Introduction
Inflammation is a broad systemic physiological
response of the human body to various entities, including
pathogens, dust particles and viruses (1). There are essentially two main types
of inflammation: Acute and chronic. Acute inflammation is a
temporary process that lasts from minutes to days, and is an
important component of the immune response (2). Its main role is to deliver leukocytes
and plasma mediators to the site of injury to eliminate
inflammatory stimuli (2). However,
because these leukocytes and associated plasma mediators fail to
discriminate between microbial and host targets, it inevitably
results in collateral host tissue damage (3,4).
Chronic inflammation results from long-term stimulation by
inflammatory agents, represented by a protracted course, usually
measured from months to years. Chronic inflammation generally
occurs when acute responses fail to eliminate pathogenic agents or
repair tissue damage at the site of inflammation, or in response to
stimuli that provoke low-intensity, asymptomatic reactions
(2). In contrast to acute
inflammation, chronic inflammation promotes angiogenesis, fibrosis
and tissue destruction at the affected site (5). Evidence is growing that inflammation
serves a key role in promoting the development of a set of chronic
diseases (1).
With a better understanding of the aetiologies of
various diseases, inflammation has become increasingly recognized
as having a critical role in the development and continuation of a
number of diseases, such as atherosclerosis, chronic liver disease
and sepsis. No method has been developed to date that can delay or
stop the progression of inflammation effectively (6). Anti-inflammatory therapies have been
widely applied to treat various diseases; although they are
effective when used to treat primary inflammatory dysregulation and
autoimmune diseases, they have notable limitations, including
osteoporosis caused by long-term glucocorticoids and
gastrointestinal injury induced by non-steroidal anti-inflammatory
drugs (7). Thus, research has been
dedicated to identifying more effective therapeutic targets for
anti-inflammatory interventions.
Stabilin-1, also referred to as Feel-1=, belongs to
the scavenger receptor class H family. As a large multidomain
molecule, Stabilin-1 contains multiple atypical epidermal growth
factor (EGF)-like repeats; tandem fasciclin-like domains; and a
membrane-proximal link module, a conserved C-type lectin-like
domain that binds to matrix glycosaminoglycans, hyaluronic acid
(HA) and chondroitin sulphate (8).
Stabilin-1 expression occurs in a highly tissue-specific manner;
under homeostatic conditions, Stabilin-1 is highly expressed by
discontinuous sinusoidal endothelial cells (SECs) of the spleen,
lymph nodes and liver, whereas expression in conventional vascular
endothelial cells is induced only under angiogenic and
proinflammatory stimuli (9,10).
Stabilin-1 is also expressed by certain tissue-resident macrophages
and M2-like monocyte subsets (11,12).
Stabilin-1 serves a notable role in various diseases
related to inflammation, such as Listeria monocytogenes (Lm)
infection and chronic liver disease. For example, in the case of Lm
infection, Stabilin-1 protects the host by facilitating macrophage
phagocytosis of Lm, and modulating the production of cytokines such
as TNF-α, IL-6 and IL-10, as well as chemokines including CCL7 and
CXCL10 (13). In a study on
atherosclerosis, mice with upregulated Stabilin-1 exhibited
markedly reduced aortic plaque formation with no readily apparent
side effects, such as liver fibrosis or renal injury (14). Moreover, in studies on diseases
including sepsis and chronic liver disease, the level of Stabilin-1
has been shown to strongly affect disease progression and prognosis
(15,16). Thus, Stabilin-1 may be considered a
novel therapeutic target for inflammation-related diseases.
The present study collected and organized relevant
literature up to December 2025 by searching databases such as
Pubmed (https://pubmed.ncbi.nlm.nih.gov/), Embase (https://www.embase.com/), Cochrane (https://www.cochranelibrary.com/), CINAHL
(https://search.ebscohost.com/) and Web
of Science (https://www.webofscience.com/), using core keywords
and snowball method The search strategy employed a combination of
medical subject headings and free-text terms: ‘Stabilin-1’ or
‘Clever-1’ or ‘Feel-1’, alongside ‘inflammation’, ‘inflammatory
response’, ‘cytokines’, ‘tissue homeostasis’, ‘receptor’,
‘scavenger’, ‘macrophage’, ‘specific immunity’, ‘cancer’, ‘tumour’,
‘atherosclerosis’, ‘viral myocarditis’, ‘hepatitis’, ‘sepsis’ and
‘Listeria infection’. The inclusion criteria were as
follows: i) The type of literature included basic experimental
research, preclinical research, literature reviews, meta-analyses
and case reports (based on clinical cases of Stabilin-1 with
irregular expression and inflammation-related diseases) and was not
limited by language restrictions; ii) The research clearly
elaborated on the molecular structure, distribution of expression
and signal transduction pathways of Stabilin-1, or directly
investigated the regulatory role of Stabilin-1 in the generation,
development and reduction of inflammation or certain diseases, or
the mechanisms by which Stabilin-1 participates in the maintenance
of tissue homeostasis (such as tissue repair, immune cells and
metabolic homeostasis); iii) The research contained complete data,
with clear conclusions, providing direct evidence to support the
relationships between Stabilin-1 and inflammation or tissue
homeostasis. The exclusion criteria were as follows: i) Research
literature that did not involve Stabilin-1, studies that only
described the gene names or protein expression, and did not explore
the mechanism of action; ii) duplicate publications (in this case,
the latest, most complete version was selected); and iii) abstract
literature published for conferences, dissertations without
complete research data and literature that had direct discrepancies
in conclusions that were not supported by any valid literature, or
non-research literature.
At present, research mainly focuses on the function
of Stabilin-1 as a scavenger receptor. Park et al (17) reported that, in acidic
environments, Ets-2/JNK signalling can promote Stabilin-1
expression to improve macrophage phagocytic ability. With the
advancement of research, Park and Kim (18) further indicated that Stabilin-1, as
a receptor for phosphatidylserine (PS), is involved in the
phagocytosis of apoptotic cells. Research on the pathological
importance of Stabilin-1 has mainly focussed on cancer. Gurung
et al (19) elucidated the
immunosuppressive effect of Stabilin-1 on tumour-associated
macrophages (TAMs) and its potential as a cancer immunotherapy
target. Existing research has been limited to a single mechanism or
disease of Stabilin-1. Therefore, the present review aims to
systematically expand the research perspective and scope. To the
best of our knowledge, based on the existing research, the present
review is the first to construct a biological mechanism framework
of ‘Stabilin-1 > innate immunity + adaptive immunity >
multi-disease regulation’. In terms of research scope, aside from
not being limited to the scavenger activity of Stabilin-1, the
present study also comprehensively analyses the multi-link
regulatory functions of Stabilin-1 in non-specific and specific
immunity. Moreover, in terms of pathological importance, aside from
atherosclerosis, viral myocarditis, chronic hepatitis, sepsis,
Listeria infection and cancer, the review systematically
summarizes the diversity of the functional roles of Stabilin-1, as
well as their common regulatory mechanism in different diseases.
Finally, aside from proposing verifiable research hypotheses for
important scientific questions that have not yet been answered in
this research area, the present review may fill a gap in the
research that exists between basic mechanistic studies and clinical
translation applications, providing a more systematic and
comprehensive theoretical basis for in-depth studies on Stabilin-1,
as well as for developing targeted therapeutic strategies for
related diseases.
Innate immunity
Regulation of monocyte/macrophage
activation and recruitment by Stabilin-1
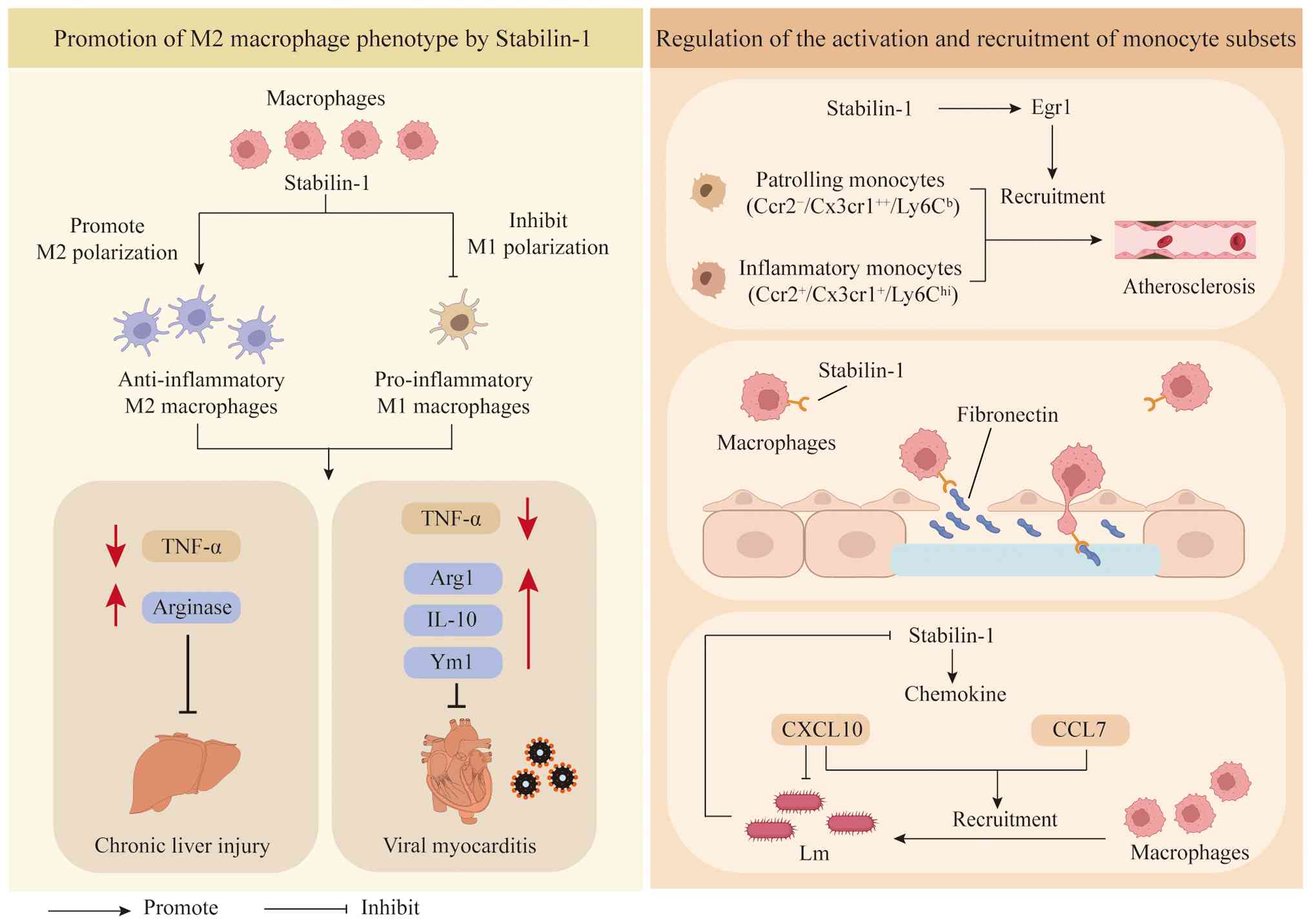
Promotion of the M2 macrophage
phenotype by Stabilin-1
The recruitment and activation of macrophages are
closely associated with the initiation and progression of
inflammation. Macrophages can be divided into M1 (classically
activated) and M2 (alternatively activated) types (20). M1 macrophages exhibit high
expression of IL-12 and IL-23, low expression of IL-10, and
generate reactive oxygen compounds, nitrogen compounds and
inflammatory cytokines, such as TNF-α and IL-6 (21). M1 macrophages are key contributors
to the T-helper cell (Th)1 immune response, and they also exhibit
strong microbicidal and tumoricidal activity (22). On the other hand, M2 macrophages
exhibit low expression of IL-12 and IL-23, and high expression of
IL-10 (21). M2 macrophages are
key components in the processes of tissue repair and regeneration,
inflammation resolution, apoptotic cell clearance and elimination
of extracellular parasites (22,23).
Macrophages expressing Stabilin-1 display a phenotypic bias towards
the M2 subset (24). When
polarization occurs towards the M2 phenotype occurs, human
monocytes maintain surface expression of Stabilin-1, whereas they
lose Stabilin-1 expression upon M1 induction, a finding consistent
with the functional immunosuppressive role of Stabilin-1 (25) (Fig.
1).
Stabilin-1 regulates the activation and
recruitment of monocyte subsets
Single-cell RNA sequencing has revealed that
Stabilin-1 deficiency can markedly alter the transcriptomic
characteristics of circulating monocytes (14) (Fig.
1). Stabilin-1 knockout (KO) in mice has been reported to
result in the dysregulated expression of 231 proteins in the
plasma, among which 41 proteins were shown to differ across
ApoE-KO, ApoE-Stabilin-1-KO and ApoE-Stabilin-2-KO mice (14). The interaction between Stabilin-1
and the extra-domain A (EDA) of fibronectin is a key mechanism
through which monocytes are recruited to the site of inflammation
(26) (Fig. 1). Stabilin-1 directly binds to
fibronectin through its extracellular fascicular domain (P9
fragment). In addition, chemokines are key signalling molecules
that regulate immune cell migration. At the level of chemokine
expression, Stabilin-1 upregulates the expression of chemokines
such as CCL2, CXCL10 and CCL7 (13). CXCL10 not only mediates immune cell
migration but also directly reduces Lm activity, and its
upregulation may further increase the anti-infective capacity of
the host (27). As a ligand for
the CCR2 receptor, CCL7 is indispensable for monocyte recruitment
during inflammation (28). Thus,
Stabilin-1 may mediate myeloid cell recruitment by regulating the
expression of CCL7 and CXCL10 (Fig.
1).
Scavenger function of Stabilin-1 in
macrophages
Stabilin-1 mediates the
internalization of extracellular secreted protein acidic and rich
in cysteine (SPARC) and its transport to lysosomes
SPARC is an evolutionarily highly conserved protein
with a molecular weight of 43 kDa (29), which is produced by endothelial
cells, fibroblasts and macrophages during wound healing, foreign
body responses and angiogenesis (30–32).
SPARC is involved in collagen mineralization, wound healing and
organ fibrosis (33), and it
interacts with various extracellular matrix (ECM) proteins,
regulating their functions and affecting the composition of the ECM
(34,35). Specifically, it stimulates TGF-β,
leading to increased synthesis of type I collagen (36). Furthermore, SPARC inhibits the
pro-angiogenic pathway (37). The
ablation of SPARC expression has been shown to diminish apoptosis
and inflammation (38).
Stabilin-1 mediates the effective uptake and
intracellular delivery of SPARC by M2 macrophages, and acetylated
low-density lipoprotein (acLDL) competes with SPARC for efficient
internalization mediated by Stabilin-1 (38) (Fig.
2). In addition, both acLDL and SPARC follow the endocytic
pathway into lysosomes (38).
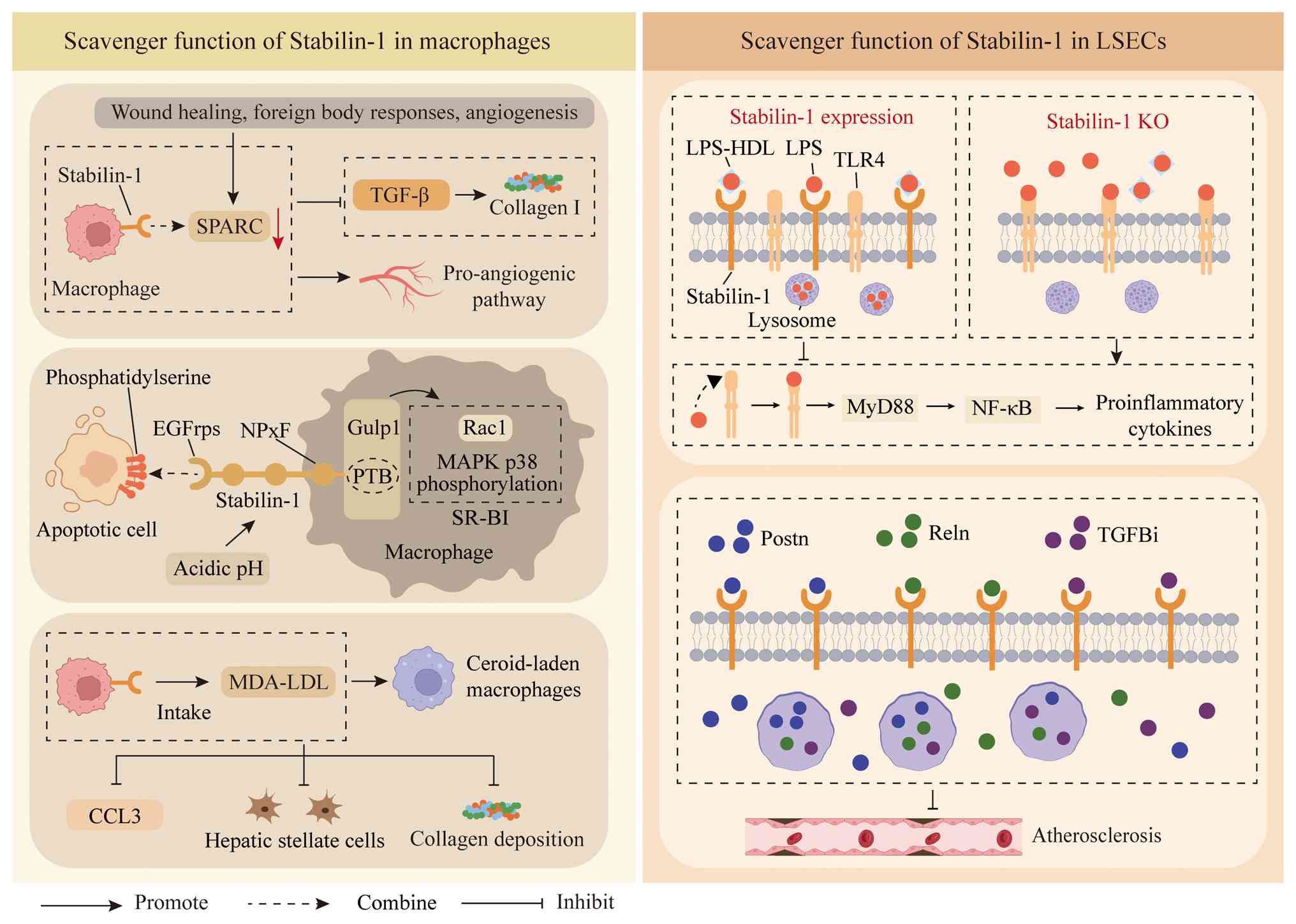 | Figure 2.Scavenger function of Stabilin-1 in
macrophages and LSECs. Stabilin-1 mediates the effective uptake and
intracellular delivery of SPARC by M2 macrophages. Stabilin-1
directly binds to phosphatidylserine through its EGFRps, thereby
mediating macrophage clearance of apoptotic cells. Acidic pH values
can also upregulate the expression of Stabilin-1. Stabilin-1 can
specifically recognize oxidative stress products, such as MDA-LDL,
form waxy loaded macrophages and thereby inhibit the release of the
proinflammatory chemokine CCL3. Normal expression of Stabilin-1
inhibits the TLR4 signalling pathway by mediating LPS clearance.
Stabilin-1 reduces the risk of atherosclerosis by mediating the
endocytosis of circulating ligands such as Postn, Reln and TGFBi.
The ‘combine’ arrow refers to the relationship in which two
substances at both ends of the arrow are combined together. EGFRp,
EGF-like repeat domain; HDL, high-density lipoprotein; KO,
knockout; LPS, lipopolysaccharide; LSEC, liver sinusoidal
endothelial cell; MDA-LDL, malondialdehyde-modified low-density
lipoprotein; MyD88, myeloid differentiation factor 88; NPxF,
asparagine-proline-x-phenylalanine; Postn, Periostin; PTB,
phosphotyrosine-binding; Reln, Reelin; SPARC, secreted protein
acidic and rich in cysteine; SR-BI, scavenger receptor class B
member 1; TGFBi, TGF-β-induced protein; TLR4, Toll-like receptor
4. |
Stabilin-1 mediates the clearance of
apoptotic cells by macrophages
Macrophage clearance of apoptotic cells is important
for peripheral tolerance during homeostasis in healthy tissues and
serves a crucial role in reducing inflammation (39). Dying cells release ‘find-me’
signals to activate phagocytes. Then, phagocytes distinguish
apoptotic cells from healthy viable cells via specific phagocytic
receptors that recognize ‘eat-me’ signals on dying cells.
Phagocytes undergo extensive cytoskeletal rearrangement to
internalize apoptotic cells. The ingested cargo is processed and
elicits specific phagocyte responses, predominantly the release of
anti-inflammatory mediators.
To date, PS is the best-studied ‘eat-me’ signal and
has been well characterized (40);
during apoptosis, PS is present on the outer surface of the cell
membrane and acts as an ‘eat-me’ signal (41). PS receptors can directly bind to PS
on apoptotic cells, including TIM, Bai1, Stabilin-1 receptors and
CD300 receptors. In addition, other receptors can indirectly bind
to apoptotic cells through soluble PS binding proteins (such as
Mfge8 and Gas6), including Tyro3/Axl/Mer receptors (Tyro3, Axl, and
Mer) and integrin receptors (αvβ3 and αvβ5) (42,43).
Under physiological conditions, macrophages bind to apoptotic
cells, PS is recognized by specific receptors, and apoptotic cells
are cleared by macrophages. This process induces macrophages to
secrete anti-inflammatory mediators and downregulates the
production of proinflammatory factors (44).
The EGF-like domains of Stabilin-1 function as
recognition receptors for PS (45). It has been shown that the ectopic
expression of Stabilin-1 endows mouse fibroblast L cells with the
ability to phagocytose damaged red blood cells in a PS-dependent
manner (46). In macrophages
cocultured with apoptotic cells, Stabilin-1 is recruited to the
sites of apoptotic cell recognition and phagocytosis, and
colocalizes with ingested apoptotic bodies in early phagosomes
(46). Blocking Stabilin-1 with
anti-Stabilin-1 antibodies has been shown to markedly inhibit
macrophage phagocytosis of apoptotic cells (46). Researchers have prepared
microspheres coated with PS to simulate apoptotic cells; most of
these PS-coated beads are bound and phagocytosed by
Stabilin-1-expressing cells, with the EGF-like repeat domain
(EGFrp) of Stabilin-1 directly binding to PS-coated beads (46). Collectively, these findings
indicate that Stabilin-1 may act as a receptor that directly binds
to PS via the EGFrp, thereby mediating the clearance of apoptotic
cells by macrophages (46)
(Fig. 2).
The Gulp1-dependent signalling pathway for apoptotic
cell clearance is essential for Stabilin-1-mediated phagocytosis
(47,48). Gulp1 consists of a
phosphotyrosine-binding (PTB) domain, a leucine zipper domain and a
proline-rich domain. Acting as an adaptor protein, it transduces
cytoskeletal rearrangement signals among several phagocytic
receptors, including MEGF10, LDL receptor-related protein 1,
platelet endothelial aggregation receptor 1 and scavenger receptor
class B member 1 (SR-BI) (49–52).
The asparagine-proline-x-phenylalanine (NPxF) motif of Stabilin-1,
where × is any amino acid, binds to the PTB domain of Gulp1
(18). Gulp1 functions downstream
of the receptor Stabilin-1, and its knockdown has been reported to
markedly cripple the phagocytic functions mediated by Stabilin-1 in
PS-exposed red blood cells (46).
Taken together, these findings suggest that Stabilin-1 promotes
apoptotic cell phagocytosis by engaging Gulp1 as an interacting
partner, which facilitates anti-inflammatory processes. Gulp1 acts
as a signalling hub to recruit and activate more downstream
molecules. On the one hand, Gulp1 activates MAPK p38 via
phosphorylation, which is involved in the regulation of endothelial
cell proliferation, migration and the inflammatory response
(49). On the other hand, Gulp1
activates the Rac family small GTPase 1 (Rac1) protein via
intermediate signalling molecules (which are not currently fully
understood). The Rac1 protein acts as a central regulatory
component in cytoskeleton reorganization, and promotes the
polymerization and reorganization of actin, providing the driving
force for the decisive phases of angiogenesis, such as endothelial
migration and vascular luminal formation (47,48).
It has been shown that a reduction in the phagocytic ability of
PS-exposed erythrocytes is mediated by the Stabilin-1 protein after
endogenous Gulp1 protein is knocked down, whereas overexpressing
the Gulp1 protein may result in an exaggerated response, where the
phagocytic ability of PS-exposed erythrocytes mediated by
Stabilin-1 is increased (47). An
important cause of vascular fibrosis and instability is
endothelial-to-mesenchymal transition (EndMT), in which the
inflammatory microenvironment serves a pivotal role. The signalling
pathway involving Stabilin-1/Gulp can inhibit the activation of
transcription factors related to the EndMT (such as Snail and
Twist) by removing apoptotic bodies, thus preventing the conversion
of endothelial cells into mesenchymal cells, preserving the
phenotype of the vascular wall, and preventing vascular fibrosis
and stenosis (53).
In addition to Gulp1, the cytoplasmic domain of
Stabilin-1 contains a DXXLL motif that binds to the
Golgi-localizing, γ-adaptin ear domain homology, ARF-binding
proteins (GGA) family of adaptor proteins (GGA1-3) and takes part
in the transport of Stabilin-1 from the trans-Golgi network to
endosomes (54). Moreover, sorting
nexin 17 can bind to the NPxF motif of Stabilin-1, thereby
regulating the expression of the latter on vascular endothelial
cell surfaces (55). These
interactions collectively constitute the regulatory network of the
Stabilin-1 signalling cascade, wherein Gulp is primarily
responsible for phagocytosis-related signal transmission, and GGA
and SNX17 are involved in intracellular transport and the
regulation of surface homeostasis of receptors (47).
Stabilin-1 mediates macrophage clearance
of oxidative stress products
Oxidative stress is a state of imbalance between
oxidants and antioxidants, in which either a lack of antioxidant
defence or overproduction of free radicals can turn into toxic
compounds, which are associated with cellular and biomolecular
damage (56). Notably,
inflammation is a typical example of the pathogenesis of oxidative
stress, and a combination of oxidative stress and inflammation has
been reported in several chronic diseases (57). In the context of chronic liver
injury, the oxidative stress produced by injured hepatocytes
results in the generation of toxic products, including oxidized LDL
(oxLDL), of which malondialdehyde-modified LDL (MDA-LDL) is a key
proinflammatory mediator (16).
Stabilin-1 is able to specifically recognize and internalize these
oxidative stress products, giving rise to ceroid-laden macrophages
and leading to a subsequent reduction in the release of the
proinflammatory chemokine CCL3 (16). This can be an effective way to
reduce the activation of hepatic stellate cells and collagen
deposition, thereby ameliorating the development of liver fibrosis
(58) (Fig. 2).
Scavenger function of Stabilin-1 in liver
SECs (LSECs)
The main source of circulating lipopolysaccharide
(LPS) molecules is the intestinal microbiome (59). When intestinal permeability
changes, a small amount of LPS crosses the mucosal barrier into the
portal vein and is then transported to the liver (60). The liver can clear 80% of the LPS
injected into the systemic circulation within several minutes
(61), among which ~75% of LPS
clearance is mediated by LSECs (62).
In this process, circulating high-density
lipoprotein (HDL) serves as the carrier of LPS (61). HDL promotes the endocytosis of LPS
and prevents LSECs from producing inflammatory cytokines (62). LSECs express a limited number of
surface receptors that can bind the LPS-HDL complex (62). Therefore, the clearance of LPS-HDL
by LSECs is a receptor-mediated process. All LPS-HDL bound to LSECs
is internalized through endocytosis (62), and once internalized, the LPS-HDL
complex is degraded in the lysosomes of LSECs (62). By contrast, LPS that escapes
hepatic elimination acts as a major pathogen triggering systemic
inflammation (63–65), and this process is involved in
cellular and humoral immune responses mediated by Toll-like
receptor 4 (TLR4) (66,67).
The TLR4 signalling pathway is tightly linked to the
occurrence and progression of inflammation (68). The TLR family consists of type I
transmembrane receptors involved in innate immunity that can
recognize pathogen-associated molecular patterns and
damage-associated molecular patterns, serving as crucial links
between adaptive immunity and innate immunity (69,70).
When signals from LPS act on TLR4, it undergoes oligomerization to
intracellularly transmit signals, activating the myeloid
differentiation factor 88 (MyD88)-dependent signalling pathway
(71). MyD88, together with IL
receptor-associated kinase 1 and 4, and TNF receptor-associated
factor 6, serves a decisive role in the activation state of several
transcription factors, such as NF-κB and activator protein 1.
Additionally, several transcription factors have the potential to
induce the secretion of proinflammatory mediators via the MAPK
pathway (72).
It has been shown that scavenger receptors are
involved in the clearance of plasma endotoxins (73). Deficiency of the Stabilin-1
receptor leads to reduced systemic clearance and endocytosis of LPS
by LSECs, but increased production of systemic inflammatory
cytokines (62). Research has
indicated that Stabilin-1-deficient mice are highly sensitive to
LPS, suggesting that TLR4 and Stabilin-1 act as functionally
antagonistic receptors in the immune response to LPS: TLR4 senses
LPS to activate inflammatory signalling, leading to increased
cytokine secretion, whereas Stabilin-1 clears LPS and thus
indirectly regulates the production of inflammatory cytokines
(62) (Fig. 2). Stabilin-1 and Stabilin-2 exhibit
similar functions during LPS clearance. Although the two Stabilin
receptors share structural similarity (55% homology), Stabilin-1
serves a more protective role against endotoxin-mediated injury
compared with Stabilin-2, and Stabilin-1 is the major contributor
to long-term LPS clearance (62).
Adaptive immunity
Regulation of adaptive immune cell
expression by Stabilin-1
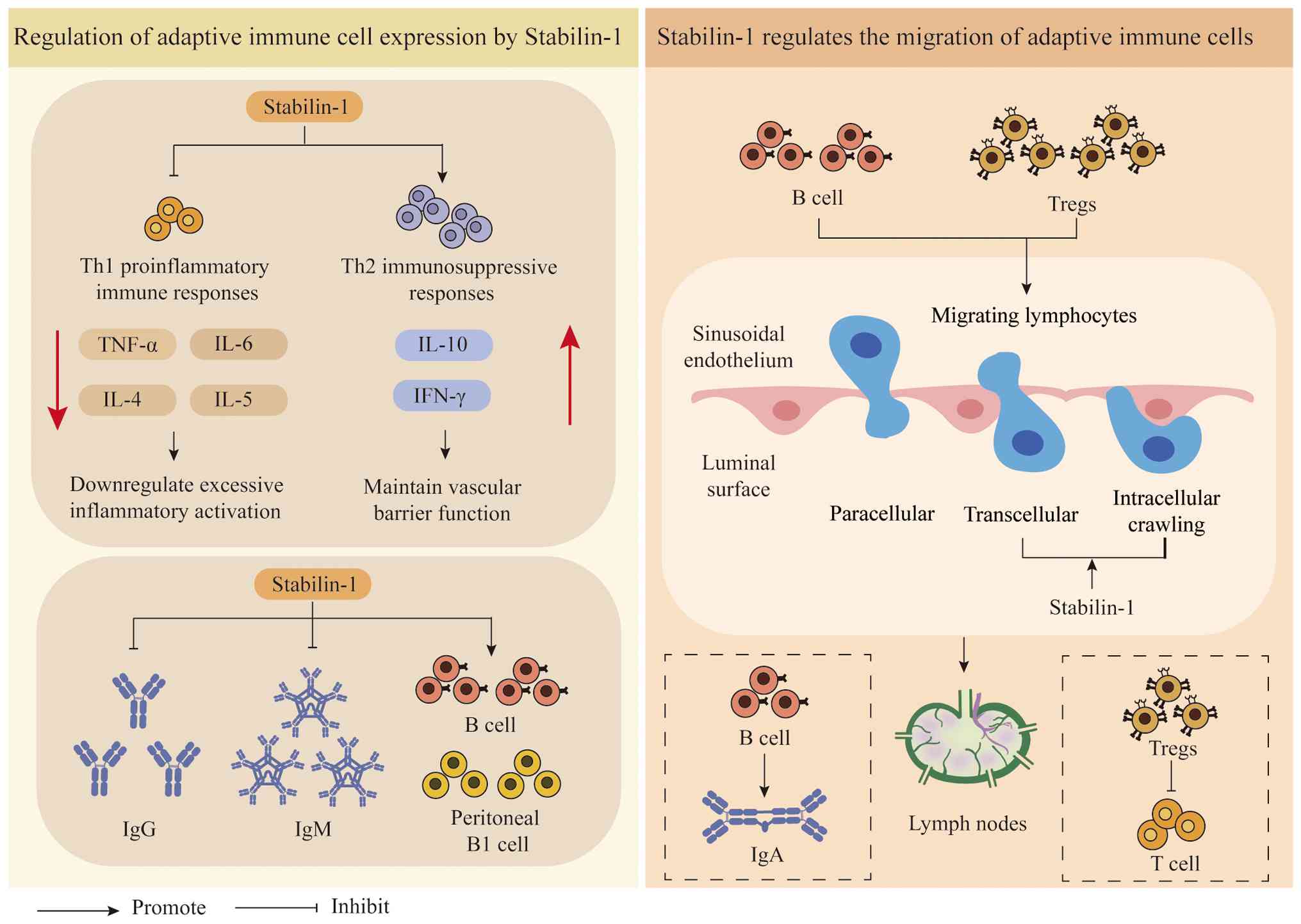
In monocytes, Stabilin-1 functions as an
immunosuppressive factor, thereby inhibiting lymphocyte activation
(25). Stabilin-1 downregulation
is associated with the upregulation of proinflammatory genes and
Stabilin-1 in human monocytes controls the activation of several
proinflammatory genes (25). Under
normal conditions, Stabilin-1 is expressed in
CD14+CD16+ and
CD14+CD16− cell populations, and
CD14+ monocytes with high Stabilin-1 expression exhibit
reduced proinflammatory potential (25). Furthermore, Stabilin-1 expression
promotes the induction of Th2 immunosuppressive responses and
inhibits the formation of Th1 proinflammatory immune responses
(25). It has been demonstrated
that, in the early stage of sepsis, Stabilin-1 may downregulate
excessive inflammatory activation by inhibiting Th1-type immune
responses, whereas in the late stage, it serves a protective role
by maintaining vascular barrier function (25) (Fig.
3). Knockdown of Stabilin-1 in monocytes leads to increased
production of inflammatory cytokines (such as TNF-α), inhibits the
production of interferon (IFN)-γ by T lymphocytes, and promotes
their secretion of IL-4 and IL-5 (25).
Stabilin-1 regulates the activity of B cells through
the production of inflammatory cytokines, such as TNF-α (74). In the absence of Stabilin-1, plasma
IgM and IgG levels are elevated, accompanied by reduced B
lymphocyte generation and a decrease in peritoneal B1 cells
(74) (Fig. 3).
Stabilin-1 regulates the migration of
adaptive immune cells
Stabilin-1 expression demonstrates unique
tissue-specific and inflammation-dependent regulatory features.
Under normal conditions, Stabilin-1 is constitutively expressed in
lymphatic endothelial cells but not in vascular endothelial cells
in skin tissue. In chronic inflammatory skin diseases, such as
psoriasis and lichen planus, vascular endothelial cells induce the
expression of Stabilin-1, and the expression level is positively
associated with the degree of lymphocyte infiltration (75). This makes Stabilin-1 a key molecule
that links the vascular and lymphatic systems in immune trafficking
(75).
In vascular endothelial cells, Stabilin-1 regulates
transendothelial migration under shear stress. The extravasation of
lymphocytes from the bloodstream follows a multistep adhesion
cascade, including rolling, firm adhesion and transendothelial
migration. Under physiologically relevant laminar shear stress
conditions, Stabilin-1 on the surface of vascular endothelial cells
is involved primarily in the transendothelial migration step, and
has only a minor regulatory effect on the rolling process without
influencing firm adhesion (75).
Stabilin-1-mediated transendothelial migration is independent of
binding to HA (75).
In lymphatic endothelial cells, Stabilin-1 mediates
transendothelial migration under static conditions. The mechanism
underlying lymphocyte trafficking in the lymphatic system has long
remained elusive, and Stabilin-1 is the first adhesion molecule
demonstrated to be involved in the lymphatic endothelial
cell-mediated transendothelial migration of peripheral blood
mononuclear cells (PBMCs) (75).
Blood flow velocity within lymphatic vessels is slow, and in
vitro simulation experiments have shown that Stabilin-1 on the
surface of lymphatic endothelial cells has no notable effect on the
firm adhesion of PBMCs but can mediate the transendothelial
migration process (75).
Furthermore, Stabilin-1 mediates the binding of lymphocytes to the
efferent lymphatic vessels of lymph nodes, suggesting that it
regulates lymphocyte trafficking at multiple nodes of the lymphatic
circulation (75).
In a proinflammatory microenvironment that simulates
liver injury, Stabilin-1 facilitates the transendothelial migration
of T cells and B cells to draining lymph nodes (76). Stabilin-1 preferentially mediates
the migration of regulatory T cells (Tregs) and B cells (77–79)
(Fig. 3). This selective
recruitment mechanism has notable pathological implications: Tregs
can promote tumour immune escape by inhibiting effector T-cell
responses, whereas IgA secreted by B cells impairs
anti-hepatocellular carcinoma immune responses, both collectively
driving the initiation and progression of liver tumours (80,81).
Pathological implications
Atherosclerosis
Novel ligands for Stabilin-1 include Periostin
(Postn), Reelin (Reln) and TGF-β-induced protein (TGFBi) (14). All of these proteins are markedly
increased in the absence of Stabilin-1 and are similarly targeted
to lysosomes for degradation through their binding to the fasciclin
domain of Stabilin-1 (14,82). Among these, Postn has been
demonstrated to be involved in the pathological process of
atherosclerosis because of its role as a macrophage
migration-regulating factor, and genetic deficiency (knockout of
Postn) could mitigate atherosclerosis (83). Furthermore, TGFBi is highly
expressed in smooth muscle cells and macrophages in human
atherosclerotic plaques (84), and
Reln deficiency can reduce atherosclerotic risk by decreasing
leukocyte-endothelial cell adhesion (85). Although Stabilin-1 is expressed in
aortic endothelial cells and plaque stromal cells, which likely
contain macrophages, haematopoietic system-specific Stabilin-1
deficiency has been reported to have no effect on the degree of
atherosclerosis in LDL receptor-KO mice (86). These findings suggest that
Stabilin-1 controls the progression of atherosclerosis primarily
through its ability to clear circulating ligands mediated by LSECs,
rather than through local cell-autonomous effects (14). Moreover, Stabilin-1 deficiency did
not markedly influence plasma lipid levels, further confirming that
traditional lipid metabolism pathways are not involved in its mode
of action (14) (Fig. 2).
Chronic liver injury
Stabilin-1 acts via its antifibrotic effect on
macrophages through scavenging, clearing oxidative stress products
and blocking proinflammatory mediators. In relation to chronic
hepatocellular injury, oxidative stress products from injured
hepatocytes, such as oxLDL, particularly MDA-LDL, which acts as a
proinflammatory mediator, have toxic effects. Stabilin-1
specifically targets and removes products of oxidative stress,
resulting in macrophages containing waxy materials, thereby
blocking the proinflammatory chemokine CCL3, hepatic stellate cell
activation and collagen deposition, thereby reducing liver fibrosis
(62,87). This regulatory pathway has been
validated by studies using Stabilin-1 KO mice, which exhibited
changes from anti-inflammatory to proinflammatory programming of
macrophages, with increased levels of the proinflammatory cytokine
TNF-α and decreased arginase levels, and a lower representation of
repair macrophages with low Ly6C expression, resulting in increased
liver fibrosis with delayed repair (16,88,89).
In addition, Stabilin-1 takes part in the intracellular routing
function of macrophages through taking up and processing other
proteins, such as SPARC, into lysosomes, thereby controlling tissue
repair (38,90).
Viral myocarditis
Stabilin-1 neither inhibits viral replication nor
has anti-inflammatory effects, but relieves myocardial injury by
modulating immune responses, resulting in an anti-inflammatory
effect in pathological viral myocarditis. The recruitment of
monocytes and macrophage polarization serve crucial roles in immune
responses in viral myocarditis (26). On the one hand, Stabilin-1 may
preserve the balance of myocardial immunity by promoting
anti-inflammatory macrophage polarization and suppressing
hypertrophy of adaptive immune responses; this will also upregulate
the expression of Ym1, Arg1 and IL-10, thereby improving viral
myocarditis (26) (Fig. 1). On the other hand, Stabilin-1
directly interacts with fibronectin via the extracellular fasciclin
domain (P9 fragment). Binding between Stabilin-1 and the EDA of
fibronectin is the most important mechanism through which monocytes
are recruited to the inflammatory area (26). To date, for viral myocarditis,
there is no specific treatment strategy, and broad-spectrum
immunosuppressive agents induce viral rebound (91). Broad-spectrum immunosuppressants
indiscriminately suppress the function of all immune cells, and
cannot distinguish between ‘overactivated damaging immune cells’
and ‘normal antiviral immune cells’. When using such drugs,
although they can to some extent suppress excessive inflammatory
reactions and reduce myocardial immune damage, they can also weaken
the body's antiviral immune ability. When the ability to clear
viruses decreases due to broad-spectrum immune suppression, the
virus that was originally suppressed by the immune system will
replicate in large quantities again in myocardial cells, resulting
in viral rebound.
Listeria infection
Stabilin-1 can facilitate the phagocytosis and
uptake of Lm by macrophages, mainly by regulating the
internalization of Lm by phagocytic cells rather than reducing
bacterial adhesion ability (13).
Stabilin-1 may regulate the strength of the immune response through
the balance between inflammatory and anti-inflammatory mediators,
not merely through the promotion of inflammatory cytokine
production (13). The effective
recruitment of immune cells to the site of infection is a core
aspect of controlling the spread of Lm, and chemokines are key
signalling molecules that regulate the migration of immune cells.
At the level of chemokine expression, Stabilin-1 upregulates the
expression of chemokines such as CCL2, CXCL10 and CCL7. CXCL10 not
only mediates the migration of immune cells but also has direct
anti-Lm activity, and upregulating its expression may further
increase the ability of the host to resist infection (27). Because CCL7 is a ligand for the
CCR2 receptor, it is crucial for the recruitment of inflammatory
monocytes (28). Stabilin-1 can be
a mediator of myeloid cell recruitment, which is induced by
regulation of the expression of CCL7 and CXCL10. Moreover, it is a
regulatory factor in the early immune cell influx during
inflammation.
For successful infection to take place, Lm must be
able to evade immune clearance mechanisms by controlling the
expression and function of immune proteins. Research has shown that
infection of mice with the pathogenic form of Lm leads to
suppression of the expression of Stabilin-1 in macrophages,
endothelial cells and spleen tissue (13). Notably, the infection caused by Lm
results in the translocation of Stabilin-1 proteins from host cell
membranes to the intracellular compartment (13).
Future studies should aim to more precisely identify
the virulence factors of Lm that control Stabilin-1 expression and
distribution, determine the synergistic impact of Stabilin-1 with
other receptors, such as scavenger receptor-A and macrophage
receptor with collagenous structure, in Lm infection and
investigate the possibility of a novel approach using Stabilin-1 to
increase immune protection against infection (13).
Sepsis
The low pH environment induced by sepsis can
upregulate the expression of Stabilin-1 through the Ets-2 and JNK
signalling pathways (17).
Stabilin-1 mediates the phagocytosis of apoptotic endothelial cells
by macrophages through binding to PS, clears apoptotic cells in the
inflammatory microenvironment, prevents their lysis and release of
proinflammatory factors such as IL-1β, IL-6 and TNF-α, and creates
conditions for the regeneration of vascular endothelial cells,
thereby maintaining vascular homeostasis (26). Furthermore, Stabilin-1 can prevent
tissue damage from excessive inflammatory responses by regulating
the activation state of phagocytic cells. In the early stage of
sepsis, Stabilin-1 may exert a protective effect by inhibiting
Th1-type immune responses and downregulating excessive inflammatory
activation, whereas in the later stage, it primarily exerts a
protective effect by maintaining vascular barrier function
(25). Hepatocellular nuclear
factor 4α(HNF4A) is a transcription factor that is notably
downregulated in the lung tissue and alveolar macrophages of septic
mice; HNF4A promotes the transcriptional expression of nuclear
receptor co activator 2 (NCOA2) by directly binding to its promoter
region; NCOA2, as a co activator of glucocorticoid receptor (GR),
can bind to GR monomers or homodimers, enhancing GR mediated gene
transcription regulation (92).
Studies have shown that HNF4A promotes the polarization of
macrophages towards the M2 phenotype by upregulating the
NCOR2/GR/Stabilin-1 axis, thereby alleviating sepsis-related lung
injury (92).
High mobility group box 1 (HMGB1) is an important
proinflammatory mediator in sepsis (93), which can bind competitively to PS
on the surface of apoptotic cells, interfering with the recognition
of PS by Stabilin-1 and thus inhibiting the phagocytosis of
macrophages (94,95). This constitutes a vicious cycle in
sepsis: Increased HMGB1 release inhibits Stabilin-1-mediated
phagocytosis, resulting in the accumulation of apoptotic cells and
increased release of proinflammatory factors, thus exacerbating the
inflammatory response. These findings identify a new target for
sepsis treatment. Neutralizing antibodies against HMGB1 have been
shown to effectively relieve the functional inhibition of
Stabilin-1 by HMGB1, and improve vascular integrity and survival
rate in mice with sepsis (96).
Cancer
TAMs, which represent the greatest number of innate
immune cells in the tumour microenvironment (TME), are highly prone
to differentiate into proinflammatory M1 or anti-inflammatory M2
macrophages in response to signals from the TME (97). In the majority of cancers, these
cells have been shown to comprise mainly the M2 macrophage subset,
with the ability to facilitate tumour progression through the
secretion of immunosuppressive cytokines and support tumour
angiogenesis (98). Various
studies have shown that infiltrates of Stabilin-1+ TAMs
are strongly associated with cancer aggressiveness and poor patient
survival rates in different types of cancer (7,99,100). A notable negative effect on
patient survival rates has been observed to be related to the
infiltration of CD68+Stabilin-1+ macrophages
in breast cancer (99). In
addition, high numbers of Stabilin-1+ TAMs have been
shown to predict adverse cancer progression outcomes in patients
with early-stage gastric adenocarcinoma (7). In colorectal cancer, the presence and
number of Stabilin-1+ TAMs could be used as a prognostic
marker for disease staging; a high number of Stabilin-1+
TAMs within tumours in patients with stage IV disease is associated
with shortened survival (100).
Similar phenomena have been reported in bladder cancer and acute
myeloid leukaemia (AML) confirming the ubiquitous role of
Stabilin-1+ TAMs in promoting cancer (101,102).
Mechanistic functions include promoting the
phenotypic polarization of macrophages to the M2 phenotype. On the
basis of experimental studies in animals with Stabilin-1 KO, the
phenotypic polarization of TAMs shifts from the M2 phenotype to the
M1 phenotype, indicating the secretion of the inflammatory factors
IL-1β, TNF-α and IL-12p70, as well as the expression of the
chemokines CCL3 and CCL4 (11,16,101,103,104). On the other hand, the high
expression level of Stabilin-1 suppresses the antitumour efficacy
mediated by T cells through the inhibition of antigen presentation
and a reduction in the expression levels of the major
histocompatibility complex, further exacerbating the augmentation
of the immunosuppressive microenvironment through the activation of
Tregs (105). Furthermore,
Stabilin-1 serves a role in metabolic and functional
plasticity-regulating processes in macrophages through the
modulation of the mTOR pathway and the composition of the lysosome
population (11,106).
Stabilin-1 is produced by the lymphatic endothelial
cells and vascular endothelial cells of tumour vessels. It promotes
the lymphatic and haematogenous metastasis of tumour cells by
facilitating the interaction between tumour cells and endothelial
cells (107,108). In squamous cell carcinoma of the
head and neck and breast cancer, a positive association has been
identified between metastatic potential and the density of
Stabilin-1+ lymphatic vessels (106,109). In liver metastatic melanoma,
Stabilin-1 enhances tumour angiogenic remodelling by modifying
Postn and TGF-β1 expression to induce immune cell infiltration
(110). The aforementioned
evidence indicates that Stabilin-1 serves a regulatory role in
promoting tumour metastasis via modification of the tumour
angiogenesis-metastasis cascade. The matrix metalloproteinase
(MMPs) family comprises a vast number of endopeptidases with varied
substrate specificities (111).
By hydrolysing intraprotein peptide bonds, MMPs can dismantle the
vast majority of proteins that make up connective tissue, such as
collagen, elastin, cellulose, gelatine and casein (112). Abnormal expression levels of MMPs
are commonly observed during tumour neovascularization, and tumour
invasion or metastasis (113,114). MMPs enhance angiogenesis through
multiple mechanisms, increasing the bioavailability of
proangiogenic mediators bound to the ECM, and promoting endothelial
cell migration and detachment from the surrounding growing blood
vessels (115,116). Stabilin-1 can regulate the
expression balance of MMPs and tissue inhibitors of
metalloproteinases, affecting the degradation and remodelling of
the ECM (117).
The primary or acquired resistance of some patients
to immune checkpoint inhibitors [such as anti-programmed death-1
(PD-1)/cytotoxic T-lymphocyte-associated protein 4 antibodies]
limits their clinical application, and enrichment of
Stabilin-1+ TAMs is among the key mechanisms leading to
resistance (118). The best-known
immune escape mechanism used by tumour cells is mediated by the
PD-1/programmed death ligand 1 (PD-L1) pathway. PD-1 is a-type I
transmembrane glycoprotein that is a member of the B7/CD28 receptor
family and is expressed on T lymphocytes in humans. Following
T-cell receptor (TCR) activation, PD-1 interacts with PD-L1 and
PD-L2, which are expressed on antigen-presenting cells or
non-hemopoietic tissues, in an attempt to stimulate proinflammatory
cytokines; this is termed the ‘PD-L1/PD-1 axis’. The PD-1/PD-L1
system is an immune regulatory mechanism mediated by intracellular
inhibitory signalling (119).
Stabilin-1+ TAMs inhibit the function of CD8+
T cells through the secretion of IL-10 and TGF-β, while
upregulating the expression of PD-L1 on tumour cells and forming an
immunosuppressive microenvironment that dampens the effects of
immunotherapy (120). Tumour
inflammation can lead to the production of Stabilin-1 as a soluble
form of Clever-1 (sClever-1) following a serine protease-mediated
cleavage process along the IFN-γ/LPS signalling pathway (106). As an isolated immunosuppressive
mediator, sClever-1 selectively interacts with the insulin-like
growth factor 2 receptor (IGF2R) surface receptor on activated T
cells via a mannose-6-phosphate (M6P) interaction (106) and thereby suppresses Y394
phosphorylation of the tyrosine-protein kinase Lck in the TCR
signalling pathway, impairing Th1 cell expansion and promoting the
differentiation of FoxP3+ suppressor CD8+ T
cells (106). Moreover, sClever-1
has been reported to interact with or bind to extracellular
vesicles (EVs) derived from macrophages and further improve the
suppressive effect on T cells by targeting T cells through EV
delivery, decreasing its responsiveness to anti-PD-1 treatment
(106).
In addition to immune regulation, Stabilin-1 can
directly affect the biological properties of tumour cells. In AML,
Stabilin-1 facilitates the polarization of M2 macrophages through
the activation of the IKK/NF-κB signalling pathway, increases the
apoptosis of tumour cells, suppresses cell proliferation and
increases the chemoresistance of tumour cells (101). In papillary thyroid carcinoma,
Stabilin-1 acts as a shield against immune tolerance to support
tumour cell proliferation through the regulation of
CD4+/CD8+ T cells (103). Furthermore, Stabilin-1
participates in ECM tissue remodelling by abolishing the tumour
suppressor factor SPARC from the TME to indirectly support tumour
cell invasion and migration (121,122).
Conclusions and future perspectives
An imbalance in inflammation and disruption of
tissue homeostasis are the key mechanisms involved in the
development and progression of different chronic diseases.
Uncovering key regulatory factors may serve an essential role in
the development of effective strategies for anti-inflammatory
therapeutic intervention (1,6).
Stabilin-1, a functional immune regulator and scavenger receptor
with different functions, has shown high potential in regulating
different immune events and various inflammatory diseases related
to non-specific immunity, specific immunity and other processes.
The intricate mechanisms related to its multifunctional modulation
are inadequately understood, and have an important role in
uncovering different unanswered questions regarding its therapeutic
potential.
In the process of innate immunity, Stabilin-1 has
been reported to favour M2 macrophage polarization and mediate the
recruitment of monocytes via interaction with fibronectin (24,26).
However, in cells other than macrophages, such as SECs and
lymphatic endothelial cells, the details of how ligand binding
triggers the intracellular signalling of Stabilin-1 remain unknown.
In SECs, Stabilin-1 suppresses the TLR4 pathway by mediating LPS
clearance (62). However, whether
this process involves cross-talk with other signalling molecules,
such as negative regulators of NF-κB or specific post-translational
modifications, including phosphorylation and ubiquitination of
Stabilin-1, has not been reported. On this basis, it may be
speculated that, in SECs, by binding to the LPS-HDL complex,
Stabilin-1 can recruit the adaptor protein Gulp1 through its NPxF
motif, and then inhibit the dimerization and nuclear translocation
of NF-κB by activating the MAPKp38 pathway, thus suppressing the
proinflammatory response mediated by TLR4 (18,49).
In the future, researchers may consider detecting the interaction
between Stabilin-1/Gulp1 protein and NF-κB regulatory protein
through co-immunoprecipitation, and changes in NF-κB activity can
be measured in SECs with Stabilin-1 overexpression or knockout
under LPS stimulation. In adaptive immunity, Stabilin-1 modulates
the migration of distinctive immune cells (75,76).
Nevertheless, the mechanism through which Stabilin-1 participates
in the process of transendothelial migration in Tregs, B cells and
lymphatic endothelial cells is not fully understood. Several
reports have shown that Stabilin-1 binds to the GGA protein SNX17,
and inhibits Stabilin-1 intracellular trafficking (54,55).
Therefore, the proposed hypothesis is that in lymphatic endothelial
cells, after binding to Tregs/B-cell surface ligands, Stabilin-1
may form a complex with GGA1 and integrin αvβ3, promoting
cytoskeleton rearrangement through the Rac1 pathway and mediating
transendothelial migration (48).
This hypothesis may be confirmed by analysing transendothelial
migration of lymphatic endothelial cells with GGA1 or integrin αvβ3
knockdown in vitro, as well as by detecting changes in the
binding ability of Stabilin-1 to Tregs/B cells. In the case of
chronic liver disease, Stabilin-1 displays a dual mechanism, which
can facilitate immune evasion in tumours and inhibit fibrosis of
the liver simultaneously (76,87).
On the basis of the current findings, the multiligand binding
property might be associated with the mechanism of its duality,
although the crucial ligands and pathways involved in regulating
its functionality are still unknown. Because Stabilin-1 can
facilitate the clearance of oxidative stress products, such as
MDA-LDL, and recruit Tregs, it may be hypothesized that in the
early phase of chronic injury in the liver, relatively high
expression of MDA-LDL in the microenvironment markedly promotes the
scavenger function of Stabilin-1, thus passively inhibiting liver
fibrosis (16). In the subsequent
phase of injury, the abundant expression of surface ligands on
Tregs may enhance the immune regulatory capabilities of Stabilin-1,
thus leading to tumour progression (78). This hypothesis could be
substantiated through the examination of the expression levels of
MDA-LDL and surface ligands on Tregs in the distinct stages of
chronic inflammation in the liver, in addition to analysing the
functionality of Stabilin-1 following the blocking of respective
ligands.
In sepsis, HMGB1 and Stabilin-1 constitute a
vicious cycle in the HMGB1-Stabilin-1 pathway, increasing the
degree of inflammation. Nevertheless, he regulatory role of
Stabilin-1 gene expression under low pH conditions in sepsis
remains unknown (94,95). Because low pH in sepsis can induce
Stabilin-1 gene upregulation through the Ets-2 and JNK pathways, it
may be hypothesized that HMGB1 further augments the gene
transcription process of Stabilin-1 through increased
phosphorylation of Ets-2 and JNK, which could negatively regulate
Stabilin-1 gene transcription and control excessive inflammation
for homeostasis (17). This
assumption could be proven by assessing changes in Ets-2 and JNK
phosphorylation in HMGB1-treated macrophages in a low pH
environment and investigating Stabilin-1 gene promoter
transcription through a luciferase gene reporter assay.
In tumours, Stabilin-1+ TAMs promote
immune escape through the inhibition of T-cell function, and the
secretion of sClever-1 further enhances immune suppression
(105,106,120). However, the exact mechanism
through which sClever-1 controls the IGF2R-Lck pathway in T cells
remains unclear. It may be proposed that sClever-1 can bind to
IGF2R on the surface of activated T cells through M6P, inducing the
internalization and degradation of Lck and thus suppressing the
phosphorylation of Lck at the Y394 site and hindering T-cell
activation (106). This
hypothesis could be confirmed by detecting the degree of
degradation of Lck in T cells treated with sClever-1, as well as
the degree of recovery after the overexpression of Lck.
In conclusion, Stabilin-1 is known to act as an
immunoregulatory scavenger receptor in the inflammatory
microenvironment and in tissue homeostasis. In-depth research into
the underlying mechanisms of Stabilin-1 activity may not only
strengthen the theoretical basis of immunological regulation in
inflammation, but also offer new targets for and approaches to
different inflammation-related illnesses.
Acknowledgements
Not applicable.
Funding
The present study was supported by the 2024 Zhejiang Province
Basic Public Welfare Research Project (grant no. TGY24H290023) and
the 2024 Zhejiang Province University Student Science and
Technology Innovation Activity Plan (New Talent Plan) (grant no.
2024R410A005).
Availability of data and materials
Not applicable.
Authors' contributions
DLC and STQ conceptualized and designed the
research, outlined the manuscript and prepared the manuscript. STQ
searched for literature and wrote an early version of this
manuscript, and participated in the writing of the current review,
focusing on the relationship between Stabilin-1 and
inflamation-related diseases. XLX contributed to the design and
writing of the original manuscript, and drew the illustrations. XTY
participated in the writing and editing of the original manuscript.
YTW and CC contributed to the editing of the manuscript and
reviewed the literature. CYY translated the manuscript and helped
edit the review. Data authentication is not applicable. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Arulselvan P, Fard MT, Tan WS, Gothai S,
Fakurazi S, Norhaizan ME and Kumar SS: Role of antioxidants and
natural products in inflammation. Oxid Med Cell Longev.
2016:52761302016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Markiewski MM and Lambris JD: The role of
complement in inflammatory diseases from behind the scenes into the
spotlight. Am J Pathol. 171:715–727. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Medzhitov R: Origin and physiological
roles of inflammation. Nature. 454:428–435. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nathan C: Points of control in
inflammation. Nature. 420:846–852. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sohrab SS, Raj R, Nagar A, Hawthorne S,
Paiva-Santos AC, Kamal MA, El-Daly MM, Azhar EI and Sharma A:
Chronic inflammation's transformation to cancer: A nanotherapeutic
paradigm. Molecules. 28:44132023. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Leńska-Mieciek M, Madetko-Alster N, Alster
P, Królicki L, Fiszer U and Koziorowski D: Inflammation in multiple
system atrophy. Front Immunol. 14:12146772023. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yin SP, Gao Y, Xie XS, Xu DD, Riabov V and
Du WD: Accumulation of stabilin-1 positive macrophages in the early
stage of gastric cancer is associated with short cumulative
survival. Oncol Lett. 19:2404–2412. 2020.PubMed/NCBI
|
|
8
|
Prevo R, Banerji S, Ni J and Jackson DG:
Rapid plasma membrane-endosomal trafficking of the lymph node sinus
and high endothelial venule scavenger receptor/homing receptor
stabilin-1 (FEEL-1/CLEVER-1). J Biol Chem. 279:52580–52592. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Weigel JA, Raymond RC, McGary C, Singh A
and Weigel PH: A blocking antibody to the hyaluronan receptor for
endocytosis (HARE) inhibits hyaluronan clearance by perfused liver.
J Biol Chem. 278:9808–9812. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Politz O, Gratchev A, McCourt PA,
Schledzewski K, Guillot P, Johansson S, Svineng G, Franke P,
Kannicht C, Kzhyshkowska J, et al: Stabilin-1 and −2 constitute a
novel family of fasciclin-like hyaluronan receptor homologues.
Biochem J. 362:155–164. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Viitala M, Virtakoivu R, Tadayon S,
Rannikko J, Jalkanen S and Hollmén M: Immunotherapeutic blockade of
macrophage Clever-1 reactivates the CD8+ T-cell response against
immunosuppressive tumors. Clin Cancer Res. 25:3289–3303. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Riabov V, Yin S, Song B, Avdic A,
Schledzewski K, Ovsiy I, Gratchev A, Llopis Verdiell M, Sticht C,
Schmuttermaier C, et al: Stabilin-1 is expressed in human breast
cancer and supports tumor growth in mammary adenocarcinoma mouse
model. Oncotarget. 7:31097–31110. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pombinho R, Pinheiro J, Resende M,
Meireles D, Jalkanen S, Sousa S and Cabanes D: Stabilin-1 plays a
protective role against Listeria monocytogenes infection
through the regulation of cytokine and chemokine production and
immune cell recruitment. Virulence. 12:2088–2103. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Manta CP, Leibing T, Friedrich M, Nolte H,
Adrian M, Schledzewski K, Krzistetzko J, Kirkamm C, David Schmid C,
Xi Y, et al: Targeting of scavenger receptors Stabilin-1 and
Stabilin-2 ameliorates atherosclerosis by a plasma proteome switch
mediating Monocyte/Macrophage suppression. Circulation.
146:1783–1799. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee W, Park SY, Yoo Y, Kim SY, Kim JE, Kim
SW, Seo YK, Park EK, Kim IS and Bae JS: Macrophagic Stabilin-1
restored disruption of vascular integrity caused by sepsis. Thromb
Haemost. 118:1776–1789. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rantakari P, Patten DA, Valtonen J,
Karikoski M, Gerke H, Dawes H, Laurila J, Ohlmeier S, Elima K,
Hübscher SG, et al: Stabilin-1 expression defines a subset of
macrophages that mediate tissue homeostasis and prevent fibrosis in
chronic liver injury. Proc Natl Acad Sci. 113:9298–9303. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Park SY, Bae DJ, Kim MJ, Piao ML and Kim
IS: Extracellular low pH modulates phosphatidylserine-dependent
phagocytosis in macrophages by increasing stabilin-1 expression. J
Biol Chem. 287:11261–11271. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Park SY and Kim IS: Stabilin receptors:
Role as phosphatidylserine receptors. Biomolecules. 9:3872019.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gurung JL, Tamang RL, Madduri L, Bennett
RG, Harris EN, Denton PW and McVicker B: Stabilin-1 in
Tumor-associated macrophages: A potential therapeutic target in
cancer immunotherapy. Biology. 14:11982025. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhuang Z, Yoshizawa-Smith S, Glowacki A,
Maltos K, Pacheco C, Shehabeldin M, Mulkeen M, Myers N, Chong R,
Verdelis K, et al: Induction of M2 macrophages prevents bone loss
in murine periodontitis models. J Dent Res. 98:200–208. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mantovani A, Biswas SK, Galdiero MR, Sica
A and Locati M: Macrophage plasticity and polarization in tissue
repair and remodelling. J Pathol. 229:176–185. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sica A and Mantovani A: Macrophage
plasticity and polarization: In vivo veritas. J Clin Invest.
122:787–795. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Canton J, Neculai D and Grinstein S:
Scavenger receptors in homeostasis and immunity. Nat Rev Immunol.
13:621–634. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
David C, Nance JP, Hubbard J, Hsu M,
Binder D and Wilson EH: Stabilin-1 expression in tumor associated
macrophages. Brain Res. 1481:71–78. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Palani S, Elima K, Ekholm E, Jalkanen S
and Salmi M: Monocyte Stabilin-1 suppresses the activation of Th1
lymphocytes. J Immunol. 196:115–123. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Carai P, Papageorgiou AP, Van Linthout S,
Deckx S, Velthuis S, Lutgens E, Wijnands E, Tschöpe C,
Schmuttermaier C, Kzhyshkowska J, et al: Stabilin-1 mediates
beneficial monocyte recruitment and tolerogenic macrophage
programming during CVB3-induced viral myocarditis. J Mol Cell
Cardiol. 165:31–39. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cole AM, Ganz T, Liese AM, Burdick MD, Liu
L and Strieter RM: Cutting edge: IFN-inducible ELR-CXC chemokines
display defensin-like antimicrobial activity. J Immunol.
167:623–627. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jia T, Serbina NV, Brandl K, Zhong MX,
Leiner IM, Charo IF and Pamer EG: Additive roles for MCP-1 and
MCP-3 in CCR2-mediated recruitment of inflammatory monocytes during
Listeria monocytogenes infection. J Immunol. 180:6846–6853.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Termine JD, Kleinman HK, Whitson SW, Conn
KM, McGarvey ML and Martin GR: Osteonectin, a bone-specific protein
linking mineral to collagen. Cell. 26:99–105. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Iruela-Arispe ML, Lane TF, Redmond D,
Reilly M, Bolender RP, Kavanagh TJ and Sage EH: Expression of SPARC
during development of the chicken chorioallantoic membrane:
Evidence for regulated proteolysis in vivo. Mol Biol Cell.
6:327–343. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bradshaw AD, Reed MJ and Sage EH:
SPARC-null mice exhibit accelerated cutaneous wound closure. J
Histochem Cytochem. 50:1–10. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Barker TH, Framson P, Puolakkainen PA,
Reed M, Funk SE and Sage EH: Matricellular homologs in the foreign
body response: Hevin suppresses inflammation, but hevin and SPARC
together diminish angiogenesis. Am J Pathol. 166:923–933. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Rosset EM and Bradshaw AD:
SPARC/osteonectin in mineralized tissue. Matrix Biol. 52–54. 78–87.
2016.PubMed/NCBI
|
|
34
|
Kelleher CM, McLean SE and Mecham RP:
Vascular extracellular matrix and aortic development. Curr Top Dev
Biol. 62:153–188. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bradshaw AD: The role of SPARC in
extracellular matrix assembly. J Cell Commun Signal. 3:239–246.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wrana JL, Overall CM and Sodek J:
Regulation of the expression of a secreted acidic protein rich in
cysteine (SPARC) in human fibroblasts by transforming growth factor
beta. Comparison of transcriptional and post-transcriptional
control with fibronectin and type I collagen. Eur J Biochem.
197:519–528. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rivera LB, Bradshaw AD and Brekken RA: The
regulatory function of SPARC in vascular biology. Cell Mol Life
Sci. 68:3165–3173. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kzhyshkowska J, Workman G, Cardó-Vila M,
Arap W, Pasqualini R, Gratchev A, Krusell L, Goerdt S and Sage EH:
Novel function of alternatively activated macrophages:
Stabilin-1-mediated clearance of SPARC. J Immunol. 176:5825–5832.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lech M, Gröbmayr R, Weidenbusch M and
Anders HJ: Tissues use resident dendritic cells and macrophages to
maintain homeostasis and to regain homeostasis upon tissue injury:
The immunoregulatory role of changing tissue environments.
Mediators Inflamm. 2012:9513902012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fadok VA, Voelker DR, Campbell PA, Cohen
JJ, Bratton DL and Henson PM: Exposure of phosphatidylserine on the
surface of apoptotic lymphocytes triggers specific recognition and
removal by macrophages. J Immunol. 148:2207–2216. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Leventis PA and Grinstein S: The
distribution and function of phosphatidylserine in cellular
membranes. Annu Rev Biophys. 39:407–427. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Park SY and Kim IS: Engulfment signals and
the phagocytic machinery for apoptotic cell clearance. Exp Mol Med.
49:e331. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hochreiter-Hufford A and Ravichandran KS:
Clearing the dead: Apoptotic cell sensing, recognition, engulfment,
and digestion. Cold Spring Harb Perspect Biol. 5:a0087482013.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Rigotti A, Acton SL and Krieger M: The
class B scavenger receptors SR-BI and CD36 are receptors for
anionic phospholipids. J Biol Chem. 270:16221–16224. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Park SY, Kim SY, Jung MY, Bae DJ and Kim
IS: Epidermal growth factor-like domain repeat of stabilin-2
recognizes phosphatidylserine during cell corpse clearance. Mol
Cell Biol. 28:5288–5298. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Park SY, Jung MY, Lee SJ, Kang KB,
Gratchev A, Riabov V, Kzhyshkowska J and Kim IS: Stabilin-1
mediates phosphatidylserine-dependent clearance of cell corpses in
alternatively activated macrophages. J Cell Sci. 122:3365–3373.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Park SY, Kim SY, Kang KB and Kim IS:
Adaptor protein GULP is involved in stabilin-1-mediated
phagocytosis. Biochem Biophys Res Commun. 398:467–472. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Park SY, Kang KB, Thapa N, Kim SY, Lee SJ
and Kim IS: Requirement of adaptor protein GULP during
stabilin-2-mediated cell corpse engulfment. J Biol Chem.
283:10593–10600. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Osada Y, Sunatani T, Kim IS, Nakanishi Y
and Shiratsuchi A: Signalling pathway involving GULP, MAPK and Rac1
for SR-BI-induced phagocytosis of apoptotic cells. J Biochem.
145:387–394. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hamon Y, Trompier D, Ma Z, Venegas V,
Pophillat M, Mignotte V, Zhou Z and Chimini G: Cooperation between
engulfment receptors: The case of ABCA1 and MEGF10. PLoS One.
1:e1202006. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Su HP, Nakada-Tsukui K, Tosello-Trampont
AC, Li Y, Bu G, Henson PM and Ravichandran KS: Interaction of
CED-6/GULP, an adapter protein involved in engulfment of apoptotic
cells with CED-1 and CD91/low density lipoprotein receptor-related
protein (LRP). J Biol Chem. 277:11772–11779. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Sullivan CS, Scheib JL, Ma Z, Dang RP,
Schafer JM, Hickman FE, Brodsky FM, Ravichandran KS and Carter BD:
The adaptor protein GULP promotes Jedi-1-mediated phagocytosis
through a clathrin-dependent mechanism. Mol Biol Cell.
25:1925–1936. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Freitag A, Wessler I and Racké K:
Phosphodiesterase inhibitors suppress alpha2-adrenoceptor-mediated
5-hydroxytryptamine release from tracheae of newborn rabbits. Eur J
Pharmacol. 354:67–71. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kzhyshkowska J, Gratchev A, Martens JH,
Pervushina O, Mamidi S, Johansson S, Schledzewski K, Hansen B, He
X, Tang J, et al: Stabilin-1 localizes to endosomes and the
trans-Golgi network in human macrophages and interacts with GGA
adaptors. J Leukoc Biol. 76:1151–1161. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Adachi H and Tsujimoto M: Adaptor protein
sorting nexin 17 interacts with the scavenger receptor
FEEL-1/stabilin-1 and modulates its expression on the cell surface.
Biochim Biophys Acta. 1803:553–563. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Kurutas EB: The importance of antioxidants
which play the role in cellular response against
oxidative/nitrosative stress: Current state. Nutr J. 15:712015.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Bayarsaikhan G, Bayarsaikhan D, Lee J and
Lee B: Targeting scavenger receptors in inflammatory disorders and
oxidative stress. Antioxidants. 11:9362022. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Bataller R and Brenner DA: Liver fibrosis.
J Clin Invest. 115:209–218. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Erridge C, Attina T, Spickett CM and Webb
DJ: A high-fat meal induces low-grade endotoxemia: Evidence of a
novel mechanism of postprandial inflammation. Am J Clin Nutr.
86:1286–1292. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Bowman JD, Surani S and Horseman MA:
Endotoxin, Toll-like receptor-4, and atherosclerotic heart disease.
Curr Cardiol Rev. 13:86–93. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Yao Z, Mates JM, Cheplowitz AM, Hammer LP,
Maiseyeu A, Phillips GS, Wewers MD, Rajaram MV, Robinson JM,
Anderson CL and Ganesan LP: Blood-borne lipopolysaccharide is
rapidly eliminated by liver sinusoidal endothelial cells via
High-Density lipoprotein. J Immunol. 197:2390–2399. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Cabral F, Al-Rahem M, Skaggs J, Thomas TA,
Kumar N, Wu Q, Fadda P, Yu L, Robinson JM, Kim J, et al: Stabilin
receptors clear LPS and control systemic inflammation. iScience.
24:1033372021. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Uhde M, Ajamian M, Caio G, De Giorgio R,
Indart A, Green PH, Verna EC, Volta U and Alaedini A: Intestinal
cell damage and systemic immune activation in individuals reporting
sensitivity to wheat in the absence of coeliac disease. Gut.
65:1930–1937. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Stevens BR, Goel R, Seungbum K, Richards
EM, Holbert RC, Pepine CJ and Raizada MK: Increased human
intestinal barrier permeability plasma biomarkers zonulin and FABP2
correlated with plasma LPS and altered gut microbiome in anxiety or
depression. Gut. 67:1555.2–1557. 2018. View Article : Google Scholar
|
|
65
|
Tulkens J, Vergauwen G, Van Deun J,
Geeurickx E, Dhondt B, Lippens L, De Scheerder MA, Miinalainen I,
Rappu P, De Geest BG, et al: Increased levels of systemic
LPS-positive bacterial extracellular vesicles in patients with
intestinal barrier dysfunction. Gut. 69:191–193. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Ospelt C and Gay S: TLRs and chronic
inflammation. Int J Biochem Cell Biol. 42:495–505. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Aderem A and Ulevitch RJ: Toll-like
receptors in the induction of the innate immune response. Nature.
406:782–787. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Chen Z, Dong WH, Wu Q and Wang J:
Two-layer regulation of TRAF6 mediated by both TLR4/NF-kB signaling
and miR-589-5p increases proinflammatory cytokines in the pathology
of severe acute pancreatitis. Am J Transl Res. 12:2379–2395.
2020.PubMed/NCBI
|
|
69
|
Umetsu D: Cell mechanics and cell-cell
recognition controls by Toll-like receptors in tissue morphogenesis
and homeostasis. Fly (Austin). 16:233–247. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Ropert C: How toll-like receptors reveal
monocyte plasticity: The cutting edge of antiinflammatory therapy.
Cell Mol Life Sci. 76:745–755. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
De Oliveira AA, Webb RC and Nunes KP:
Toll-like receptor 4 and Heat-shock protein 70: Is it a new target
pathway for diabetic vasculopathies? Curr Drug Targets. 20:51–59.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Giordano W, Ricciardi G, Casciaro M,
Fiorentino V, Pizzimenti C, Viola A, Martini M, Tuccari G and Ieni
A: Role of IL-33/ST2 pathway in inflammatory bowel disease: An
overview and future perspectives. Gastrointest Disord. 6:446–460.
2024. View Article : Google Scholar
|
|
73
|
Hampton RY, Golenbock DT, Penman M,
Krieger M and Raetz CRH: Recognition and plasma clearance of
endotoxin by scavenger receptors. Nature. 352:342–344. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Dunkel J, Viitala M, Karikoski M,
Rantakari P, Virtakoivu R, Elima K, Hollmén M, Jalkanen S and Salmi
M: Enhanced antibody production in Clever-1/Stabilin-1-Deficient
mice. Front Immunol. 9:22572018. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Irjala H, Elima K, Johansson E, Merinen M,
Kontula K, Alanen K, Grenman R, Salmi M and Jalkanen S: The same
endothelial receptor controls lymphocyte traffic both in vascular
and lymphatic vessels. Eur J Immunol. 33:815–824. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Nahmias J, Aziz E and Geraci L:
Praziquantel in the treatment of schistosomiasis. Harefuah.
105:214–215. 1983.(In Hebrew). PubMed/NCBI
|
|
77
|
Shetty S, Bruns T, Weston CJ, Stamataki Z,
Oo YH, Long HM, Reynolds GM, Pratt G, Moss P, Jalkanen S, et al:
Recruitment mechanisms of primary and malignant B cells to the
human liver. Hepatology. 56:1521–1531. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Patten DA, Wilson GK, Bailey D, Shaw RK,
Jalkanen S, Salmi M, Rot A, Weston CJ, Adams DH and Shetty S: Human
liver sinusoidal endothelial cells promote intracellular crawling
of lymphocytes during recruitment: A new step in migration.
Hepatology. 65:294–309. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Shetty S, Weston CJ, Oo YH, Westerlund N,
Stamataki Z, Youster J, Hubscher SG, Salmi M, Jalkanen S, Lalor PF
and Adams DH: Common lymphatic endothelial and vascular endothelial
Receptor-1 mediates the transmigration of regulatory T cells across
human hepatic sinusoidal endothelium. J Immunol. 186:4147–4155.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Shalapour S, Lin XJ, Bastian IN, Brain J,
Burt AD, Aksenov AA, Vrbanac AF, Li W, Perkins A, Matsutani T, et
al: Inflammation-induced IgA+ cells dismantle anti-liver cancer
immunity. Nature. 551:340–345. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Tang Q and Bluestone JA: Regulatory T-cell
physiology and application to treat autoimmunity. Immunol Rev.
212:217–237. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Conway S and Molkentin J: Periostin as a
heterofunctional regulator of cardiac development and disease. Curr
Genomics. 9:548–555. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Schwanekamp JA, Lorts A, Vagnozzi RJ,
Vanhoutte D and Molkentin JD: Deletion of periostin protects
against atherosclerosis in mice by altering inflammation and
extracellular matrix remodeling. Arterioscler Thromb Vasc Biol.
36:60–68. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
O'Brien ER, Bennett KL, Garvin MR, Zderic
TW, Hinohara T, Simpson JB, Kimura T, Nobuyoshi M, Mizgala H,
Purchio A and Schwartz SM: Beta ig-h3, a transforming growth
factor-beta-inducible gene, is overexpressed in atherosclerotic and
restenotic human vascular lesions. Arterioscler Thromb Vasc Biol.
16:576–584. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Ding Y, Huang L, Xian X, Yuhanna IS,
Wasser CR, Frotscher M, Mineo C, Shaul PW and Herz J: Loss of
Reelin protects against atherosclerosis by reducing
leukocyte-endothelial cell adhesion and lesion macrophage
accumulation. Sci Signal. 9:ra292016. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Nahon JE, Hoekstra M, Van Hulst S, Manta
C, Goerdt S, Geerling JJ, Géraud C and Van Eck M: Hematopoietic
Stabilin-1 deficiency does not influence atherosclerosis
susceptibility in LDL receptor knockout mice. Atherosclerosis.
281:47–55. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Patten DA and Shetty S: The role of
stabilin-1 in lymphocyte trafficking and macrophage scavenging in
the liver microenvironment. Biomolecules. 9:2832019. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Ramachandran P, Pellicoro A, Vernon MA,
Boulter L, Aucott RL, Ali A, Hartland SN, Snowdon VK, Cappon A,
Gordon-Walker TT, et al: Differential Ly-6C expression identifies
the recruited macrophage phenotype, which orchestrates the
regression of murine liver fibrosis. Proc Natl Acad Sci.
109:E3186–E3195. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Karlmark KR, Weiskirchen R, Zimmermann HW,
Gassler N, Ginhoux F, Weber C, Merad M, Luedde T, Trautwein C and
Tacke F: Hepatic recruitment of the inflammatory Gr1+ monocyte
subset upon liver injury promotes hepatic fibrosis. Hepatology.
50:261–274. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Kzhyshkowska J, Mamidi S, Gratchev A,
Kremmer E, Schmuttermaier C, Krusell L, Haus G, Utikal J,
Schledzewski K, Scholtze J and Goerdt S: Novel stabilin-1
interacting chitinase-like protein (SI-CLP) is up-regulated in
alternatively activated macrophages and secreted via lysosomal
pathway. Blood. 107:3221–3228. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Tschöpe C, Ammirati E, Bozkurt B, Caforio
ALP, Cooper LT, Felix SB, Hare JM, Heidecker B, Heymans S, Hübner
N, et al: Myocarditis and inflammatory cardiomyopathy: Current
evidence and future directions. Nat Rev Cardiol. 18:169–193. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Yang YH, Wen R, Huang XM, Zhang T, Yang N,
Liu CF and Zhang TN: HNF4A mitigates sepsis-associated lung injury
by upregulating NCOR2/GR/STAB1 axis and promoting macrophage
polarization towards M2 phenotype. Cell Death Dis. 16:1202025.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Bae JS: Role of high mobility group box 1
in inflammatory disease: Focus on sepsis. Arch Pharm Res.
35:1511–1523. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Friggeri A, Yang Y, Banerjee S, Park YJ,
Liu G and Abraham E: HMGB1 inhibits macrophage activity in
efferocytosis through binding to the alphavbeta3-integrin. Am J
Physiol Cell Physiol. 299:C1267–C1276. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Liu G, Wang J, Park YJ, Tsuruta Y, Lorne
EF, Zhao X and Abraham E: High mobility group protein-1 inhibits
phagocytosis of apoptotic neutrophils through binding to
phosphatidylserine. J Immunol. 181:4240–4246. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Bogatcheva NV and Verin AD: The role of
cytoskeleton in the regulation of vascular endothelial barrier
function. Microvasc Res. 76:202–207. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Aizaz M, Khan A, Khan F, Khan M, Musad
Saleh EA, Nisar M and Baran N: The cross-talk between macrophages
and tumor cells as a target for cancer treatment. Front Oncol.
13:12590342023. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Larionova I, Kiselev A, Kazakova E, Liu T,
Patysheva M, Iamshchikov P, Liu Q, Mossel DM, Riabov V, Rakina M,
et al: Tumor-associated macrophages respond to chemotherapy by
detrimental transcriptional reprogramming and suppressing
stabilin-1 mediated clearance of EGF. Front Immunol.
14:10004972023. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Ammar A, Mohammed RAA, Salmi M, Pepper M,
Paish EC, Ellis IO and Martin SG: Lymphatic expression of CLEVER-1
in breast cancer and its relationship with lymph node metastasis.
Anal Cell Pathol. 34:67–78. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Ålgars A, Irjala H, Vaittinen S, Huhtinen
H, Sundström J, Salmi M, Ristamäki R and Jalkanen S: Type and
location of tumor-infiltrating macrophages and lymphatic vessels
predict survival of colorectal cancer patients. Int J Cancer.
131:864–873. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Yin J, Luo J, Wang L and Liu L and Liu L:
STAB1 promotes acute myeloid leukemia progression by activating the
IKK/NF-κB pathway and increasing M2 macrophage polarization. Cancer
Sci. 116:1508–1521. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Boström MM, Irjala H, Mirtti T, Taimen P,
Kauko T, Ålgars A, Jalkanen S and Boström PJ: Tumor-associated
macrophages provide significant prognostic information in
urothelial bladder cancer. PLoS One. 10:e01335522015. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Spourquet C, Delcorte O, Lemoine P,
Dauguet N, Loriot A, Achouri Y, Hollmén M, Jalkanen S, Huaux F,
Lucas S, et al: BRAFV600E expression in thyrocytes causes
recruitment of immunosuppressive STABILIN-1 macrophages. Cancers.
14:46872022. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Karikoski M, Marttila-Ichihara F, Elima K,
Rantakari P, Hollmén M, Kelkka T, Gerke H, Huovinen V, Irjala H,
Holmdahl R, et al: Clever-1/stabilin-1 controls cancer growth and
metastasis. Clin Cancer Res. 20:6452–6464. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Mutka M, Virtakoivu R, Joensuu K, Hollmén
M and Heikkilä P: Clever-1 positive macrophages in breast cancer.
Breast Cancer Res Treat. 195:237–248. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Hollmén M, Figueiredo CR and Jalkanen S:
New tools to prevent cancer growth and spread: A ‘Clever’ approach.
Br J Cancer. 123:501–509. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Irjala H, Alanen K, Grénman R, Heikkilä P,
Joensuu H and Jalkanen S: Mannose receptor (MR) and common
lymphatic endothelial and vascular endothelial receptor (CLEVER)-1
direct the binding of cancer cells to the lymph vessel endothelium.
Cancer Res. 63:4671–4676. 2003.PubMed/NCBI
|
|
108
|
Salmi M, Koskinen K, Henttinen T, Elima K
and Jalkanen S: CLEVER-1 mediates lymphocyte transmigration through
vascular and lymphatic endothelium. Blood. 104:3849–3857. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Liu T, Larionova I, Litviakov N, Riabov V,
Zavyalova M, Tsyganov M, Buldakov M, Song B, Moganti K, Kazantseva
P, et al: Tumor-associated macrophages in human breast cancer
produce new monocyte attracting and pro-angiogenic factor YKL-39
indicative for increased metastasis after neoadjuvant chemotherapy.
Oncoimmunology. 7:e14369222018. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Häfele V, Dietsch B, Straub N, Goerdt S
and Géraud C: Deficiency of Stabilin-1 in the context of hepatic
melanoma metastasis. Cancers (Basel). 16:4412024. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Choi YC and Dalakas MC: Expression of
matrix metalloproteinases in the muscle of patients with
inflammatory myopathies. Neurology. 54:65. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Koh Y and Jaoude J: Matrix
metalloproteinases in exercise and obesity. Vasc Health Risk Manag.
12:287–295. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Gobin E, Bagwell K, Wagner J, Mysona D,
Sandirasegarane S, Smith N, Bai S, Sharma A, Schleifer R and She
JX: A pan-cancer perspective of matrix metalloproteases (MMP) gene
expression profile and their diagnostic/prognostic potential. BMC
Cancer. 19:5812019. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Cui N, Hu M and Khalil RA: Biochemical and
biological attributes of matrix metalloproteinases. Prog Mol Biol
Transl Sci. 147:1–73. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Ballato M, Germanà E, Ricciardi G,
Giordano WG, Tralongo P, Buccarelli M, Castellani G, Ricci-Vitiani
L, D'Alessandris QG, Giuffrè G, et al: Understanding
neovascularization in glioblastoma: Insights from the current
literature. Int J Mol Sci. 26:27632025. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Rundhaug JE: Matrix metalloproteinases and
angiogenesis. J Cell Mol Med. 9:267–285. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Parks WC, Wilson CL and López-Boado YS:
Matrix metalloproteinases as modulators of inflammation and innate
immunity. Nat Rev Immunol. 4:617–629. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Rannikko JH, Verlingue L, de Miguel M,
Pasanen A, Robbrecht D, Skytta T, Iivanainen S, Shetty S, Ma YT,
Graham DM, et al: Bexmarilimab-induced macrophage activation leads
to treatment benefit in solid tumors: The phase I/II first-in-human
MATINS trial. Cell Rep Med. 4:1013072023. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Fiorentino V, Pizzimenti C, Franchina M,
Pepe L, Russotto F, Tralongo P, Micali MG, Militi GB and Lentini M:
Programmed cell death ligand 1 immunohistochemical expression and
cutaneous melanoma: A controversial relationship. Int J Mol Sci.
25:6762024. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Yu K, Cao Y, Zhang Z, Wang L, Gu Y, Xu T,
Zhang X, Guo X, Shen Z and Qin J: Blockade of CLEVER-1 restrains
immune evasion and enhances anti-PD-1 immunotherapy in gastric
cancer. J Immunother Cancer. 13:e0110802025. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Adachi H and Tsujimoto M: FEEL-1, a novel
scavenger receptor with in vitro bacteria-binding and
angiogenesis-modulating activities. J Biol Chem. 277:34264–34270.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Kzhyshkowska J: Multifunctional receptor
stabilin-1 in homeostasis and disease. Sci World J. 10:2039–2053.
2010. View Article : Google Scholar
|