Introduction
Hepatocellular carcinoma (HCC) is the third leading
cause of cancer-related deaths worldwide (1). Pathogenesis is predominantly
associated with chronic liver disease, driven by a vicious cycle of
persistent hepatic injury, chronic inflammation and aberrant
hepatocyte regeneration (2,3). The
disease is characterized by rapid progression and poor prognosis,
posing challenges for clinical management. Current treatment
primarily involves surgical resection, often combined with adjuvant
drug therapy (4). However, surgery
entails substantial risks and the recurrence rate within one year
post-surgery is as high as 50% (5), severely limiting long-term outcomes.
Therefore, the development of safer and more effective therapeutic
strategies is an urgent priority in HCC research.
Tumors can evade immune recognition and elimination
primarily by secreting various immunosuppressive factors that
dampen host antitumor immune responses, thereby achieving immune
escape (6,7) (Fig.
1A). Accumulating evidence indicates that, beyond the intrinsic
biological properties of tumor cells, the tumor immune
microenvironment (TIME) serves a pivotal role in regulating the
initiation, progression and recurrence of HCC (8,9).
Notably, the liver and gut are anatomically and functionally
interconnected through the portal venous and biliary systems,
forming the ‘gut-liver axis’. The portal vein transports
gut-derived microbial metabolites to the liver, whereas bile and
certain liver-synthesized immune factors are returned to the
intestine via the biliary tract (10), establishing a bidirectional
communication network. This intimate crosstalk suggests that
dysbiosis or functional alterations in the gut microbiota influence
hepatic immunity and inflammation. Consequently, modulating the gut
microbiota and its metabolic activities represents a novel and
promising therapeutic strategy for HCC intervention (11,12).
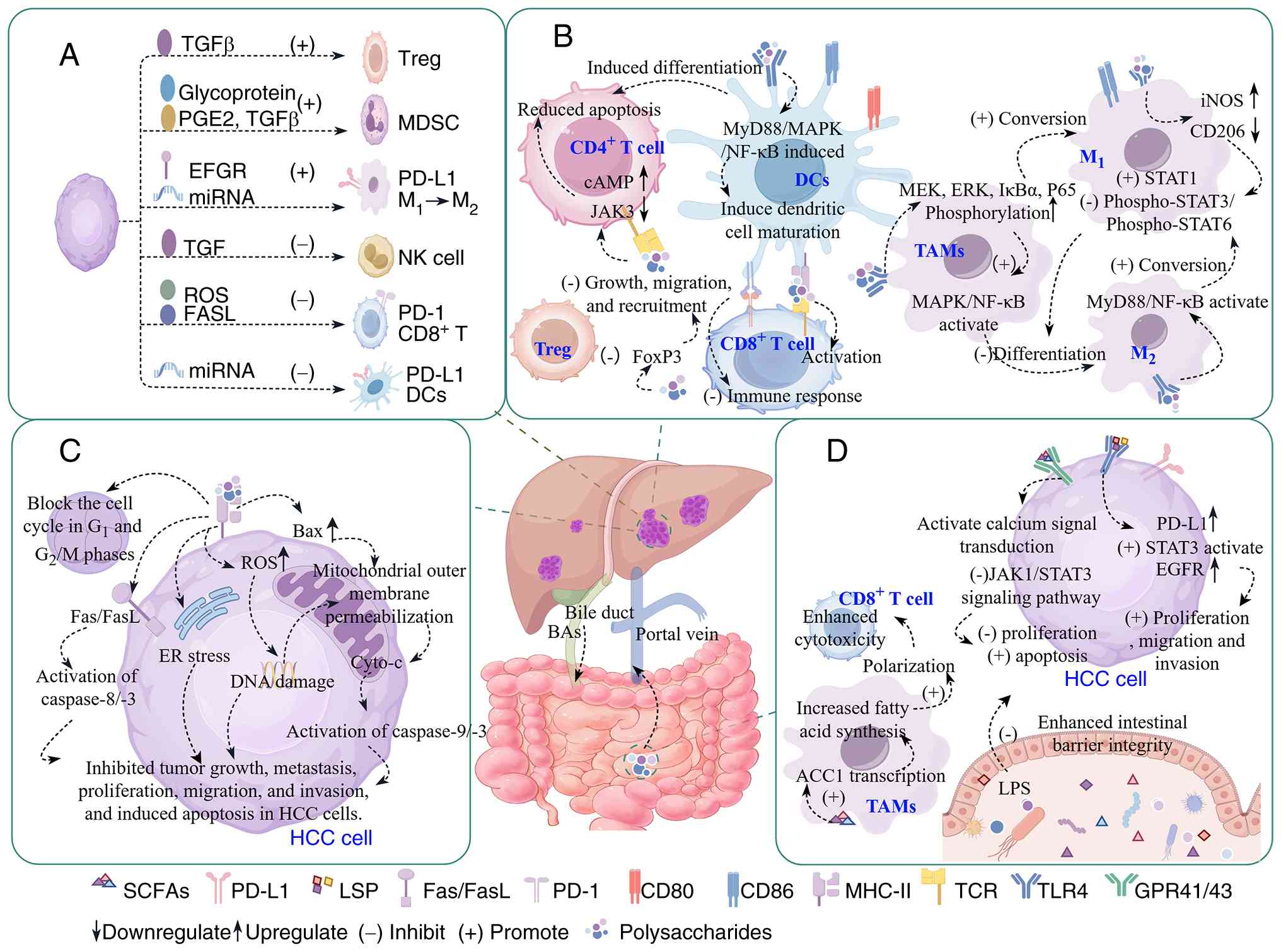 | Figure 1.Overview of the molecular and
cellular mechanisms by which polysaccharides exert anti-HCC
activity. (A) Pro-tumorigenic cellular response in HCC. (B)
Polysaccharide-mediated modulation of tumor cell behavior. (C)
Polysaccharide modulation of the tumor immune microenvironment. (D)
Regulation of gut microbiota and their metabolites by
polysaccharides. Materials provided by FigDraw2.0 (https://www.figdraw.com/). ACC1, acetyl-CoA
carboxylase 1; BAs, bile acids; Cyto-c, cytochrome c; DC,
dendritic cell; ER, endoplasmic reticulum; FASL, Fas and its
ligand; GPR41/43, G protein-coupled receptor 41/43; HCC,
hepatocellular carcinoma; iNOS, inducible nitric oxide synthase;
JAK, Janus kinase; LPS, lipopolysaccharide; MDSC, myeloid-derived
suppressor cells; MHC-II, major histocompatibility complex class
II; miRNA, microRNA; MyD88, myeloid differentiation primary
response 88; NK, natural killer; PD-1, programmed death-1; PD-L1,
programmed death-ligand 1; PGE2, prostaglandin E2; ROS, reactive
oxygen species; SCFA, short-chain fatty acid; TAM, tumor-associated
macrophage; TCR, T cell receptor; TLR4, toll-like receptor 4; Treg,
regulatory T cell. |
Natural polysaccharides have attracted widespread
attention (13–15) due to their antitumor activity.
Following delivery to the liver via the portal vein, these
polysaccharides not only directly target HCC cells by modulating
multiple key signaling pathways, thereby inhibiting proliferation,
migration and invasion, as well as inducing apoptosis (16), but also exert indirect antitumor
effects by remodeling the TIME and enhancing immune surveillance
(17). The fraction of
polysaccharides that remains unabsorbed in the gastrointestinal
tract can interact with the gut microbiota, modulate microbial
composition and metabolic activity, improve intestinal barrier
integrity, and activate specific immune or metabolic pathways, such
as the IL-6/Janus kinase (JAK)1/STAT3 signaling axis, thereby
generating systemic and distal anti-HCC effects (18,19).
The search strategy was as follows: PubMed
(https://pubmed.ncbi.nlm.nih.gov/), Web
of Science (https://www.webofscience.com/), China National
Knowledge Infrastructure (CNKI, http://www.cnki.net/) and Wanfang Data Knowledge
Service Platform (http://www.wanfangdata.com.cn/) were among the major
English and Chinese databases in which a thorough literature search
was carried out. The search covered the period from database
inception to October 2025. The literature search was conducted
using a combination of keywords, primarily focusing on
‘polysaccharides’, ‘HCC’ and ‘clinical research’. More specific
terms, such as ‘gut-liver axis’, ‘clinical trials’ and ‘mechanism’,
were also incorporated to refine the search. Studies involving
treatment were eligible for inclusion if they were conducted on
patients with pathologically or radiologically confirmed HCC, or in
relevant HCC cellular and animal models, and investigated either a
single purified natural polysaccharide or a formulation primarily
composed of natural polysaccharides. The exclusion criteria
included the following: i) Studies that used complex formulas
(rather than single polysaccharide components) or chemically
synthesized drugs as the primary intervention; ii) studies that
focused only on extraction technology without pharmacodynamic
evaluation; and iii) studies involving polysaccharides that did not
assess HCC prevention or treatment.
In summary, the present article provides a
systematic review of current research advances regarding natural
polysaccharides for the treatment of HCC.
Polysaccharide-mediated modulation of tumor
cell behavior
Upon reaching the liver, polysaccharides derived
from plants and fungi can suppress the initiation and progression
of HCC through multi-target, multi-pathway mechanisms. These
compounds exert antitumor effects by inhibiting tumor cell
proliferation, migration and invasion, as well as by inducing
programmed cell death through specific molecular pathways.
The 4°C-extracted Grifola frondosa
polysaccharide (GFG-4) suppresses HepG2 cell proliferation by
engaging the membrane-bound Fas/Fas ligand death receptor system
and activating the downstream caspase-8/-3 pathway (13). In the intrinsic (mitochondrial)
apoptotic pathway, the acid-soluble polysaccharide from Grifola
frondosa upregulates the expression of the pro-apoptotic
protein Bax, thereby inducing the opening of the mitochondrial
permeability transition pore. This facilitates the release of
cytochrome c (cyto-c) from the mitochondria into the
cytosol, resulting in sequential activation of caspase-9 and
caspase-3. The ensuing caspase cascade drives apoptosis in HepG2
cells (14) (Fig. 1C).
In addition to modulating classical apoptotic
pathways, polysaccharides inhibit the progression of HCC through
multiple mechanisms, including cell cycle regulation, suppression
of epithelial-mesenchymal transition (EMT), induction of
endoplasmic reticulum (ER) stress and modulation of redox
homeostasis. Regarding oxidative stress, polysaccharides from
Ulva lactuca and Lactarius deliciosus increase
intracellular reactive oxygen species (ROS) levels, leading to DNA
damage and dissipation of the mitochondrial membrane potential
(20,21). These effects result in the
inhibition of HepG2/H22 cell proliferation and promotion of
apoptosis (20,21). In the context of cell cycle arrest,
Polygonatum sibiricum polysaccharides induce G1
phase arrest in HepG2 cells, thereby reducing the proportion of
cells progressing to the S phase (22). By contrast, daylily polysaccharide
triggers G2/M phase arrest by inhibiting the
Wnt/β-catenin signaling pathway, ultimately suppressing tumor cell
proliferation and promoting apoptosis (23) (Fig.
1C). The in vitro and in vivo anti-HCC effects of
polysaccharides are summarized in Table SI (13,14,21–49).
Numerous polysaccharides have been shown to exhibit
anti-migratory and anti-invasive effects, and thus, to inhibit
tumor metastasis. Codonopsis pilosula polysaccharide
suppresses EMT, reduces cancer stemness and impairs intercellular
adhesion by inhibiting cyclin-dependent kinase 1 activity and
modulating the pyruvate dehydrogenase kinase 1/β-catenin signaling
pathway, thereby markedly attenuating malignant phenotypic traits
in Huh-7 HCC cells (39).
Similarly, basil polysaccharide targets hypoxia-inducible factor-1α
(HIF-1α), alleviates tumor hypoxia and reverses EMT by
downregulating mesenchymal markers (such as β-catenin, N-cadherin
and vimentin) and upregulating epithelial markers [such as
E-cadherin, vacuole membrane protein 1 and zonula occludens-1
(ZO-1)], leading to cytoskeletal reorganization and suppression of
HCC cell invasion and metastasis (33). Furthermore, Astragalus
polysaccharide (APS) enhances ER stress and promotes apoptosis in
HCC cells by reducing protein O-GlcNAcylation and increasing
O-GlcNAcase expression (50).
However, it is imperative to acknowledge that the
antitumor efficacy of polysaccharides is strictly
structure-dependent, and not all polysaccharide fractions exhibit
bioactivity. Variations in molecular weight, glycosidic linkages
and branching degrees can lead to distinct biological outcomes. For
instance, polysaccharide fractions with extremely low molecular
weights may fail to effectively recognize cell surface receptors,
such as toll-like receptors (TLRs) or scavenger receptors,
resulting in a loss of immunomodulatory or pro-apoptotic activity
(51). Similarly, specific
glycosidic bond configurations (for example, α-vs. β-linkages) or
the absence of essential functional groups (such as sulfate or
carboxyl groups) can render a polysaccharide biologically inert
against HCC cells (52). This
structural specificity suggests that the ‘negative’ findings often
reported in polysaccharide research may not indicate a failure of
the therapeutic class per se, but rather reflect the
critical necessity of precise structural features for
bioactivity.
Overall, while the structural heterogeneity of
polysaccharides dictates their specific molecular targets and
efficacy, those possessing optimal structural features consistently
exhibit potent bioactivity against HCC, including suppression of
cell proliferation, induction of apoptosis, induces cell cycle
arrest, inhibition of metastasis and remodeling of the tumor
microenvironment. These pleiotropic effects highlight the
synergistic potential of polysaccharides as multicomponent,
multi-target and multi-pathway therapeutic interventions.
Polysaccharide modulation of the TIME
Polysaccharide interactions with
adaptive immune cells
The T cells within the TIME primarily comprise
CD8+ cells, CD4+ cells and regulatory T cells
(Tregs) (53). CD8+ T
cells exert cytotoxic effects by recognizing tumor-specific
antigens presented by major histocompatibility complex class I
(MHC-I) molecules on antigen-presenting cells (APCs). By contrast,
CD4+ T cells recognize antigenic peptides presented by
MHC class II (MHC-II) molecules, thereby coordinating and
modulating adaptive immune responses to enhance antitumor immunity.
Tregs, which express high levels of the transcription factor
forkhead box protein P3 (FoxP3), mediate immunosuppressive
functions by inhibiting the activation and proliferation of
effector T cells, thereby promoting tumor immune evasion (54,55)
(Fig. 1B). Notably,
polysaccharides can be internalized by APCs via endocytic pathways
and subsequently degraded into low-molecular-weight carbohydrates
via intracellular nitric oxide (NO)-dependent processes. These
processed fragments are then loaded onto MHC-II molecules and
presented to the T-cell receptor, leading to the activation of
antigen-specific CD4+ T-cell responses (56).
CD4+CD25+ Tregs have been
shown to serve a pivotal role in suppressing CD8+ T-cell
activity within the tumor margin regions of HCC (15). APS counteracts this
immunosuppressive effect by downregulating FoxP3 mRNA expression in
the local TIME, thereby inhibiting the proliferation and expansion
of CD4+CD25+ Tregs (Fig. 1B). Additionally, APS disrupts the
C-X-C motif chemokine receptor 4 (CXCR4)/C-X-C motif chemokine
ligand 12 signaling axis, blocking stromal-derived
factor-1-mediated recruitment of Tregs into the HCC
microenvironment, thus reducing their infiltration and
immunosuppressive activity (57).
Similarly, Ganoderma lucidum spore polysaccharide (GLSP)
suppresses HCC growth in tumor-bearing mice by upregulating
microRNA (miR)-125b expression. This leads to inhibition of the
Notch1 signaling pathway and reduced FoxP3 expression, thereby
alleviating Treg-mediated suppression of effector T cell
proliferation (58).
In the context of immune checkpoint regulation,
engagement of programmed death-1 (PD-1) on T cells with its ligand
programmed death-ligand 1 (PD-L1), which is expressed on tumor or
immune cells, results in T cell exhaustion and impaired effector
function (59). In HCC, IFN-γ
secreted by activated immune cells induces acetylation of myocyte
enhancer factor 2D, leading to the upregulation of PD-L1 expression
and subsequent attenuation of CD8+ T cell-mediated
antitumor immunity (60). APS
mitigates this immunosuppressive mechanism by modulating the
miR-133a-3p/moesin signaling axis, which enhances the infiltration
of CD4+ and CD8+ T cells into tumor tissues
(17,61). Furthermore, APS promotes the
generation and long-term persistence of CD122-positive C-X-C motif
chemokine receptor 3-positive PD-1-negative memory-like T cells,
and enhances their migration to tumor sites, boosting the cytotoxic
activity of CD8+ chimeric antigen receptor T cells
against HCC (62).
In addition to modulating Treg cells and immune
checkpoints, other polysaccharides enhance antitumor immunity
through alternative mechanisms. Dandelion polysaccharides increase
the proportion of CD4+ T cells in the spleen and improve
T-cell infiltration into tumor tissues. In H22 tumor-bearing mice,
these immunomodulatory actions mediated by T-cell homeostasis exert
antitumor activity by inhibiting IL-6-activated JAK/STAT signaling
and modulating hepcidin expression (63,64).
Salvianolic acid polysaccharides ameliorate CD4+ T cell
apoptosis and serum cytokine dysregulation induced by H22 tumor
engraftment. Salvianolic acid polysaccharides reduce intracellular
cyclic adenosine monophosphate levels in CD4+ T cells,
downregulate JAK3 protein expression, promote STAT5 phosphorylation
and facilitate transcriptional activation of downstream
anti-apoptotic genes. Collectively, these effects enhance the
cytotoxic function of natural killer (NK) cells and CD8+
T cells (65).
Despite the potential of polysaccharides in
enhancing T-cell infiltration and cytotoxicity, a critical
challenge lies in the potential induction of T-cell exhaustion and
immune tolerance. The tumor microenvironment is characterized by
persistent antigen exposure, and while polysaccharides amplify
T-cell activation, this sustained hyperstimulation may drive
CD8+ T cells into a state of terminal exhaustion, marked
by the upregulation of multiple immune checkpoints [such as PD-1, T
cell immunoglobulin- and mucin-domain-containing 3 (TIM-3) and
lymphocyte-activation gene 3 (LAG-3)] and loss of effector function
(66). Additionally, the liver's
complex immunosuppressive network, characterized by the recruitment
of regulatory T cells and myeloid-derived suppressor cells, may
counteract the initial stimulatory effects of polysaccharides,
leading to a state of adaptive immune resistance (67). This suggests that monotherapy with
immunostimulatory polysaccharides may face limitations due to the
development of compensatory inhibitory mechanisms, highlighting the
necessity of combination strategies or structural optimization to
maximize therapeutic windows.
Polysaccharide interactions with
innate immune cells
Dendritic cells (DCs)
DCs serve as central sentinel cells in immune
surveillance within the TIME and serve a pivotal role in initiating
adaptive immune responses against pathogens and malignant cells,
particularly HCC cells. This is achieved by presenting antigenic
peptides via MHC-I and MHC-II molecules to CD4+ and
CD8+ T cells, respectively (68). Notably, DC-derived exosomes also
express MHC-I, MHC-II, and co-stimulatory molecules such as CD80
and CD86, and possess potent antigen-presenting capacity. These
exosomes can effectively activate both naïve and memory T cells,
making them promising candidates for eliciting specific antitumor
immunity (69,70).
Studies have shown that polysaccharides can bind
specifically to TLR4 on DCs, triggering downstream signaling
cascades that promote DC maturation and functional activation. This
enhances DC-T cell interactions and critically contributes to
anti-HCC immune responses (71).
Notably, polysaccharide components isolated from Portulaca
oleracea and Carica papaya leaves have been demonstrated
to trigger the TLR4/myeloid differentiation primary response 88
(MyD88)/MAPK/NF-κB signaling cascade. In this pathway, the
toll/interleukin-1 receptor/resistance protein (TIR) domain of the
MyD88 adaptor protein engages with the TIR domain of TLR4,
facilitating the recruitment and activation of downstream signaling
intermediates and kinases. This cascade culminates in the nuclear
translocation of the NF-κB transcription factor, which binds to
specific promoter sequences to upregulate the expression of
maturation-associated genes, thereby driving the phenotypic and
functional maturation of DCs (72,73).
Polysaccharides from Alhagi sparsifolia and
APS upregulate the expression of costimulatory molecules (CD80 and
CD86), MHC-I and MHC-II in DCs and their subsets. Concurrently,
they enhance the secretion of key pro-inflammatory cytokines,
including IL-12, TNF-α and IL-6 (74,75).
These changes markedly improve the capacity of DCs for antigen
uptake, processing and presentation, enabling them to efficiently
drive the differentiation of naïve T cells into T helper 1 (Th1)
cells and enhance the cytotoxic activity of CD8+ T
cells, ultimately resulting in suppression of tumor growth
(73). Pretreatment with TAK-242,
a selective TLR4 inhibitor, attenuates polysaccharide-induced
upregulation of DC maturation markers and impairs Th1-polarized
immune responses. This pharmacological blockade confirms the
central role of TLR4 in mediating the immunomodulatory effects of
p-SGP (the purified acidic polysaccharide from Sarcandra
glabra) (76) (Fig. 1B).
Beyond direct T-cell activation, the
immunostimulatory effects of polysaccharides on DCs initiate
broader immune cascade responses. Mature DCs function as central
hubs in the immune network, promoting the activation and
recruitment of NK cells and macrophages, thereby orchestrating a
multi-layered, synergistic antitumor defense within the tumor
microenvironment (77).
Macrophages
Tumor-associated macrophages (TAMs) are among the
most abundant immune cell populations in the TIME and exhibit high
plasticity, enabling their polarization into two functionally
distinct phenotypes: Pro-inflammatory, antitumor M1 macrophages and
anti-inflammatory, pro-tumorigenic M2 macrophages (78). Accumulating evidence indicates that
tumor cells actively suppress M1 polarization and promote the
transition of TAMs toward the M2 phenotype through multiple
mechanisms (79,80). This shift fosters an
immunosuppressive microenvironment that facilitates tumor growth,
invasion and metastasis (81,82).
Polysaccharides have emerged as promising immunomodulatory agents
capable of reversing TAM polarization imbalance. These natural
compounds not only enhance the phagocytic capacity and
antigen-presenting function of macrophages, but also precisely
regulate their polarization at the molecular level, promoting M1
activation while inhibiting M2 differentiation, thereby exerting
potent anti-HCC effects (83–85).
Mechanistic investigations have demonstrated that
polysaccharides exert their effects by activating the
TLR4/MAPK/MyD88/NF-κB signaling cascade. This leads to the
upregulation of pro-inflammatory cytokines (TNF-α, IL-1β and IL-6)
and phosphorylation of critical downstream effectors, including
MEK, ERK, IκBα and p65. Simultaneously, the expression of
M2-associated markers, including IL-10 and arginase-1 (Arg-1), is
downregulated. These molecular events culminate in the phenotypic
reprogramming of M2-polarized TAMs into an M1-like state (85–87).
TLR4 serves as a central receptor mediating the
immunomodulatory effects of polysaccharides on TAMs. It has been
reported that in tumor-bearing mouse models with genetic deletion
of TLR4 or MyD88, APS fails to induce the production of
inflammatory mediators such as NO, TNF-α, IL-1β and IL-6, and the
activation of TNF receptor associated factor 6 and NF-κB is
impaired (83). These findings
underscore the essential role of TLR4-dependent signaling in the
antitumor activity of polysaccharides (Fig. 1B).
In addition to activating the TLR4 signaling
pathway, polysaccharides shift the polarization balance of TAMs by
upregulating M1 markers (including CD86 and inducible nitric oxide
synthase) and pro-inflammatory cytokines, while downregulating M2
markers (such as CD206 and Arg-1) and anti-inflammatory factors.
This phenotypic reprogramming is associated with enhanced
activation of STAT1 and suppression of STAT3 and STAT6
phosphorylation, effectively blocking the transcriptional program
that drives M2 polarization (84,85,88).
In summary, polysaccharides not only enhance the
phagocytic and antigen-presenting functions of TAMs but also
orchestrate their polarization through coordinated modulation of
multiple signaling pathways. By promoting M1 repolarization and
counteracting M2-mediated immunosuppression, they facilitate the
activation of effector immune cells, including T cells and DCs,
ultimately contributing to the formation of a robust antitumor
immune network. These properties highlight the multitarget,
high-efficacy potential of polysaccharides as immunomodulatory
agents in HCC therapy.
However, the potent immunomodulatory capacity of
polysaccharides also presents a double-edged sword, necessitating
careful consideration of the risk of immune hyperactivation and
cytokine storms. Given that TLR4 signaling serves as the primary
mechanism for polysaccharide recognition, excessive or uncontrolled
stimulation of this pathway can lead to the overproduction of
pro-inflammatory mediators, such as TNF-α, IL-1β and IL-6 (89). While these cytokines are essential
for antitumor responses, their systemic elevation may trigger a
‘cytokine storm’, leading to collateral tissue damage and
endotoxin-like toxicity. Furthermore, the dynamic balance between
M1 and M2 macrophage polarization is delicate; while
polysaccharides aim to repolarize TAMs towards the M1 phenotype,
persistent and excessive M1 activation may paradoxically contribute
to chronic inflammation (90) and
immune exhaustion (91) within the
TIME, potentially undermining long-term therapeutic efficacy.
NK cells
NK cells have emerged as promising candidates for
cancer immunotherapy. As key effector cells of the innate immune
system, NK cells serve a pivotal role in antitumor immune
surveillance (92,93). Accumulating evidence indicates that
various polysaccharides can effectively enhance NK cell function
and antitumor activity. For instance, in vivo studies have
demonstrated that Salvia miltiorrhiza polysaccharide,
Panax notoginseng neutral polysaccharide and Angelica
sinensis polysaccharide (ASP) increase NK cell cytotoxicity,
boost splenic lymphocyte activity, elevate the levels of critical
cytokines such as IL-2 and TNF-α, and modulate peripheral blood
lymphocyte subsets, collectively contributing to tumor suppression
and exerting anti-HCC effects (65,94,95).
Despite these findings, the molecular mechanisms by which
polysaccharides specifically regulate NK cells in HCC remain poorly
understood, and systematic investigations in this area are
lacking.
NK cell function is frequently impaired in patients
with HCC, which is a phenomenon linked to both immunosenescence and
an immunosuppressive tumor microenvironment (96,97).
Notably, macrophage migration inhibitory factor interacts with its
receptor CXCR4 to upregulate immunoglobulin-like transcript 2
(ILT2) in NK cells. Elevated ILT2 expression is strongly associated
with diminished NK cell effector functions, including reduced
cytotoxicity and antibody-dependent cellular cytotoxicity. Thus,
ILT2 serves not only as a phenotypic marker of exhausted NK cells
in HCC but also as a promising therapeutic target; blockade of the
ILT2 pathway may restore NK cell antitumor activity (98). Advances in research on engineered
NK (eNK) cells have further highlighted their therapeutic
potential. Functionally enhanced eNK cells exhibit high surface
expression of antitumor molecules such as TNF-related
apoptosis-inducing ligand, CD226 and CD16, along with increased
intracellular levels of perforin, granzyme B, TNF-α and IFN-γ.
These eNK cells exhibit potent cytotoxic activity against multiple
HCC cell lines, including HepG2, HuH7 and SNU-423 cells (99), highlighting the feasibility of
harnessing NK cells for HCC immunotherapy.
Growing evidence highlights the critical role of
microbiota in regulating NK cell function and antitumor immunity
(100). Although the influence of
gut microbiota-derived metabolites on NK cells is increasingly
recognized, the contribution of intratumoral bacteria to NK
cell-mediated responses remains unclear (101). For example, Streptococcus
species within the HCC tumor microenvironment activate the NF-κB
pathway, leading to upregulation of peroxiredoxin 1 (PRDX1)
expression. This induces excessive lactate accumulation, which
suppresses CD8+ T-cell cytotoxicity, and upregulates
immune checkpoint molecules TIM-3 and LAG-3 (102). In vivo experiments have
demonstrated that Streptococcus colonization reduced the
efficacy of PD-1 blockade therapy by 43%, whereas inhibition of
PRDX1 reversed this resistance (102). These findings suggest that PRDX1
is a central node in bacteria-driven metabolic reprogramming and a
key mediator of HCC immune evasion.
Notably, NK cells in the HCC microenvironment are
often functionally impaired or susceptible to cell death (101). By contrast, Bifidobacterium
parabrevis exerts protective effects. B. parabrevis
promotes lipolysis to generate acetyl-CoA, thereby inhibiting NK
cell ferroptosis, and prolonging cell survival and effector
functions (103). Furthermore,
B. parabrevis drives NK cell differentiation toward an
adaptive, highly cytotoxic phenotype characterized by elevated heat
shock protein expression, while suppressing terminal exhaustion,
enhancing anti-HCC activity (103).
In summary, NK cells serve an indispensable role in
the immune response against HCC. As natural immunomodulators,
polysaccharides exhibit the potential to activate NK cells, and
further have the unique capacity to modulate both gut and
intratumoral microbiota (104).
Future research should prioritize the elucidation of synergistic
mechanisms within the multidimensional polysaccharide-microbiota-NK
cell interaction network. Specifically, investigations should focus
on how this axis reshapes the TIME, reverses NK cell exhaustion and
potentiates antitumor immunity. Such efforts will provide novel
insights for the development of precise immunotherapeutic
strategies targeting HCC via NK cell-mediated mechanisms.
Regulation of gut microbiota and their
metabolites by polysaccharides
Characteristics of the gut microbiota
in patients with HCC
The gut microbiota serves a pivotal role in the
initiation, progression, postoperative recovery and recurrence of
HCC. Accumulating evidence indicates that both patients with HCC
and murine models exhibit dysbiosis compared with healthy
individuals (11,105,106). Specifically, there is a marked
reduction in the relative abundance of beneficial bacterial genera
capable of producing short-chain fatty acids (SCFAs), including
Lactobacillus, Bifidobacterium, Bacteroides, Faecalibacterium,
Ruminococcus and Fusobacterium, along with enrichment of
pro-inflammatory lipopolysaccharide (LPS)-producing pathogens
(106). This microbial imbalance
compromises intestinal barrier integrity, increases gut
permeability and promotes a ‘leaky gut’ state, facilitating the
translocation of microbial products such as LPS into the portal
circulation (107). Once in the
liver, these metabolites exacerbate hepatic inflammation and
injury, thereby contributing to HCC development (11,106).
Alterations in specific gut microbial taxa are
closely associated with clinical outcomes in HCC. Depletion of
Lactobacillus, Bacteroides and Bifidobacterium is
associated with an increased risk of early tumor recurrence and
delayed postoperative recovery (108,109). Notably, exogenous probiotic
supplementation has been shown to reshape the gut microbiome,
increase hepatic C-X-C motif chemokine receptor 6-positive NK T
cells and reduce TAM infiltration, effectively suppressing
progression from non-alcoholic steatohepatitis (NASH) to HCC
(110). These findings suggest
that modulation of the gut microbiota may represent a promising
strategy for HCC prevention and intervention.
LPS is a key mediator of the gut-derived influence
on HCC progression and acts through multiple molecular pathways.
First, LPS activates TLR4 in the liver, reinforcing its own
expression via a lin-28 RNA binding posttranscriptional regulator
A/let-7g-mediated positive feedback loop, which subsequently
triggers the NF-κB signaling pathway and drives the release of
pro-inflammatory cytokines, promoting tumor progression (111,112). Second, LPS enhances the stability
and expression of GNAS mRNA through N6-methyladenosine
(m6A) RNA methylation, leading to accumulation of the G
protein α subunit. This suppresses the inhibitory effect of the
long non-coding RNA TPTEP1 on STAT3, resulting in sustained STAT3
activation and accelerated HCC cell proliferation and invasiveness
(113). Furthermore, we found
that the gut microbiota dysbiosis was associated with upregulated
EGFR expression in HCC tissues (Fig.
1D), which may subsequently enhance the proliferation,
migration and invasion of tumor cells (114). LPS also promotes tumor immune
evasion through epigenetic regulation. LPS upregulates the
methyltransferase METTL14, increasing m6A methylation of
lncRNA MIR155HG. This modified transcript is stabilized by the
reader protein ELAV like RNA binding protein 1 and subsequently
modulates the miR-223/STAT1 signaling axis, leading to increased
PD-L1 expression. As a result, T-cell function is suppressed,
facilitating immune escape in HCC (115).
Substantial evidence has demonstrated that
polysaccharides exert protective and therapeutic effects against
HCC by modulating gut microbiota composition and restoring
intestinal barrier function. For example, oral administration of
GLSP or Echinacea purpurea polysaccharides (EPP) enriches
beneficial taxa, such as members of the Muribaculaceae family,
Coprococcus, Clostridium, Roseburia, Bifidobacterium,
Lactobacillus and Bacteroides, and elevates levels of
SCFAs, including acetate, propionate, butyrate and D-lactate, in
HCC-bearing mice relative to the model group (116,117). These metabolites promote
intestinal epithelial cell proliferation, enhance the expression of
tight junction proteins (such as occludin, claudin-1 and ZO-1) and
alleviate oxidative stress, collectively reinforcing the integrity
of the intestinal mucosal barrier (118,119). By reducing systemic LPS
translocation, polysaccharides suppress aberrant activation of the
TLR4/NF-κB signaling pathway, downregulate inflammatory cytokines
(such as IL-6 and TNF-α) and inhibit migration-associated factors
such as MMP-2. Consequently, they attenuate liver injury, inhibit
HCC cell proliferation and induce apoptosis (118,119).
Collectively, these findings indicate that the gut
microbiota and its metabolites act as ‘distal drivers’ in the
pathogenesis of HCC. Polysaccharides exert systemic, multi-target
regulation by reshaping microbial homeostasis, reinforcing the
intestinal barrier and blocking inflammatory signaling cascades.
This mechanism offers a novel perspective for the prevention and
treatment of HCC.
Promotion of SCFA production
In patients with HCC and corresponding animal
models, dysbiosis of the gut microbiota is frequently associated
with reduced levels of SCFAs (11,106,120). Growing evidence indicates that
the anti-HCC effects of polysaccharides are closely associated with
their ability to modulate both the composition of the gut
microbiota and its metabolic output. For example, administration of
EPP, GFG-4 and GLSP alters colonic concentrations of acetate,
propionate, butyrate and D-lactate in HCC-bearing mice compared
with model controls. Concurrently, these treatments upregulate
butyrate metabolism-related pathways in tumor cells (118,119,121). These metabolic changes not only
reflect selective remodeling of gut microbial communities by
polysaccharides but also suggest their potential to indirectly
suppress HCC initiation and progression through enhanced SCFA
production and distribution.
Among SCFAs, acetate exerts multiple antitumor
effects upon reaching the liver. Acetate promotes histone
acetylation in the promoter region of acetyl-CoA carboxylase 1
(ACC1), a key enzyme in fatty acid synthesis, thereby enhancing its
transcriptional activity. This increases de novo
lipogenesis, which modulates TAM polarization and enhances
CD8+ T cell-mediated immune responses, ultimately
suppressing HCC growth (109)
(Fig. 1D). Additionally, acetate
inhibits histone deacetylase activity, leading to increased
acetylation of the transcription factor Sox13 at lysine 30. This
post-translational modification reduces Sox13 protein stability and
suppresses secretion of the pro-inflammatory cytokine IL-17A by
group 3 innate lymphoid cells, thereby alleviating chronic
inflammation in the tumor microenvironment (120). Furthermore, acetate binds to the
G protein-coupled receptor 43 on hepatocytes, inhibiting activation
of the IL-6/JAK1/STAT3 signaling pathway. This suppression blocks
proliferation and induces apoptosis of non-alcoholic fatty liver
disease (NAFLD)-associated HCC cells, effectively impeding
NAFLD-HCC progression (18)
(Fig. 1D).
Butyrate also exhibits potent anti-HCC activity.
Butyrate triggers intracellular calcium dyshomeostasis and robust
ROS accumulation by activating calcium signaling pathways, leading
to mitochondrial dysfunction. This cascade inhibits the
proliferative and metastatic capacities of HCC cells (122). In contrast, Valerate, another
SCFA, delays malignant transformation from NAFLD to HCC by
enhancing intestinal barrier integrity and binding to G
protein-coupled receptor 41/43 on hepatocytes, which in turn
suppresses activation of Rho-GTPase signaling (19).
Collectively, SCFA depletion due to gut microbiota
dysbiosis is a critical pathophysiological event in HCC
development. Polysaccharides can restore metabolic and
immunological homeostasis at the host-microbiota interface by
selectively modulating gut microbial composition and function,
particularly through promotion of SCFA production, thereby exerting
systemic anti-HCC effects. Notably, this regulatory effect is
highly structure-dependent; variations in monosaccharide
composition, glycosidic linkage type and molecular conformation
result in distinct microbial modulation profiles among different
polysaccharides (123). Thus,
deciphering the molecular interactions between polysaccharides and
the gut microbiota clarifies their anti-HCC mechanisms and paves
the way for novel therapeutic strategies based on the ‘gut-liver
axis’.
Regulation of bile acid (BA)
metabolism
BAs are amphipathic metabolites synthesized from
cholesterol in the liver. As important signaling molecules, they
regulate key physiological processes, such as BA homeostasis, lipid
and glucose metabolism, and inflammatory responses, primarily
through the activation of the nuclear receptor farnesoid X receptor
(FXR) and G protein-coupled BA receptor 1 (124). A study has shown that
FXR-deficient mice exhibited increased susceptibility to HCC when
exposed to chemical carcinogens or metabolic liver disease, with a
higher tumor burden than controls (125). These findings highlight the
critical protective role of FXR in suppressing HCC development and
progression.
A complex bidirectional crosstalk exists between the
gut microbiota and BAs, jointly maintaining the dynamic equilibrium
of the ‘gut-liver axis’. During HCC development, gut dysbiosis
disrupts this balance, leading to dysregulated BA metabolism that
promotes hepatic inflammation, fibrosis and malignant
transformation (126,127). For example, secondary BAs such as
deoxycholic acid (DOCA) and lithocholic acid are cytotoxic, and can
induce DNA damage, oxidative stress and mitochondrial dysfunction.
These compounds also modulate the infiltration and function of
immune cells within the TIME, thereby facilitating HCC progression
(128). Alterations in microbial
taxa involved in BA metabolism, such as members of the phylum
Bacteroidetes and the genus Lactobacillus, are associated
with disease progression in NASH-related HCC, changes in serum BA
profiles and severity of liver injury (129). Notably, certain commensal
bacteria, including specific strains of Lactobacillus and
Bifidobacterium, express bile salt hydrolases, enabling them
to deconjugate primary BAs into their free forms. This limits their
conversion into more carcinogenic secondary BAs by other gut
microbes, potentially reducing disease risk (130). Thus, understanding the
interactive network between BAs and the gut microbiota is essential
for elucidating the pathogenesis of HCC.
Polysaccharides have emerged as promising modulators
of gut microbiota composition and BA metabolism due to their
prebiotic-like properties and metabolic regulatory functions
(131). Crude fucoidan derived
from Sargassum, which exhibits a strong BA-binding capacity
and reduces intracellular total cholesterol levels in HepG2 cells
without compromising viability, holds antitumor potential via
modulation of the cholesterol-BA metabolic axis (132). Polysaccharides from Gardenia
jasminoides (Gardenia polysaccharide) and Artemisia
capillaris (Yinchenhao polysaccharide) have exhibited
protective effects in cholestatic liver injury models. Gardenia
polysaccharides upregulate hepatic FXR expression and enrich
butyrate-producing bacteria such as Roseburia and
Faecalibacterium. This enhances FXR activation by
microbially derived SCFAs, reverses BA homeostasis imbalance,
suppresses TLR4/NF-κB signaling, reduces pro-inflammatory cytokine
expression (including TNF-α and IL-6 expression) and alleviates
hepatic inflammation (133,134). The Artemisia capillaris
Thunb. polysaccharide promotes nuclear translocation of nuclear
factor erythroid 2-related factor 2 (Nrf2) and upregulates BA
efflux transporters [for example, bile salt export pump and
multidrug resistance-associated protein 2 (MRP2)] and detoxifying
enzymes (for example, UDP-glucuronosyltransferases and
sulfotransferases), thereby improving BA excretion. Concurrently,
Artemisia capillaris Thunb. polysaccharide increases the
abundance of SCFA-producing bacteria, further potentiating the
Nrf2-mediated antioxidant pathway and mitigating oxidative
stress-induced hepatocyte injury (135).
Polysaccharides have been widely recognized to
modulate immune responses in the tumor microenvironment by
suppressing polarization of M2-type TAMs and promoting their shift
toward a pro-inflammatory (M1-like) phenotype (136). Specifically, studies on Huaier
polysaccharide (HP) have demonstrated that it effectively induces
this phenotypic transition, enhancing the capacity of macrophages
to secrete pro-inflammatory cytokines, thereby strengthening
antitumor immunity (137).
Notably, antibiotic-mediated depletion of the gut microbiota
attenuated the antitumor efficacy of HP, accompanied by reduced
expression of key pro-inflammatory factors such as TNF-α and IL-6.
Further analysis suggested that chenodeoxycholic acid, a secondary
BA produced by gut microbial metabolism, may act as a critical
mediator of the immunomodulatory effects of HP (137).
In summary, the dynamic interplay between the gut
microbiota and BAs is crucial for hepatic homeostasis (126,127). Dysregulation of BA metabolism not
only directly increases HCC risk (125,128) but also establishes a vicious
cycle by reshaping gut microbial composition, thereby exacerbating
liver pathology (126,127,129). Polysaccharides exert
multi-target, multi-pathway regulation of the gut-liver axis,
modulating BA synthesis and signaling while restoring the microbial
balance (131–137), offering unique therapeutic
advantages in HCC prevention and treatment. Future research should
focus on deciphering the molecular mechanisms of the
polysaccharide-microbiota-BA axis to pave the way for precise
interventions and clinical translation.
Development and utilization of
hepatocyte-targeted polysaccharides
Polysaccharide-based targeted
formulations
Polysaccharide-based liver-targeted drug delivery
systems have emerged as promising candidates for cancer therapy
(138). Polysaccharides exhibit
potent immunomodulatory activities, are capable of activating
various immune cells and reshape the TIME. However, their clinical
translation is limited by unfavorable pharmacokinetic properties,
including high molecular weight, low bioavailability and rapid
systemic clearance (139).
Selenium (Se), an essential trace element, serves
critical roles in antioxidant defense, DNA repair and the
regulation of apoptosis, and is closely associated with the
development and progression of multiple cancer types, including HCC
(140). Emerging evidence
suggests that conjugating polysaccharides with Se to form
polysaccharide-Se nanoparticles (PS-SeNPs) not only overcomes the
limitations of the individual components but also generates
synergistic antitumor effects (140). This approach represents a
promising strategy for developing novel, multifunctional anticancer
agents.
Studies have shown that PS-SeNPs induce HCC
apoptosis by co-activating mitochondrial and death receptor
pathways. Specifically, Polyporus umbellatus
polysaccharide-SeNPs and fermented Lactarius deliciosus
polysaccharide-SeNPs were found to upregulate the Bax/Bcl-2 ratio
and activate the cyto-c/caspase-8/9/3 cascade (20,141).
In addition, certain PS-SeNPs inhibit HCC cell
proliferation by modulating cell cycle progression. For example,
Marsdenia tenacissima polysaccharide (MTP70)-SeNPs, composed
of a heteropolysaccharide MTP70 isolated from the stem of
Marsdenia tenacissima (Roxb.) Wight et Arn., simultaneously
activate the Bax/Bcl-2/caspase apoptotic pathway and the
p21/Akt/cyclin A2 cell cycle regulatory axis, inducing S-phase cell
cycle arrest, and effectively suppressing tumor cell proliferation,
invasion and metastasis (142).
Similarly, APS-SeNPs induce morphological changes in HepG2 cells
via the mitochondrial pathway and cause S-phase arrest, exerting
pro-apoptotic effects (143).
In addition to direct cytotoxicity, PS-SeNPs
exhibit immunomodulatory properties and can actively reshape the
TIME. Certain polysaccharide-based nanocomposites selectively
target immune cells, particularly macrophages, promoting their
polarization toward the antitumor M1 phenotype. For instance,
Pholiota adiposa polysaccharide (PAP)-based gold NPs enhance
macrophage secretion of NO, TNF-α and IL-12p70, thereby augmenting
immunomodulatory and antitumor responses (144). By contrast, PAP-SeNPs exhibit
dual targeting capabilities, accumulating in both tumor tissues and
key immune organs. These NPs enter macrophages via
lysosome-dependent phagocytosis and activate the TLR4/MyD88/NF-κB
signaling pathway in a time-dependent manner. This drives
repolarization of immunosuppressive M2 macrophages toward the
pro-inflammatory M1 phenotype, initiating robust antitumor
immunity. Furthermore, PAP-SeNPs increase the proportion of
peripheral CD3+CD4+ and
CD3+CD8+ T cells, further amplifying the
anti-HCC immune response (145).
Additionally, selenium nanoparticles synthesized from Fructus
corni acidic polysaccharide-3 (FCP-3-SeNPs), prepared from an
acidic polysaccharide extracted from Fructus corni
(Cornus officinalis), exhibit strong immunomodulatory
activity. They enhance NO, TNF-α and IL-12p70 production by
macrophages, increase the CD4+/CD8+ T-cell
ratio in peripheral blood, and indirectly induce HCC cell apoptosis
(146). An in vivo study
has demonstrated that FCP-3-SeNPs reduce tumor volume, with
superior efficacy compared with FCP-3 alone, indicating that the Se
nanoformulation markedly enhances the antitumor activity of the
polysaccharides (146).
In summary, polysaccharide-Se nanocomposites combat
HCC via multiple mechanisms, including direct induction of tumor
cell apoptosis, cell cycle arrest and modulation of the TIME,
underscoring their therapeutic potential. Future research should
prioritize optimization of the targeting efficiency and stability
of these nanocarriers, clarify their in vivo
pharmacokinetics and underlying molecular mechanisms, and
accelerate their clinical translation.
Polysaccharides as nanocarriers for
drug delivery
Natural polysaccharides have emerged as promising
materials for the design of nanoscale drug delivery systems due to
their abundant natural sources, excellent biocompatibility and
multivalent functional groups that allow diverse chemical
modifications (147). In the
field of cancer diagnosis and therapy, polysaccharides offer
notable advantages, including high stability, low systemic
toxicity, and the ability to be functionally engineered for
targeted delivery and combination therapies (148). These features enhance the
therapeutic efficacy of anticancer agents while minimizing
off-target side effects.
Certain polysaccharides exhibit intrinsic
liver-targeting properties that make them ideal candidates for
hepatic drug delivery. For instance, polysaccharide fractions
isolated from vinegar-baked Radix Bupleuri (VBRB) exhibit marked
liver-specific accumulation (149). A study has shown that VBRB
polysaccharides enhance both the relative uptake efficiency and the
relative targeting efficiency of drugs in the liver. This improved
hepatic targeting is associated with upregulation of hepatocyte
nuclear factor 4α and organic cation transporter 1, coupled with
downregulation of MRP2, collectively promoting favorable drug
metabolism and transport (149).
Furthermore, a novel VBRB-derived polysaccharide (VRP), VRP3-4, can
self-assemble into stable NPs in aqueous solution and specifically
target breast cancer resistance proteins (BCRP), such as
ATP-binding cassette sub-family G member 2, as well as MRP2. By
suppressing the expression of these efflux transporters, VRP3-4
enhances the inhibitory effect of methotrexate against HCC cells,
exhibiting strong potential as a chemosensitizing agent (150). These findings highlight the role
of VBRB polysaccharides as functional carriers for overcoming drug
resistance in HCC therapy.
ASP, a representative plant-derived polysaccharide,
exhibits excellent water solubility, biocompatibility and inherent
liver-targeting capabilities. By conjugating hydrophobic moieties
such as DOCA or ursodeoxycholic acid (UDCA) to ASP, amphiphilic
conjugates (ASP-DOCA and ASP-UDCA) can be synthesized. These
conjugates spontaneously self-assemble into NPs in aqueous media.
When loaded with therapeutic agents such as oridonin (ORI) or
doxorubicin (DOX), they form drug delivery nanosystems, including
ORI/ASP-DOCA NPs, DOX/ASP-DOCA NPs and ORI/ASP-UDCA NPs. In
vivo studies have demonstrated that these nanotherapeutics are
efficiently internalized by hepatocytes via asialoglycoprotein
receptor (ASGPR)-mediated endocytosis, resulting in enhanced
cellular uptake and cytotoxicity (147,151,152). Compared with free drugs, these
formulations exhibit superior ASGPR-dependent tumor targeting.
Notably, ORI/ASP-DOCA and DOX/ASP-DOCA NPs exhibit markedly
improved antitumor efficacy compared with free ORI and DOX,
respectively (147,151,152).
Another study developed a redox-responsive
amphiphilic polymer, ASP-disulfide-berberine (BBR), by conjugating
ASP with BBR through a disulfide linker and further encapsulating
the mitochondria-targeting agent honokiol (HNK) to construct
ASP-BBR-PM@HNK NPs. This system was efficiently internalized by
HepG2 cells, enhancing hepatic delivery of HNK and its functional
impact on HCC mitochondria (153). Additionally, a biomimetic
nanoplatform, glycyrrhetinic acid-APS-disulfide-curcumin-Cur@RBCm
NPs, has been fabricated by coating erythrocyte membranes onto
glycyrrhetinic acid-APS-disulfide-curcumin micelles. This design
enabled prolonged circulation and enhanced tumor accumulation in
vivo (154). Compared with
hyaluronic acid-based nanocarriers, ASP-based systems exhibit
superior antitumor efficacy and targeting specificity in HCC models
(154), underscoring the unique
advantages of ASP in constructing high-performance liver-targeted
nanotherapeutics.
Scrophularia ningpoensis polysaccharide
(SNP) has been conjugated with UDCA to form SNP-UDCA, which was
used to encapsulate bufalin (BF), yielding BF/SNP-UDCA NPs. In
vitro uptake assays revealed time-dependent internalization of
these NPs in HepG2 cells, which was inhibited by pre-incubation
with free SNP, suggesting that cellular uptake was mediated by
specific recognition of SNP. This finding supports
receptor-mediated targeting (155). For instance, a recent study
demonstrated that polysaccharide-based nanoplatforms not only
enhance drug accumulation in HCC tissues and improve tumor
suppression but also actively modulate the tumor microenvironment.
Specifically, that study showed these systems promote the
repolarization of TAMs from the pro-tumorigenic M2 phenotype to the
antitumor M1 phenotype and effectively recruit NK cells (156).
In conclusion, natural polysaccharide-based
nanocarriers have considerable potential for the precise treatment
of HCC. Their native liver-targeting properties, tunable structures
and potential for multifunctional designs warrant further
investigation.
Current challenges and limitations of
polysaccharide-based nanotherapeutics
Despite the promising advances in
polysaccharide-based nanotherapeutics for HCC, significant
challenges and limitations remain to be addressed before clinical
translation can be realized. A primary concern lies in the
pharmacokinetic instability and potential immunotoxicity of these
formulations. While polysaccharides exhibit intrinsic liver
targeting, their high molecular weight and complex structures often
lead to rapid clearance by the reticuloendothelial system,
resulting in short circulation half-lives (157). Furthermore, although generally
considered biocompatible, the potent immunostimulatory effects of
polysaccharides, mediated primarily through TLR4, pose a risk of
unintended systemic inflammation or immune overactivation when
administered in NP form, necessitating rigorous evaluation of their
immunotoxicological profiles (158).
Another critical limitation is the lack of
standardization and scalability in the preparation of
polysaccharide nanocarriers. Natural polysaccharides are inherently
heterogeneous; their molecular weight, branching degree and
monosaccharide composition can vary depending on the source,
extraction method and season. This batch-to-batch variability poses
a major hurdle for Good Manufacturing Practice compliance and
reproducible drug loading (159).
Additionally, the stability of self-assembled NPs is highly
sensitive to environmental conditions (for example, pH and ionic
strength). A number of polysaccharide-based formulations exhibit
limited stability in the acidic environment of the stomach or the
high-salt environment of the bloodstream, leading to premature drug
leakage and off-target toxicity (160).
While receptor-mediated targeting (for example, via
ASGPR) is a hallmark of these systems, the targeting efficiency
remains suboptimal. The ‘protein corona’ formed upon contact with
blood plasma can mask the targeting ligands on the NP surface,
reducing their specific uptake by hepatocytes (161). Furthermore, the enhanced
permeability and retention effect in human HCC tumors is often less
pronounced than that in murine xenograft models, suggesting that
passive targeting strategies alone may be insufficient for deep
tumor penetration in clinical settings (162). Addressing these limitations
through structural engineering, surface modification and rigorous
preclinical validation will be crucial for the future development
of polysaccharide-based nanomedicines.
Considering the challenges posed by the protein
corona and other biopharmaceutical limitations of monotherapy,
there is a compelling rationale to explore combinatorial
strategies. Combining polysaccharide-based nanotherapeutics with
conventional chemotherapeutic or targeted agents may offer a
synergistic approach to overcome these delivery barriers and
enhance therapeutic efficacy (163).
Polysaccharides in combination therapy
Although targeted therapy and conventional
chemotherapy remain central to the clinical management of HCC,
their long-term efficacy is often limited by declining therapeutic
response, emergence of acquired resistance and significant systemic
toxicity (4,5). These challenges severely compromise
treatment outcomes and patient survival. Polysaccharides have
attracted growing interest due to their excellent biocompatibility,
low toxicity, immunomodulatory properties (57,61)
and potential to enhance the efficacy of conventional anticancer
therapies. As a class of bioactive macromolecules, polysaccharides
are increasingly recognized as promising adjuvants for optimizing
HCC treatment, offering strategies to overcome drug resistance,
reduce adverse effects and improve therapeutic outcomes. In
addition to their use as components of targeted drug delivery
systems or nanocarriers, research has increasingly focused on
combining polysaccharides with established antitumor agents,
including targeted and chemotherapeutic drugs, to achieve
synergistic effects (107).
Polysaccharides can enhance the sensitivity of HCC
cells to targeted agents by modulating key signaling pathways. For
example, fucoidan restores sensitivity to sorafenib in
sorafenib-resistant HepG2-SR cells. Mechanistically, fucoidan
directly interacts with EGFR on the cell membrane, inhibiting its
redistribution into lipid raft microdomains and thereby suppressing
downstream pro-survival signaling. Notably, this synergistic effect
is markedly diminished when EGFR is blocked by a neutralizing
antibody or silenced via gene knockdown (164), confirming that EGFR is a critical
mediator of this combinatorial action. In vivo,
co-administration of fucoidan and sorafenib leads to expanded
regions of tumor apoptosis and necrosis, reduces the expression of
the proliferation marker Ki67 and the angiogenesis marker CD34, and
increases activation of executioner caspase-3 (165). These findings highlight that the
combination not only enhances antitumor activity but also induces
multidimensional pathological improvements.
Polysaccharides have exhibited strong
chemosensitizing and protective effects in combination with
chemotherapy. Cisplatin-induced cell death is impaired in apoptotic
peptidase activating factor 1 (Apaf1)-knockout HepG2 cells,
highlighting the essential role of Apaf1 in mediating
chemosensitivity. When combined with cisplatin, lentinan
upregulates Apaf1 transcription, induces mitochondrial membrane
depolarization and restores the apoptotic capacity. Lentinan
reduces the effective dose of cisplatin required for cytotoxicity
(166) while preserving its
efficacy and minimizing the risk of toxicity and resistance.
Similarly, Ulva lactuca polysaccharides enhance the
antitumor effects of 5-fluorouracil by downregulating serpin family
H member 1, thereby suppressing collagen secretion and disrupting
extracellular matrix deposition. This inhibits tumor cell migration
and invasion, while concurrently alleviating chemotherapy-induced
oxidative stress (167). APS acts
synergistically with DOX by inhibiting O-GlcNAcylation, which
exacerbates ER stress and activates apoptotic pathways, ultimately
promoting DOX-induced apoptosis in HCC cells (50). Additionally, tramete
polysaccharides enhance the sensitivity of HCC cells and xenograft
models to oxaliplatin by modulating the miR-224-5p/ATP binding
cassette subfamily B member 1/P-glycoprotein (P-gp) axis, resulting
in reduced expression of the drug efflux pump P-gp and improved
intracellular drug retention (168).
The synergistic potential of polysaccharides
extends beyond conventional drugs. For instance, asparagus
polysaccharide combined with HIF-1α RNA interference
synergistically suppresses proliferation, migration and invasion in
SK-Hep1 and Hep-3B HCC cell lines. This combination co-inhibits the
PI3K/AKT and MAPK signaling pathways, and effectively reduces tumor
angiogenesis (169),
demonstrating the promise of multimodal combinatorial strategies in
HCC therapy.
In summary, polysaccharides not only exert
synergistic antitumor effects by modulating the tumor
microenvironment, enhancing drug sensitivity and reversing
resistance mechanisms, but also effectively mitigate the toxic side
effects associated with chemotherapy and targeted therapy (164–167). These multifaceted properties make
polysaccharides promising adjuvant interventions for the
comprehensive treatment of HCC, offering novel insights and
experimental foundations for optimizing existing therapeutic
regimens.
Current clinical evidence and gaps
The existing literature (170) and database searches indicate that
clinical research on natural polysaccharides for HCC treatment lags
behind that on modern antineoplastic agents, such as immune
checkpoint inhibitors (ICIs), resulting in a fragile evidence base.
This disparity is primarily attributed to a longstanding structural
imbalance characterized by intense basic research but limited
clinical translation, manifesting as a distinct ‘inverted pyramid’
pattern.
Basic research is robust, as evidenced by extensive
in vitro and in vivo studies on representative
polysaccharides such as lentinan, Ganoderma lucidum and
Astragalus polysaccharides. These studies have provided
profound insights into molecular mechanisms, including
immunomodulation, apoptosis induction, metastasis suppression, and
regulation of the gut-liver axis (38,58,166).
However, high-quality clinical evidence remains
limited. There is a critical lack of high-level evidence-based
medicine studies, specifically phase III, multicenter, randomized,
double-blind controlled trials, dedicated to treating HCC with
single purified polysaccharides. Currently, polysaccharide-based
agents are predominantly used as adjunctive therapies in
combination with transarterial chemoembolization (TACE),
chemotherapy or targeted/immune therapies. Furthermore, existing
applications largely rely on compound formulations or crude
extracts rather than structurally defined, quality-controlled
monomeric polysaccharides (171).
This limitation hinders precise elucidation of their mechanisms of
action and standardized evaluation of their efficacy.
Nevertheless, emerging clinical evidence suggests
the value of polysaccharide-based combination therapies. For
instance, a recent prospective study by Zou et al (170) evaluated the efficacy of
low-molecular-weight fucoidan (LMF), a sulfated polysaccharide
derived from brown algae, in combination with TACE for HCC
treatment. The results demonstrated that, compared with the control
group, the LMF combination group achieved a significantly higher
disease control rate (DCR; 95.24 vs. 80.00%; P=0.035) and a
markedly lower disease progression rate (4.76 vs. 20.00%).
Regarding safety and hepatic function preservation, the LMF group
exhibited superior outcomes, with improved liver function retention
(P=0.029) and a higher proportion of patients with Child-Pugh Class
A status (80.95 vs. 62.50%) (170). These findings demonstrate that
LMF not only enhances tumor control but also effectively mitigates
TACE-associated hepatotoxicity, thereby extending the therapeutic
window for patients while maintaining a favorable safety
profile.
In conclusion, although clinical research on
natural polysaccharides remains insufficient, preliminary evidence
underscores their unique advantages in improving prognosis and
achieving synergistic effects with reduced toxicity (170). There is an urgent need for
large-scale, rigorously designed clinical trials to further
validate their clinical value and facilitate their translation into
standardized therapeutic regimens.
Challenges and limitations
While the specific engineering challenges
associated with polysaccharide-based nanocarriers are discussed
earlier in the manuscript, the broader translation of natural
polysaccharides into clinical oncology faces even more fundamental
bottlenecks. These challenges span across the entire drug
development pipeline, from the elucidation of chemical structures
to regulatory approval. These macro-level limitations spanning
chemistry, biology and translational medicine are categorized and
described subsequently.
Complexity of the material basis and
challenges in structure-activity relationship (SAR)
elucidation
Unlike chemically synthesized small-molecule drugs,
natural polysaccharides are inherently heterogeneous mixtures
characterized by high polydispersity, and microheterogeneity in
molecular weight distribution, degree of branching and
monosaccharide composition (51,52).
Their biological activity depends strongly on fine structural
features ranging from primary to higher-order structures (including
glycosidic bond configuration, branching positions and triple-helix
spatial conformation), rendering establishment of clear SARs
difficult (172). Although
analytical techniques such as nuclear magnetic resonance and mass
spectrometry have advanced considerably, they still face
limitations in resolving precise sequence connectivity and
three-dimensional conformations, hindering identification of key
‘active fragments’ or ‘pharmacophores’ (173). Furthermore, polysaccharide
structures are highly sensitive to extraction, purification and
storage conditions, and even minor structural perturbations (for
example, debranching, degradation or conformational transitions)
can trigger drastic fluctuations in biological activity, further
complicating SAR modeling (174).
While the characterization of primary structures has reached
relative maturity for some polysaccharides, research into
higher-order structures, dynamic SARs and rational structural
modification remains in its infancy.
Biopharmaceutical deficiencies: Dual
barriers of low intestinal permeability and enzymatic
instability
Natural polysaccharides generally possess
physicochemical properties such as high molecular weight, high
viscosity and complex branched architectures, which hinder their
ability to cross the intestinal epithelial barrier into systemic
circulation (139). Following
oral administration, they are prone to enzymatic degradation or
exhibit extremely low bioavailability, severely limiting in
vivo efficacy (175).
Although strategies such as nanocarrier encapsulation, liposomal
wrapping and chemical modifications (for example, sulfation and
acetylation) can improve absorption profiles, these processes often
modify native conformations, potentially reducing intrinsic
activity or introducing unforeseen toxicological risks (140). Balancing improved bioavailability
with preservation of native bioactive structures remains a key
challenge.
Source heterogeneity and lack of
standardized production protocols
The quality of natural polysaccharides is
influenced by multiple factors related to raw material sources (for
example, geographical origin, harvest season, cultivation patterns
and processing methods) and extraction parameters, resulting in
significant batch-to-batch variability (176). Polysaccharides are highly
sensitive to temperature, pH and extraction duration. Traditional
water extraction followed by alcohol precipitation may lead to
structural degradation or residual impurities (including proteins
and nucleic acids). Modern assisted extraction techniques, such as
ultrasound- or microwave-assisted methods, may induce glycosidic
bond cleavage and shifts in molecular weight distribution if not
carefully controlled (177). The
absence of internationally recognized standard operating procedures
for specific polysaccharides hinders cross-study comparability,
reproducibility and industrial translation.
Incomplete safety evaluation systems
and potential risks
Although polysaccharides are generally regarded as
low-toxicity agents, current safety evaluations are largely limited
to acute toxicity and short-term studies. There is a notable lack
of systematic data on long-term chronic toxicity, reproductive
toxicity and carcinogenicity, particularly at high doses or with
prolonged administration (178).
As macromolecules, certain structurally modified polysaccharides or
crude extracts containing protein impurities may exhibit
immunogenicity, potentially triggering allergic reactions or
neutralizing antibody production. Additionally, incomplete removal
of residual solvents, heavy metals, pesticides and endotoxins
during extraction poses significant safety hazards (179). Polysaccharides of animal origin
require rigorous evaluation of viral inactivation and prion
contamination. Furthermore, data on pharmacokinetic drug-drug
interactions between polysaccharides and other antineoplastic
agents remain limited (40).
Obstacles in clinical translation and
regulatory adaptation
A substantial gap exists between laboratory
research and clinical application. Clinical trial design is
challenging due to the lack of well-defined pharmacokinetic
profiles and specific analytes for quantification, complicating
compliance with U.S. Food and Drug Administration/European
Medicines Agency standards (170). Additionally, the unique taste and
viscosity of polysaccharides complicate placebo matching,
increasing the risk of unblinding. Indication positioning remains
ambiguous: Polysaccharides typically exhibit ‘multi-target,
weak-effect’ profiles, making them more suitable for chronic
management or adjuvant therapy, which contrasts with the
‘single-target, strong-effect’ paradigm favored in drug approval
systems (180). Furthermore,
limitations in administration routes persist. Low oral
bioavailability necessitates injectable formulations for some
compounds, increasing production complexity and clinical risks
(181). Finally, global
regulatory frameworks differ markedly. In China, polysaccharides
may be classified as novel traditional Chinese medicine (TCM)
drugs, health food ingredients or food additives, each with
distinct requirements. In Europe and the US, they are often
categorized as dietary supplements, complicating drug approval
unless comprehensive chemistry, manufacturing and control data are
provided (182). Current
pharmacopoeial standards largely rely on total sugar content assays
(such as the phenol-sulfuric acid method), which cannot distinguish
active components from impurities and lack robust fingerprinting
methods (183). Although policies
supporting TCM innovation have been introduced in China, specific
regulatory guidelines governing polysaccharide-based macromolecular
drugs (for example, guidance on methods for evaluating
bioequivalence) remain underdeveloped, limiting industrial
investment and clinical translation.
Limited research perspective: Lack of
systemic immune network analyses
Most studies have focused on changes in isolated
immune cell populations, such as TAMs, DCs, T cells and
myeloid-derived suppressor cells, in terms of quantity, phenotype
or single signaling pathways (57,74,80).
However, they often neglect the dynamic crosstalk among diverse
immune cells within the TIME. Given the multitarget nature of
polysaccharides, future research should shift from ‘single-cell
effects’ to ‘immune network modulation’, aiming to construct
comprehensive maps of systemic immune responses using advanced
tools such as single-cell sequencing and spatial
transcriptomics.
Unclear receptor recognition
mechanisms and insufficient targeting studies
Polysaccharides initiate downstream signaling by
binding to pattern recognition receptors on immune cells, such as
TLRs, C-type lectin receptors and NOD-like receptors (51). However, most studies have focused
on terminal signaling events rather than the molecular details of
polysaccharide-receptor interactions, including binding
specificity, kinetics and conformational dynamics. Elucidating the
identities of key receptors and their functional roles is crucial
for understanding how polysaccharides precisely regulate immune
cell fate and for enabling rational drug design.
Incomplete understanding of gut-liver
axis regulation
The gut microbiota serves a critical role in HCC
pathogenesis. Dysbiosis can promote LPS translocation, impair APC
functions (including the function of DCs and TAMs), and drive
hepatic inflammation and tumorigenesis (107,113). Emerging evidence indicates that
polysaccharides enrich beneficial bacteria (such as
Lactobacillus and Bifidobacterium), suppress
pathogenic species, maintain gut homeostasis and generate
metabolites such as SCFA. These metabolites can activate hepatic
immune cells (including CD8+ T cells and DCs), thereby
reshaping the TIME (118,119,121). Therefore, investigating the
‘polysaccharide-gut microbiota-metabolite-immune cell’ axis
represents a pivotal direction for uncovering the systemic
antitumor mechanisms of these compounds.
Scarcity of high-quality clinical
evidence
Although preclinical data from in vitro and
animal studies are abundant, high-level clinical evidence,
particularly from multicenter, randomized, double-blind,
placebo-controlled trials, is lacking (22–30).
Most existing clinical studies are limited to the Chinese
population, which restricts their generalizability and
international acceptance (170).
International multicenter trials are required to validate the
efficacy and safety across diverse patient cohorts.
Slow pharmacological kinetics: A
mismatch with aggressive tumor phenotypes
Polysaccharides exert their effects primarily
through indirect immunomodulatory mechanisms, resulting in a
relatively slow onset of therapeutic action. Therefore, they are
not ideal as monotherapies for patients with advanced, rapidly
progressing HCC. Instead, they are better suited for early
intervention, postoperative adjuvant therapy and combination
regimens with conventional treatments (164,166,167).
Future directions
The development of natural polysaccharide
therapeutics is currently at a critical juncture, transitioning
from the empirical application of crude extracts to modern
precision medicine characterized by well-defined structures and
controllable quality. To overcome existing bottlenecks and
accelerate modernization and global integration of polysaccharides
for HCC treatment, multidisciplinary convergence is imperative.
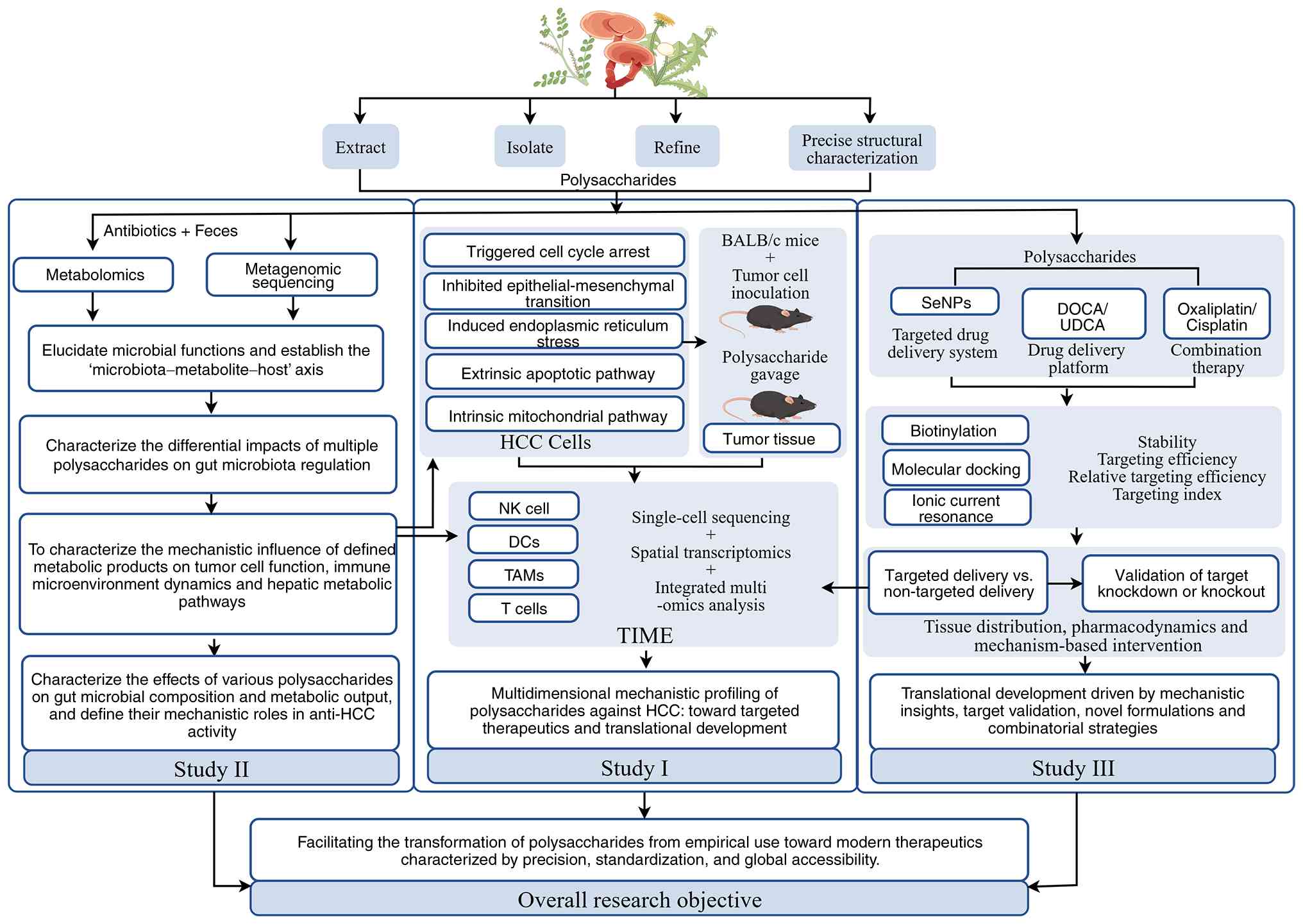
Fig. 2 presents the overall design
framework.
Multi-omics integration for
deciphering systemic regulatory networks
To overcome limitations of traditional
‘single-pathway’ studies, this strategy (Fig. 2, Study I) aims to provide a
comprehensive elucidation of the mechanisms by which
polysaccharides remodel the TIME. This involves collecting HCC
tissue and blood samples before and after polysaccharide treatment
for multidimensional sequencing, including transcriptomics,
proteomics, metabolomics and epigenomics. Advanced bioinformatics
tools [such as MOFA+ (184) and
iCluster (185)] can be used to
integrate these datasets and construct a dynamic interaction
network linking polysaccharides, metabolites, immune factors and
signaling pathways. By directly correlating polysaccharide-induced
metabolic shifts (for example, increased SCFAs) with immune cell
gene expression profiles (such as IFN-γ pathway activation), key
hub nodes can be identified (60,109). This approach avoids the ‘blind
men and an elephant’ trial-and-error paradigm, facilitating a
paradigm shift from isolated mechanistic discovery to a
systems-level understanding.
Microbiomics to decode indirect
gut-liver axis-mediated effects
This direction (Fig.
2, Study II) aims to unravel the ‘black box’ of oral
polysaccharide administration, specifically determining whether
efficacy is mediated through a cascade of ‘oral ingestion-gut
microbiota modulation-hepatic delivery of active metabolites’.
Metagenomic sequencing can provide strain-level resolution and
functional gene profiling (186).
Causality can be validated using germ-free mouse models and fecal
microbiota transplantation. Stable isotope tracing technologies can
further enable real-time tracking of polysaccharide-derived
metabolites (such as butyrate) as they are generated via
microbiota-mediated conversion and transported to the liver to
modulate immune responses (187).
Distinguishing between ‘direct systemic absorption’ and ‘indirect
microbiota-mediated mechanisms’ will inform optimization of
administration routes (oral vs. injection) and formulation
design.
Single-cell and spatial multiomics for
mapping intercellular communication
This approach (Fig.
2, Study I) aims to characterize, at single-cell resolution,
how polysaccharides reshape immune cell states and intercellular
interactions. Combined application of single-cell RNA sequencing
and single-cell T/B cell receptor sequencing enables analysis of
clonal expansion and reversal of exhaustion in tumor-infiltrating
lymphocytes. Coupled with spatial transcriptomics, this allows
in situ mapping of CD8+ T-cell infiltration
trajectories into the tumor core and their spatial proximity to
tumor and stromal cells following polysaccharide treatment
(188). Computational tools such
as CellChat (189) and
CellPhoneDB (190) can be used to
predict and validate enhanced ligand-receptor interactions. This
strategy aims to precisely lock onto the specific ‘effector cell’
subsets targeted by polysaccharides, providing high-resolution
evidence for developing targeted combination therapies.
Artificial intelligence (AI)-enabled
rational drug design and structural
To address structural complexity and unclear SARs,
this strategy optimization (Fig. 2, Study I) aims to transition from
‘empirical extraction’ to ‘precision design’. A structured database
encompassing primary structures (monosaccharide composition and
glycosidic linkages), higher-order conformations (helical
conformation and branching degree) and biological activities
(immunostimulatory potency and receptor affinity) can be developed.
Machine learning models (such as graph neural networks or
transformers) can predict the immunological activity of novel
polysaccharides. AI-driven molecular docking can identify key
structural fragments with high affinity for pattern recognition
receptors (such as TLR4), guiding rational chemical modification
(including precise control of sulfation degrees and molecular
weight tailoring) and synthesis of optimized derivatives (191).
Synergistic combinations with ICIs to
overcome resistance
This strategy leverages the immunomodulatory
properties of polysaccharides to reverse resistance to ICIs. Based
on multiomics and single-cell data, polysaccharides may enhance
tumor immunogenicity (for example, upregulating MHC-I expression or
reducing immunosuppressive signals) (188). The efficacy of combination
therapies (such as polysaccharide + anti-PD-1) should be evaluated
in humanized HCC models, focusing on long-term immune memory.
Early-phase clinical trials (phase Ib/II) using endpoints such as
objective response rate and DCR can rapidly validate these
synergistic effects, accelerating the bench-to-bedside
translation.
Development of advanced nanodelivery
systems and alternative administration routes
This strategy (Fig.
2, Study III) addresses challenges such as low oral
bioavailability and poor targeting. Approaches include: i)
Constructing NPs via amphiphilic modification or self-assembly with
hydrophobic drugs/metal ions (such as Se and Zn) to enhance
stability and absorption (140);
ii) using liposomes or PEGylated micelles to prolong circulation
and improve tumor accumulation via the enhanced permeability and
retention effect (162); and iii)
incorporating targeting ligands (such as galactose for ASGPR
receptor targeting on hepatocytes and RGD peptides for tumor
vasculature) and designing chemical bonds sensitive to the HCC
microenvironment (low pH, high glutathione and specific enzymes
such as hyaluronidase that are overexpressed or exhibit increased
activity) to achieve smart, site-specific release (192). Alternative administration routes,
such as TACE, intraperitoneal hyperthermic perfusion or transdermal
systems, may further enhance therapeutic efficacy by bypassing
first-pass metabolism.
Through integration of these strategies, research
can transition from exploratory approaches to precision-driven
development. By leveraging multiomics technologies for target
identification, AI-assisted approaches for rational drug design and
combination immunotherapy for clinical transition, this integrated
framework provides a promising pathway to accelerate the
development and global adoption of polysaccharide-based therapies
for HCC.
Conclusion
In conclusion, natural polysaccharides possess
unique advantages in the comprehensive management of HCC due to
their multi-target regulation, low toxicity and potent
immunomodulatory properties. They are particularly well suited as
adjuncts to surgery, locoregional therapies, targeted therapy or
immunotherapy, offering benefits in enhancing efficacy, improving
quality of life and preventing recurrence (57,58,164,193). Nevertheless, their clinical
translation faces significant challenges, including mechanistic
ambiguity, lack of standardization, suboptimal formulations and
insufficient high-quality clinical evidence. Future efforts should
focus on deeper mechanistic exploration, precise structural
characterization, formulation innovation and robust clinical
validation to drive the transition of polysaccharides from
empirical applications to precision medicine and global clinical
adoption. By combining traditional insights with modern biomedical
advancements, natural polysaccharides offer a promising pathway for
developing novel HCC immunotherapies and personalized treatment
strategies.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science and Technology
Key Project of the ‘Revealing the List and Taking the Lead’
Initiative in Heilongjiang Province (grant no. 2023ZXJ02C02) and
the Basic Research Foundation for Outstanding Young Teachers in
Heilongjiang Province (grant no. YQJH2023150).
Availability of data and materials
Not applicable.
Authors' contributions
CW was involved in conceptualization, data curation
(the systematic screening, organization, and extraction of relevant
literature retrieved from electronic databases), and prepared the
original draft. JD was involved in supervision and project
administration, and reviewed and edited the manuscript. XZ acquired
funding, was involved in project administration and supervision,
and reviewed and edited the manuscript. Data authentication is not
applicable. All authors have read and approved the final version of
the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
HCC
|
hepatocellular carcinoma
|
|
TIME
|
tumor immune microenvironment
|
|
APS
|
Astragalus polysaccharide
|
|
TACE
|
transarterial chemoembolization
|
|
ICI
|
immune checkpoint inhibitor
|
|
LMF
|
low-molecular-weight fucoidan
|
|
DCR
|
disease control rate
|
|
SCFA
|
short-chain fatty acid
|
|
DC
|
dendritic cell
|
|
TAM
|
tumor-associated macrophage
|
|
NK
|
natural killer
|
|
EMT
|
epithelial-mesenchymal transition
|
|
ER
|
endoplasmic reticulum
|
|
ROS
|
reactive oxygen species
|
|
HIF-1α
|
hypoxia-inducible factor-1α
|
|
PRDX1
|
peroxiredoxin 1
|
|
TLR4
|
toll-like receptor 4
|
|
MyD88
|
myeloid differentiation primary
response 88
|
|
FXR
|
farnesoid X receptor
|
|
BA
|
bile acid
|
References
|
1
|
Rich NE: Changing epidemiology of
hepatocellular carcinoma within the united states and worldwide.
Surg Oncol Clin N Am. 33:1–12. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Han S, Yang Z, Zhang T, Ma J, Chandler K
and Liangpunsakul S: Epidemiology of Alcohol-associated liver
disease. Clin Liver Dis. 25:483–492. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chidambaranathan-Reghupaty S, Fisher PB
and Sarkar D: Hepatocellular carcinoma (HCC): Epidemiology,
etiology and molecular classification. Adv Cancer Res. 149:1–61.
2021.PubMed/NCBI
|
|
4
|
Khan S, Chidi A, Hrebinko K, Kaltenmeier
C, Nassour I, Hoehn R, Geller D, Tsung A and Tohme S: Readmission
after surgical resection and transplantation for hepatocellular
carcinoma: A retrospective cohort study. Am Surg. 88:83–92. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Simsek C, Kim A, Ma M, Danis N, Gurakar M,
Cameron AM, Philosophe B, Garonzik-Wang J, Ottmann S, Gurakar A and
Saberi B: Recurrence of hepatocellular carcinoma following deceased
donor liver transplantation: Case series. Hepatoma Res.
6:112020.PubMed/NCBI
|
|
6
|
Montella L, Sarno F, Ambrosino A, Facchini
S, D'Antò M, Laterza MM, Fasano M, Quarata E, Ranucci RAN, Altucci
L, et al: The role of immunotherapy in a tolerogenic environment:
Current and future perspectives for hepatocellular carcinoma.
Cells. 10:19092021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Volponi C, Gazzillo A and Bonavita E: The
tumor microenvironment of hepatocellular carcinoma: Untying an
intricate immunological network. Cancers (Basel). 14:61512022.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Donne R and Lujambio A: The liver cancer
immune microenvironment: Therapeutic implications for
hepatocellular carcinoma. Hepatology. 77:1773–1796. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li Z, Wang Y, Xing R, Zeng H, Yu XJ, Zhang
YJ, Xu J and Zheng L: Cholesterol efflux drives the generation of
immunosuppressive macrophages to promote the progression of human
hepatocellular carcinoma. Cancer Immunol Res. 11:1400–1413. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Albillos A, de Gottardi A and Rescigno M:
The gut-liver axis in liver disease: Pathophysiological basis for
therapy. J Hepatol. 72:558–577. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ma J, Li J, Jin C, Yang J, Zheng C, Chen
K, Xie Y, Yang Y, Bo Z, Wang J, et al: Association of gut
microbiome and primary liver cancer: A two-sample Mendelian
randomization and case-control study. Liver Int. 43:221–233. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhao R, Li J, Chen B, Zhao J, Hu L, Huang
K, Chen Q, Yao J, Lin G, Bao L, et al: The enrichment of the gut
microbiota Lachnoclostridium is associated with the presence of
intratumoral tertiary lymphoid structures in hepatocellular
carcinoma. Front Immunol. 14:12897532023. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao J, Liang K, Zhong H, Liu S, Sun P and
He R: A cold-water polysaccharide-protein complex from Grifola
frondosa exhibited antiproliferative activity via mitochondrial
apoptotic and Fas/FasL pathways in HepG2 cells. Int J Biol
Macromol. 218:1021–1032. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu J, Liu C, Ji HY and Liu AJ: The
caspases-dependent apoptosis of hepatoma cells induced by an
acid-soluble polysaccharide from Grifola frondosa. Int J
Biol Macromol. 159:364–372. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang XH, Yamagiwa S, Ichida T, Matsuda Y,
Sugahara S, Watanabe H, Sato Y, Abo T, Horwitz DA and Aoyagi Y:
Increase of CD4+ CD25+ regulatory T-cells in the liver of patients
with hepatocellular carcinoma. J Hepatol. 45:254–262. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wen H, Kuang Y, Lian X, Li H, Zhou M, Tan
Y, Zhang X, Pan Y, Zhang J and Xu J: Physicochemical
characterization, antioxidant and anticancer activity evaluation of
an acidic polysaccharide from alpinia officinarum hance.
Molecules. 29:18102024. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
He L, Xu K, Niu L and Lin L: Astragalus
polysaccharide (APS) attenuated PD-L1-mediated immunosuppression
via the miR-133a-3p/MSN axis in HCC. Pharm Biol. 60:1710–1720.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Song Q, Zhang X, Liu W, Wei H, Liang W,
Zhou Y, Ding Y, Ji F, Ho-Kwan Cheung A, Wong N and Yu J:
Bifidobacterium pseudolongum-generated acetate suppresses
non-alcoholic fatty liver disease-associated hepatocellular
carcinoma. J Hepatol. 79:1352–1365. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lau HC, Zhang X, Ji F, Lin Y, Liang W, Li
Q, Chen D, Fong W, Kang X, Liu W, et al: Lactobacillus acidophilus
suppresses non-alcoholic fatty liver disease-associated
hepatocellular carcinoma through producing valeric acid.
EBioMedicine. 100:1049522024. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu Q, Wang X, Hao S, Wu Y, Zhang W, Chen
L, Yan C, Lu Y, Chen Y and Ding Z: Synergetic effects and
inhibition mechanisms of the polysaccharide-selenium nanoparticle
complex in human hepatocarcinoma cell proliferation. J Sci Food
Agric. 104:5124–5138. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Qiu Y, Xu J, Liao W, Wen Y, Jiang S, Wen J
and Zhao C: Suppression of hepatocellular carcinoma by Ulva
lactuca ulvan via gut microbiota and metabolite interactions. J
Adv Res. 52:103–117. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li M, Liu Y, Zhang H, Liu Y, Wang W, You
S, Hu X, Song M, Wu R and Wu J: Anti-cancer potential of
polysaccharide extracted from Polygonatum sibiricum on HepG2
cells via cell cycle arrest and apoptosis. Front Nutr.
9:9382902022. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sang T, Fu YJ and Song L: Polysaccharides
from Hemerocallis citrina baroni inhibit the growth of
hepatocellular carcinoma cells by regulating the Wnt/β-catenin
pathway. Nutr Cancer. 75:1658–1672. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu C, Dai KY, Ji HY, Jia XY and Liu AJ:
Structural characterization of a low molecular weight Bletilla
striata polysaccharide and antitumor activity on H22 tumor-bearing
mice. Int J Biol Macromol. 205:553–562. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kong H, Yang J, Wang X, Mamat N, Xie G,
Zhang J, Zhao H and Li J: The combination of Brassica rapa
L. polysaccharides and cisplatin enhances the anti liver cancer
effect and improves intestinal microbiota and metabolic disorders.
Int J Biol Macromol. 265:1307062024. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huang WH, Liao WR and Sun RX: Astragalus
polysaccharide induces the apoptosis of human hepatocellular
carcinoma cells by decreasing the expression of Notch1. Int J Mol
Med. 38:551–557. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang J, Li Z, Yang X, Qiao Y, Feng C, Yu
S, Jing H, Liu W, Ren L, Duan Q, et al: The antitumor role of a
newly discovered α-d-glucan from Holotrichia diomphalia Bates as a
selective blocker of aldolase A. Carbohydr Polym. 255:1175322021.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Su J, Liao D, Su Y, Liu S, Jiang L, Wu J,
Liu Z and Wu Y: Novel polysaccharide extracted from Sipunculus
nudus inhibits HepG2 tumour growth in vivo by enhancing immune
function and inducing tumour cell apoptosis. J Cell Mol Med.
25:8338–8351. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tan L, Liu S, Li X, He J, He L, Li Y, Yang
C, Li Y, Hua Y and Guo J: The large molecular weight polysaccharide
from wild cordyceps and its antitumor activity on H22 Tumor-bearing
mice. Molecules. 28:33512023. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yue Q, Liu Y, Li F, Hong T, Guo S, Cai M,
Zhao L, Su L, Zhang S, Zhao C and Li K: Antioxidant and anticancer
properties of fucoidan isolated from Saccharina Japonica brown
algae. Sci Rep. 15:89622025. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cho Y, Cho EJ, Lee JH, Yu SJ, Kim YJ, Kim
CY and Yoon JH: Fucoidan-induced ID-1 suppression inhibits the in
vitro and in vivo invasion of hepatocellular carcinoma cells.
Biomed Pharmacother. 83:607–616. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ren F, Li J, Wang Y, Wang Y, Feng S, Yuan
Z and Qian X: The effects of Angelica sinensis
polysaccharide on tumor growth and iron metabolism by regulating
hepcidin in Tumor-bearing mice. Cell Physiol Biochem. 47:1084–1094.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Feng B, Zhu Y, Sun C, Su Z, Tang L, Li C
and Zheng G: Basil polysaccharide inhibits hypoxia-induced
hepatocellular carcinoma metastasis and progression through
suppression of HIF-1α-mediated epithelial-mesenchymal transition.
Int J Biol Macromol. 137:32–44. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Qiu Y, Xu J, Liao W, Yang S, Wen Y, Farag
MA, Zheng L and Zhao C: Ulvan derived from Ulva lactuca
suppresses hepatocellular carcinoma cell proliferation through
miR-542-3p-mediated downregulation of SLC35F6. Int J Biol Macromol.
308:1422522025. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li S, Gao H, Zhao J, Wang G and Gu G:
Galactomannan tetrasaccharide targets mannose receptor to suppress
hepatocellular carcinoma growth through ROS/JNK signaling-induced
apoptosis and ROS-mediated autophagy-dependent cell death. Bioorg
Chem. 165:1090122025. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cheng W, Cheng Z, Weng L, Xing D and Zhang
M: Asparagus polysaccharide inhibits the Hypoxia-induced migration,
invasion and angiogenesis of hepatocellular carcinoma cells partly
through regulating HIF1α/VEGF expression via MAPK and PI3K
signaling pathway. J Cancer. 12:3920–3929. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cheng W, Cheng Z, Xing D and Zhang M:
Asparagus polysaccharide suppresses the migration, invasion, and
angiogenesis of hepatocellular carcinoma cells partly by targeting
the HIF-1α/VEGF signalling pathway in vitro. Evid Based Complement
Alternat Med. 2019:37698792019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
You J, Wu Q, Li Y, Li X, Lin Z, Huang J,
Xue Y, Gulimiran A and Pan Y: Lentinan induces apoptosis of mouse
hepatocellular carcinoma cells through the EGR1/PTEN/AKT signaling
axis. Oncol Rep. 50:1422023. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li N, Yang C, Xia J, Wang W and Xiong W:
Molecular mechanisms of Codonopsis pilosula in inhibiting
hepatocellular carcinoma growth and metastasis. Phytomedicine.
128:1553382024. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang SX, Zhu C, Ba Y, Chen D, Zhou XL,
Cao R, Wang L.P..Ren Y and Wu XZ: Gekko-sulfated glycopeptide
inhibits tumor angiogenesis by targeting basic fibroblast growth
factor. J Biol Chem. 287:13206–13215. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yang Z, Liu Z, Xu J, Zhu J, Pu Y and Bao
Y: Study on the physicochemical properties and immunomodulatory
anti-tumor effect of the Pholiota adiposa polysaccharide.
Food Funct. 13:5153–5165. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang X, Gan J, Han M, Wu Y, Liu L, Zhao Y
and Zhao R: Comparison of structure and the synergistic
anti-hepatocellular carcinoma effect of three polysaccharides from
vinegar-baked Radix Bupleuri. Int J Biol Macromol. 282:1367552024.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yang A, Fan H, Zhao Y, Chen X, Zhu Z, Zha
X, Zhao Y, Chai X, Li J, Tu P and Hu Z: An immune-stimulating
proteoglycan from the medicinal mushroom Huaier up-regulates NF-κB
and MAPK signaling via Toll-like receptor 4. J Biol Chem.
294:2628–2641. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang H, Sun B, Zhang Z, Chen J, Hao Q, Sun
Y, Yang Y, Wang Z and Pei J: Effects of Acanthopanax senticosus
polysaccharide on the proliferation, apoptosis and cell cycle in
human HepG2 cells. Pharmazie. 71:201–204. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chen P, Liu HP, Ji HH, Sun NX and Feng YY:
A cold-water soluble polysaccharide isolated from Grifola
frondosa induces the apoptosis of HepG2 cells through
mitochondrial passway. Int J Biol Macromol. 125:1232–1241. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hu T, Zhang K, Pan D, Pan X, Yang H, Xiao
J, Shen X and Luo P: Inhibition effect of dictyophora
polysaccharides on human hepatocellular carcinoma cell line
HCC-LM3. Med Sci Monit. 26:e9188702020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ying Y, Yuan C, Jiao Z, Zheng L, Qun W,
Hao G, Xiaofei S, Mingdian Y and Nan S: Pachymaran regulates
pyroptosis of liver cancer cells via SQLE/NLRP3/GSDMD signaling
pathway. Chin J Pathophysiol. 40:444–455. 2024.
|
|
48
|
Weiping L, Jianfen C, Jingi A, Yi L and
Fengbin L: Mechanism of sulfated polysaccharide from Undaria
pinnatifidaon proliferation and migration of hepatocellular
carcinoma cells. Chin Pharmacol Bull. 40:670–678. 2024.
|
|
49
|
Cai L, Zhou S, Wang Y, Xu X, Zhang L and
Cai Z: New insights into the anti-hepatoma mechanism of
triple-helix β-glucan by metabolomics profiling. Carbohydr Polym.
269:1182892021. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Li M, Duan F, Pan Z, Liu X, Lu W, Liang C,
Fang Z, Peng P and Jia D: Astragalus polysaccharide promotes
Doxorubicin-induced apoptosis by reducing O-GlcNAcylation in
hepatocellular carcinoma. Cells. 12:8662023. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Nam J, Kim A, Kim K, Moon J.H..Baig J,
Phoo M, Moon JJ and Son S: Engineered polysaccharides for
controlling innate and adaptive immune responses. Nat Rev Bioeng.
2:733–751. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Adhikari E, Liu Q, Burton C,
Mockabee-Macias A, Lester DK and Lau E: L-fucose, a sugary
regulator of antitumor immunity and immunotherapies. Mol Carcinog.
61:439–453. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Garris CS and Luke JJ: Dendritic cells,
the T-cell-inflamed tumor microenvironment, and immunotherapy
treatment response. Clin Cancer Res. 26:3901–3907. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Liang R, Li L and Wang Z: Progress in
clinical research on tumor-infiltrating lymphocytes insolid tumors.
Chin J Clin Oncol. 48:1168–1172. 2021.
|
|
55
|
Zheng X, Jin W, Wang S and Ding H:
Progression on the roles and mechanisms of Tumor-Infiltrating T
lymphocytes in patients with hepatocellular carcinoma. Front
Immunol. 12:7297052021. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Cobb BA, Wang Q, Tzianabos AO and Kasper
DL: Polysaccharide processing and presentation by the MHCII
pathway. Cell. 117:677–687. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Li Q, Bao JM, Li XL, Zhang T and Shen XH:
Inhibiting effect of Astragalus polysaccharides on the functions of
CD4+CD25 highTreg cells in the tumor microenvironment of human
hepatocellular carcinoma. Chin Med J (Engl). 125:786–793.
2012.PubMed/NCBI
|
|
58
|
Li A, Shuai X, Jia Z, Li H, Liang X, Su D
and Guo W: Ganoderma lucidum polysaccharide extract inhibits
hepatocellular carcinoma growth by downregulating regulatory T
cells accumulation and function by inducing microRNA-125b. J Transl
Med. 13:1002015. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Sangro B, Melero I, Wadhawan S, Finn RS,
Abou-Alfa GK, Cheng AL, Yau T, Furuse J, Park JW, Boyd Z, et al:
Association of inflammatory biomarkers with clinical outcomes in
nivolumab-treated patients with advanced hepatocellular carcinoma.
J Hepatol. 73:1460–1469. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Xiang J, Zhang N, Sun H, Su L, Zhang C, Xu
H, Feng J, Wang M, Chen J, Liu L, et al: Disruption of SIRT7
increases the efficacy of checkpoint inhibitor via MEF2D regulation
of programmed cell death 1 ligand 1 in hepatocellular carcinoma
cells. Gastroenterology. 158:664–678.e24. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zechen L, Heran Z, Yazhen Z, Jinhua L and
Jue W: Effects of astragalus polysaccharide on inhibitory ability
of PD-1 inhibitor in H22 Tumor-bearing mice. Chin J Tradit Med Sci
Technol. 31:217–222. 2024.
|
|
62
|
Zhang Q, Su C, Luo Y, Zheng F, Liang CL,
Chen Y, Liu H, Qiu F, Liu Y, Feng W, et al: Astragalus
polysaccharide enhances antitumoral effects of chimeric antigen
receptor-engineered (CAR) T cells by increasing
CD122+CXCR3+PD-1-memory T cells. Biomed Pharmacother.
179:1174012024. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Ren F, Zhang Y, Qin Y, Shang J, Wang Y,
Wei P, Guo J, Jia H and Zhao T: Taraxasterol prompted the
anti-tumor effect in mice burden hepatocellular carcinoma by
regulating T lymphocytes. Cell Death Discov. 8:2642022. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Ren F, Yang Y, Wu K, Zhao T, Shi Y, Song M
and Li J: The effects of dandelion polysaccharides on iron
metabolism by regulating hepcidin via JAK/STAT signaling pathway.
Oxid Med Cell Longev. 2021:71847602021. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Shu G, Zhao W, Yue L, Su H and Xiang M:
Antitumor immunostimulatory activity of polysaccharides from Salvia
chinensis Benth. J Ethnopharmacol. 168:237–247. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Vatsavai N, Kaur Bhinder S, Shaik R,
Mahira S, Kapoor S, Ali MS, Verma D, Singh J, Badavenkatappa Gari
S, Upadhyay P, et al: Advances and challenges in cancer
immunotherapy: Mechanisms, clinical applications, and future
directions. Front Pharmacol. 16:16025292025. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Li X, Ruan Q, Yang W, Tian H, Wu N, Qadir
J, Wang J, Hu H, Liu Y, Cai M, et al: Polysaccharide isolated from
Grifola frondosa eliminates myeloid-derived suppressor cells
and inhibits tumor growth by enhancing T cells responses. Int J
Biol Sci. 20:664–679. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Wang Y, Xiang Y, Xin VW, Wang XW, Peng XC,
Liu XQ, Wang D, Li N, Cheng JT, Lyv YN, et al: Dendritic cell
biology and its role in tumor immunotherapy. J Hematol Oncol.
13:1072020. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Wang X, He L, Huang X, Zhang S, Cao W, Che
F, Zhu Y and Dai J: Recent progress of exosomes in multiple
myeloma: Pathogenesis, diagnosis, prognosis and therapeutic
strategies. Cancers (Basel). 13:16352021. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Tarte K and Klein B: Dendritic cell-based
vaccine: A promising approach for cancer immunotherapy. Leukemia.
13:653–663. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Zhang W, Park HB, An EK, Kim SJ, Ryu D,
Kim D, Lim D, Hwang J, Kwak M, You S, et al: Fucoidan from
Durvillaea Antarctica enhances the anti-cancer effect of
anti-PD-L1 antibody by activating dendritic cells and T cells. Int
J Biol Macromol. 280:1359222024. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Jia G, Shao X, Zhao R, Zhang T, Zhou X,
Yang Y, Li T, Chen Z and Liu Y: Portulaca oleracea L.
polysaccharides enhance the immune efficacy of dendritic cell
vaccine for breast cancer. Food Funct. 12:4046–4059. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Kim WS, Han JM, Song HY, Byun EH, Lim ST
and Byun EB: Annona muricata L.-Derived polysaccharides as a
potential adjuvant to a dendritic Cell-based vaccine in a
Thymoma-bearing model. Nutrients. 12:16022020. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Lim SM, Park HB and Jin JO: Polysaccharide
from Astragalus membranaceus promotes the activation of human
peripheral blood and mouse spleen dendritic cells. Chin J Nat Med.
19:56–62. 2021.PubMed/NCBI
|
|
75
|
Wusiman A, He J, Cai G, Zhu T, Bo R, Liu
Z, Hu Y and Wang D: Alhagi honey polysaccharides encapsulated into
PLGA nanoparticle-based pickering emulsion as a novel adjuvant to
induce strong and long-lasting immune responses. Int J Biol
Macromol. 202:130–140. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Liu W, Gong X, Luo J, Jiang L, Lu W, Pan
C, Yao W, Gao X and Tian H: A purified acidic polysaccharide from
Sarcandra glabra as vaccine adjuvant to enhance anti-tumor
effect of cancer vaccine. Carbohydr Polym. 263:1179672021.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Zhang W, An EK, Park HB, Hwang J,
Dhananjay Y, Kim SJ, Eom HY, Oda T, Kwak M, Lee PC and Jin JO:
Ecklonia cava fucoidan has potential to stimulate natural killer
cells in vivo. Int J Biol Macromol. 185:111–121. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Jiang Y, Han Q, Zhao H and Zhang J:
Promotion of epithelial-mesenchymal transformation by
hepatocellular carcinoma-educated macrophages through
Wnt2b/β-catenin/c-Myc signaling and reprogramming glycolysis. J Exp
Clin Cancer Res. 40:132021. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Wu H, Li J, Yao R, Liu J, Su L and You W:
Focusing on the interplay between tumor-associated macrophages and
tumor microenvironment: From mechanism to intervention.
Theranostics. 15:7378–7408. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Chen D, Zhang X, Li Z and Zhu B: Metabolic
regulatory crosstalk between tumor microenvironment and
tumor-associated macrophages. Theranostics. 11:1016–1030. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Hu Y, Li Y, Xiong H, Zhang Y, Wang F, Zhuo
W, Zeng Z, Zhao Y, Wang H, Hu P, et al: Exosomal
SLC16A1-AS1-induced M2 macrophages polarization facilitates
hepatocellular carcinoma progression. Int J Biol Sci. 20:4341–4363.
2024. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Jiang M, Wang D, Su N, Lou W, Chen Y, Yang
H, Chen C, Xi F, Chen Y, Deng L, et al: TRIM65 knockout inhibits
the development of HCC by polarization tumor-associated macrophages
towards M1 phenotype via JAK1/STAT1 signaling pathway. Int
Immunopharmacol. 128:1114942024. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Zhou L, Liu Z, Wang Z, Yu S, Long T, Zhou
X and Bao Y: Astragalus polysaccharides exerts immunomodulatory
effects via TLR4-mediated MyD88-dependent signaling pathway in
vitro and in vivo. Sci Rep. 7:448222017. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Song M, Li ZH, Gu HS, Tang RY, Zhang R,
Zhu YL, Liu JL, Zhang JJ and Wang LY: Ganoderma lucidum
spore polysaccharide inhibits the growth of hepatocellular
carcinoma cells by altering macrophage polarity and induction of
apoptosis. J Immunol Res. 2021:66966062021. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Li GL, Tang JF, Tan WL, Zhang T, Zeng D,
Zhao S, Ran JH, Li J, Wang YP and Chen DL: The anti-hepatocellular
carcinoma effects of polysaccharides from Ganoderma lucidum
by regulating macrophage polarization via the MAPK/NF-κB signaling
pathway. Food Funct. 14:3155–3168. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Su H, He L, Yu X, Wang Y, Yang L, Wang X,
Yao X, Luo P and Zhang Z: Structural characterization and
mechanisms of macrophage immunomodulatory activity of a novel
polysaccharide with a galactose backbone from the processed
Polygonati Rhizoma. J Pharm Anal. 14:1009742024. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Wang YJ, Wan DL, Li QM, Zha XQ and Luo JP:
Structural characteristics and immunostimulatory activities of a
new polysaccharide from Dendrobium fimbriatum Hook. Food
Funct. 12:3057–3068. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Guo W, Liu X, Guo J, Gao R, Xiang X, An X
and Bai L: Polysaccharides of Brassica rapa L. attenuate
tumor growth via shifting macrophages to M1-like phenotype.
Phytother Res. 36:3957–3968. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Zhang Y, Cui Y, Feng Y, Jiao F and Jia L:
Lentinus edodes polysaccharides alleviate acute lung injury by
inhibiting oxidative stress and inflammation. Molecules.
27:73282022. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Delprat V, Tellier C, Demazy C, Raes M,
Feron O and Michiels C: Cycling hypoxia promotes a pro-inflammatory
phenotype in macrophages via JNK/p65 signaling pathway. Sci Rep.
10:8822020. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Huang R, Kang T and Chen S: The role of
tumor-associated macrophages in tumor immune evasion. J Cancer Res
Clin Oncol. 150:2382024. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Barrow AD and Colonna M: Tailoring Natural
Killer cell immunotherapy to the tumour microenvironment. Semin
Immunol. 31:30–36. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Ghaedrahmati F, Esmaeil N and Abbaspour M:
Targeting immune checkpoints: How to use natural killer cells for
fighting against solid tumors. Cancer Commun (Lond). 43:177–213.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Liu YH, Qin HY, Zhong YY, Li S, Wang HJ,
Wang H, Chen LL, Tang X, Li YL, Qian ZY, et al: Neutral
polysaccharide from Panax notoginseng enhanced cyclophosphamide
antitumor efficacy in hepatoma H22-bearing mice. BMC Cancer.
21:372021. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Dong XD, Liu YN, Zhao Y, Liu AJ, Ji HY and
Yu J: Structural characterization of a water-soluble polysaccharide
from Angelica dahurica and its antitumor activity in H22
tumor-bearing mice. Int J Biol Macromol. 193:219–227. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Li Z, Gong Y, Ye L and Liu W: The dual
roles of natural killer cells in liver immunity and tolerance:
Implications for health and disease. Hepatol Commun. 10:e009232026.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Lu X, Luo Y, Huang Y, Zhu Z, Yin H and Xu
S: Cellular senescence in hepatocellular carcinoma: Immune
microenvironment insights via machine learning and in vitro
experiments. Int J Mol Sci. 26:7732025. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Sakata T, Yoshio S, Yamazoe T, Mori T,
Kakazu E, Aoki Y, Aoyanagi N, Okamoto T, Ito T, Toyoda H, et al:
Immunoglobulin-like transcript 2 as an impaired anti-tumor
cytotoxicity marker of natural killer cells in patients with
hepatocellular carcinoma. Front Immunol. 15:13894112024. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Nakamura M, Tanaka Y, Hakoda K, Ohira M,
Kobayashi T, Kurachi K, Tamura K and Ohdan H: Antitumor effects of
natural killer cells derived from gene-engineered human-induced
pluripotent stem cells on hepatocellular carcinoma. Cancer Immunol
Immunother. 74:992025. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Tian P, Yang W, Guo X, Wang T, Tan S, Sun
R, Xiao R, Wang Y, Jiao D, Xu Y, et al: Early life gut microbiota
sustains liver-resident natural killer cells maturation via the
butyrate-IL-18 axis. Nat Commun. 14:17102023. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Fanijavadi S, Hansen TF and Zedan AH: NK
Cell-microbiota interaction biomarker strategy: Advancing prostate
cancer management. Biomolecules. 15:2732025. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Zhang H, Lan X, Cai L, Gao X, Gao F, Yu D,
Zhang J, Zhang J and Tai Q: Tumor-associated bacteria activate
PRDX1-driven glycolysis to promote immune evasion and PD-1 antibody
resistance in hepatocellular carcinoma. Front Microbiol.
16:15996912025. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Pan B, Zhang X, Ye D, Yao Y, Zhang Z, Luo
Y, Wu H, Wang X and Tang N: Intratumoral Brevibacillus parabrevis
enhances antitumor immunity by inhibiting NK cell ferroptosis in
hepatocellular carcinoma. Cell Death Dis. 16:4072025. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Álvarez-Mercado AI and Plaza-Diaz J:
Dietary polysaccharides and gut microbiota ecosystem. Nutrients.
14:42852022. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Wang XY, Zhang Y and Liu FF: Influence of
Pholiota adiposa on gut microbiota and promote tumor cell
apoptosis properties in H22 tumor-bearing mice. Sci Rep.
12:85892022. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Huo R and Chen Y, Li J, Xu Q, Guo J, Xu H,
You Y, Zheng C and Chen Y: Altered gut microbiota composition and
its potential association in patients with advanced hepatocellular
carcinoma. Curr Oncol. 30:1818–1830. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Dong J, Ping L, Cao T, Sun L, Liu D, Wang
S, Huo G and Li B: Immunomodulatory effects of the
Bifidobacterium longum BL-10 on lipopolysaccharide-induced
intestinal mucosal immune injury. Front Immunol. 13:9477552022.
View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Zheng C, Lu F, Chen B, Yang J, Yu H, Wang
D, Xie H, Chen K, Xie Y, Li J, et al: Gut microbiome as a biomarker
for predicting early recurrence of HBV-related hepatocellular
carcinoma. Cancer Sci. 114:4717–4731. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Ma H, Yang L, Liang Y, Liu F, Hu J, Zhang
R, Li Y, Yuan L and Feng F: B. thetaiotaomicron-derived acetic acid
modulate immune microenvironment and tumor growth in hepatocellular
carcinoma. Gut Microbes. 16:22978462024. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Li T, Lin X, Shen B, Zhang W, Liu Y, Liu
H, Wang Y, Zheng L and Zhi F: Akkermansia muciniphila suppressing
nonalcoholic steatohepatitis associated tumorigenesis through
CXCR6(+) natural killer T cells. Front Immunol. 13:10475702022.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Beyoğlu D and Idle JR: The gut
microbiota-A vehicle for the prevention and treatment of
hepatocellular carcinoma. Biochem Pharmacol. 204:1152252022.
View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Chen IT, Cheng AC, Liu YT, Yan C, Cheng
YC, Chang CF and Tseng PH: Persistent TLR4 activation promotes
hepatocellular carcinoma growth through positive feedback
regulation by LIN28A/Let-7g miRNA. Int J Mol Sci. 23:84192022.
View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Ding H, Zhang X, Su Y, Jia C and Dai C:
GNAS promotes inflammation-related hepatocellular carcinoma
progression by promoting STAT3 activation. Cell Mol Biol Lett.
25:82020. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Kubo T, Nishimura N, Kaji K, Tomooka F,
Shibamoto A, Iwai S, Suzuki J, Kawaratani H, Namisaki T, Akahane T,
et al: Role of epiregulin on lipopolysaccharide-induced
hepatocarcinogenesis as a mediator via EGFR signaling in the cancer
microenvironment. Int J Mol Sci. 25:44052024. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Peng L, Pan B, Zhang X, Wang Z, Qiu J,
Wang X and Tang N: Lipopolysaccharide facilitates immune escape of
hepatocellular carcinoma cells via m6A modification of lncRNA
MIR155HG to upregulate PD-L1 expression. Cell Biol Toxicol.
38:1159–1173. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Zhang XT, Yang Y, Ji C, Fu Y, Pu X and Xu
G: Ganoderma lucidum polysaccharides reduce the severity of
acute liver injury by improving the diversity and function of the
gut microbiota. Heliyon. 10:e355592024. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Jiang W, Zhu H, Liu C, Hu B, Guo Y, Cheng
Y and Qian H: In-depth investigation of the mechanisms of
Echinacea purpurea polysaccharide mitigating alcoholic liver
injury in mice via gut microbiota informatics and liver
metabolomics. Int J Biol Macromol. 209:1327–1338. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Lei Y, Chao S and Manxu Z: Regulation of
intestinal floras and their metabolism functions by Ganoderma
lucidum plysaceharide in mice with HepG2 cell-induced implanted
cancer. J Practical Hepatol. 24:476–479. 2021.
|
|
119
|
Jing G, Xu W, Ma W, Yu Q, Zhu H, Liu C,
Cheng Y, Guo Y and Qian H: Echinacea purpurea polysaccharide
intervene in hepatocellular carcinoma via modulation of gut
microbiota to inhibit TLR4/NF-κB pathway. Int J Biol Macromol.
261:1299172024. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Hu C, Xu B, Wang X, Wan WH, Lu J, Kong D,
Jin Y, You W, Sun H, Mu X, et al: Gut microbiota-derived
short-chain fatty acids regulate group 3 innate lymphoid cells in
HCC. Hepatology. 77:48–64. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Zhao J, He R, Zhong H, Liu S, Liu X,
Hussain M and Sun P: A cold-water extracted polysaccharide-protein
complex from Grifola frondosa exhibited anti-tumor activity
via TLR4-NF-κB signaling activation and gut microbiota modification
in H22 tumor-bearing mice. Int J Biol Macromol. 239:1242912023.
View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Che Y, Chen G, Guo Q, Duan Y, Feng H and
Xia Q: Gut microbial metabolite butyrate improves anticancer
therapy by regulating intracellular calcium homeostasis.
Hepatology. 78:88–102. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Bai G, Xie Y, Gao X, Xiao C, Yong T, Huang
L, Cai M, Liu Y, Hu H and Chen S: Selective impact of three
homogenous polysaccharides with different structural
characteristics from Grifola frondosa on human gut microbial
composition and the structure-activity relationship. Int J Biol
Macromol. 269:1321432024. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Wu L, Feng J, Li J, Yu Q, Ji J, Wu J, Dai
W and Guo C: The gut microbiome-bile acid axis in
hepatocarcinogenesis. Biomed Pharmacother. 133:1110362021.
View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Sun L, Cai J and Gonzalez FJ: The role of
farnesoid X receptor in metabolic diseases, and gastrointestinal
and liver cancer. Nat Rev Gastroenterol Hepatol. 18:335–347. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Song Y, Lau HC, Zhang X and Yu J: Bile
acids, gut microbiota, and therapeutic insights in hepatocellular
carcinoma. Cancer Biol Med. 21:144–162. 2023.PubMed/NCBI
|
|
127
|
Petrick JL, Florio AA, Koshiol J, Pfeiffer
RM, Yang B, Yu K, Chen CJ, Yang HI, Lee MH and McGlynn KA:
Prediagnostic concentrations of circulating bile acids and
hepatocellular carcinoma risk: REVEAL-HBV and HCV studies. Int J
Cancer. 147:2743–2753. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Ohtani N and Hara E: Gut-liver
axis-mediated mechanism of liver cancer: A special focus on the
role of gut microbiota. Cancer Sci. 112:4433–4443. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Sydor S, Best J, Messerschmidt I, Manka P,
Vilchez-Vargas R, Brodesser S, Lucas C, Wegehaupt A, Wenning C,
Aßmuth S, et al: Altered microbiota diversity and bile acid
signaling in cirrhotic and noncirrhotic NASH-HCC. Clin Transl
Gastroenterol. 11:e001312020. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Shen R, Ke L, Li Q, Dang X, Shen S, Shen
J, Li S, Liang L, Peng B, Kuang M, et al: Abnormal bile
acid-microbiota crosstalk promotes the development of
hepatocellular carcinoma. Hepatol Int. 16:396–411. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Wang A, Xiong W, Li J, Hu Y, Zou L and Liu
Y: The regulatory effects of bioactive polysaccharides on
intestinal function and bile acids: Chemical structures,
bioactivities, and mechanisms. Front Nutr. 11:14959932024.
View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Lin P, Chen S and Zhong S: Nutritional and
chemical composition of Sargassum zhangii and the physical
and chemical characterization, binding bile acid, and
cholesterol-lowering activity in HepG2 cells of its fucoidans.
Foods. 11:17712022. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Wang T, Tian T, Zhu Z, Fang S, Zhang L,
Peng X, Shi R, Li Y, Wu J and Ma Y: Gardenia jasminoides
Ellis. Polysaccharides alleviated cholestatic liver injury by
increasing the production of butyric acid and FXR activation.
Phytother Res. 38:5363–5375. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Fang S, Wang T, Li Y, Xue H, Zou J, Cai J,
Shi R, Wu J and Ma Y: Gardenia jasminoides Ellis
polysaccharide ameliorates cholestatic liver injury by alleviating
gut microbiota dysbiosis and inhibiting the TLR4/NF-κB signaling
pathway. Int J Biol Macromol. 205:23–36. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Cai J, Zhu Z, Li Y, Li Q, Tian T, Meng Q,
Wang T, Ma Y and Wu J: Artemisia capillaris Thunb.
Polysaccharide alleviates cholestatic liver injury through gut
microbiota modulation and Nrf2 signaling pathway activation in
mice. J Ethnopharmacol. 327:1180092024. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Wei J, Dai Y, Zhang N, Wang Z, Tian X, Yan
T, Jin X and Jiang S: Natural plant-derived polysaccharides
targeting macrophage polarization: A promising strategy for cancer
immunotherapy. Front Immunol. 15:14083772024. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Li X, Zhang H, Deng Y, Fang Q, Zhang X,
Ding S, Hou X and Du H: Huaier polysaccharides inhibits
hepatocellular carcinoma via gut microbiota mediated M2 macrophage
polarization. Int J Biol Macromol. 293:1393572025. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Liu X, Wu Z, Guo C, Guo H, Su Y, Chen Q,
Sun C, Liu Q, Chen D and Mu H: Hypoxia responsive nano-drug
delivery system based on angelica polysaccharide for liver cancer
therapy. Drug Deliv. 29:138–148. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Liu HM, Cheng J, Wang XY, Jiang Y, Ni J,
Zhang Y and Wang W: Structure identification of Ganoderma
lucidum spore polysaccharides and their antitumor activity in
vivo. Molecules. 29:23482024. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Wu BK, Chen QH, Pan D, Chang B and Sang
LX: A novel therapeutic strategy for hepatocellular carcinoma:
Immunomodulatory mechanisms of selenium and/or selenoproteins on a
shift towards anti-cancer. Int Immunopharmacol. 96:1077902021.
View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Gao X, Li X, Mu J, Ho CT, Su J, Zhang Y,
Lin X, Chen Z, Li B and Xie Y: Preparation, physicochemical
characterization, and anti-proliferation of selenium nanoparticles
stabilized by Polyporus umbellatus polysaccharide. Int J Biol
Macromol. 152:605–615. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Zhang S, Gao R, Ding B, Li J, Wang T, Chen
J, Li C, Jiao Y and Song L: Antihepatoma activity of Marsdenia
tenacissima polysaccharide-decorated selenium nanoparticles by
regulating the Bax/Bcl-2/caspases and p21/Akt/cyclin A2 signaling
pathways. Int J Biol Macromol. 279:1349812024. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Jiao J, Yu J, Ji H and Liu A: Synthesis of
macromolecular Astragalus polysaccharide-nano selenium complex and
the inhibitory effects on HepG2 cells. Int J Biol Macromol.
211:481–489. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Yang Z, Liu Z, Zhu J, Xu J, Pu Y and Bao
YL: Green synthesis and characterization of gold nanoparticles from
Pholiota adiposa and their anticancer effects on hepatic
carcinoma. Drug Deliv. 29:997–1006. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Xu J, Liu Z, Zhang S, Xiang J, Lan H and
Bao Y: Anti-hepatoma immunotherapy of Pholiota adiposa
polysaccharide-coated selenium nanoparticles by reversing M2-like
tumor-associated macrophage polarization. Int J Biol Macromol.
277:1336672024. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Lan H, Xu J, Zong W, Zhou L, Yang J, Xia Y
and Bao Y: Immunomodulatory effects of Fructus corni acidic
polysaccharide and its selenium nanoparticles composites in
hepatocellular carcinoma. Int J Biol Macromol. 282:1368182024.
View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Sun H, Nai J, Deng B, Zheng Z, Chen X,
Zhang C, Sheng H and Zhu L: Angelica sinensis
Polysaccharide-based nanoparticles for Liver-targeted delivery of
oridonin. Molecules. 29:7312024. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Anda-Flores YD, Rascón-Chu A, Campa-Mada
AC, Lizardi-Mendoza J, Tanori-Cordova J and Carvajal-Millan E:
Chapter 17-Polysaccharides nanoparticles as oral drug delivery
systems, in Natural Polysaccharides in Drug Delivery and Biomedical
Applications. Hasnain MS and Nayak AK: Academic Press; pp. 399–417.
2019
|
|
149
|
Wu Y, Liu L, Zhao Y and Zhao R:
Polysaccharides of vinegar-baked radix bupleuri promote the hepatic
targeting effect of oxymatrine by regulating the protein expression
of HNF4α, Mrp2, and OCT1. J Ethnopharmacol. 267:1134712021.
View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Wang X, Gan J, Han M, Wu Y, Liu L, Zhao Y
and Zhao R: Nano-pectic polysaccharide from vinegar-baked Radix
Bupleuri enhances methotrexate efficacy against hepatocellular
carcinoma via BCRP and MRP2 downregulation. Int J Biol Macromol.
318:1450462025. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Zhang Y, Cui Z, Mei H, Xu J, Zhou T, Cheng
F and Wang K: Angelica sinensis polysaccharide nanoparticles
as a targeted drug delivery system for enhanced therapy of liver
cancer. Carbohydr Polym. 219:143–154. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Sun H, Gao C, Yang Z, Song R, Zhu T, Sun
Y, Sheng H and Zhu L: Preparation and evaluation of
anti-hepatocellular carcinoma activity of oridonin nanoparticles
using Angelica sinensis polysaccharide and ursodeoxycholic
acid as carrier. Int J Biol Macromol. 327:1474692025. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Wang B, Lv B, Li H, Zhang J, Ding Y, Zhou
J, Bu M, Fan L and Han C: Design of self-assembled micelles based
on natural dual-targeting strategies and evaluation of their
anti-liver cancer effects as drug delivery systems. NPJ Precis
Oncol. 9:822025. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Guo C, Hou X, Liu Y, Zhang Y, Xu H, Zhao F
and Chen D: Novel Chinese angelica polysaccharide biomimetic
nanomedicine to curcumin delivery for hepatocellular carcinoma
treatment and immunomodulatory effect. Phytomedicine.
80:1533562021. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Zheng Z, Deng BQ, Chen XM, Zhu LQ and
Sheng HG: Preparation, characterization, and in vitro anti-liver
tumor activity of bufalin nanoparticles with Scrophularia
ningpoensis polysaccharide and ursodeoxycholic acid as carriers.
Zhongguo Zhong Yao Za Zhi. 50:3013–3023. 2025.(In Chinese).
PubMed/NCBI
|
|
156
|
Qin Y, Cao Z, Wang J, Liu C, Liao C, Huang
B, Liu Q, Xia B, Ning Q, Wei H and Yu CY: A Pectin-based delivery
nanoplatform with an optimized tradeoff between active targeting
and drug loading for hepatocellular carcinoma treatment. Mol Pharm.
22:5555–5566. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Li PL, Li CX, Xue YT, Li HH, Liu HB, He
XX, Yu GL and Guan HS: An HPLC method for microanalysis and
pharmacokinetics of marine sulfated polysaccharide PSS-loaded poly
lactic-co-glycolic acid (PLGA) nanoparticles in rat plasma. Mar
Drugs. 11:1113–1125. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Torres FG, Troncoso OP, Pisani A, Gatto F
and Bardi G: Natural polysaccharide nanomaterials: An overview of
their immunological properties. Int J Mol Sci. 20:50922019.
View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Serrano-Sevilla I, Artiga Á, Mitchell SG,
De Matteis L and de la Fuente JM: Natural polysaccharides for siRNA
Delivery: Nanocarriers based on chitosan, hyaluronic acid, and
their derivatives. Molecules. 24:25702019. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Sun Y, Jing X, Ma X, Feng Y and Hu H:
Versatile types of Polysaccharide-based drug delivery systems: From
Strategic design to cancer therapy. Int J Mol Sci. 21:91592020.
View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Ahmad A, Georgiou PG, Pancaro A, Hasan M,
Nelissen I and Gibson MI: Polymer-tethered glycosylated gold
nanoparticles recruit sialylated glycoproteins into their protein
corona, leading to off-target lectin binding. Nanoscale.
14:13261–13273. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Sharifi M, Cho WC, Ansariesfahani A,
Tarharoudi R, Malekisarvar H, Sari S, Bloukh SH, Edis Z, Amin M,
Gleghorn JP, et al: An updated review on EPR-based solid tumor
targeting nanocarriers for cancer treatment. Cancers (Basel).
14:28682022. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Shin SW, Jung W, Choi C, Kim SY, Son A,
Kim H, Lee N and Park HC: Fucoidan-Manganese dioxide nanoparticles
potentiate radiation therapy by Co-Targeting tumor hypoxia and
angiogenesis. Mar Drugs. 16:5102018. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Luo J, Li L, Zhu Z, Chang B, Deng F, Wang
D, Lu X, Zuo D, Chen Q and Zhou J: Fucoidan inhibits EGFR
redistribution and potentiates sorafenib to overcome
sorafenib-resistant hepatocellular carcinoma. Biomed Pharmacother.
154:1136022022. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Abdollah MRA, Ali AA, Elgohary HH and
Elmazar MM: Antiangiogenic drugs in combination with seaweed
fucoidan: A mechanistic in vitro and in vivo study exploring the
VEGF receptor and its downstream signaling molecules in hepatic
cancer. Front Pharmacol. 14:11089922023. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Wang Z, Qu K, Zhou L, Ren L, Ren B, Meng
F, Yu W, Wang H and Fan H: Apaf1 nanoLuc biosensors identified
lentinan as a potent synergizer of cisplatin in targeting
hepatocellular carcinoma cells. Biochem Biophys Res Commun.
577:45–51. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Liao W, Shan S, Xu J, Chen Z, Wu Y, Wen Y,
Chen W and Zhao C: Synergistic inhibition of hepatocarcinogenesis
by green alga Ulva lactuca polysaccharide and 5-fluorouracil
targeted SERPINH1. Phytomedicine. 148:1572662025. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Gou Y, Zheng X, Li W, Deng H and Qin S:
Polysaccharides produced by the mushroom Trametes
robiniophila murr boosts the sensitivity of hepatoma cells to
oxaliplatin via the miR-224-5p/ABCB1/P-gp axis. Integr Cancer Ther.
21:153473542210902212022. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Zhu T, Cheng Z, Peng X, Xing D and Zhang
M: HIF-1α RNAi combined with asparagus polysaccharide exerts an
antiangiogenesis effect on hepatocellular carcinoma in vitro and in
vivo. Evid Based Complement Alternat Med.
2021:99873832021.PubMed/NCBI
|
|
170
|
Zou Y, Wu S.Y..Zhang W, Zhang W, Shen X,
Qu X and Yao Q: Fucoidan improves tumour control and liver function
in TACE for unresectable hepatocellular carcinoma: A randomised
trial. Liver Int. 45:e703472025. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Wang W, Zhou H, Sen A, Zhang P, Yuan L and
Zhou S: Recent advances in the mechanisms and applications of
astragalus polysaccharides in liver cancer treatment: An overview.
Molecules. 30:27922025. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Wang D, Zhang Z, Zhao L, Yang L and Lou C:
Recent advances in natural polysaccharides against hepatocellular
carcinoma: A review. Int J Biol Macromol. 253:1267662023.
View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Tu J, Liu X, Li K, Liu H, Li J, Zhu J, Xia
N and Wang Q: A novel polysaccharide from Citrus aurantium
L.: Structural properties and antitumor activities in vitro and in
vivo. J Ethnopharmacol. 347:1197252025. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Dai KY, Liu C, Ji HY and Liu AJ:
Structural characteristics and anti-tumor activity of
alkali-extracted acidic polysaccharide extracted from Panax
ginseng. Int J Biol Macromol. 305:1412302025. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Yang J, Xiong K, Li T, Zhang M, Li Z, Wen
Z and Jiang Y: Anti-inflammatory effects of natural
polysaccharides: Molecular mechanisms and nanotherapeutic
applications. Front Immunol. 16:17233462025. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Pu W, Yu Y, Shi X, Shao Y, Ye B, Chen Y,
Song Q, Shen J and Li H: The effect of seasonal and annual
variation on the quality of Polygonatum cyrtonema hua
rhizomes. Plants (Basel). 13:34592024.PubMed/NCBI
|
|
177
|
Zhang R, Deng D, Song H, Liu H and Yang S:
Impact of extraction methods on soybean hull polysaccharides:
Structure and functional properties analysis. Food Chem X.
31:1030492025. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Velusami CC, Boddapati SR, Hongasandra
Srinivasa S, Richard EJ, Joseph JA, Balasubramanian M and Agarwal
A: Safety evaluation of turmeric polysaccharide extract: Assessment
of mutagenicity and acute oral toxicity. Biomed Res Int.
2013:1583482013. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Mututuvari TM and Tran CD: Synergistic
adsorption of heavy metal ions and organic pollutants by
supramolecular polysaccharide composite materials from cellulose,
chitosan and crown ether. J Hazard Mater. 264:449–459. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Cha M, Yan S, Zhang Y and Wang P: Progress
in the application of marine polysaccharide drug delivery systems
in tumor immunotherapy: Multiple mechanisms and material forms. Mar
Drugs. 23:3842025. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Yu Z, Guo F, Guo Y, Zhang Z, Wu F and Luo
X: Optimization and evaluation of astragalus polysaccharide
injectable thermoresponsive in-situ gels. PLoS One.
12:e01739492017. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Bergallo P, Castagnari V, Fernández A and
Mejía R: Regulatory initiatives to reduce sugar-sweetened beverages
(SSBs) in Latin America. PLoS One. 13:e02056942018. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Garfias Silva V, Cordova Aguilar MS,
Ascanio G, Aguayo JP, Pérez-Salas KY and Susunaga Notario ADC: Acid
hydrolysis of pectin and mucilage from cactus (Opuntia ficus) for
Identification and Quantification of Monosaccharides. Molecules.
27:58302022. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Argelaguet R, Arnol D, Bredikhin D, Deloro
Y, Velten B, Marioni JC and Stegle O: MOFA+: A statistical
framework for comprehensive integration of multi-modal single-cell
data. Genome Biol. 21:1112020. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Chalise P, Kwon D, Fridley BL and Mo Q:
Statistical methods for integrative clustering of Multi-omics data.
Methods Mol Biol. 2629:73–93. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Bozzi D, Neuenschwander S, Cruz Dávalos
DI, Sousa da Mota B, Schroeder H, Moreno-Mayar JV, Allentoft ME and
Malaspinas AS: Towards predicting the geographical origin of
ancient samples with metagenomic data. Sci Rep. 14:217942024.
View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Fang H, Kukje Zada D, Barra NG, Rodrigues
ELR and Schertzer JD: Protocol for determining gut microbiota
metabolites as substrates in mouse metabolism. STAR Protoc.
7:1043252026. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Xia M, Liu Q, Zhang W, Ge J and Mei Z:
Spatiotemporal dynamics of central nervous system diseases:
Advancing translational neuropathology via Single-cell and spatial
multiomics. MedComm (2020). 6:e703282025. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Zhou H, Pu Z, Lu Y, Zheng P, Yu H and Mou
L: Elucidating T cell dynamics and molecular mechanisms in
syngeneic and allogeneic islet transplantation through single-cell
RNA sequencing. Front Immunol. 15:14292052024. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Du H, Si G, Si J, Song X and Si F:
Integration of single-cell RNA and bulk RNA sequencing revealed
malignant ductal cell heterogeneity and prognosis signatures in
pancreatic cancer. Front Immunol. 16:15791842025. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Koirala M, Yan L, Mohamed Z and DiPaola M:
AI–Integrated QSAR modeling for enhanced drug discovery: From
classical approaches to deep learning and structural insight. Int J
Mol Sci. 26:93842025. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
D'Souza AA and Devarajan PV:
Asialoglycoprotein receptor mediated hepatocyte
targeting-Strategies and applications. J Control Release.
203:126–139. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Wang M, Li C, Li J, Hu W, Yu A, Tang H, Li
J, Kuang H and Zhang H: Extraction, purification, structural
characteristics, biological activity and application of
polysaccharides from Portulaca oleracea L. (Purslane): A
review. Molecules. 28:48132023. View Article : Google Scholar : PubMed/NCBI
|