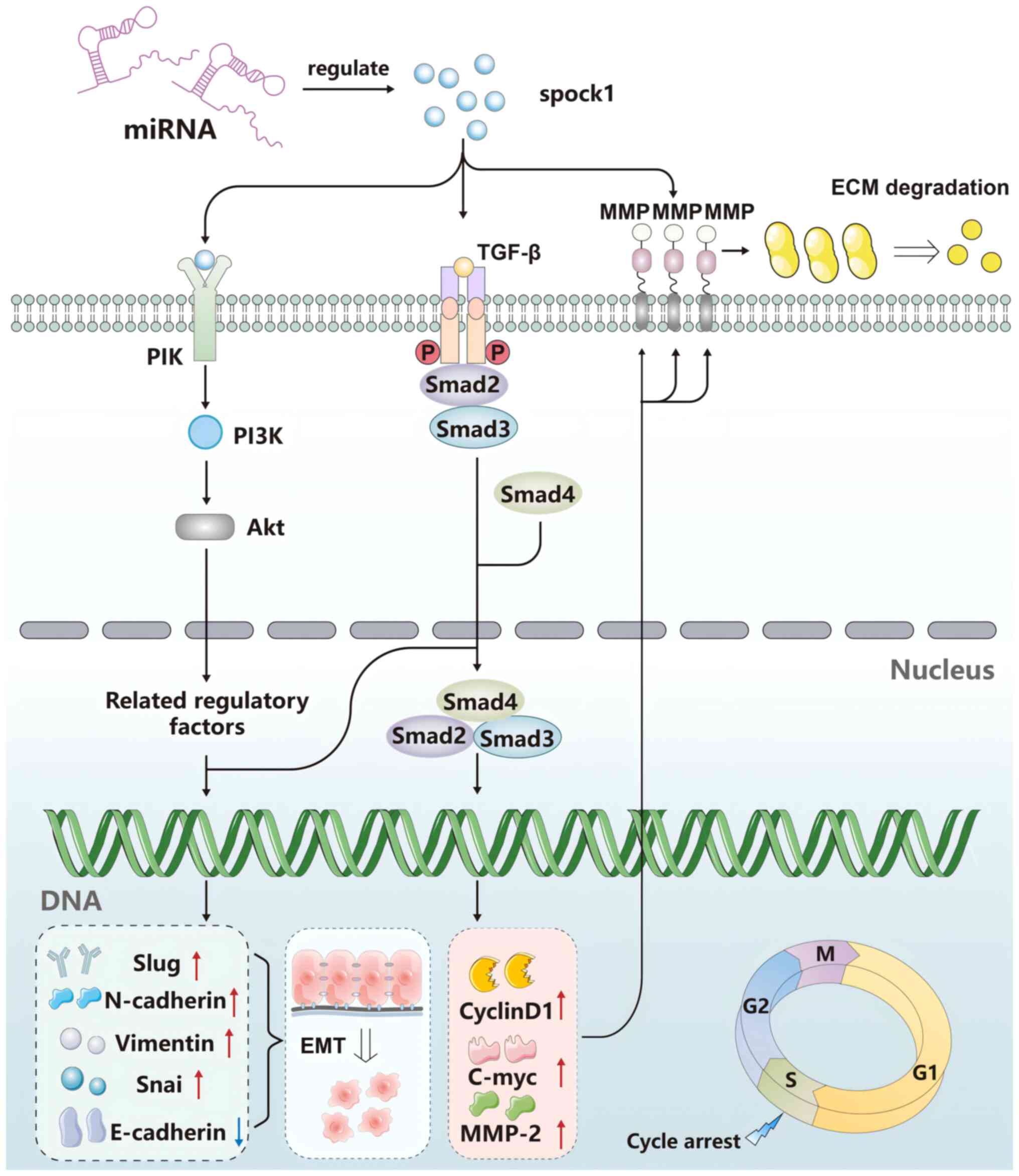
SPOCK1 in hepatocellular carcinoma
(HCC)
Li et al (77) previously examined the SPOCK1
expression profile in 135 pairs of HCC and adjacent paraneoplastic
tissues, revealing that 60% of the HCC samples had increased
expression levels of SPOCK1 mRNA and protein compared with
those in paraneoplastic tissues. In addition, OS and disease-free
survival (DFS) rates decreased significantly when SPOCK1
expression was increased. In terms of the mechanism, SPOCK1
was found to prevent cytochrome c release, and to inhibit
HCC cell apoptosis by activating Akt and enhance MMP9 expression
and activity, rendering HCC cells more invasive (Fig. 2).
SPOCK1 is regulated by numerous
miRNAs
SPOCK1 can serve as a potential target for
numerous miRNAs, where its elevation leads to cancer
deterioration.
SPOCK1 in non-small cell lung cancer
(NSCLC)
Yu et al (78) and Xu et al (79) previously identified SPOCK1
in three cell types using TargetScan and miRDB, two typical online
target gene prediction programs. This previous study investigated
the role of miR-130a-3p and its target gene SPOCK1 in the
etiopathogenesis of tobacco smoke-induced human lung cancer
(78). The results showed that
SPOCK1 expression was high in BEAS-2B cells with unexposed
bronchial epithelium. In addition, significant inhibition of cell
migration was observed in transformed S30 cells exposed to
cigarette smoke, regardless of SPOCK1 silencing. In lung
cancer A549 and H1299 cells transformed using S30, SPOCK1
expression was reduced following miR-130a-3p upregulation,
suggesting that cigarette-transformed cells exhibit a behavior
comparable to lung cancer cells and have a reduced ability to
migrate. In addition, SPOCK1 was previously found to be
overexpressed in human NSCLC cells and tissues. NSCLC cell
proliferation, colony formation, migration and invasion were also
reported to be significantly inhibited by the downregulation of
SPOCK1 expression in vitro. In addition, silencing
SPOCK1 may inhibit Wnt/β-catenin pathway activation
(12,80).
SPOCK1 in head and neck squamous cell
carcinoma (HNSCC)
In HNSCC cells, Koshizuka et al (81) found that SPOCK1 expression
was controlled by miR-150-5p and miR-150-3p. In particular, when
SPOCK1 expression is knocked down in HNSCC cells, they
tended to exhibit a more aggressive behavior. High SPOCK1
expression was then subsequently confirmed in clinical specimens of
HNSCC. According to The Cancer Genome Atlas database, patients
expressing higher SPOCK1 exhibited a substantially shorter
OS rate.
SPOCK1 in esophageal squamous cell
carcinoma (ESCC)
Osako et al (82) previously found significantly
diminished pre-miR-150-5p and miR-150-3p expression levels in ESCC
tissues analyzed using RNA sequencing-based methods. Further
studies into miRNA target genes using a combination of genome-wide
gene expression analysis and database searches subsequently
revealed that SPOCK1 may serve as a candidate target for
miR-150-5p and miR-150-3p in ESCC cells. Luciferase reporter gene
assays provided further evidence demonstrating the direct
modulation of SPOCK1 by these aforementioned miRNAs.
Silencing SPOCK1 expression was then found to suppress
cancer cell migration and invasion. In summary, downregulation of
the pre-miR-150 chain, leading to SPOCK1 overexpression in
ESCC, may be a pathogenic mechanism of this cancer (82,83).
Regulation of SPOCK1 by miRNAs in
HCC
Li et al (77) and Zhu et al (84) detected differentially expressed
miRNAs in HCC to assess the impact of their expression on the
proliferation, invasion and apoptosis in HCC cells, which found
miR-139-5p, miR-940 and miR-193a-5p. The overexpression of
SPOCK1, a common target gene of all three aforementioned
genes in HCC, contributed to a further increase in HCC cell
malignancy, proliferation and invasion, whilst suppressing
apoptosis. Therefore, miR-139-5p, miR-940 and miR-193a-5p may
target SPOCK1 and inhibit HCC development. (Fig. 3).
Role of SPOCK2 in tumor
development
SPOCK2 holds particular promise and it is
becoming a subject of intense research. Initially, it was
considered to serve as a tumor suppressor that is closely
associated with methylation. SPOCK2 does not inhibit
membrane-type matrix metalloproteinases (MT-MMPs) but instead
antagonizes the inhibition of MT-MMPs by other SPOCK family
members. Furthermore, an imbalance in SPOCK2 has been
frequently reported to result in the expression of oncogenes.
SPOCK2 in prostate cancer
Prostate cancer is a common malignancy in men,
where genetic factors and exposure to interventions increase its
risk. According to a previous study by Verma et al (85), SPOCK2 and non-structural
maintenance of chromosomes element 1 homolog (NSE1) can
regulate methylation in prostate cancer. Combining NSE1 and
SPOCK2 hypermethylation was found to increase the ability to
distinguish tumors from normal tissues with 80% sensitivity and 95%
specificity (86). Prostate cancer
tissues were also found to exhibit lower SPOCK2 expression
levels compared with benign prostatic hyperplasia tissues. In
addition, SPOCK2 was observed to suppress the expression of
MT1-MMPs and MMP2s, whilst inhibiting MMP2 activation in DU145 and
LNCaP cells. Upregulation of SPOCK2 can inhibit prostate
cancer cell (DU145 and LNCaP) invasion and migration (87). Another previous study (88) proposed that SPOCK2
methylation may be a candidate biomarker for prostate, colon,
ovarian (89) and breast cancers
compared to normal paracancerous tissues. Further research should
focus on characterizing SPOCK2 as a potential biomarker or a
treatment target for prostate cancer.
SPOCK2 in astrocytoma
Astrocytoma is the most common primary brain
malignancy. It typically occurs in neuroectodermal tumor tissues
and grows rapidly with high degrees of malignancy (90). The most common histological type is
glioblastoma, a highly heterogeneous and invasive malignancy with a
median survival period of 12–15 months after surgery and
chemotherapy (91). Tobey et
al previously analyzed a microarray dataset containing
pediatric and adult astrocytoma samples to screen for
differentially expressed genes. SPOCK2 expression was found
to be downregulated in all tests. Therefore, dysregulation of
SPOCK2 may serve a role in the development of malignant
astrocytomas and represent a novel therapeutic target. However,
further studies are necessary (92).
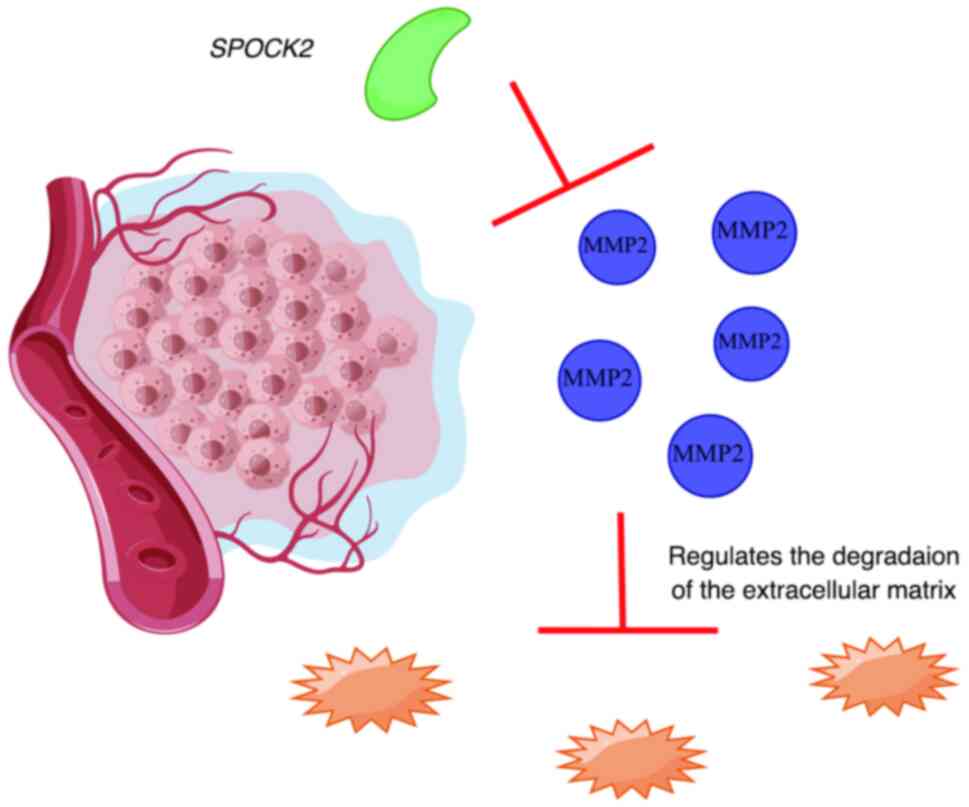
SPOCK2 in endometrial cancer
Endometrial cancer is a representative type of
uterine cancer. Owing to insufficient early symptoms and signs,
lack of testing methods and poor survival rate after late-stage
detection, finding early biomarkers of endometrial cancer is
crucial. Ren et al (93)
have previously found that the SPOCK2 gene may serve as a
biomarker for endometrial cancer. Specifically, these
aforementioned studies concluded that patients with endometrial
cancer have lower SPOCK2 expression compared with that in
healthy individuals. Lower SPOCK2 protein expression levels
were also found to be associated with distant metastasis and
myometrial infiltration. By upregulating SPOCK2, endometrial cancer
cells were observed to exhibit stunted proliferation, invasion and
adhesion, whereas apoptosis was increased. Mechanistically, it
restricted MT1-MMP and MMP2 expression, whilst also inhibiting MMP2
activation (Fig. 4).
SPOCK2 in ovarian cancer
Lou et al (19) previously reported significantly
elevated expression levels of SPOCK2 in ovarian cancer and
suggested that the dysregulation of hsa-miR-363-3p/SPOCK2
may worsen the progression of this cancer. The
hsa-miR-363-3p/SPOCK2 axis is involved in regulating the
actin cytoskeleton. Mechanistically, SPOCK2 may regulate the
actin cytoskeleton, thereby affecting cell adhesion, invasion,
migration and ultimately ovarian cancer progression (94). These extensive studies suggest that
the SPOCK2 gene is not merely a suppressor gene but can also
serve as an oncogene in ovarian cancer, which requires further
exploration.
Role of SPOCK3 in tumor
development
The SPOCK3 gene has not been explored in
depth compared with SPOCK2 and SPOCK1. This may be
due to its similarity with SPOCK1 in terms of structure and
function. By contrast, studies on SPOCK1 have been more
comprehensive, which may partially reflect some of the possible
functions of SPOCK3. The N-Tes splice variant of
SPOCK3 (7,95) was previously found to inhibit
MT1-MMP- and MT3-MMP-mediated MMP2 activation, with the key
sequence located at amino acid residues 33–84 after the N-terminal
signal peptide (96).
Luo et al (30) previously compared tumor and control
tissues of patients with prostate cancer and found a clear positive
association between SPOCK3 expression and DFS of patients
with prostate cancer. In addition, patients with low SPOCK3
expression levels had inferior DFS compared with those with high
SPOCK3 expression levels (44,97).
This suggests that SPOCK3 is a potential prognostic marker
for treatment or prognosis (98).
Similarly, in previous studies on glioma, SPOCK3
downregulation was found to be associated with the inhibition of
glioma cell migration and invasion. SPOCK3 was previously
found to serve an important role in regulating the physiology of
glioma cells (95,99). However, to the best of our
knowledge, relatively few studies have been conducted on the
function of SPOCK3 and the regulatory relationship between
SPOCK3 and malignancies.
Outlook
In summary, the present review summarized existing
research on the expression profile of the most important members of
the SPOCK family, the signaling pathways they were associated with
and their possible roles in a variety of malignancies (Table I). Cancers typically develop in
multiple steps, covering multiple stages, including the activation
of multiple proto-oncogenes and inactivation of tumor suppressor
genes. The specific mechanism underlying the role of the SPOCK
family in the apoptosis, invasion and metastasis of tumor cells
require further investigation. It is hypothesized that the
SPOCK family of genes may regulate apoptosis through the
PI3K/Akt and Wnt/β-catenin signaling pathways in malignant tumors.
This may in turn regulate the EMT process, causing the tumor cells
to either gain or lose their metastatic and invasive abilities on
the one hand, whereas by regulating the activity of Bax and Bad
(pro-apoptotic proteins) on the other hand. However, the SPOCK
subfamily may yet have specific relevance to certain cancers, such
that large-scale fundamental and clinical studies are required
before the SPOCK family can be exploited as a biomarker for
clinical applications. In non-malignant diseases, overexpression of
SPOCK1 may promote stellate cell activation, proliferation
and migration through activation of the integrin α5β1/PI3K/Akt
signaling pathway, thereby enhancing liver fibrosis (100). In bronchopulmonary dysplasia, the
expression of SPOCK2 is gradually upregulated in a
time-dependent manner during the transition from AT2 to AT1 cells,
which can be used as one of the key markers for this transition
(101). Further research on the
SPOCK family may provide a breakthrough in diagnosing and treating
specific malignant diseases in the near future.
 | Table I.Summary of information on the
distribution of the SPOCK family, signaling pathways and associated
diseases. |
Table I.
Summary of information on the
distribution of the SPOCK family, signaling pathways and associated
diseases.
| Item | SPOCK1 | SPOCK2 | SPOCK3 | (Refs.) |
|---|
| Main expression
locations | Brain, prostate,
testis | Brain, lungs,
testicles, kidneys | Brain, ovaries,
prostate | (35–38) |
| Related signaling
pathways | PI3K/Akt signaling
pathway, Wnt/β-catenin signaling pathways, Akt/mTOR signaling
pathway, mTOR-S6K signaling pathway | Unknown | Unknown | (19,28,68,70,79,
103,105–107) |
| Upregulated
expression-related diseases | Liver fibrosis,
liver cancer, stomach cancer, oesophageal cancer, pancreatic
cancer, gallbladder cancer, colorectal cancer, lung cancer,
prostate cancer, urothelial cancer, ovarian cancer, AD brain
tumour, breast cancer, oral submucosal fibrosis, squamous cell
carcinoma of the head, osteosarcoma | BPD, ovarian
cancer | ADHD, BPD | (19,28,35–38,79,103,104,106, 107) |
| Downregulated
expression-related diseases | Unknown | Pancreatic cancer,
lung cancer, prostate cancer, endometrial cancer, brain tumor | Prostate
cancer | (19,30,89,108) |
Acknowledgements
Not applicable.
Funding
The present study was supported by Shenzhen Clinical Medical
Research Center for Otolaryngology Diseases (grant no.
20220819120540004) and Sanming Project of Medicine in Shenzhen
(grant no. SZSM202111016) and the Shenzhen Fund for Guangdong
Provincial High level Clinical Key Specialties (grant no.
SZGSP004).
Availability of data and materials
Not applicable.
Authors' contributions
MX contributed to the study design, literature
search and selection and analysis of the literature/information. JX
and EJ were involved in the writing process, including manuscript
drafting, editing and reviewing, as well as the creation of figures
and tables. All authors have read and approved the final
manuscript. Data authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Carioli G, Malvezzi M, Bertuccio P,
Boffetta P, Levi F, La Vecchia C and Negri E: European cancer
mortality predictions for the year 2021 with focus on pancreatic
and female lung cancer. Ann Oncol. 32:478–487. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dolgin E: Cancer's new normal. Nat Cancer.
2:1248–1250. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cheek DM and Naxerova K: Mapping the long
road to cancer. Cell. 185:939–940. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Siegel RL, Miller KD, Wagle NS and Jemal
A: Cancer statistics, 2023. CA Cancer J Clin. 73:17–48. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bonnet F, Perin JP, Maillet P, Jolles P
and Alliel PM: Characterization of a human seminal plasma
glycosaminoglycan-bearing polypeptide. Biochem J. 288((Pt 2)):
565–569. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Charbonnier F, Périn JP, Roussel G,
Nussbaum JL and Alliel PM: Cloning of testican/SPOCK in man and
mouse. Neuromuscular expression perspectives in pathology. C R
Seances Soc Biol Fil. 191:127–133. 1997.(In French). PubMed/NCBI
|
|
7
|
Hausser HJ, Decking R and Brenner RE:
Testican-1, an inhibitor of pro-MMP-2 activation, is expressed in
cartilage. Osteoarthritis Cartilage. 12:870–877. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Perurena N, Zandueta C, Martinez-Canarias
S, Moreno H, Vicent S, Almeida AS, Guruceaga E, Gomis RR,
Santisteban M, Egeblad M, et al: EPCR promotes breast cancer
progression by altering SPOCK1/testican 1-mediated 3D growth. J
Hematol Oncol. 10:232017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhong J, Krawczyk SA, Chaerkady R, Huang
H, Goel R, Bader JS, Wong GW, Corkey BE and Pandey A: Temporal
profiling of the secretome during adipogenesis in humans. J
Proteome Res. 9:5228–5238. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chiarini A, Onorati F, Marconi M, Pasquali
A, Patuzzo C, Malashicheva A, Irtyega O, Faggian G, Pignatti PF,
Trabetti E, et al: Studies on sporadic non-syndromic thoracic
aortic aneurysms: II. Alterations of extra-cellular matrix
components and focal adhesion proteins. Eur J Prev Cardiol. 25
(Suppl 1):S51–S58. 2018. View Article : Google Scholar
|
|
11
|
Váncza L, Tátrai P, Reszegi A, Baghy K and
Kovalszky I: SPOCK1 with unexpected function. The start of a new
career. Am J Physiol Cell Physiol. 322:C688–c693. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gao Y, Yu M, Ma M, Zhuang Y, Qiu X, Zhao
Q, Dai J, Cai H and Yan X: SPOCK1 contributes to the
third-generation EGFR tyrosine kinase inhibitors resistance in lung
cancer. J Cell Biochem. 120:12566–12573. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Charbonnier F, Périn JP, Mattei MG,
Camuzat A, Bonnet F, Gressin L and Alliel PM: Genomic organization
of the human SPOCK gene and its chromosomal localization to 5q31.
Genomics. 48:377–380. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Alliel PM, Perin JP, Jolles P and Bonnet
FJ: Testican, a multidomain testicular proteoglycan resembling
modulators of cell social behaviour. Eur J Biochem. 214:347–350.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hohenester E, Maurer P, Hohenadl C, Timpl
R, Jansonius JN and Engel J: Structure of a novel extracellular
Ca(2+)-binding module in BM-40. Nat Struct Biol. 3:67–73. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Molina F, Bouanani M, Pau B and Granier C:
Characterization of the type-1 repeat from thyroglobulin, a
cysteine-rich module found in proteins from different families. Eur
J Biochem. 240:125–133. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen TK, Surapaneni AL, Arking DE,
Ballantyne CM, Boerwinkle E, Chen J, Coresh J, Köttgen A, Susztak
K, Tin A, et al: APOL1 Kidney Risk Variants and Proteomics. Clin J
Am Soc Nephrol. 17:684–692. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kamioka M, Imamura J, Komatsu N, Daibata M
and Sugiura T: Testican 3 expression in adult T-cell leukemia. Leuk
Res. 33:913–918. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lou W, Ding B, Zhong G, Du C, Fan W and Fu
P: Dysregulation of pseudogene/lncRNA-hsa-miR-363-3p-SPOCK2 pathway
fuels stage progression of ovarian cancer. Aging (Albany NY).
11:11416–11439. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lal CV and Ambalavanan N: Genetic
predisposition to bronchopulmonary dysplasia. Semin Perinatol.
39:584–591. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ye Z, Chen J, Hu X, Yang S, Xuan Z, Lu X
and Zhao Q: SPOCK1: A multi-domain proteoglycan at the crossroads
of extracellular matrix remodeling and cancer development. Am J
Cancer Res. 10:3127–3137. 2020.PubMed/NCBI
|
|
22
|
Okato A, Arai T, Kojima S, Koshizuka K,
Osako Y, Idichi T, Kurozumi A, Goto Y, Kato M, Naya Y, et al: Dual
strands of pre-miR-150 (miR-150-5p and miR-150-3p) act as antitumor
miRNAs targeting SPOCK1 in naïve and castration-resistant prostate
cancer. Int J Oncol. 51:245–256. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Reichard J and Zimmer-Bensch G: The
epigenome in neurodevelopmental disorders. Front Neurosci.
15:7768092021. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pimentel-Santos FM, Ligeiro D, Matos M,
Mourão AF, Costa J, Santos H, Barcelos A, Godinho F, Pinto P, Cruz
M, et al: Whole blood transcriptional profiling in ankylosing
spondylitis identifies novel candidate genes that might contribute
to the inflammatory and tissue-destructive disease aspects.
Arthritis Res Ther. 13:R572011. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Vannahme C, Schübel S, Herud M, Gösling S,
Hülsmann H, Paulsson M, Hartmann U and Maurer P: Molecular cloning
of testican-2: Defining a novel calcium-binding proteoglycan family
expressed in brain. J Neurochem. 73:12–20. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li D, Farrell JJ, Mez J, Martin ER, Bush
WS, Ruiz A, Boada M, de Rojas I, Mayeux R, Haines JL, et al: Novel
loci for Alzheimer's disease identified by a genome-wide
association study in Ashkenazi Jews. Alzheimers Dement.
19:5550–5562. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yu F, Li G, Gao J, Sun Y, Liu P, Gao H, Li
P, Lei T, Chen Y, Cheng Y, et al: SPOCK1 is upregulated in
recurrent glioblastoma and contributes to metastasis and
Temozolomide resistance. Cell Prolif. 49:195–206. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xu M, Zhang X, Zhang S, Piao J, Yang Y,
Wang X and Lin Z: SPOCK1/SIX1axis promotes breast cancer
progression by activating AKT/mTOR signaling. Aging (Albany NY).
13:1032–1050. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shengxiao X, Xinxin S, Yunxiang Z, Zhijie
T and Xiaofei T: Identification of a basement membrane-related gene
signature for predicting prognosis, immune infiltration, and drug
sensitivity in colorectal cancer. Front Oncol. 14:14281762024.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Luo J, Lai C, Xu X, Shi J, Hu J, Guo K,
Mulati Y, Xiao Y, Kong D, Liu C and Xu K: Mechanism of prognostic
marker SPOCK3 affecting malignant progression of prostate cancer
and construction of prognostic model. BMC Cancer. 23:7412023.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ma D, Fetahu IS, Wang M, Fang R, Li J, Liu
H, Gramyk T, Iwanicki I, Gu S, Xu W, et al: The fusiform gyrus
exhibits an epigenetic signature for Alzheimer's disease. Clin
Epigenetics. 12:1292020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Herreros-Pomares A, Llorens C, Soriano B,
Bagan L, Moreno A, Calabuig-Fariñas S, Jantus-Lewintre E and Bagan
J: Differentially methylated genes in proliferative verrucous
leukoplakia reveal potential malignant biomarkers for oral squamous
cell carcinoma. Oral Oncol. 116:1051912021. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jensen LD, Hot B, Ramsköld D, Germano RFV,
Yokota C, Giatrellis S, Lauschke VM, Hubmacher D, Li MX, Hupe M, et
al: Disruption of the Extracellular Matrix Progressively Impairs
Central Nervous System Vascular Maturation Downstream of β-Catenin
Signaling. Arterioscler Thromb Vasc Biol. 39:1432–1447. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chai T, Tian M, Yang X, Qiu Z, Lin X and
Chen L: Genome-Wide identification of associations of circulating
molecules with spontaneous coronary artery dissection and aortic
aneurysm and dissection. Front Cardiovasc Med. 9:8749122022.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Martinez JR, Dhawan A and Farach-Carson
MC: Modular Proteoglycan Perlecan/HSPG2: Mutations, phenotypes, and
functions. Genes (Basel). 9:5562018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gordts PLSM and Esko JD: The heparan
sulfate proteoglycan grip on hyperlipidemia and atherosclerosis.
Matrix Biol. 71–72. 262–282. 2018.PubMed/NCBI
|
|
37
|
Pintér A, Hevesi Z, Zahola P, Alpár A and
Hanics J: Chondroitin sulfate proteoglycan-5 forms perisynaptic
matrix assemblies in the adult rat cortex. Cell Signal.
74:1097102020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Schiffer D, Mellai M, Boldorini R, Bisogno
I, Grifoni S, Corona C, Bertero L, Cassoni P, Casalone C and
Annovazzi L: The significance of chondroitin sulfate proteoglycan 4
(CSPG4) in human gliomas. Int J Mol Sci. 19:27242018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Swenarchuk LE: Nerve, Muscle, and
Synaptogenesis. Cells. 8:14482019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gridley DS, Mao XW, Tian J, Cao JD, Perez
C, Stodieck LS, Ferguson VL, Bateman TA and Pecaut MJ: Genetic and
apoptotic changes in lungs of mice flown on the STS-135 mission in
space. In vivo. 29:423–433. 2015.PubMed/NCBI
|
|
41
|
Zhang L, An XP, Liu XR, Fu MZ, Han P, Peng
JY, Hou JX, Zhou ZQ, Cao BY and Song YX: Characterization of the
transcriptional complexity of the receptive and pre-receptive
endometria of dairy goats. Sci Rep. 5:142442015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hadchouel A, Durrmeyer X, Bouzigon E,
Incitti R, Huusko J, Jarreau PH, Lenclen R, Demenais F,
Franco-Montoya ML, Layouni I, et al: Identification of SPOCK2 as a
susceptibility gene for bronchopulmonary dysplasia. Am J Respir
Crit Care Med. 184:1164–1170. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Buga AM, Scholz CJ, Kumar S, Herndon JG,
Alexandru D, Cojocaru GR, Dandekar T and Popa-Wagner A:
Identification of new therapeutic targets by genome-wide analysis
of gene expression in the ipsilateral cortex of aged rats after
stroke. PLoS One. 7:e509852012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Weber H, Scholz CJ, Jacob CP, Heupel J,
Kittel-Schneider S, Erhardt A, Hempel S, Schmidt B, Kiel T, Gessner
A, et al: SPOCK3, a risk gene for adult ADHD and personality
disorders. Eur Arch Psychiatry Clin Neurosci. 264:409–421. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Karcher NR, Rogers BP and Woodward ND:
Functional connectivity of the striatum in schizophrenia and
psychotic bipolar disorder. Biol Psychiatry Cogn Neurosci
Neuroimaging. 4:956–965. 2019.PubMed/NCBI
|
|
46
|
Walker WH II, Walton JC, DeVries AC and
Nelson RJ: Circadian rhythm disruption and mental health. Transl
Psychiatry. 10:282020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Haukvik UK, Tamnes CK, Soderman E and
Agartz I: Neuroimaging hippocampal subfields in schizophrenia and
bipolar disorder: A systematic review and meta-analysis. J
Psychiatr Res. 104:217–226. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Weissflog L, Scholz CJ, Jacob CP, Nguyen
TT, Zamzow K, Gross-Lesch S, Renner TJ, Romanos M, Rujescu D,
Walitza S, et al: KCNIP4 as a candidate gene for personality
disorders and adult ADHD. Eur Neuropsychopharmacol. 23:436–447.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Bocock JP, Edgell CJ, Marr HS and Erickson
AH: Human proteoglycan testican-1 inhibits the lysosomal cysteine
protease cathepsin L. Eur J Biochem. 270:4008–4015. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
van de Merbel AF, van der Horst G, Buijs
JT and van der Pluijm G: Protocols for Migration and Invasion
Studies in Prostate Cancer. Methods Mol Biol. 1786:67–79. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Morillo-Bernal J, Fernandez LP and
Santisteban P: FOXE1 regulates migration and invasion in thyroid
cancer cells and targets ZEB1. Endocr Relat Cancer. 27:137–151.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Pal A, Haliti P, Dharmadhikari B, Qi W and
Patra P: Manipulating extracellular matrix organizations and
parameters to control local cancer invasion. IEEE/ACM Trans Comput
Biol Bioinform. 18:2566–2576. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Quan Q, Wang X, Lu C, Ma W, Han J, Xia G,
Yang G and Wang C: Association of extracellular matrix
microarchitecture and three-dimensional collective invasion of
cancer cells. Biotech Histochem. 95:605–612. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Bakir B, Chiarella AM, Pitarresi JR and
Rustgi AK: EMT, MET, plasticity, and tumor metastasis. Trends Cell
Biol. 30:764–776. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Pastushenko I and Blanpain C: EMT
transition states during tumor progression and metastasis. Trends
Cell Biol. 29:212–226. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Ohtsu A: Current status and future
prospects of chemotherapy for metastatic gastric cancer: A review.
Gastric Cancer. 8:95–102. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Chen D, Zhou H, Liu G, Zhao Y, Cao G and
Liu Q: SPOCK1 promotes the invasion and metastasis of gastric
cancer through Slug-induced epithelial-mesenchymal transition. J
Cell Mol Med. 22:797–807. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Ge R, Wang Z, Montironi R, Jiang Z, Cheng
M, Santoni M, Huang K, Massari F, Lu X, Cimadamore A, et al:
Epigenetic modulations and lineage plasticity in advanced prostate
cancer. Ann Oncol. 31:470–479. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Chen Q, Yao YT, Xu H, Chen YB, Gu M, Cai
ZK and Wang Z: SPOCK1 promotes tumor growth and metastasis in human
prostate cancer. Drug Des Devel Ther. 10:2311–2321. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Chien MH, Lin YW, Wen YC, Yang YC, Hsiao
M, Chang JL, Huang HC and Lee WJ: Targeting the SPOCK1-snail/slug
axis-mediated epithelial-to-mesenchymal transition by apigenin
contributes to repression of prostate cancer metastasis. J Exp Clin
Cancer Res. 38:2462019. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Chen ML, Ho CJ, Yeh CM, Chen SL, Sung WW,
Wang SC and Chen CJ: High SPOCK1 expression is associated with
advanced stage, T value, and gleason grade in prostate cancer.
Medicina (Kaunas). 55:3432019. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Ma LJ, Wu WJ, Wang YH, Wu TF, Liang PI,
Chang IW, He HL and Li CF: SPOCK1 overexpression confers a poor
prognosis in urothelial carcinoma. J Cancer. 7:467–476. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Fan LC, Jeng YM, Lu YT and Lien HC: SPOCK1
Is a novel transforming growth factor-β-induced myoepithelial
marker that enhances invasion and correlates with poor prognosis in
breast cancer. PLoS One. 11:e01629332016. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Veenstra VL, Damhofer H, Waasdorp C,
Steins A, Kocher HM, Medema JP, ven Laarhoven HW and Bijlsma MF:
Stromal SPOCK1 supports invasive pancreatic cancer growth. Mol
Oncol. 11:1050–1064. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Roberts DD: Emerging functions of
matricellular proteins. Cell Mol Life Sci. 68:3133–3136. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Gerarduzzi C, Hartmann U, Leask A and
Drobetsky E: The matrix revolution: Matricellular proteins and
restructuring of the cancer microenvironment. Cancer Res.
80:2705–2717. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Yang J, Yang Q, Yu J, Li X, Yu S and Zhang
X: SPOCK1 promotes the proliferation, migration and invasion of
glioma cells through PI3K/AKT and Wnt/β-catenin signaling pathways.
Oncol Rep. 35:3566–3576. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Li Y, Chen L, Chan TH, Liu M, Kong KL, Qiu
JL, Li Y, Yuan YF and Guan XY: SPOCK1 is regulated by CHD1L and
blocks apoptosis and promotes HCC cell invasiveness and metastasis
in mice. Gastroenterology. 144:179–191.e4. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Zhang J, Zhi X, Shi S, Tao R, Chen P, Sun
S, Bian L, Xu Z and Ma L: SPOCK1 is up-regulated and promotes tumor
growth via the PI3K/AKT signaling pathway in colorectal cancer.
Biochem Biophys Res Commun. 482:870–876. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Kong L, Wu Q, Zhao L, Ye J, Li N and Yang
H: Identification of messenger and long noncoding RNAs associated
with gallbladder cancer via gene expression profile analysis. J
Cell Biochem. 120:19377–19387. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Shu YJ, Weng H, Ye YY, Hu YP, Bao RF, Cao
Y, Cao Y, Wang XA, Zhang F, Xiang SS, et al: SPOCK1 as a potential
cancer prognostic marker promotes the proliferation and metastasis
of gallbladder cancer cells by activating the PI3K/AKT pathway. Mol
Cancer. 14:122015. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Vincent A, Herman J, Schulick R, Hruban RH
and Goggins M: Pancreatic cancer. Lancet. 378:607–620. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Ilic M and Ilic I: Epidemiology of
pancreatic cancer. World J Gastroenterol. 22:9694–9705. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Li J, Ke J, Fang J and Chen JP: A
potential prognostic marker and therapeutic target: SPOCK1 promotes
the proliferation, metastasis, and apoptosis of pancreatic ductal
adenocarcinoma cells. J Cell Biochem. 121:743–754. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Chen C, Xie L, Ren T, Huang Y, Xu J and
Guo W: Immunotherapy for osteosarcoma: Fundamental mechanism,
rationale, and recent breakthroughs. Cancer Lett. 500:1–10. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Ritter J and Bielack SS: Osteosarcoma. Ann
Oncol. 21 Suppl 7:vii320–vii325. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Li P, Xiao Z, Luo J, Zhang Y and Lin L:
MiR-139-5p, miR-940 and miR-193a-5p inhibit the growth of
hepatocellular carcinoma by targeting SPOCK1. J Cell Mol Med.
23:2475–2488. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Yu XF, Wang J, OUYang N, Guo S, Sun H,
Tong J, Chen T and Li J: The role of miR-130a-3p and SPOCK1 in
tobacco exposed bronchial epithelial BEAS-2B transformed cells:
Comparison to A549 and H1299 lung cancer cell lines. J Toxicol
Environ Health A. 82:862–869. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Xu Y, Zhao P, Xu X, Zhang S, Xia B and Zhu
L: T790M mutation sensitizes non-small cell lung cancer cells to
radiation via suppressing SPOCK1. Biochem Biophys Rep.
38:1017292024.PubMed/NCBI
|
|
80
|
Wang T, Liu X, Tian Q, Liang T and Chang
P: Reduced SPOCK1 expression inhibits non-small cell lung cancer
cell proliferation and migration through Wnt/β-catenin signaling.
Eur Rev Med Pharmacol Sci. 22:637–644. 2018.PubMed/NCBI
|
|
81
|
Koshizuka K, Hanazawa T, Kikkawa N, Katada
K, Okato A, Arai T, Idichi T, Osako Y, Okamoto Y and Seki N:
Antitumor miR-150-5p and miR-150-3p inhibit cancer cell
aggressiveness by targeting SPOCK1 in head and neck squamous cell
carcinoma. Auris Nasus Larynx. 45:854–865. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Osako Y, Seki N, Koshizuka K, Okato A,
Idichi T, Arai T, Omoto I, Sasaki K, Uchikado Y, Kita Y, et al:
Regulation of SPOCK1 by dual strands of pre-miR-150 inhibit cancer
cell migration and invasion in esophageal squamous cell carcinoma.
J Hum Genet. 62:935–944. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Song X, Han P, Liu J, Wang Y, Li D, He J,
Gong J, Li M, Tu W, Yan W, et al: Up-regulation of SPOCK1 induces
epithelial-mesenchymal transition and promotes migration and
invasion in esophageal squamous cell carcinoma. J Mol Histol.
46:347–356. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Zhu X, Jiang S, Wu Z, Liu T, Zhang W, Wu
L, Xu L and Shao M: Long non-coding RNA TTN antisense RNA 1
facilitates hepatocellular carcinoma progression via regulating
miR-139-5p/SPOCK1 axis. Bioengineered. 12:578–588. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Verma M, Patel P and Verma M: Biomarkers
in prostate cancer epidemiology. Cancers (Basel). 3:3773–3798.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Sambuudash O, Kim HS and Cho MY: Lack of
aberrant methylation in an adjacent area of left-sided colorectal
cancer. Yonsei Med J. 58:749–755. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Liu G, Ren F and Song Y: Upregulation of
SPOCK2 inhibits the invasion and migration of prostate cancer cells
by regulating the MT1-MMP/MMP2 pathway. PeerJ. 7:e71632019.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Chang AJ, Autio KA, Roach M III and Scher
HI: High-risk prostate cancer-classification and therapy. Nat Rev
Clin Oncol. 11:308–323. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Ren F, Wang DB, Li T, Chen YH and Li Y:
Identification of differentially methylated genes in the malignant
transformation of ovarian endometriosis. J Ovarian Res. 7:732014.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Taphoorn MJB, Dirven L, Kanner AA,
Lavy-Shahaf G, Weinberg U, Taillibert S, Toms SA, Honnorat J, Chen
TC, Sroubek J, et al: Influence of treatment with tumor-treating
fields on health-related quality of life of patients with newly
diagnosed glioblastoma: A secondary analysis of a randomized
clinical trial. JAMA Oncol. 4:495–504. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Wessels PH, Weber WE, Raven G, Ramaekers
FC, Hopman AH and Twijnstra A: Supratentorial grade II astrocytoma:
Biological features and clinical course. Lancet Neurol. 2:395–403.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
MacDonald TJ, Pollack IF, Okada H,
Bhattacharya S and Lyons-Weiler J: Progression-associated genes in
astrocytoma identified by novel microarray gene expression data
reanalysis. Methods Mol Biol. 377:203–222. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Ren F, Wang D, Wang Y, Chen P and Guo C:
SPOCK2 affects the biological behavior of endometrial cancer cells
by regulation of MT1-MMP and MMP2. Reprod Sci. 27:1391–1399. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
94
|
She S, Yang M, Hu H, Hu P, Yang Y and Ren
H: Proteomics based identification of autotaxin as an
anti-hepatitis B virus factor and a promoter of hepatoma cell
invasion and migration. Cell Physiol Biochem. 45:744–760. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Nakada M, Yamada A, Takino T, Miyamori H,
Takahashi T, Yamashita J and Sato H: Suppression of membrane-type 1
matrix metalloproteinase (MMP)-mediated MMP-2 activation and tumor
invasion by testican 3 and its splicing variant gene product,
N-Tes. Cancer Res. 61:8896–8902. 2001.PubMed/NCBI
|
|
96
|
Nakada M, Miyamori H, Yamashita J and Sato
H: Testican 2 abrogates inhibition of membrane-type matrix
metalloproteinases by other testican family proteins. Cancer Res.
63:3364–3369. 2003.PubMed/NCBI
|
|
97
|
Yu G, Tang Z, Chen H, Chen Z, Wang L, Cao
H, Wang G, Xing J, Shen H, Cheng Q, et al: Long-term exposure to 4G
smartphone radiofrequency electromagnetic radiation diminished male
reproductive potential by directly disrupting Spock3-MMP2-BTB axis
in the testes of adult rats. Sci Total Environ. 698:1338602020.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Wang LY, Cui JJ, Zhu T, Shao WH, Zhao Y,
Wang S, Zhang YP, Wu JC and Zhang L: Biomarkers identified for
prostate cancer patients through genome-scale screening.
Oncotarget. 8:92055–92063. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Lee SJ, Kim SJ, Seo HH, Shin SP, Kim D,
Park CS, Kim KT, Kim YH, Jeong JS and Kim IH: Over-expression of
miR-145 enhances the effectiveness of HSVtk gene therapy for
malignant glioma. Cancer Lett. 320:72–80. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Du Z, Lin Z, Wang Z, Liu D, Tian D and Xia
L: SPOCK1 overexpression induced by platelet-derived growth
factor-BB promotes hepatic stellate cell activation and liver
fibrosis through the integrin α5β1/PI3K/Akt signaling pathway. Lab
Invest. 100:1042–1056. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Morales Johansson H, Newman DR and Sannes
PL: Whole-genome analysis of temporal gene expression during early
transdifferentiation of human lung alveolar epithelial type 2 cells
in vitro. PLoS One. 9:e934132014. View Article : Google Scholar : PubMed/NCBI
|