Introduction
Primary Sjögren's syndrome (pSS) is a systemic
autoimmune disorder characterized by chronic progressive
inflammation, lymphocytic infiltration, exocrine gland destruction
and production of disease-specific autoantibodies (1). It has been reported that 70–98% of
patients with pSS develop clinically notable dry eye disease
globally (2). In the absence of
effective intervention, persistent ocular surface inflammation may
progress to severe epithelial damage and irreversible visual
impairment, markedly compromising quality of life of patients.
Progressive lacrimal gland destruction represents a central
pathogenic event in pSS-associated aqueous-deficient dry eye
(3). Therefore, elucidating the
molecular mechanisms underlying lacrimal gland injury is key to
developing targeted therapeutic strategies for severe and
treatment-refractory disease.
Increasing evidence indicates that a sustained
inflammatory microenvironment is a principal driver of target organ
damage in pSS. Immune dysregulation, including the type I
interferon (IFN) signature (4) and
activation of toll-like receptors, promotes the production of
proinflammatory cytokines (5).
Among these mediators, interleukin (IL)-17 is consistently elevated
and contributes to tissue inflammation through amplification of
downstream inflammatory cascades, partly via activation of nuclear
factor κB (NF-κB) signaling. Furthermore, aberrant expression of
costimulatory molecules, including members of the B7 family
(6,7), and activation of the NLR family pyrin
domain containing 3 inflammasome (8) further exacerbate immune cell
infiltration and glandular injury. Although these
immunopathological networks have been extensively investigated in
salivary glands (9,10), the specific signaling mechanisms
governing lacrimal gland damage remain incompletely defined.
Current evidence regarding the lacrimal gland microenvironment
indicates that stressed epithelial cells release specific
chemokines to recruit dendritic cells and macrophages.
Specifically, the secretion of C-X-C motif chemokine ligand 10
drives the recruitment of T helper (Th)1 cells, while the release
of CCL20 and CCL19 facilitates the homing of dendritic cells and
Th17 subsets. This process initiates the marked infiltration and
homing of autoreactive T lymphocytes, predominantly the Th1 and
Th17 subsets. The subsequent local accumulation of these immune
cells leads to a robust release of proinflammatory cytokines,
including IFN-γ and IL-17, which directly drive lacrimal gland
destruction (11,12). Recent studies have highlighted
potential therapeutic strategies for pSS-related dry eye, such as
restoring aquaporin 5 function (13) and suppressing T-cell activation via
cyclosporine A (14). However, the
molecular mechanisms and specific inflammatory cascades directly
driving lacrimal gland pathogenesis remain incompletely elucidated,
necessitating further mechanistic investigation in future
research.
The non-obese diabetic (NOD)/Ltj mouse is a
well-established experimental model of pSS that recapitulates
autoantibody production, lymphocytic infiltration and impaired
exocrine gland secretion (15–17).
Compared with non-autoimmune wild-type control mice, the genetic
background of this specific strain is characterized by multiple
distinct susceptibility loci and a unique major histocompatibility
complex (MHC) class II haplotype. Specifically, the NOD/Ltj strain
carries the H2g7 haplotype, which is characterized by
structural alterations in the I-Aβ chain and defective expression
of the Ea gene, resulting in the absence of functional I-E
molecules. In addition to MHC-related features, the NOD/Ltj strain
harbors multiple susceptibility loci outside the MHC region,
including variants in the IL2 gene associated with reduced
IL-2 production, as well as functional polymorphisms in
CTLA4. This polygenic genotype fundamentally impairs central
and peripheral immune tolerance. Therefore, autoreactive
lymphocytes escape deletion and undergo spontaneous activation
directly driving the autoimmune exocrinopathy. The present study
aimed to delineate the molecular pathways contributing to lacrimal
gland injury in pSS-associated dry eye. Specifically, the present
study examined the contribution of NF-κB-mediated Th17 responses to
local inflammation and tissue damage. Lastly, the present study
evaluated whether pharmacological inhibition of NF-κB using JSH-23
could attenuate inflammatory cytokine production, reduce apoptosis
and restore tear secretion, thereby establishing a mechanistic
association between NF-κB activation and lacrimal gland dysfunction
in pSS.
Materials and methods
Animal experiments
To investigate the role of NF-κB signaling in
lacrimal gland injury, 20 male NOD/Ltj mice (12 weeks old, weighing
24–28 g) were obtained from Beijing Huafukang Biotechnology Co.,
Ltd. [Experimental Animal Production License No.
SCXK(Jing)2019-0008] and 16 age-matched male Institute of Cancer
Research (ICR) mice (weighing 35–40 g) were obtained from Shanghai
SLAC Laboratory Animal Co., Ltd. [Experimental Animal Production
License No. SCKK(HU)2022-0004]. Animals were housed under specific
pathogen-free conditions at 18–22°C and 40–60% relative humidity
with a 12-h light/dark cycle and ad libitum access to
standard food and water. Mice were maintained until 20 weeks of age
prior to experimental analysis.
The systemic dosage of JSH-23 (MedChemExpress) was
set at 3 mg/kg daily based on previously validated pharmacological
protocols. This dosage has been reported to effectively inhibit
NF-κB activation and its downstream proinflammatory cascades in
various murine models. Previous studies have confirmed that this
specific dose attenuates inflammatory injury in vital organs,
including the lung liver and kidney, without inducing notable
systemic toxicity (18,19). Therefore, 3 mg/kg JSH-23 was
selected to investigate its mechanistic impact on lacrimal gland
injury in the present study. In the NOD/Ltj + JSH-23 treatment
group (n=6), mice received the NF-κB inhibitor JSH-23 (3 mg/kg) by
oral gavage once daily for 4 weeks. Untreated NOD/Ltj mice and ICR
control mice received an equal volume of sterile normal saline by
oral gavage at the same frequency to control for handling-related
stress. At the end of the treatment period, mice were euthanized by
cervical dislocation and lacrimal glands were harvested for
subsequent analyses. All procedures were conducted in accordance
with the Animal Research: Reporting of In Vivo Experiments
guidelines (20) and complied with
the National Institutes of Health Guide for the Care and Use of
Laboratory Animals (21). The
experimental protocol was approved by the Animal Care and Welfare
Committee of Zhejiang University (approval no. 2022-190; Zhejiang,
China).
Tear production measurement
To evaluate lacrimal gland secretory function, tear
production was measured every 5 days during the initial assessment,
and every 4 days beginning at treatment initiation (20 weeks of
age) using the phenol red-impregnated cotton thread test (Tianjin
Jingming New Technological Development Co. Ltd.) (14). Measurements were continued until 24
weeks of age. During each assessment, mice were gently restrained
without anesthesia to avoid suppression of basal tear secretion.
The lower eyelid was carefully retracted and one end of the phenol
red cotton thread was placed into the lateral canthus of the
conjunctival fornix. After 1 min, the length of the wetted red
portion of the thread was measured in millimeters (mm) using a
calibrated ruler with 0.5-mm precision. Each eye was tested three
times consecutively at the same time of day to minimize diurnal
variation. All measurements were performed independently by two
investigators blinded to group allocation. The final tear secretion
value for each eye was calculated as the mean of the measurements
obtained by the two observers.
Histopathological evaluation and
immunohistochemistry (IHC)
To evaluate histopathological alterations and
determine in situ expression levels of target proteins and
immune cell infiltration in lacrimal glands, hematoxylin and eosin
(H&E) staining and IHC were performed. Lacrimal gland tissues
were fixed in 4% paraformaldehyde (PFA) for 24 h at room
temperature, embedded in paraffin and sectioned (thickness, 4 µm).
Paraffin sections were deparaffinized and rehydrated through graded
ethanol solutions (100% ethanol, two changes, 3 min each; 95%
ethanol, 2 min; 70% ethanol, 2 min), followed by rinsing in
distilled water.
For H&E staining, sections were stained with
hematoxylin for 5 min and counterstained with eosin for 2 min at
room temperature to visualize the overall tissue structure and cell
morphology. Stained sections were visualized and images were
captured using a light microscope (TS100; Nikon Corporation).
For IHC analysis, lacrimal gland tissues were fixed
in 4% PFA for 24 h at room temperature and sectioned at a thickness
of 4 µm. Antigen retrieval was performed in citrate buffer (pH,
6.0) at 95–100°C for 15 min, followed by washing with PBS.
Endogenous peroxidase activity was blocked with 3% hydrogen
peroxide at room temperature for 15 min. Sections were then blocked
with 5% bovine serum albumin (BSA) containing 0.3% Triton X-100
(for permeabilization) for 30 min at room temperature and incubated
overnight at 4°C with the following primary antibodies: Anti-CARD14
(cat. no. A16569; 1:50; ABclonal Biotech Co., Ltd.), anti-CCL19
(cat. no. PA5-109488; 1:50; Invitrogen; Thermo Fisher Scientific,
Inc.), anti-CD4 (cat. no. MA1-146; 1:50; Invitrogen; Thermo Fisher
Scientific, Inc.) and anti-IL-17A (cat. no. sc-374218; 1:50; Santa
Cruz Biotechnology, Inc.). After washing with phosphate-buffered
saline (PBS), sections were incubated with horseradish
peroxidase-conjugated secondary antibody (cat. no. 31460; 1:500;
Invitrogen; Thermo Fisher Scientific, Inc.) for 1 h at room
temperature. Immunoreactivity was visualized using
3,3′-diaminobenzidine substrate, followed by hematoxylin
counterstaining at room temperature for 2 min. Sections were
dehydrated, cleared and mounted with coverslips. Images were
captured using a light microscope (TS100; Nikon Corporation) at
×200 magnification. For quantitative analysis, at least 3 randomly
selected non-overlapping fields per section were analyzed using
ImageJ software (version 1.48v; National Institutes of Health). The
mean integrated optical density or positive staining area per field
was calculated for each sample.
mRNA sequencing (mRNA-seq) and
bioinformatics analysis
To identify transcriptomic alterations associated
with lacrimal gland dysfunction, mRNA-seq was performed on lacrimal
gland tissues obtained from NOD/Ltj mice (n=3) and ICR mice (n=3).
Total RNA was extracted using TRIzol® reagent (cat. no.
15596026; Invitrogen; Thermo Fisher Scientific, Inc.), according to
the manufacturer's protocol. RNA concentration and purity were
assessed using a NanoDrop™ spectrophotometer (Thermo
Fisher Scientific, Inc.) and RNA integrity was evaluated using an
Agilent 2100 Bioanalyzer (Agilent Technologies, Inc.). Samples with
an RNA integrity number ≥7.0 were used for library preparation.
Poly(A) mRNA was enriched using oligo(dT) magnetic beads and
fragmented into short segments. Complementary DNA (cDNA) was
synthesized using the NEBNext® Ultra™ II RNA
Library Prep Kit for Illumina® (cat. no. E7770S; New
England BioLabs, Inc.). The concentration of the final libraries
was measured using a Qubit 4.0 Fluorometer (Thermo Fisher
Scientific, Inc.), and the final library was loaded at a
concentration of 1.5 pM. Libraries were amplified by PCR and
validated prior to sequencing. Paired-end sequencing [2×150 bp
(PE150)] was performed on an Illumina NovaSeq™ 6000
platform (Illumina, Inc.) using the NovaSeq 6000 S4 Reagent Kit
(cat. no. 20028312; Illumina, Inc.).
Raw sequencing reads were filtered to remove adapter
sequences and low-quality reads prior to downstream analysis.
Differential gene expression analysis between groups was performed
using the ‘DESeq2’ package (version 1.46.0; http://bioconductor.org/packages/DESeq2/) in R
software. Genes with |log2 fold change (FC)|>1 and an
adjusted P-value (false discovery rate) <0.05 were considered
significantly differentially expressed. To functionally annotate
the identified differentially expressed genes (DEGs), Gene Ontology
(GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway
enrichment analysis were conducted using the ‘clusterProfiler’
package (version 4.14.6; http://bioconductor.org/packages/clusterProfiler/) in
R software (R Development Core Team). GO enrichment analysis
categorized the DEGs into biological processes, cellular components
and molecular functions. KEGG pathway analysis was performed to
identify significantly enriched signaling pathways. Pathways
containing >2 genes and an adjusted P<0.05 was considered to
indicate a statistically significant difference. Transcription
factor-associated regulatory networks were constructed using
weighted correlation network analysis based on the top 2,000 most
variable genes to identify key regulatory modules associated with
disease phenotype.
The raw mRNA-seq data generated in the present study
have been deposited in the National Center for Biotechnology
Information (NCBI) Gene Expression Omnibus database (https://www.ncbi.nlm.nih.gov/geo/) under
accession number GSE306385.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
To validate differential gene expression identified
by mRNA-seq, total RNA was extracted from lacrimal gland tissues
using the RNeasy Mini Kit (Qiagen GmbH) according to the
manufacturer's instructions. cDNA was synthesized using the
iScript™ cDNA Synthesis Kit (Bio-Rad Laboratories, Inc.)
according to the manufacturer's protocol (25°C for 5 min, 46°C for
20 min and 95°C for 1 min). qPCR was performed using
iTaq™ Universal SYBR® Green SuperMix (Bio-Rad
Laboratories, Inc.). Target-specific primers were designed using
NCBI Primer-Basic Local Alignment Search Tool (BLAST) to span at
least one exon-exon junction. Primer sequences are listed in
Table I. Thermocycling conditions
were as follows: Initial denaturation at 95°C for 30 sec, followed
by 40 cycles of denaturation at 95°C for 5 sec and
annealing/extension at 60°C for 30 sec. A melting curve analysis
(65–95°C) was performed at the end of each run to confirm
amplification specificity. Relative gene expression levels were
calculated using the 2−ΔΔCq method (22), with β-actin serving as the internal
reference gene. The specific primers for mouse β-actin were
designed using the NCBI Primer-BLAST software based on the
reference cDNA sequence (GenBank accession no. NM_007393.5).
 | Table I.Primer sequences used for reverse
transcription-quantitative PCR (mouse). |
Table I.
Primer sequences used for reverse
transcription-quantitative PCR (mouse).
| Gene | Primer sequences
(5′-3′) |
|---|
| CARD14 | F:
CAGCACTTTCAGCGGTCTC |
|
| R:
CATCTCGTCCTTCAGTTTCAG |
| TNFSF14 | F:
ATTGGTGGACCTCTGTTATGG |
|
| R:
CTAACTCCTTCGGGTAGCG |
| TNFSF13 | F:
GGTGATGTGGCAACCAGTA |
|
| R:
CCTTGTCCTTCCCGAGATA |
| CCL19 | F:
CTGCCTCAGATTATCTGCCAT |
|
| R:
CTTCCGCATCATTAGCACCC |
| CCL20 | F:
GTGGGTTTCACAAGACAGA |
|
| R:
CTCTTAGGCTGAGGAGGTT |
| CCL12 | F:
CACTTCTATGCCTCCTGCTC |
|
| R:
TGGCTGCTTGTGATTCTCC |
| β-actin | F:
CCCAACTTGATGTATGAAGG |
|
| R:
TTTGTGTAAGGTAAGGTGTGC |
Western blotting
To evaluate the activation of the NF-κB signaling
pathway and the expression levels of downstream inflammatory
mediators at the protein level, lacrimal gland tissues were
homogenized and lysed in radioimmunoprecipitation assay buffer
(cat. no. 87787; Thermo Fisher Scientific, Inc.) supplemented with
protease and phosphatase inhibitor cocktails. Protein
concentrations were determined using a bicinchoninic acid protein
assay kit (Thermo Fisher Scientific, Inc.). Equal amounts of total
protein (30 µg/lane) were separated in 10% gels using sodium
dodecyl sulfate-polyacrylamide gel electrophoresis and transferred
onto polyvinylidene difluoride membranes (MilliporeSigma).
Membranes were blocked with 5% BSA in Tris-buffered saline (TBS)
containing 0.1% Tween-20 (TBST) for 1 h at room temperature and
incubated overnight at 4°C with primary antibodies. After washing
with TBST, membranes were incubated with the appropriate infrared
dye-conjugated secondary antibodies for 1 h at room temperature in
the dark. Protein bands were detected using an Odyssey®
Infrared Imaging System (LI-COR Biosciences). Band intensities were
quantified using ImageJ software (Version 1.48v; National
Institutes of Health). Relative protein expression levels were
normalized to β-actin.
Primary antibodies against NF-κB p65 (cat. no.
8242T), phosphorylated (p-) NF-κB p65 (cat. no. 3033T), inhibitor
of κB kinase (IKK)α (cat. no. 11930T), IKKβ (cat. no. 8943T),
p-IKKα/β (cat. no. 2697T), inhibitor of κBα (IκBα; cat. no. 4814T)
and p-IκBα (cat. no. 2859T) were purchased from Cell Signaling
Technology, Inc., and used at a dilution of 1:1,000. Antibodies
against tumor necrosis factor-α (TNF-α; cat. no. 60291-1-1G) was
obtained from Proteintech Group, Inc. and used at a dilution of
1:1,000. The β-actin antibody (cat. no. 20536-1-AP; Proteintech
Group, Inc.) was used at a dilution of 1:5,000. The secondary
antibodies used were IRDye 800CW Goat anti-Rabbit IgG (cat. no.
926-32211; 1:10,000; LI-COR Biosciences) and IRDye 680RD Goat
anti-Mouse IgG (cat. no. 926-68070; 1:10,000; LI-COR
Biosciences).
Flow cytometry and cell sorting
At the experimental endpoint, peripheral whole blood
samples (~0.8 ml per mouse) were collected from the mice in each
experimental group (ICR controls, NOD/Ltj and NOD/Ltj + JSH-23) via
retro-orbital venous plexus bleeding. No other clinical procedures
or experimental manipulations were performed on the mice on the day
of sacrifice prior to blood collection. All samples were obtained
as the primary procedure under deep anesthesia. Specifically,
anesthesia was induced using 3–4% inhaled isoflurane and maintained
at 1.5–2% isoflurane. Deep anesthesia was confirmed by the complete
loss of the pedal withdrawal reflex and corneal reflex before
initiating the blood collection. The blood was immediately
collected into tubes containing EDTA. Peripheral blood mononuclear
cells (PBMCs) were isolated using density gradient centrifugation
with Ficoll-Paque™ PLUS (cat. no. 17-1440-02; GE
Healthcare) according to standard protocols. Briefly, the collected
whole blood samples were diluted 1:1 with PBS and carefully layered
onto lymphocyte separation medium. Samples were centrifuged at 800
× g for 30 min at room temperature. After centrifugation, the
mononuclear cell layer was collected, transferred to a 15 ml tubes
and washed twice with PBS at 250 × g for 10 min at room
temperature. For intracellular cytokine staining, PBMCs were
stimulated with Cell Stimulation Cocktail plus protein transport
inhibitors (cat. no. 00-4975-93; 1:500; eBioscience; Thermo Fisher
Scientific, Inc.) for 6 h at 37°C in a 5% CO2 incubator.
Cells were filtered through a 30 µm CELLTrics® filter
(Sysmex Corporation) and resuspended in staining buffer. Surface
staining was performed using CD4 antibody (cat. no. MA1-146; 1:100;
Invitrogen; Thermo Fisher Scientific, Inc.) for 30 min at 4°C in
the dark. After washing, the cells were fixed and permeabilized
using the Intracellular Fixation & Permeabilization Buffer Set
(cat. no. 88-8824-00; eBioscience; Thermo Fisher Scientific, Inc.)
for 30 min at room temperature. Subsequently, the cells were
incubated with IL-17 antibodies (cat. no. 45-7177-80; 1:100;
Invitrogen; Thermo Fisher Scientific, Inc.) were added to the cell
suspension, mixed gently by pulse vortex and incubated for 30 min
in the dark at 4°C. After washing, PBMCs were centrifuged at 300 ×
g for 5 min at 4°C and resuspended in PBS for analysis and cell
sorting.
Flow cytometric analysis and cell sorting were
performed using a BD FACSAria™ III cell sorter (BD
Biosciences) equipped with a 100 µm nozzle at 20 psi. Compensation
was performed using single-stained controls. Dead cells were
excluded using 4′,6-diamidino-2-phenylindole (DAPI; Thermo Fisher
Scientific, Inc.) staining at room temperature for 5 min.
Fluorescence signals were detected using the 488 nm laser with a
530/30 nm filter for Alexa Fluor™ 488 (CD4), a 695/40 nm
filter for PerCP-Cy5.5 (IL-17A), and the 355 nm laser with a 450/50
nm filter for DAPI. Data were analyzed using FlowJo software
(version 10.8.1; FlowJo, LLC; BD Biosciences). Gating was
established using isotype controls and fluorescence minus one
controls. CD4+IL-17A+ cells were gated within the lymphocyte
population based on forward and side scatter characteristics.
Sorted cells were collected into 1.5 ml RNase-free
tubes pre-coated with 1% BSA in PBS containing RNasin®
Plus RNase inhibitor (1:25 dilution) for downstream analysis.
Enzyme-linked immunosorbent assay
(ELISA)
To quantitatively assess systemic inflammatory
cytokine levels, whole blood samples were collected into sterile
tubes without anticoagulant and allowed to clot at room temperature
for 20 min. Samples were centrifuged at 1,000 × g for 20 min at 4°C
and the serum supernatants were carefully collected and stored at
−80°C until analysis. Serum cytokine concentrations were measured
using commercial ELISA kits (Jiangsu Meimian Industrial Co., Ltd.),
according to the manufacturer's instructions. The following kits
were used: IFN-γ (cat. no. MM-0182M1), TNF-α (cat. no. MM-0132M1),
IL-17A (cat. no. MM-0759M1) and IL-6 (cat. no. MM-0163M1). Briefly,
serum samples and standards were added to antibody-coated 96-well
microplates and incubated for 30 min at 37°C. After washing to
remove unbound proteins enzyme-conjugated detection antibodies were
added and incubated for 30 min at 37°C. Following substrate
addition, color development was conducted for 15 min at 37°C and
terminated using stop solution and absorbance was measured at 450
nm using a microplate reader. Cytokine concentrations were
calculated from standard curves generated using recombinant
standards provided in the kits. All samples were analyzed in
duplicate and mean values were used for statistical analysis.
TUNEL staining
Apoptosis in lacrimal gland tissues was assessed
using a terminal deoxynucleotidyl transferase dUTP nick end
labeling (TUNEL) assay. Tissues were fixed in 4% PFA for 24 h at
room temperature. Paraffin-embedded tissue sections (4-µm
thickness) were deparaffinized, rehydrated and permeabilized with
0.1% Triton X-100 for 5 min at room temperature. Sections were
blocked with 5% BSA for 30 min at room temperature and washed with
PBS. TUNEL staining was performed using a commercial kit (cat. no.
RC-01; http://www.recordbio.com; Shanghai
Recordbio Biological Technology Co., Ltd.) according to the
manufacturer's instructions. Sections were incubated with the TUNEL
reaction mixture for 1 h and counterstained with DAPI (1 µg/ml; 5
min at room temperature). Sections were then mounted using antifade
mounting medium (cat. no. S2100; Beijing Solarbio Science &
Technology Co., Ltd.).
Images were captured using a fluorescence microscope
(TS100-F; Nikon Corporation). Apoptosis was quantified by
calculating the percentage of TUNEL+ cells compared with
the total number of DAPI-stained nuclei. At least 3 randomly
selected non-overlapping fields per section were analyzed using
ImageJ software (Version 1.48v; National Institutes of Health).
Immunofluorescence (IF) staining
To evaluate immune cell infiltration and spatial
co-localization in lacrimal gland tissues, paraffin sections
(thickness, 4 µm) were fixed in 4% PFA for 10 min at room
temperature, permeabilized with 0.1% Triton X-100 and blocked with
5% BSA for 30 min at room temperature.
Sections were incubated overnight at 4°C with
primary antibodies against CD4 (cat. no. MA1-146; 1:20; Invitrogen;
Thermo Fisher Scientific, Inc.) and IL-17A (cat. no. sc-374218;
1:50; Santa Cruz Biotechnology, Inc.). After washing, sections were
incubated for 45 min at room temperature in the dark with Alexa
Fluor™ 488-conjugated secondary antibodies (cat. no.
A-21121; 1:500; Invitrogen; Thermo Fisher Scientific, Inc.) or
Alexa Fluor™ 594 (cat. no. A-11007; 1:500; Invitrogen;
Thermo Fisher Scientific, Inc.). Nuclei were counterstained with
DAPI (1 µg/ml) for 5 min at room temperature.
Fluorescent and phase-contrast images were captured
using a Zeiss Axiovert 200 inverted microscope equipped with
AxioCam MRm digital camera and analyzed using AxioVision software
(version 4.9.1; Carl Zeiss AG). Confocal images were acquired using
a Leica DM6000 B confocal laser scanning microscope (Leica
Microsystems GmbH).
Statistical analysis
All experiments were performed using at least three
independent biological replicates unless stated otherwise. Data are
presented as mean ± standard deviation (SD). Statistical analyses
were conducted using GraphPad Prism (version 8.0; Dotmatics). For
comparisons between two independent groups, an unpaired two-tailed
Student's t-test was used. For comparisons among three groups,
one-way analysis of variance followed by Tukey's multiple
comparisons test was performed. P<0.05 was considered to
indicate a statistically significant difference.
Results
Characterization of dry eye-like
manifestations and systemic inflammation in NOD/Ltj mice
Phenol red thread assay demonstrated significantly
reduced tear secretion in NOD/Ltj mice compared with ICR controls
at all assessed time points (all P<0.001), with a mean decrease
>2 mm (Fig. 1A). Serum cytokine
analysis by ELISA revealed significantly elevated levels of IFN-γ,
IL-17A and TNF-α in NOD/Ltj mice compared with ICR mice
(P<0.05), whereas IL-6 levels did not differ significantly
between groups (Fig. 1B).
Histopathological examination of lacrimal gland sections stained
with H&E demonstrated marked glandular atrophy and prominent
lymphocytic infiltration in NOD/Ltj mice compared with ICR controls
(Fig. 1C). These findings
confirmed the presence of lacrimal gland dysfunction and systemic
inflammatory activation in NOD/Ltj mice.
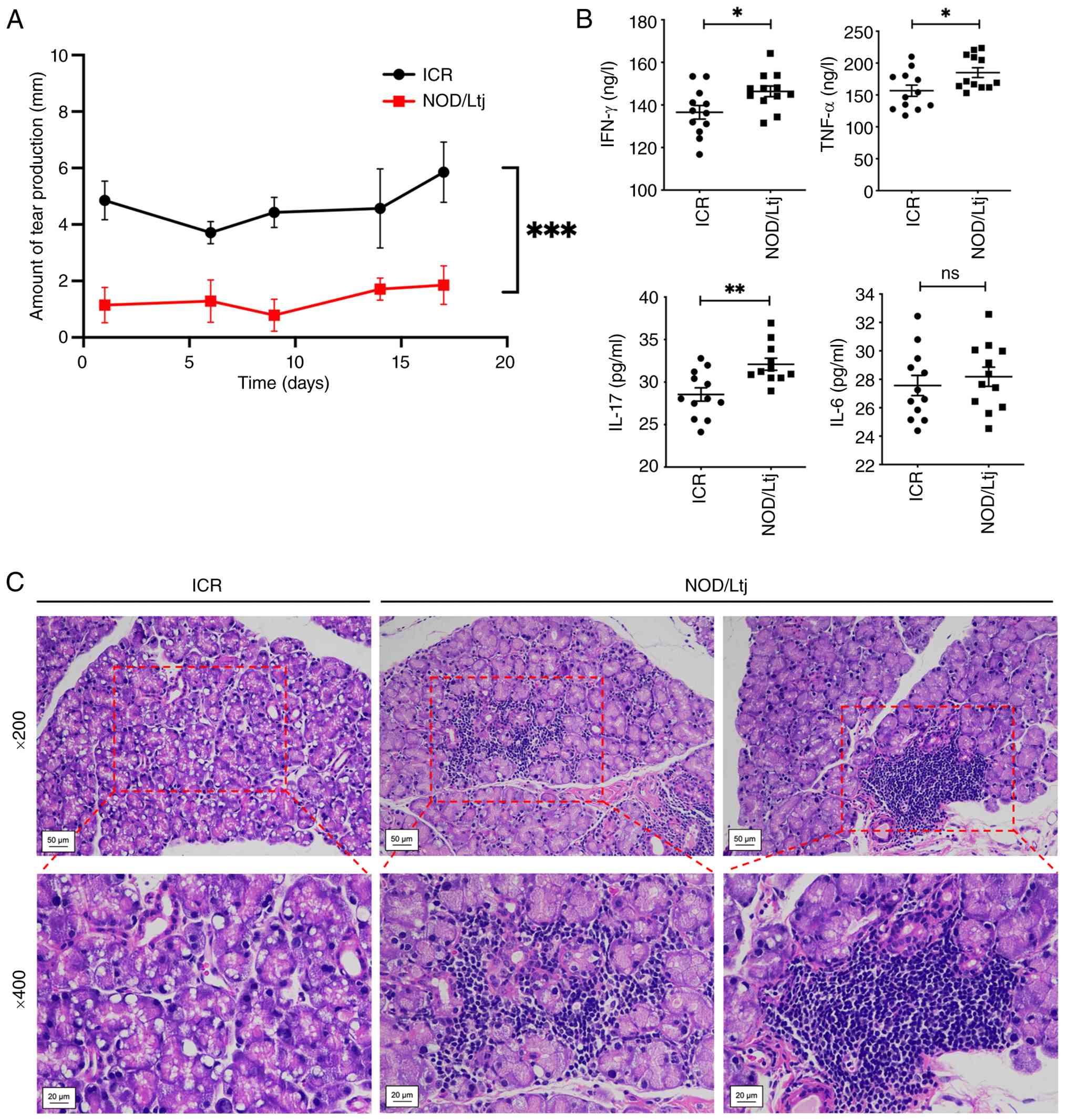 | Figure 1.Characterization of lacrimal gland
dysfunction and systemic inflammation in NOD/Ltj mice. (A) Tear
secretion measured by the phenol red thread test. (B) Serum levels
of IFN-γ, IL-17A, TNF-α and IL-6 determined by enzyme-linked
immunosorbent assay. (C) Representative hematoxylin and
eosin-stained lacrimal gland sections from ICR and NOD/Ltj mice
presenting glandular architecture and lymphocytic infiltration
(magnification, ×200 and ×400). Data are presented as mean ± SD
(n=12). *P<0.05, **P<0.01 and ***P<0.001; ns vs. ICR
control. ns, not significant; ICR, Institute of Cancer Research;
IL-17A, interleukin-17A; IFN-γ, interferon-γ; TNF-α, tumor necrosis
factor-α; SD, standard deviation; NOD, non-obese diabetic. |
Transcriptomic profiling identifies
enrichment of NF-κB signaling and Th17 differentiation pathways in
lacrimal glands
To investigate molecular mechanisms associated with
reduced tear production in NOD/Ltj mice, mRNA-seq was performed on
lacrimal gland tissues. A total of 1,109 DEGs were identified
compared with ICR controls, including 874 upregulated and 235
downregulated genes. Differential expression patterns are presented
in the heatmap (Fig. 2A) and
volcano plot (Fig. 2C). GO
enrichment analysis categorized the DEGs into biological processes,
cellular components and molecular functions, with significant
enrichment in immune and inflammatory response-related categories,
specifically including ‘cellular response to interferon-β’,
‘positive regulation of T cell proliferation’, ‘T cell
differentiation’ and ‘adaptive immune response’ (Fig. 2B). KEGG pathway analysis
demBonstrated significant enrichment of pathways associated with
‘NF-κB signaling’ and ‘Th17 cell differentiation’ (Fig. 2D). The NF-κB signaling pathway
exhibited an adjusted P-value of 6.71×10−12 with a rich
factor of 0.29, whereas the Th17 differentiation pathway
demonstrated an adjusted P-value of 3.46×10−12 with a
rich factor of 0.29.
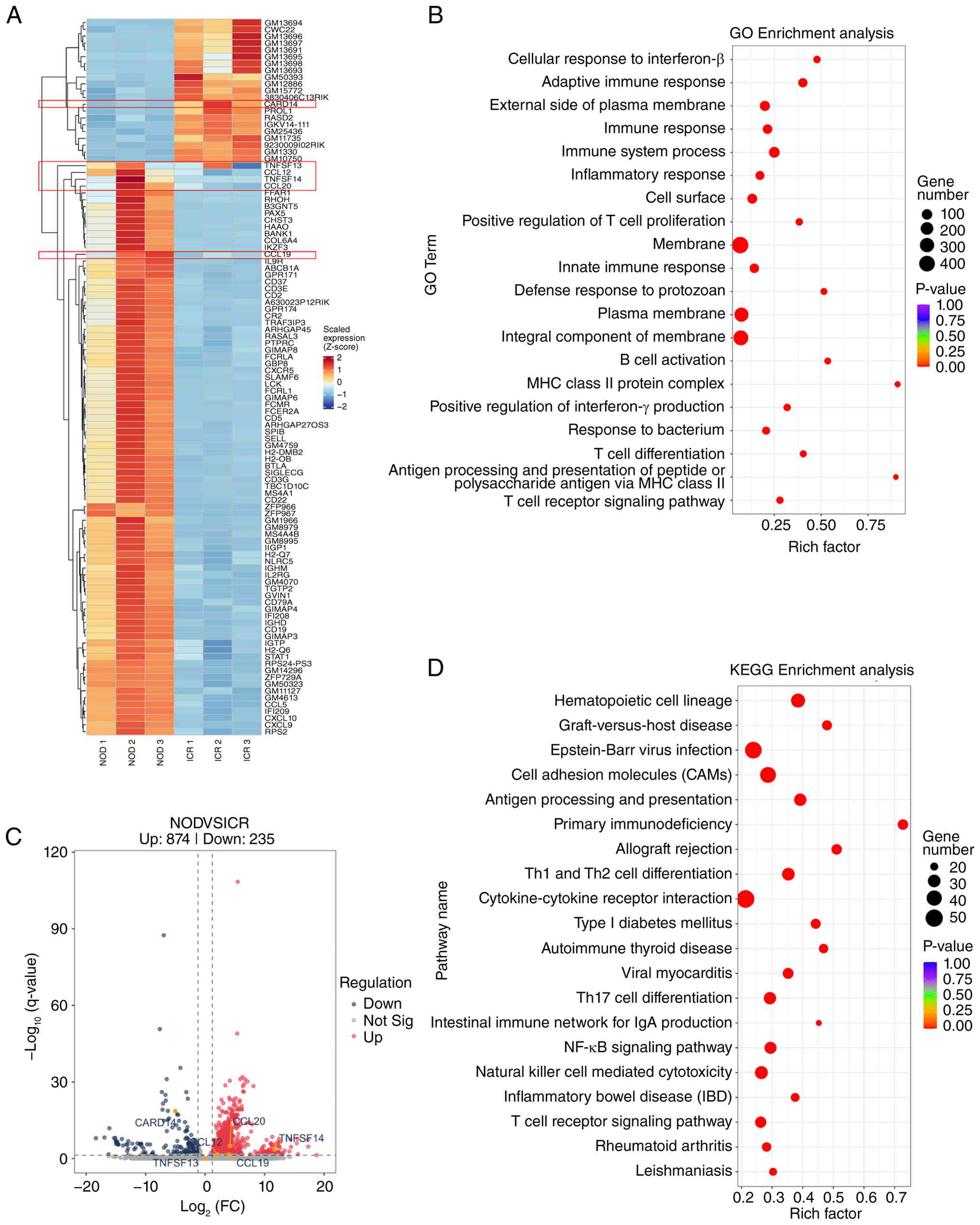 | Figure 2.Transcriptomic analysis of lacrimal
glands in NOD/Ltj mice. (A) Heatmap presenting DEGs between ICR and
NOD/Ltj mice. (B) GO enrichment analysis of DEGs. (C) Volcano plot
illustrates the distribution of upregulated and downregulated
genes. (D) Kyoto Encyclopedia of Genes and Genomes pathway
enrichment analysis. The six genes selected for subsequent reverse
transcription-quantitative PCR validation (CARD14, TNFSF14,
TNFSF13, CCL19, CCL20 and CCL12) are labeled directly in
(A) and (C). DEGs were defined as |log2 FC|>1 and
adjusted P-value <0.05. *P<0.05, **P<0.01 and ns. ns, not
significant; ICR, Institute of Cancer Research; DEGs,
differentially expressed genes; GO, Gene Ontology; FC, fold change;
NOD, non-obese diabetic. |
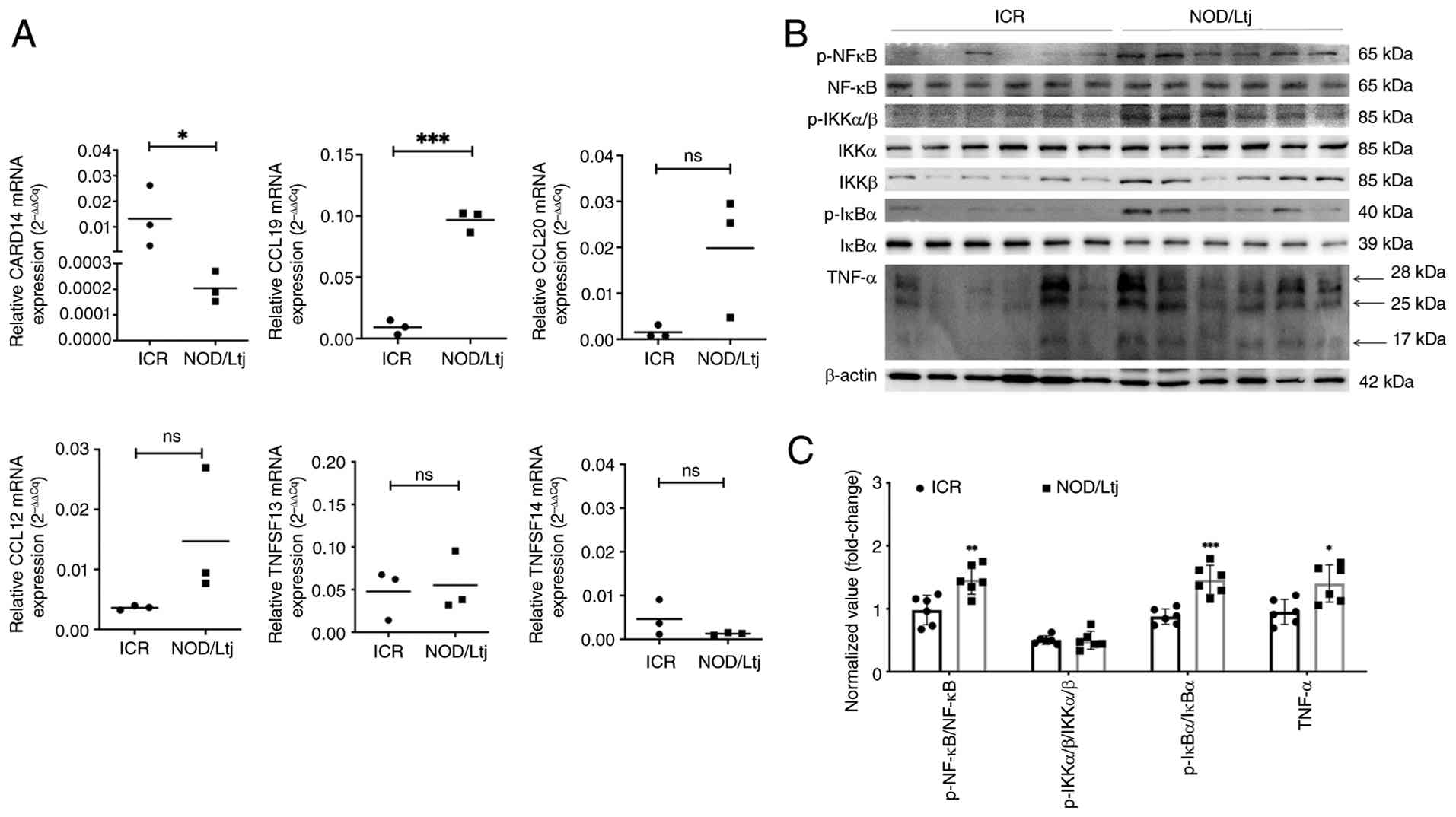
Based on pathway enrichment and differential
expression magnitude, six candidate genes were selected for
validation: One downregulated gene (CARD14:
log2FC, −5.02) and five upregulated genes
(TNFSF14: log2FC, 11.64; TNFSF13:
log2FC, 0.14; CCL19: log2FC, 3.99;
CCL20: log2FC, 4.18; CCL12:
log2FC, 2.09). Consistent with the mRNA-seq data, the
RT-qPCR analysis confirmed the significant downregulation of
CARD14 (P=0.0412) and upregulation of CCL19
(P=0.001), consistent with the sequencing results. The expression
levels of CCL12 (P=0.1464) and CCL20 (P=0.0762),
TNFSF13 (P=0.7954), and TNFSF14 (P=0.2265) also
showed trends consistent with the sequencing data, although these
changes did not reach statistical significance (Fig. 3A).
 | Figure 3.Validation of selected differentially
expressed genes and NF-κB pathway activation in lacrimal glands.
(A) Relative mRNA expression levels of CARD14, TNFSF14, TNFSF13,
CCL19, CCL20 and CCL12 determined by reverse
transcription-quantitative PCR. (B) Representative western blotting
images of NF-κB pathway-related proteins and downstream
inflammatory mediators. (C) Densitometric quantification of protein
expression normalized to β-actin. Data are presented as mean ± SD
(n=6). *P<0.05, **P<0.01 and ***P<0.005; ns vs. ICR
control. NF-κB, nuclear factor κB; ns, not significant; ICR,
Institute of Cancer Research; IKK, inhibitor of κB kinase; IκBα,
inhibitor of κB α; TNF-α, tumor necrosis factor-α; SD, standard
deviation; p, phosphorylated; NOD, non-obese diabetic. |
To further assess NF-κB pathway activation at the
protein level, western blotting analysis was performed. Expression
levels of NF-κB pathway components, specifically p-NF-κB (relative
to total NF-κB; P=0.0048) and p-IκBα (relative to total IκBα;
P=0.0003), were significantly increased in lacrimal glands of
NOD/Ltj mice compared with that of ICR controls. Furthermore, a
significantly elevated level of downstream inflammatory mediator
TNF-α was observed (P=0.0112) (Fig. 3B
and C). These findings indicated the activation of NF-κB
signaling in the lacrimal glands of NOD/Ltj mice.
Pharmacological inhibition of NF-κB
attenuates pathway activation and downstream inflammatory mediators
in NOD/Ltj mice
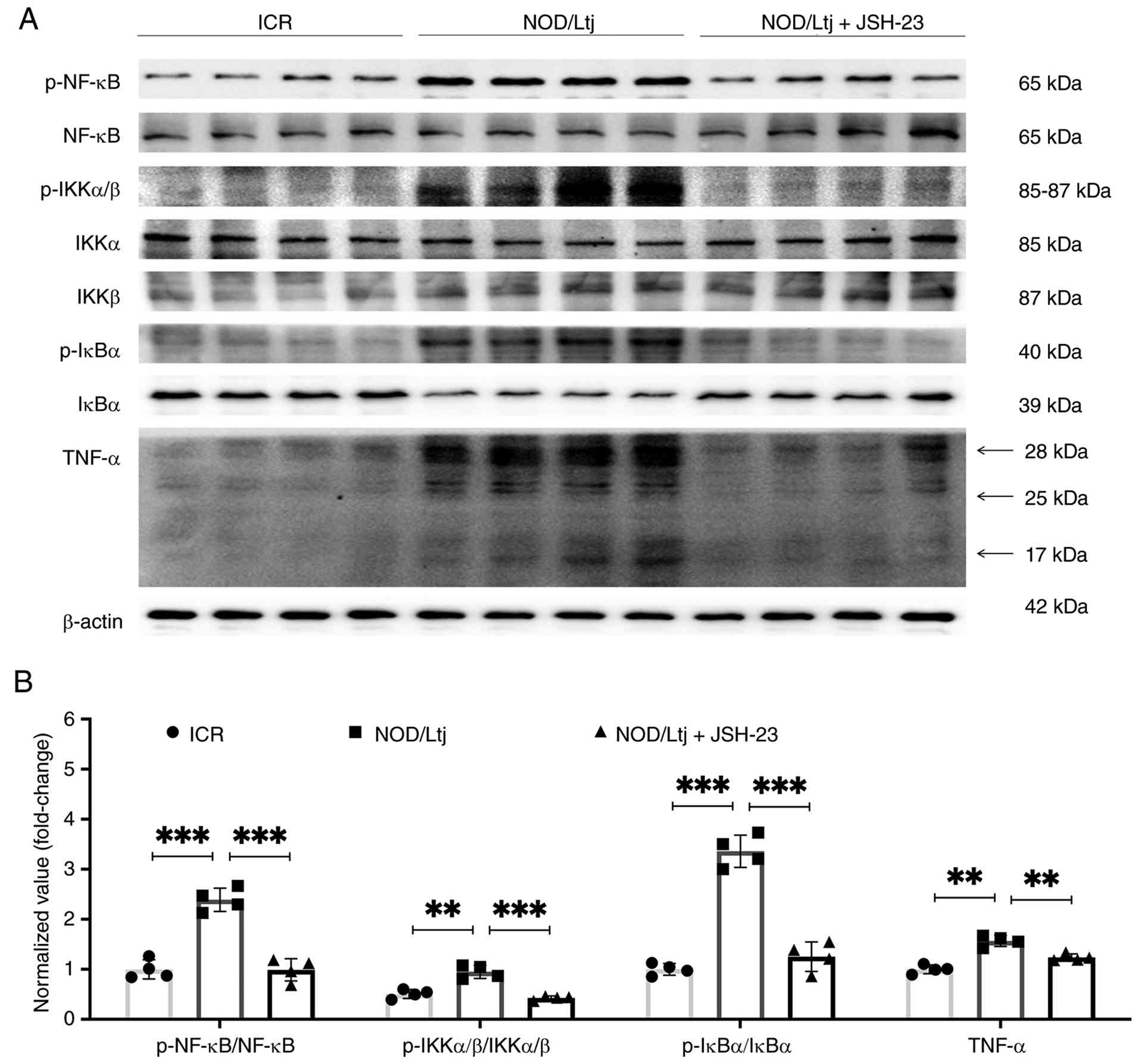
To evaluate the functional contribution of NF-κB
signaling in pSS-associated lacrimal gland pathology, the NF-κB
inhibitor JSH-23 was administered to NOD/Ltj mice. Western blotting
analysis demonstrated significantly increased p-NF-κB (P<0.001),
-IKKα/β (P=0.0004) and -IκBα (P=0.0002), accompanied by
significantly reduced total IκBα expression in NOD/Ltj mice
compared with ICR controls (Fig.
4A). Treatment with JSH-23 significantly reduced
phosphorylation of NF-κB pathway components (p-NF-κB, P=0.0003;
p-IKKα/β, P=0.0009; p-IκBα, P=0.0054 vs. NOD group) and
significantly restored IκBα expression (P=0.0023 vs. NOD group)
toward levels observed in ICR mice (compared to ICR controls:
p-NF-κB, P=0.1525; p-IKKα/β, P=0.0480; p-IκBα, P=0.5786; IκBα,
P=0.0878) (Fig. 4A and B).
Furthermore, the elevated expression level of downstream
inflammatory mediator TNF-α (P=0.0002 vs. ICR group) was
significantly reduced following JSH-23 treatment (P=0.0022 vs. NOD
group), showing no significant difference compared with the ICR
controls (P=0.0586) (Fig. 4A and
B). These findings indicated that pharmacological inhibition of
NF-κB suppresses pathway activation and downstream inflammatory
signaling in NOD/Ltj lacrimal glands.
 | Figure 4.JSH-23 suppresses NF-κB pathway
activation in lacrimal glands of NOD/Ltj mice. (A) Representative
western blotting images presenting expression levels of NF-κB
pathway-related proteins and downstream inflammatory mediators in
ICR, NOD/Ltj and NOD/Ltj + JSH-23 groups. (B) Densitometric
quantification of protein expression normalized to β-actin. Data
are presented as mean ± SD (n=4). **P<0.01 and ***P<0.001 vs.
NOD/Ltj group; ns vs. ICR control. NF-κB, nuclear factor κB; ns,
not significant; SD, standard deviation; ICR, Institute of Cancer
Research; IKK, inhibitor of κB kinase; IκBα, inhibitor of κB α;
TNF-α, tumor necrosis factor-α; p, phosphorylated; NOD, non-obese
diabetic. |
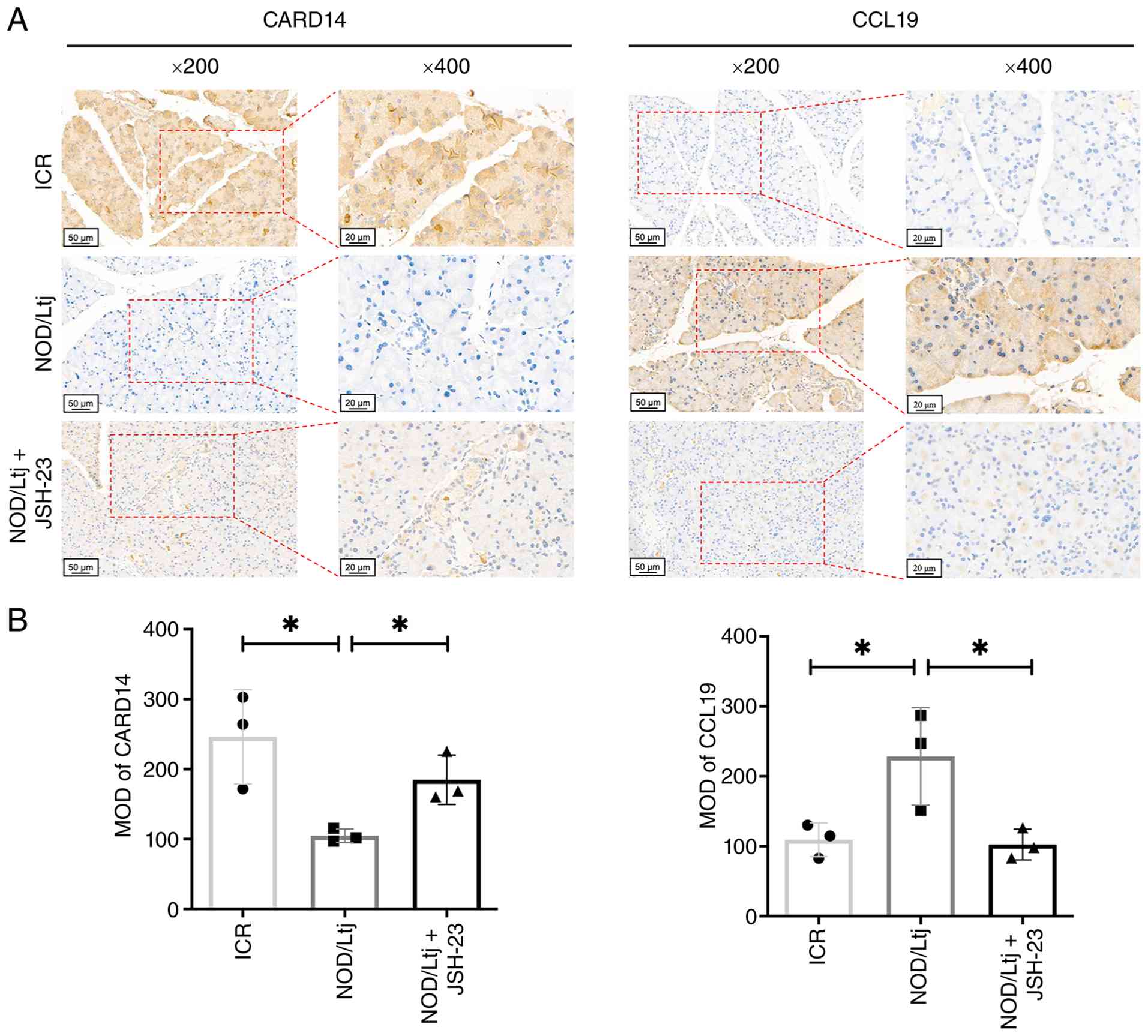
To assess in situ protein expression of
selected DEGs, immunohistochemical staining was performed (Fig. 5A). Compared with ICR mice, NOD/Ltj
mice exhibited significantly decreased CARD14 protein expression
(P=0.0228) and significantly increased CCL19 expression (P=0.0489)
in lacrimal gland tissues. Administration of JSH-23 significantly
reversed these alterations (CARD14, P=0.0192; CCL19, P=0.0405 vs.
NOD group), restoring expression levels similar to those in ICR
controls (CARD14, P=0.2349; CCL19, P=0.7356) (Fig. 5B). Therefore, these results
demonstrated that NF-κB inhibition modulates CARD14 and CCL19
expression in lacrimal gland tissues of NOD/Ltj mice.
NF-κB inhibition reduces systemic Th17
cell frequency and inflammatory cytokine production in NOD/Ltj
mice
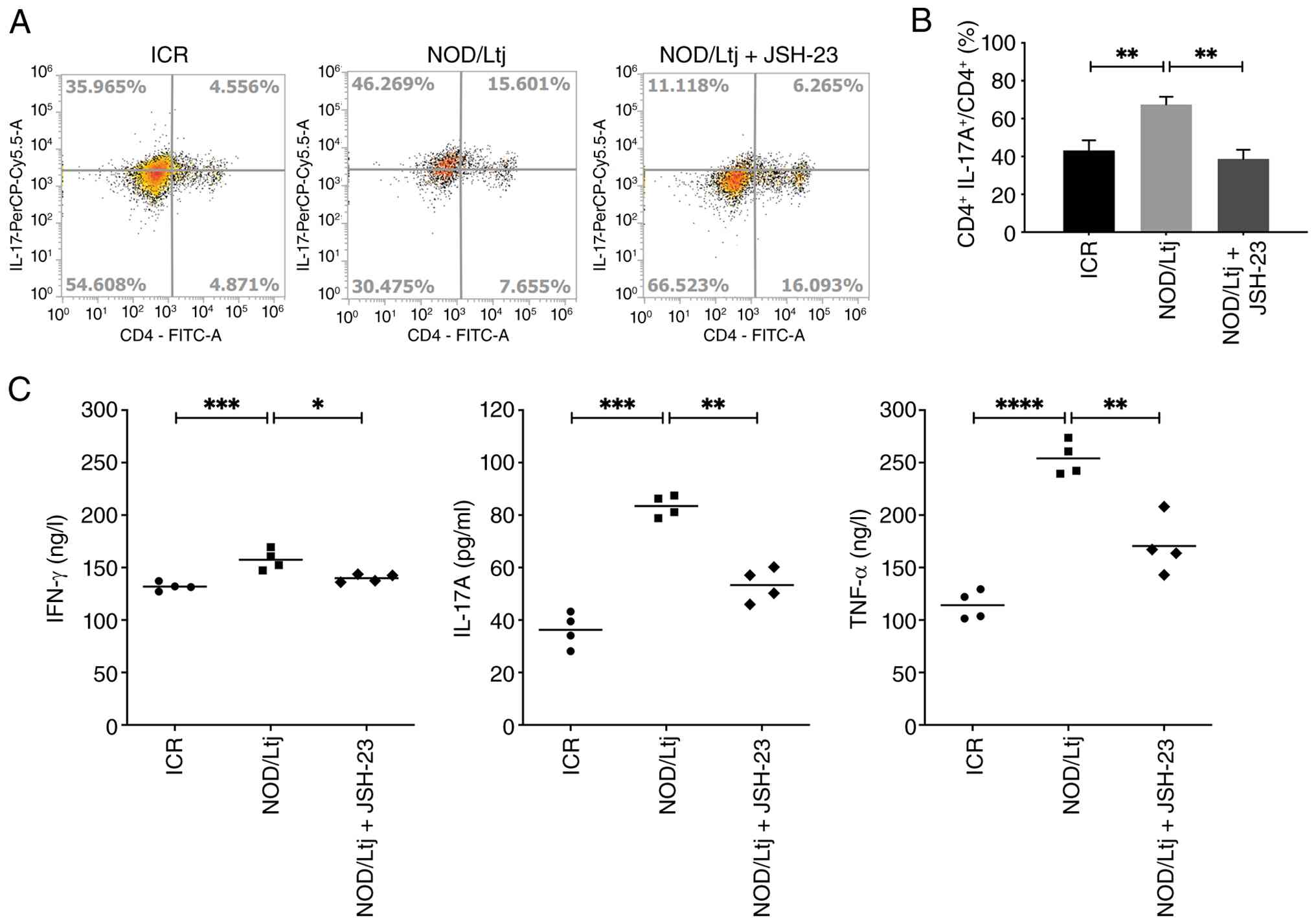
To evaluate the effect of NF-κB inhibition on
systemic immune responses, PBMCs were analyzed by flow cytometry to
quantify CD4+ T cells and CD4+IL-17A+ Th17 cells (Fig. 6A and B). The proportion of
CD4+IL-17A+ cells within the CD4+ T cell population was
significantly increased in NOD/Ltj mice compared with ICR controls
(P=0.0047), indicating the expansion of Th17 cells. Treatment with
JSH-23 significantly reduced the CD4+IL-17A+/CD4+ ratio compared
with the untreated NOD/Ltj mice (P=0.0011), restoring levels toward
those observed in ICR controls.
 | Figure 6.JSH-23 reduces systemic Th17 cell
frequency and inflammatory cytokine production in NOD/Ltj mice. (A)
Representative flow cytometry plots presenting CD4+ and CD4+IL-17A+
cells in peripheral blood mononuclear cells from ICR, NOD/Ltj and
NOD/Ltj + JSH-23 groups. (B) Quantitative analysis of the
proportion of CD4+IL-17A+ cells within the CD4+ T cell population.
(C) Serum levels of IFN-γ, IL-17A and TNF-α measured by
enzyme-linked immunosorbent assay. Data are presented as mean ± SD
(n=6). *P<0.05, **P<0.01, ***P<0.001 and ****P<0.0001
vs. NOD/Ltj group; ns, not significant vs. ICR control. SD,
standard deviation; Th17, T helper 17; NOD, non-obese diabetic;
ICR, Institute of Cancer Research; IL-17A, interleukin-17A; IFN-γ,
interferon-γ; TNF-α, tumor necrosis factor-α. |
Consistent with changes in Th17 cell frequency,
ELISA demonstrated significantly elevated serum levels of IFN-γ
(P=0.0009), IL-17A (P=0.0002) and TNF-α (P<0.0001) in NOD/Ltj
mice compared with ICR controls (Fig.
6C). Administration of JSH-23 significantly reduced the
cytokine levels compared with untreated NOD/Ltj mice (IFN-γ,
P=0.0151; IL-17A, P=0.0099; and TNF-α, P=0.0019), and restored them
to levels with no significant differences compared with the ICR
controls (P=0.0590, P=0.0599 and P=0.0502, respectively),
indicating suppression of systematic inflammatory cytokine
production.
NF-κB inhibition reduces local Th17
cell infiltration in lacrimal glands
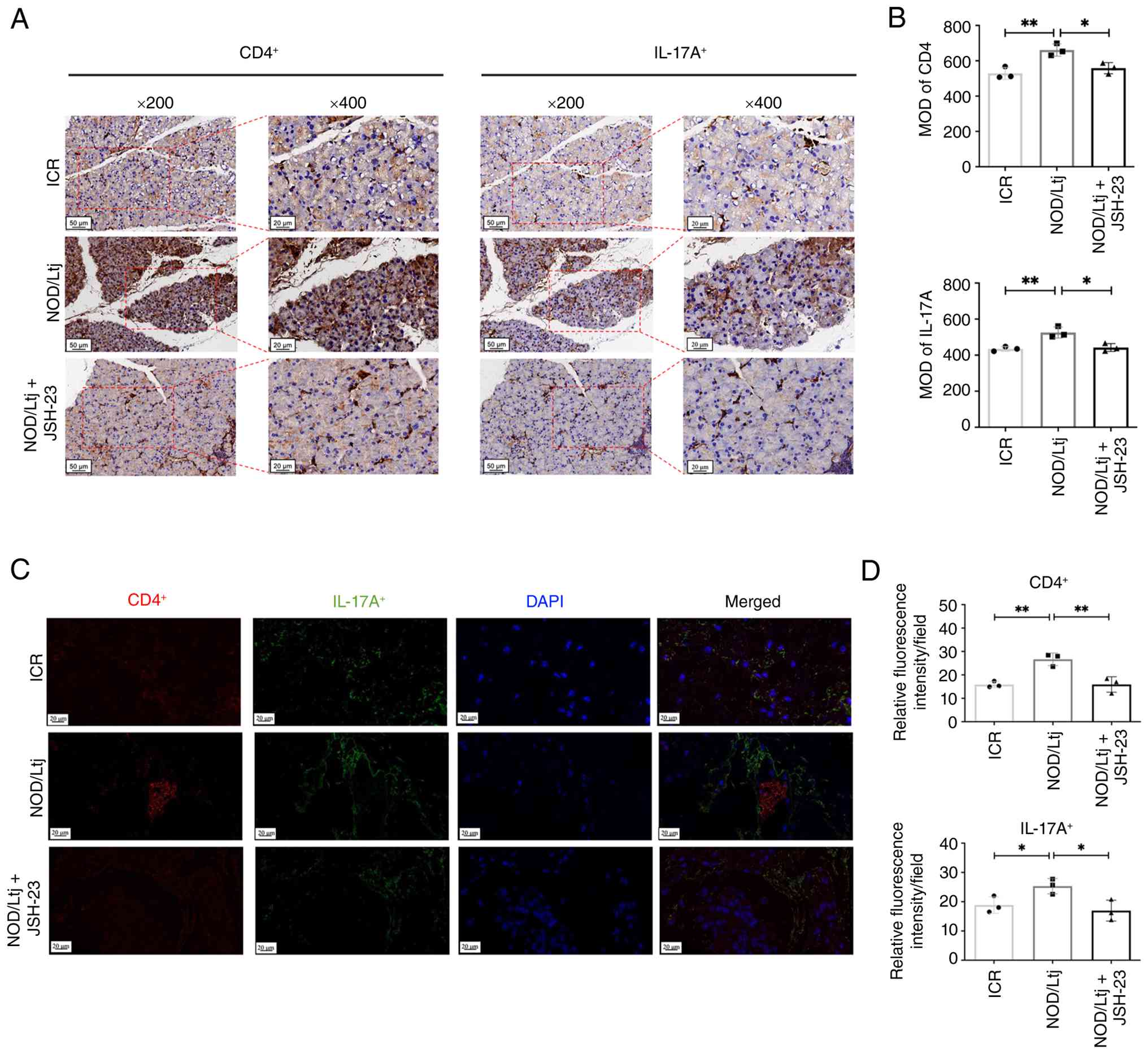
To evaluate local immune cell infiltration in
lacrimal gland tissues, IHC and IF analyses were performed
(Fig. 7). IHC demonstrated
significantly increased CD4+ T cell (P=0.0091) and IL-17A
expression (P=0.0095) in lacrimal glands of NOD/Ltj mice compared
with ICR controls (Fig. 7A and B).
Treatment with JSH-23 significantly reduced CD4+ and
IL-17A staining intensity (P=0.0190 and P=0.0170 vs. NOD group,
respectively), restoring levels toward those observed in the ICR
mice. Consistently, IF staining confirmed significantly increased
infiltration of CD4+ (P=0.0033) and IL-17A+
(P=0.0434) cells in the lacrimal gland tissues of NOD/Ltj mice,
which was significantly attenuated following JSH-23 administration
(P=0.0042 and P=0.0302 vs. NOD group, respectively), showing no
significant differences compared with the ICR baseline (P=0.9849
and P=0.5101, respectively) (Fig. 7C
and D). These findings indicated that inhibition of NF-κB
signaling reduces local Th17-associated inflammatory infiltration
in lacrimal glands of NOD/Ltj mice.
 | Figure 7.JSH-23 reduces local Th17 cell
infiltration in lacrimal glands of NOD/Ltj mice. (A) Representative
immunohistochemical staining of CD4+ and IL-17A+ cells in lacrimal
glands (magnification, ×200 and ×400). (B) Quantitative analysis of
CD4+ and IL-17A+ cell infiltration based on mean optical density
(MOD). (C) Representative immunofluorescence staining of CD4+ T
cells and IL-17A in lacrimal gland sections from ICR, NOD/Ltj and
NOD/Ltj + JSH-23 groups (scale bars, 20 µm). (D) Quantitative
analysis of CD4+ and IL-17A+ fluorescence
intensity. Data are presented as mean ± SD (n=4 per group).
*P<0.05 and **P<0.01 vs. NOD/Ltj group; ns, not significant
vs. ICR control. MOD, mean optical density; SD, standard deviation;
NOD, non-obese diabetic; ICR, Institute of Cancer Research; Th17, T
helper 17; IL-17A, interleukin-17A. |
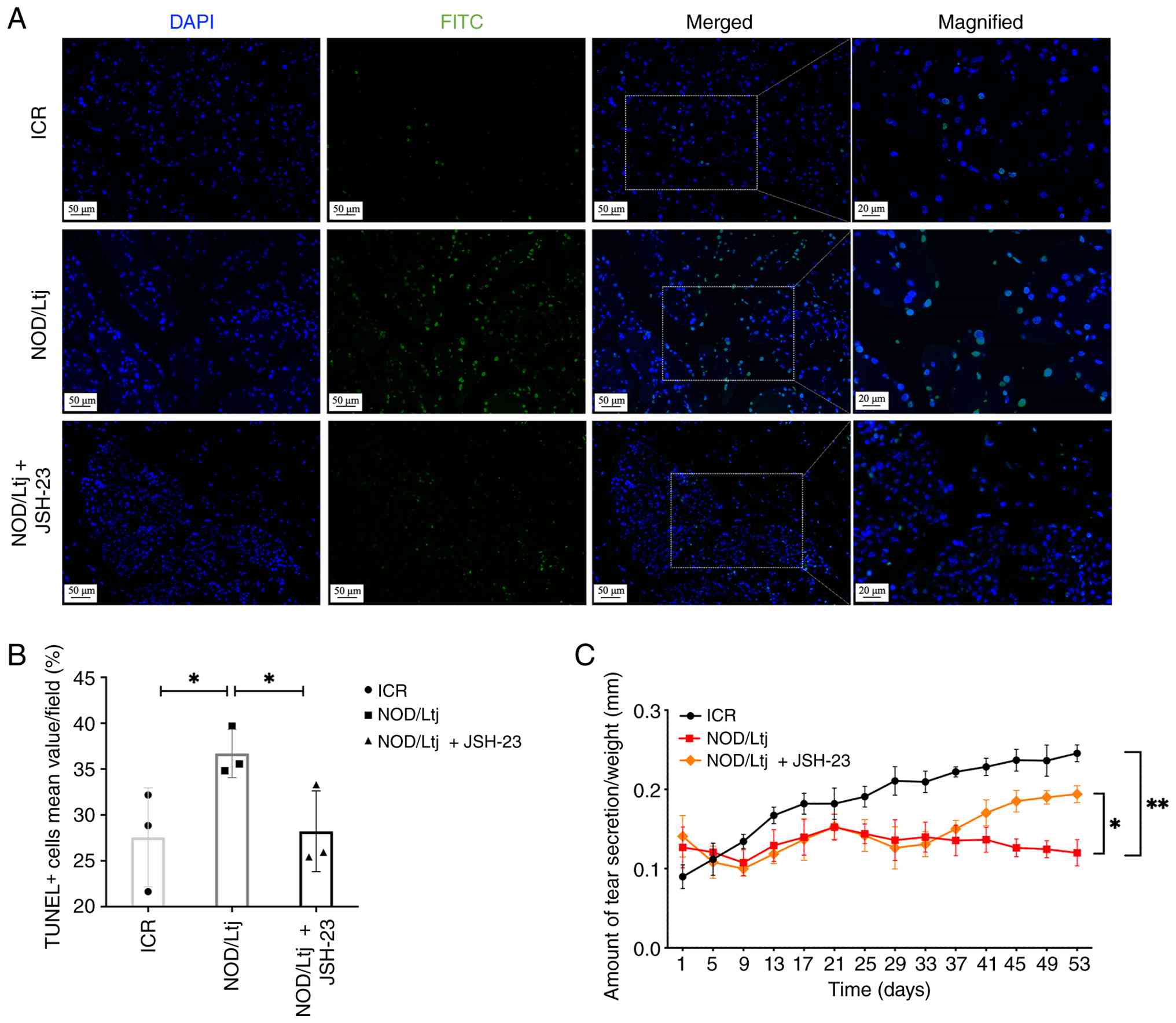
NF-κB inhibition reduces lacrimal
gland apoptosis and partially restores tear secretion
TUNEL staining demonstrated a significantly higher
proportion of apoptotic cells in lacrimal gland tissues of NOD/Ltj
mice compared with ICR controls (P=0.0476; Fig. 8A and B). Quantitative analysis
demonstrated that JSH-23 treatment reduced the percentage of
TUNEL+ cells compared with untreated NOD/Ltj mice
(P=0.0428), restoring apoptosis levels toward those observed in ICR
controls (P=0.8763).
To assess the functional impact of NF-κB inhibition
on tear secretion, phenol red thread assay was performed. Tear
production in NOD/Ltj mice was significantly reduced compared with
ICR mice at the final assessment time point (53 days; P=0.0038)
(Fig. 8C). Treatment with JSH-23
significantly increased tear production compared with untreated
NOD/Ltj mice (P=0.0320), indicating partial restoration of lacrimal
gland secretory function.
Discussion
Due to the technical challenges associated with
lacrimal gland biopsy in humans, the diagnosis of pSS and studies
of target organ injury have largely relied on minor salivary gland
biopsy specimens (23–25). Therefore, investigations on
lacrimal gland pathology in pSS-related dry eye has predominantly
depended on experimental mouse models that effectively bridge this
clinical knowledge gap. The NOD/Ltj mouse is extensively used as a
spontaneous model of pSS, because it recapitulates key clinical and
immunological features of the disease in patients, including
reduced tear secretion, lymphocytic infiltration of exocrine glands
and elevated systemic inflammatory cytokines (15). Notably, sex-specific differences
have been described in this model, with prominent dacryoadenitis
observed in men and sialadenitis in women (16). In the present study, male NOD/Ltj
mice exhibited marked lacrimal gland dysfunction characterized by
reduced tear production, extensive lymphocytic infiltration and
germinal center-like structures, accompanied by elevated serum
levels of IFN-γ, TNF-α and IL-17A. These findings were consistent
with previous studies regarding human pSS immunopathology and
confirmed the suitability of this model in investigating the
specific mechanisms of lacrimal gland injury in pSS (26–28).
Previous transcriptomic studies in pSS have
primarily focused on salivary gland tissue (29,30)
and peripheral blood (31–33), with limited analysis of lacrimal
gland-specific molecular alterations (34,35).
Although earlier microarray approaches have identified pathways
associated with dacryoadenitis in NOD mice (36), a high resolution transcriptomic
characterization of the lacrimal gland microenvironment remains to
be elucidated. In the present study, the application of
high-throughput mRNA-seq allowed for a more sensitive and
comprehensive characterization of the lacrimal gland transcriptome.
The present study data revealed significant enrichment of immune
activation and inflammatory response pathways, with prominent
involvement of ‘NF-κB signaling’ and ‘Th17 cell differentiation’
pathways. While these pathways are recognized as key drivers of pSS
pathogenesis in systemic and salivary gland contexts, the present
study findings provide specific transcriptomic evidence of their
coordinated dysregulation within the lacrimal gland.
Several key DEGs were selected for validation based
on their functional roles in the NF-κB pathway and immune
regulation. CARD14 was selected for its pivotal role as an
activator of the NF-κB pathway, mediating inflammatory responses
through signaling complex formation (37). TNFSF14 and TNFSF13
were included due to their involvement in immune regulation via
NF-κB: TNFSF14 promotes T-cell activation and Th17 responses
through NF-κB-dependent pathways (38,39),
while TNFSF13 contributes to B cell hyperactivity (40). Chemokines CCL19, CCL20 and
CCL12 were chosen for their roles in immune cell recruitment
and Th17 cell migration. Specifically, CCL20 facilitates
Th17 trafficking via CCR6 (41),
CCL19 regulates T- and dendritic cell migration through CCR7
(42), and CCL12 facilitates
monocyte recruitment via CCR2 (43).
Among the DEGs, CARD14 and CCL19 were
selected for further validation. CARD14 is a scaffold protein
predominantly expressed in epithelial cells (44) and known to facilitate NF-κB
activation through assembly of signaling complexes (45). While gain-of-function mutations in
the CARD14 can disrupt its autoinhibitory conformation and promote
aberrant NF-κB activation in inflammatory disorders such as
psoriasis (46), the present study
findings revealed a distinct expression pattern in the pSS lacrimal
gland. Specifically, reduced CARD14 expression was observed in
untreated NOD/Ltj lacrimal glands. Due to the predominant
epithelial localization of CARD14, its downregulation likely
reflects epithelial cell loss secondary to chronic inflammatory
injury rather than reduced pathway activity. Following JSH-23
administration, the reduction in glandular apoptosis was
accompanied by a marked restoration in CARD14 expression. This
restoration suggested that CARD14 levels may serve as a sensitive
indicator of epithelial structural integrity. The alignment of
CARD14 restoration with the observed decrease in TUNEL+
cells reflects an overall improvement in epithelial cell survival
following the attenuation of local inflammation.
By contrast, CCL19 was markedly upregulated
in NOD/Ltj lacrimal glands. CCL19 is a chemokine that actively
recruits lymphocytes and contributes to immune cell accumulation in
inflamed tissues (47). While the
involvement of CCL19 has been reported in the salivary glands of
patients with pSS (30), its
specific regulation within the lacrimal gland microenvironment
remains less characterized. NF-κB has been reported to regulate
CCL19 transcription under inflammatory conditions (48,49).
The present study findings supported this regulatory relationship
in the lacrimal gland, as NF-κB activation was associated with
elevated CCL19 expression and increased immune cell infiltration.
Pharmacological inhibition of NF-κB significantly reduced CCL19
levels, suggesting that suppression of chemokine-driven immune
recruitment represents a key mechanism underlying the
anti-inflammatory effects of JSH-23 within the pSS lacrimal
gland.
Increasing evidence supports a key role for NF-κB
signaling in target organ injury in pSS (50). In salivary glands, NF-κB activation
promotes lymphocytic infiltration and pro-inflammatory cytokine
production (5,51,52).
In the present study, the western blotting analysis demonstrated
hyperactivation of the canonical NF-κB pathway in lacrimal glands
of NOD/Ltj mice, as evidenced by increased phosphorylation of NF-κB
p65, IKKα/β and IκBα. These findings extend previous observations
to the lacrimal gland and support a key role for NF-κB mediated
inflammatory signaling in glandular dysfunction (50,51,53).
Therapeutic targeting of NF-κB has been proposed as a strategy to
attenuate glandular inflammation while preserving epithelial
integrity (54). Modulation of
NF-κB pathway components, including IKKε and IκBα, as well as
associations with anti-SS-related antigen A autoantibodies, have
been reported in pSS (55,56). Small-molecule inhibitors with
NF-κB-modulatory properties, such as iguratimod, have advanced to
clinical evaluation (54).
Clinical studies indicated that iguratimod, when combined with
hydroxychloroquine and glucocorticoids, improves xerostomia and
xerophthalmia, reduces disease activity and does not increase
adverse events, including gastrointestinal symptoms, liver function
abnormalities and leukopenia (57–59).
By contrast, other targeted therapies, such as the spleen tyrosine
kinase inhibitor GS-9876, have not demonstrated notable clinical
benefit in pSS (60). Although
these clinical findings supported the therapeutic potential of
NF-κB modulation, the protective mechanisms operating within the
lacrimal gland remain incompletely defined.
Dry eye in pSS is a marked lack of aqueous tears
caused by lacrimal gland destruction. During the experimental
design phase, the potential use of JSH-23 as a topical eye drop was
carefully evaluated. Previous studies have demonstrated that
targeting the NF-κB pathway effectively mitigates ocular surface
inflammation (61–63). However, those interventions
primarily target ocular surface epithelia. Anatomically, the
lacrimal gland is an extraocular exocrine gland located deep within
the orbit, producing aqueous tears that flow directionally onto the
ocular surface via excretory ducts. Due to this directional fluid
flow and deep anatomical location, topically applied eye drops
possess a limited ability to penetrate retrogradely to reach the
glandular microenvironment. Therefore, a topical formulation would
not effectively treat the primary glandular lesions characteristic
of pSS. Therefore, systemic administration was selected to address
the underlying etiology. Unlike intraocular tissues protected by
the blood ocular barrier, the extraocular lacrimal gland is highly
vascularized in the orbit. Systemic oral administration utilizes
the bloodstream to deliver the therapeutic agent directly into this
target tissue. Although precise ocular bioavailability and absolute
local drug concentrations were not directly quantified via
pharmacokinetic measurements in the present study, pharmacological
inhibition of NF-κB using JSH-23 suppressed pathway activation in
the lacrimal glands and was associated with reduced systemic levels
of IFN-γ, TNF-α and IL-17A. Previous studies suggested that
glandular epithelial cells may serve as initiators of inflammatory
cascades in pSS, contributing to a self-perpetuating cycle of
tissue injury and immune activation (64–66).
NF-κB signaling has been implicated in epithelial apoptosis and
inflammatory amplification (51).
Consistent with this framework, the present study demonstrated that
JSH-23 treatment reduced lacrimal gland apoptosis and partially
restored tear secretion. This restoration aligns with the recovered
expression level of CARD14, suggesting that attenuation of NF-κB
mediated inflammation may contribute to improved epithelial
survival and glandular function by disrupting the local
inflammatory vicious cycle.
In addition to tear volume, the biochemical
composition of tears serves a key role in ocular surface health.
The lacrimal gland secretes the aqueous layer of the tear film
containing functional proteins such as lactoferrin and lysozyme,
while mucins are primarily produced by the ocular surface
epithelium (67). In pSS,
autoimmune destruction of the lacrimal gland leads to deficiency in
these protective proteins. These alterations directly disrupt the
ocular surface microenvironment and compromise tear film stability.
Despite the high clinical relevance of these changes, the severe
aqueous deficiency in the NOD Ltj mouse model precluded the
collection of sufficient tear fluid for reliable quantitative
analysis in the present study. Future research utilizing
specialized microvolume proteomic technologies are warranted to
comprehensively evaluate these compositional changes and identify
potential biomarkers (68).
Th17 cells represent a key effector subset in
pSS-associated target organ damage. Increased Th17 cell
infiltration has been documented in salivary glands of patients
with pSS and has been associated with disease severity (69,70).
As the signature cytokine of Th17 cells, IL-17A amplifies
inflammatory cascades by promoting the production of additional
pro-inflammatory mediators, including TNF-α (71,72).
While experimental studies in pSS models further support a
pathogenic role for Th17 cells during early glandular inflammation
(70,73,74),
the specific molecular pathways that govern their recruitment to
the lacrimal gland remain less defined compared with those in the
salivary glands. In the present study model, NOD/Ltj mice exhibited
increased systemic and local Th17 cell frequency, accompanied by
elevated IL-17A expression in lacrimal glands. NF-κB signaling has
been reported to regulate transcription factors such as retinoic
acid receptor-related orphan receptor-γt and to facilitate IL-17A
production, thereby promoting Th17 cell differentiation and
sustained inflammatory responses in pSS (54). Inhibition of NF-κB with JSH-23
significantly reduced both systemic Th17 cell frequency and local
CD4+IL-17A+ cell infiltration into the lacrimal gland. These
findings supported a mechanistic association between NF-κB
activation and Th17-driven inflammation in pSS lacrimal gland
pathology.
The present study defined the transcriptomic
signatures specific to the lacrimal gland microenvironment. First,
to the best of our knowledge, the present study identified
CARD14 as a newly reported DEG in pSS lacrimal tissues. The
marked downregulation of CARD14 in the disease model of the
present study indicated a distinct function in maintaining
glandular epithelial integrity. Second, by applying high-throughput
mRNA-seq, a localized regulatory network was mapped that associated
CARD14 dysregulation and NF-κB activation with subsequent
chemokine such as CCL19 release and Th17 recruitment. Third, in
vivo experiments also demonstrated that targeted NF-κB
inhibition with JSH-23 not only slows disease progression but
directly restores local glandular secretory function and promotes
epithelial survival.
Several limitations need to be acknowledged in the
present study. First, the sample size utilized for the RNA
sequencing analysis was relatively small which may limit the
overall statistical power to capture all transcriptomic
alterations. Second, the in vivo intervention employed a
single dose of JSH-23 and longitudinal progression of lacrimal
gland pathology was not systematically evaluated. Therefore, the
dose-response relationship and long-term pharmacological safety of
NF-κB inhibition remain to be elucidated in future research. Third,
although the present study findings demonstrated decreased Th17
cell frequency following pathway inhibition, the precise molecular
mechanisms that associate NF-κB activation with Th17
differentiation were not directly investigated. Specifically,
directed gain/loss of function experiments are warranted to confirm
the roles of CARD14 and CCL19. Future longitudinal studies
incorporating optimized sample collection strategies are warranted
to clarify the long-term impact of NF-κB inhibition and
comprehensively evaluate these compositional changes. Furthermore,
integrating these experimental findings with emerging artificial
intelligence tools in medical informatics may hold notable
potential in translating preclinical findings into precision
therapies for patients with pSS (75).
In summary, the present study demonstrated that
CARD14 dysregulation and the subsequent hyperactivation of the
NF-κB signaling pathway contributes to lacrimal gland injury in a
murine model of pSS. NF-κB activation was associated with enhanced
Th17 cell infiltration, increased pro-inflammatory cytokine
production and epithelial apoptosis, resulting in impaired tear
secretion. Pharmacological inhibition of NF-κB with JSH-23
attenuated systemic and local inflammatory responses, reduced
glandular apoptosis and partially restored tear secretion. Although
further studies are warranted to optimize dosing strategies and
define long-term safety, these findings provided in vivo
evidence supporting NF-κB signaling as a potential therapeutic
target in pSS-associated severe dry eye.
Acknowledgements
The authors would like to thank Dr Changjun Wang and
Dr Yiming Sun (Eye Center of Second Affiliated Hospital, Zhejiang
University School of Medicine, Zhejiang, China) for their
assistance with statistical advice.
Funding
The present study was supported by funds from National Natural
Science Foundation of China (grant nos. 82000938 and 82330032) and
Zhejiang Provincial Key Research and Development Program (grant no.
2024C03204).
Availability of data and materials
The data generated in the present study may be found
in the National Center for Biotechnology Information (NCBI) Gene
Expression Omnibus repository under the accession number GSE306385
or at the following URL: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE306385.
The corresponding raw mRNA-seq data generated in the present study
may be found in the NCBI Sequence Read Archive under the BioProject
accession number PRJNA1310557 or at the following URL: https://www.ncbi.nlm.nih.gov/Traces/study/?acc=PRJNA1310557&o=acc_s%3Aa.
Authors' contributions
JX conceptualized the present study, devised the
methodology, conducted the investigation, acquired funding and
wrote the original draft. HZ devised the methodology, conducted the
investigation and formal analysis, and wrote the original draft. WS
devised the methodology, conducted the investigation and formal
analysis. JY conceptualized the present study, provided
supervision, acquired funding, obtained resources, participated in
project administration, reviewed and edited the manuscript. JX and
JY confirm the authenticity of all the raw data. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Animal
Ethics Committee of the Second Affiliated Hospital of Zhejiang
University School of Medicine (approval no. 2022 NO.190; Zhejiang,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
pSS
|
primary Sjögren's syndrome
|
|
NF-κB
|
nuclear factor κB
|
|
Th17
|
T helper 17
|
|
IL-17A
|
interleukin-17A
|
|
IFN-γ
|
interferon-γ
|
|
TNF-α
|
tumor necrosis factor-α
|
|
IKK
|
inhibitor of κB kinase
|
|
IκBα
|
inhibitor of κB α
|
|
PBMCs
|
peripheral blood mononuclear
cells
|
|
DEGs
|
differentially expressed genes
|
|
GO
|
Gene Ontology
|
|
KEGG
|
Kyoto Encyclopedia of Genes and
Genomes
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
ELISA
|
enzyme-linked immunosorbent assay
|
|
TUNEL
|
terminal deoxynucleotidyl transferase
dUTP nick end labeling
|
|
IF
|
immunofluorescence
|
|
IHC
|
immunohistochemistry
|
|
SD
|
standard deviation
|
References
|
1
|
Bjordal O, Norheim KB, Rødahl E, Jonsson R
and Omdal R: Primary Sjögren's syndrome and the eye. Surv
Ophthalmol. 65:119–132. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Xu D, Zhao S, Li Q, Wang YH, Zhao JL, Li
MT, Zhao Y and Zeng XF: Characteristics of Chinese patients with
primary Sjögren's syndrome: Preliminary report of a multi-centre
registration study. Lupus. 29:45–51. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bron AJ, de Paiva CS, Chauhan SK, Bonini
S, Gabison EE, Jain S, Knop E, Markoulli M, Ogawa Y, Perez V, et
al: TFOS DEWS II pathophysiology report. Ocul Surf. 15:438–510.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bodewes ILA, Björk A, Versnel MA and
Wahren-Herlenius M: Innate immunity and interferons in the
pathogenesis of Sjögren's syndrome. Rheumatology (Oxford).
60:2561–2573. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kwok SK, Cho ML, Her YM, Oh HJ, Park MK,
Lee SY, Woo YJ, Ju JH, Park KS, Kim HY and Park SH: TLR2 ligation
induces the production of IL-23/IL-17 via IL-6, STAT3 and NF-kB
pathway in patients with primary Sjogren's syndrome. Arthritis Res
Ther. 14:R642012. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li P, Yang Y, Jin Y, Zhao R, Dong C, Zheng
W, Zhang T, Li J and Gu Z: B7-H3 participates in human salivary
gland epithelial cells apoptosis through NF-κB pathway in primary
Sjögren's syndrome. J Transl Med. 17:2682019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zheng X, Wang Q, Yuan X, Zhou Y, Chu H,
Wang G and Li X, Wang Y, Wei L, Wang L and Li X: B7-H4 inhibits the
development of primary Sjögren's syndrome by regulating treg
differentiation in NOD/Ltj mice. J Immunol Res. 2020:48967272020.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zheng Q, Ren Y, Reinach PS, She Y, Xiao B,
Hua S, Qu J and Chen W: Reactive oxygen species activated NLRP3
inflammasomes prime environment-induced murine dry eye. Exp Eye
Res. 125:1–8. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mariette X and Criswell LA: Primary
Sjögren's syndrome. N Engl J Med. 378:931–939. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Brito-Zerón P, Baldini C, Bootsma H,
Bowman SJ, Jonsson R, Mariette X, Sivils K, Theander E, Tzioufas A
and Ramos-Casals M: Sjögren syndrome. Nat Rev Dis Primers.
2:160472016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ogawa Y, Shimizu E and Tsubota K:
Interferons and dry eye in Sjögren's syndrome. Int J Mol Sci.
19:35482018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liao J, Yu X, Huang Z, He Q, Yang J, Zhang
Y, Chen J, Song W, Luo J and Tao Q: Chemokines and lymphocyte
homing in Sjögren's syndrome. Front Immunol. 15:13453812024.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fu L, Zhao Z, Zhao S, Zhang M, Teng X,
Wang L and Yang T: The involvement of aquaporin 5 in the
inflammatory response of primary Sjogren's syndrome dry eye:
Potential therapeutic targets exploration. Front Med (Lausanne).
11:14398882024. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gao M, Zhao L, Liang R, Zhu Q, Zhao Q and
Kong X: Evaluation of the efficacy and safety of topical 0.05%
cyclosporine eye drops (II) in the treatment of dry eye associated
with primary Sjögren's syndrome. Ocul Immunol Inflamm.
31:1662–1668. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gao Y, Chen Y, Zhang Z, Yu X and Zheng J:
Recent advances in mouse models of Sjögren's syndrome. Front
Immunol. 11:11582020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Park YS, Gauna AE and Cha S: Mouse models
of primary Sjogren's syndrome. Curr Pharm Des. 21:2350–2364. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Humphreys-Beher MG, Hu Y, Nakagawa Y, Wang
PL and Purushotham KR: Utilization of the non-obese diabetic (NOD)
mouse as an animal model for the study of secondary Sjögren's
syndrome. Lacrimal Gland, Tear Film, and Dry Eye Syndromes: Basic
Science and Clinical Relevance. Springer; US, Boston, MA: pp.
631–636. 1994, View Article : Google Scholar
|
|
18
|
Kumar A, Negi G and Sharma SS: JSH-23
targets nuclear factor-kappa B and reverses various deficits in
experimental diabetic neuropathy: Effect on neuroinflammation and
antioxidant defence. Diabetes Obes Metab. 13:750–758. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Nassar A, Kaplanski J and Azab AN: A
selective nuclear factor-κB inhibitor, JSH-23, exhibits
antidepressant-like effects and reduces brain inflammation in rats.
Pharmaceuticals (Basel). 17:12712024. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Percie du Sert N, Hurst V, Ahluwalia A,
Alam S, Avey MT, Baker M, Browne WJ, Clark A, Cuthill IC, Dirnagl
U, et al: The ARRIVE guidelines 2.0: Updated guidelines for
reporting animal research. PLoS Biol. 18:e30004102020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
National Research Council, . Committee for
the Update of the Guide for the Care and Use of Laboratory Animals:
Guide for the care and use of laboratory animals. 8th edition.
National Academies Press US; Washington, DC: 2011
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Singh S, Jasani G, Basu S and Varma DR:
Radiological imaging of the lacrimal gland in Sjogren's syndrome: A
systematic review and meta-analysis. Curr Eye Res. 49:1115–1122.
2024. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shiboski CH, Shiboski SC, Seror R,
Criswell LA, Labetoulle M, Lietman TM, Rasmussen A, Scofield H,
Vitali C, Bowman SJ, et al: 2016 American college of
rheumatology/European league against rheumatism classification
criteria for primary Sjögren's syndrome: A consensus and
data-driven methodology involving three international patient
cohorts. Ann Rheum Dis. 76:9–16. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Daniels TE, Cox D, Shiboski CH, Schiødt M,
Wu A, Lanfranchi H, Umehara H, Zhao Y, Challacombe S, Lam MY, et
al: Associations between salivary gland histopathologic diagnoses
and phenotypic features of Sjögren's syndrome among 1,726 registry
participants. Arthritis Rheum. 63:2021–2030. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gottenberg JE, Cagnard N, Lucchesi C,
Letourneur F, Mistou S, Lazure T, Jacques S, Ba N, Ittah M,
Lepajolec C, et al: Activation of IFN pathways and plasmacytoid
dendritic cell recruitment in target organs of primary Sjögren's
syndrome. Proc Natl Acad Sci USA. 103:2770–2775. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Brkic Z and Versnel MA: Type I IFN
signature in primary Sjögren's syndrome patients. Expert Rev Clin
Immunol. 10:457–467. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen C, Liang Y, Zhang Z, Zhang Z and Yang
Z: Relationships between increased circulating YKL-40, IL-6 and
TNF-α levels and phenotypes and disease activity of primary
Sjögren's syndrome. Int Immunopharmacol. 88:1068782020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang L, Xu P, Wang X, Zhang Z, Zhao W, Li
Z, Yang G and Liu P: Identification of differentially expressed
genes in primary Sjögren's syndrome. J Cell Biochem.
120:17368–17377. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li N, Li L, Wu M, Li Y, Yang J, Wu Y, Xu
H, Luo D, Gao Y, Fei X and Jiang L: Integrated bioinformatics and
validation reveal potential biomarkers associated with progression
of primary Sjögren's syndrome. Front Immunol. 12:6971572021.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lin Y, Yao X, Yan M, Zhou L, Huang W, Xiao
Y, Wu D and Chen J: Integrated analysis of transcriptomics to
identify hub genes in primary Sjögren's syndrome. Oral Dis.
28:1831–1845. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Vitali C, Dolcino M, Del Papa N, Minniti
A, Pignataro F, Maglione W, Lunardi C and Puccetti A: Gene
expression profiles in primary Sjögren's syndrome with and without
systemic manifestations. ACR Open Rheumatol. 1:603–613. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cui J, Li H, Wang T, Shen Q, Yang Y, Yu X
and Hu H: Novel Immune-related genetic expression for primary
Sjögren's syndrome. Front Med (Lausanne). 8:7199582022. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mauduit O, Delcroix V, Umazume T, de Paiva
CS, Dartt DA and Makarenkova HP: Spatial transcriptomics of the
lacrimal gland features macrophage activity and epithelium
metabolism as key alterations during chronic inflammation. Front
Immunol. 13:10111252022. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rattner A, Heng JS, Winer BL, Goff LA and
Nathans J: Normal and Sjogren's syndrome models of the murine
lacrimal gland studied at single-cell resolution. Proc Natl Acad
Sci USA. 120:e23119831202023. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nguyen CQ, Sharma A, She JX, McIndoe RA
and Peck AB: Differential gene expressions in the lacrimal gland
during development and onset of keratoconjunctivitis sicca in
Sjögren's syndrome (SJS)-like disease of the C57BL/6.NOD-Aec1Aec2
mouse. Exp Eye Res. 88:398–409. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Israel L and Mellett M: Clinical and
genetic heterogeneity of CARD14 mutations in psoriatic skin
disease. Front Immunol. 9:22392018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ikawa T, Ichimura Y, Miyagawa T, Fukui Y,
Toyama S, Omatsu J, Awaji K, Norimatsu Y, Watanabe Y, Yoshizaki A,
et al: The contribution of LIGHT (TNFSF14) to the development of
systemic sclerosis by modulating IL-6 and T helper type 1 chemokine
expression in dermal fibroblasts. J Invest Dermatol.
142:1541–1551.e3. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Han M, Sun Y, Zhao W, Xiang G, Wang X,
Jiang Z, Xue Z and Zhou W: Comprehensive characterization of
TNFSF14/LIGHT with implications in prognosis and immunotherapy of
human gliomas. Front Immunol. 13:10252862022. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen R, Wang X, Dai Z, Wang Z, Wu W, Hu Z,
Zhang X, Liu Z, Zhang H and Cheng Q: TNFSF13 is a novel
onco-inflammatory marker and correlates with immune infiltration in
gliomas. Front Immunol. 12:7137572021. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yu Q, Lou XM and He Y: Preferential
recruitment of Th17 cells to cervical cancer via CCR6-CCL20
pathway. PLoS One. 10:e01208552015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hauser MA and Legler DF: Common and biased
signaling pathways of the chemokine receptor CCR7 elicited by its
ligands CCL19 and CCL21 in leukocytes. J Leukoc Biol. 99:869–882.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yang J, Agarwal M, Ling S,
Teitz-Tennenbaum S, Zemans RL, Osterholzer JJ, Sisson TH and Kim
KK: Diverse injury pathways induce alveolar epithelial Cell
CCL2/12, which promotes lung fibrosis. Am J Respir Cell Mol Biol.
62:622–632. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Scudiero I, Zotti T, Ferravante A,
Vessichelli M, Vito P and Stilo R: Alternative splicing of
CARMA2/CARD14 transcripts generates protein variants with
differential effect on NF-κB activation and endoplasmic reticulum
stress-induced cell death. J Cell Physiol. 226:3121–3131. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bertin J, Wang L, Guo Y, Jacobson MD,
Poyet JL, Srinivasula SM, Merriam S, DiStefano PS and Alnemri ES:
CARD11 and CARD14 are novel caspase recruitment domain
(CARD)/membrane-associated guanylate kinase (MAGUK) family members
that interact with BCL10 and activate NF-kappa B. J Biol Chem.
276:11877–11882. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Howes A, O'Sullivan PA, Breyer F, Ghose A,
Cao L, Krappmann D, Bowcock AM and Ley SC: Psoriasis mutations
disrupt CARD14 autoinhibition promoting BCL10-MALT1-dependent NF-κB
activation. Biochem J. 473:1759–1768. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Liu Z, Li F, Pan A, Xue H, Jiang S, Zhu C,
Jin M, Fang J, Zhu X, Brown MA and Wang X: Elevated CCL19/CCR7
expression during the disease process of primary Sjögren's
syndrome. Front Immunol. 10:7952019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yu HR, Sung ML, Kuo HC, Lin CH and Chen
CN: Shear stress modulates resistin-induced CC chemokine ligand 19
expression in human aortic endothelial cells. J Cell Physiol.
230:2120–2127. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Pisani LF, Tontini G, Vecchi M, Croci GA
and Pastorelli L: NF-kB pathway is involved in microscopic colitis
pathogenesis. J Int Med Res. 50:030006052210801042022. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Sisto M, Ribatti D and Lisi S:
Understanding the complexity of Sjögren's syndrome: Remarkable
progress in elucidating NF-κB mechanisms. J Clin Med. 9:28212020.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang X, Shaalan A, Liefers S, Coudenys J,
Elewaut D, Proctor GB, Bootsma H, Kroese FGM and Pringle S:
Dysregulation of NF-kB in glandular epithelial cells results in
Sjögren's-like features. PLoS One. 13:e02002122018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Lisi S, Sisto M, Lofrumento DD and D'Amore
M: Sjögren's syndrome autoantibodies provoke changes in gene
expression profiles of inflammatory cytokines triggering a pathway
involving TACE/NF-κB. Lab Invest. 92:615–624. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Shimizu T, Nakamura H and Kawakami A: Role
of the innate immunity signaling pathway in the pathogenesis of
Sjögren's syndrome. Int J Mol Sci. 22:30902021. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Pringle S, Wang X, Bootsma H, Spijkervet
FKL, Vissink A and Kroese FGM: Small-molecule inhibitors and the
salivary gland epithelium in Sjögren's syndrome. Expert Opin
Investig Drugs. 28:605–616. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Chen W, Lin J, Cao H, Xu D, Xu B, Xu L,
Yue L, Sun C, Wu G and Qian W: Local and systemic IKKε and NF-κB
signaling associated with Sjögren's syndrome immunopathogenesis. J
Immunol Res. 2015:5346482015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Sisto M, Lisi S, Lofrumento DD, Ingravallo
G, De Lucro R and D'Amore M: Salivary gland expression level of
IκBα regulatory protein in Sjögren's syndrome. J Mol Histol.
44:447–454. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Pu J, Wang X, Riaz F, Zhang T, Gao R, Pan
S, Wu Z, Liang Y, Zhuang S and Tang J: Effectiveness and safety of
iguratimod in treating primary Sjögren's Syndrome: A systematic
review and meta-analysis. Front Pharmacol. 12:6212082021.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zeng L, He Q, Yang K, Hao W, Yu G and Chen
H: A systematic review and meta-analysis of 19 randomized
controlled trials of iguratimod combined with other therapies for
Sjögren's syndrome. Front Immunol. 13:9247302022. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Long Z, Zeng L, He Q, Yang K, Xiang W, Ren
X, Deng Y and Chen H: Research progress on the clinical application
and mechanism of iguratimod in the treatment of autoimmune diseases
and rheumatic diseases. Front Immunol. 14:11506612023. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Wang B, Chen S, Li Y, Xuan J, Liu Y and
Shi G: Targeted therapy for primary Sjögren's syndrome: Where are
we now? BioDrugs. 35:593–610. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zhang X, Yin Y, Yue L and Gong L:
Selective serotonin reuptake inhibitors aggravate
depression-associated dry eye via activating the NF-κB pathway.
Invest Ophthalmol Vis Sci. 60:407–419. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Yu Z, Yazdanpanah G, Alam J, de Paiva CS
and Pflugfelder S: Induction of innate inflammatory pathways in the
corneal epithelium in the desiccating stress dry eye model. Invest
Ophthalmol Vis Sci. 64:82023. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Zhou Y, Ma B, Liu Q, Duan H, Huo Y, Zhao
L, Chen J, Han W and Qi H: Transmembrane protein CMTM6 alleviates
ocular inflammatory response and improves corneal epithelial
barrier function in experimental dry eye. Invest Ophthalmol Vis
Sci. 65:42024. View Article : Google Scholar
|
|
64
|
Katsiougiannis S, Tenta R and Skopouli FN:
Autoimmune epithelitis (Sjögren's syndrome); the impact of
metabolic status of glandular epithelial cells on
auto-immunogenicity. J Autoimmun. 104:1023352019. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Moutsopoulos HM: Sjögren's syndrome:
Autoimmune epithelitis. Clin Immunol Immunopathol. 72:162–165.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Manoussakis MN and Kapsogeorgou EK: The
role of intrinsic epithelial activation in the pathogenesis of
Sjögren's syndrome. J Autoimmun. 35:219–224. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Versura P, Bavelloni A, Grillini M,
Fresina M and Campos EC: Diagnostic performance of a tear protein
panel in early dry eye. Mol Vis. 19:1247–1257. 2013.PubMed/NCBI
|
|
68
|
Das N, Menon NG, de Almeida LGN, Woods PS,
Heynen ML, Jay GD, Caffery B, Jones L, Krawetz R, Schmidt TA and
Dufour A: Proteomics analysis of tears and saliva from Sjogren's
syndrome patients. Front Pharmacol. 12:7871932021. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Voigt A, Bohn K, Sukumaran S, Stewart CM,
Bhattacharya I and Nguyen CQ: Unique glandular ex-vivo Th1 and Th17
receptor motifs in Sjögren's syndrome patients using single-cell
analysis. Clin Immunol. 192:58–67. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Verstappen GM, Corneth OBJ, Bootsma H and
Kroese FGM: Th17 cells in primary Sjögren's syndrome: Pathogenicity
and plasticity. J Autoimmun. 87:16–25. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Veldhoen M: Interleukin 17 is a chief
orchestrator of immunity. Nat Immunol. 18:612–621. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Fei Y, Zhang W, Lin D, Wu C, Li M, Zhao Y,
Zeng X and Zhang F: Clinical parameter and Th17 related to
lymphocytes infiltrating degree of labial salivary gland in primary
Sjögren's syndrome. Clin Rheumatol. 33:523–529. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Nguyen CQ, Yin H, Lee BH, Carcamo WC,
Chiorini JA and Peck AB: Pathogenic effect of interleukin-17A in
induction of Sjögren's syndrome-like disease using
adenovirus-mediated gene transfer. Arthritis Res Ther. 12:R2202010.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Katsifis GE, Rekka S, Moutsopoulos NM,
Pillemer S and Wahl SM: Systemic and local interleukin-17 and
linked cytokines associated with Sjögren's syndrome
immunopathogenesis. Am J Pathol. 175:1167–1177. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Lin H: Artificial intelligence with great
potential in medical informatics: A brief review. Medinformatics.
1:2–9. 2024. View Article : Google Scholar
|