Epigenetics refers to heritable phenotypical changes
that occur without alterations in the underlying DNA sequence
(1). Cell differentiation is
driven by the establishment of distinct epigenetic landscapes and
transcriptional programs, which determine lineage commitment and
cell fate (2). Epigenetic
regulation encompasses diverse mechanisms, including
post-translational histone modification and non-coding (nc)RNA
expression, as well as DNA and RNA methylation (3). Since Waddington (4) proposed the concept of the epigenetic
landscape (4), the contribution of
epigenetic regulation to the development and progression of
systemic diseases has been increasingly recognized (5,6). In
numerous disorders such as hematological and cardiovascular
disease, Alzheimer's disease, cancer and musculoskeletal
conditions, epigenetic mechanisms serve key roles in pathogenesis
(7–12).
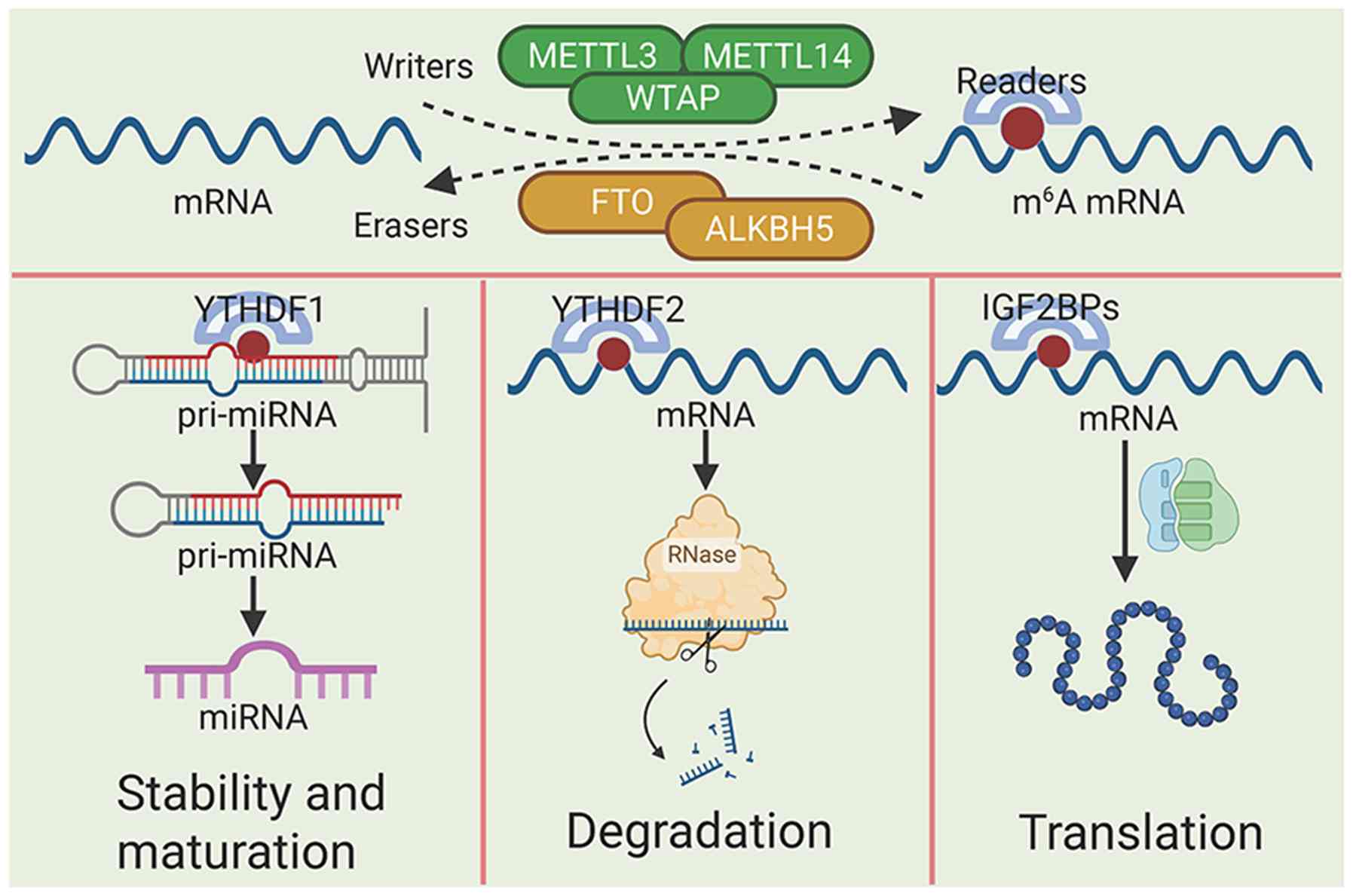
RNA modification, a key regulatory mechanism within
the epigenetic landscape, acts as an intermediary between DNA,
proteins and effector molecules, serving an important role in
post-transcriptional gene regulation (13). In addition to the canonical
nucleotides A, C, G and U, >100 distinct chemical modifications
have been identified in RNA (14).
For example, a classical mRNA contains a 5′cap structure that
enables ribosome recognition and provides protection from
degradation, as well as a 3′poly-A tail that regulates
transcription termination, facilitates nuclear export and enhances
translation efficiency (15).
Following the discovery of cap and tail modifications, numerous
internal RNA modifications have been identified, such as
N1-methyladenosine (16),
N6-methyladenosine (m6A) (17,18)
and 5-methylcytosine (19). Among
these, m6A is the most abundant and dynamically
regulated internal RNA modification in eukaryotic cells (20).
Skeletal system diseases, including osteoporosis
(OP), osteoarthritis (OA), rheumatoid arthritis (RA), ankylosing
spondylitis, lumbar disc herniation and cervical spondylosis,
represent a notable category of disorders worldwide (21). OA affected 595 million people
globally in 2020, rheumatoid arthritis 17.6 million, and lower back
pain 619 million, with all three projected to increase further by
2050; OP is common, with a pooled global prevalence of 19.7%
(22). Advances in epigenetics
have provided novel perspectives for understanding the
pathogenesis, diagnosis and treatment of skeletal disease (23). The present review aimed to
summarize m6A-mediated regulation of skeletal
development and its involvement in the diagnosis and treatment of
skeletal disorder, with the aim of providing novel insight and
therapeutic strategies for clinical management.
WTAP is a splicing-related protein encoded on
chromosome 6q25-27. It consists of 396 amino acids with a molecular
weight of ~46 kDa. WTAP regulates the cell cycle by stabilizing
cyclin A2 and CDK2 mRNA and thereby promoting G2/M and G1/S
progression (42,43). It contributes to tumorigenesis
primarily through m6A-dependent control of target
transcripts involved in proliferation and survival, including
pathways associated with glycolysis, PI3K/AKT signaling, NF-κB
activation and drug resistance. In addition, WTAP is essential for
sustaining the Sertoli cell-dependent spermatogonial stem cell
niche, and its loss impairs spermatogonial stem cell maintenance
and spermatogenesis, supporting its role in germ cell proliferation
and development (42,44). WTAP itself lacks methyltransferase
activity but serves as a regulatory scaffold of the m6A
writer complex by interacting with METTL3 and METTL14, facilitating
their recruitment to target RNAs, and promoting their accumulation
in nuclear speckles, thereby enabling efficient m6A
deposition in vivo. In the absence of WTAP, the binding of
METTL3 to mRNA is notably decreased.
YTHDC1 mediates the nuclear export of methylated RNA
(MeR). Knockout of YTHDC1 leads to nuclear accumulation of
transcripts, whereas cytoplasmic mRNA levels gradually decrease.
Conversely, overexpression of YTHDC1 has been shown to reduce
nuclear mRNA levels (69). YTHDC2
is highly expressed in mouse testicular tissue (31,70).
Its loss causes decreased testis size, degeneration of seminiferous
tubules, depletion of germ cells, loss of mature spermatozoa, and
meiotic arrest, ultimately impairing spermatogenesis and causing
male infertility. YTHDC2 facilitates transcript translation and
accelerates mRNA degradation, although the precise mechanisms
remain to be fully elucidated (31,71).
The oxidative demethylation mechanism helps prevent
methyltransferases from acting on RNA in nucleolar regions and
ensures that demethylation is not readily reversed (74). FTO exhibits substrate selectivity,
primarily catalyzing the removal of m6A modifications
within mRNA rather than at the 5′cap. In addition, FTO demethylates
m6A in nuclear mRNA and N6,2′-O-dimethyladenosine at the
5′cap in the cytoplasm (76).
ALKBH5, the second m6A demethylase identified after FTO,
belongs to the ALKB protein family, which comprises nine homologs.
ALKBH5 deficiency leads to increased m6A levels in mRNA,
resulting in abnormal testicular development and apoptosis of germ
cells in mice (36). RNA-binding
motif protein 33 (RBM33) is a key auxiliary factor for ALKBH5.
RBM33 recruits ALKBH5 to m6A-modified substrates and
activates its demethylation activity by removing minor
ubiquitin-like modifications (77). ALKBH5 is also key for oocyte
meiosis, as its deficiency causes oocyte developmental arrest,
disrupts RNA stability and leads to excessive translation (78). However, the precise mechanistic
differences between FTO and ALKBH5 remain to be fully elucidated.
Fig. 1 summarizes the regulatory
effects of m6A modification on mRNA.
ncRNA refers to RNA molecules that do not encode
proteins and are classified into two primary types: Structural and
regulatory ncRNA. Regulatory ncRNA includes long ncRNA (lncRNA),
microRNA (miRNA) and circular RNA (circRNA) (79). miRNA is a single-stranded RNA
molecule ~22 nucleotides in length. Its biogenesis begins in the
nucleus, where primary (pri)-miRNA is cleaved by Drosha in
conjunction with DiGeorge critical region 8 (DGCR8) to generate
pre-miRNA. DGCR8 facilitates localization by directing Drosha to
specific cleavage sites. The second processing step occurs in the
cytoplasm, whereby Dicer cleaves pre-miRNA to form a
double-stranded miRNA duplex. Subsequently, one strand of this
duplex associates with target mRNA to form the miRNA effector
within the miRNA-induced silencing complex, while the other strand
is released and degraded (80). At
the molecular level, m6A regulates ncRNAs primarily
through two mechanisms: It reshapes local RNA secondary structures
to alter the accessibility of RNA-binding motifs and recruits
specific RNA-binding proteins, such as DGCR8 and HNRNPA2B1, thereby
controlling ncRNA processing, stability, localization, and
function. In mammals, METTL3 deposits m6A on pri-miRNAs,
which promotes their recognition and processing by the
Microprocessor component DGCR8. Accordingly, METTL3 depletion
reduces DGCR8 binding to pri-miRNAs, leads to the accumulation of
unprocessed pri-miRNAs, and globally decreases mature miRNA
abundance (20,81,82).
In addition, HNRNPA2B1 acts as a nuclear m6A reader that
binds a subset of methylated pri-miRNAs and facilitates
microprocessor-dependent miRNA processing (20,81).
Studies have shown that the m6A reader protein heterogeneous
nuclear ribonucleoprotein A2/B1 (HNRNPA2B1) participates in
pri-miRNA processing (81,82). Mechanistically, HNRNPA2B1 binds a
subset of m6A-marked pri-miRNAs and interacts with the
microprocessor component DGCR8, thereby facilitating DGCR8
association with these pri-miRNA transcripts and promoting their
processing (81–84). Consistently, depletion of HNRNPA2B1
impairs DGCR8 binding, causes the nuclear accumulation of
unprocessed pri-miRNAs and reduces the levels of a subset of mature
miRNAs (81,82). These findings indicate that
HNRNPA2B1 functions as a nuclear m6A reader that
mediates, at least in part, METTL3/m6A-dependent
pri-miRNA maturation (82).
m6AHNRNPA2B1 participates in the processing of a subset
of pri-miRNAs rather than globally regulating all miRNAs. Knockdown
of HNRNPA2B1 decreases the levels of these HNRNPA2B1-dependent
miRNAs, at least in part because HNRNPA2B1 facilitates DGCR8
association with specific m6A-marked pri-miRNA
transcripts and thereby promotes their processing. In addition,
HNRNPA2B1 has been reported to bind m6A-modified RNAs
and mediate m6A-dependent nuclear RNA processing events
(81,82). Notably, miRNAs promote m6A
modification and influence methylation-site selection by
guiding METTL3 to target mRNAs through sequence pairing. After
Dicer-dependent maturation, some miRNAs relocalize to the nucleus,
where they pair with target transcripts near candidate m6A
motifs, facilitate METTL3 recruitment, and enhance local
m6A deposition (85,86).
Accordingly, perturbation of miRNA abundance or sequence alters m6A
formation at cognate sites, while Dicer depletion
reduces, and Dicer overexpression increases, overall m6A
levels (87). These
findings indicate a reciprocal association between miRNA and m6A
modification, whereby miRNAs regulate and are regulated
by m6A. However, m6A-dependent regulation of miRNA
biogenesis has been studied predominantly at the initiation stage,
where METTL3-mediated m6A deposition on pri-miRNAs
promotes DGCR8/microprocessor recognition and processing (81,82).
By contrast, the role of m6A in later pre-miRNA cleavage
remains less well characterized and appears to be context
dependent, with limited reports showing that m6A readers can
facilitate the processing of selected pre-miRNAs through AGO2- or
DICER-associated mechanisms (81,88).
lncRNA is typically >200 nucleotides and performs
diverse biological functions, including cis- and trans-regulation,
formation of nuclear domains and regulation of RNA function
(89). The X-inactive specific
transcript (XIST) mediates gene silencing on the X chromosome.
Methylation of XIST by the METTL3-WTAP complex is key for its
function and the m6A reader YTHDC1 promotes
XIST-mediated gene silencing by binding the methylated transcript,
demonstrating m6A involvement in lncRNA functional
regulation (90). In colorectal
cancer (CRC), METTL14 knockdown decreases m6A
modification on XIST, leading to increased XIST expression and
enhanced proliferation, migration, and invasion of CRC cells
(90,91). YTHDF2, rather than the other
m6A readers tested, recognized m6A-modified
XIST and promoted its degradation m6A m6A
(62,90–92).
circRNA lacks a free 5′cap and 3′poly-A tail, forming a covalently
closed loop structure. circRNA can be categorized into three types:
Exonic, exonic-intronic and intronic. Exonic circRNAs are the most
abundant and are primarily localized in the cytoplasm, whereas
exonic-intronic and intronic circRNAs are mainly found in the
nucleus. circRNAs perform multiple functions, including serving as
miRNA sponges, regulating transcription and translation,
facilitating protein splicing and transport, modulating
protein-protein interactions and serving as templates for protein
translation (93). Yang et
al (94) demonstrated that
m6A modification promotes the translational function of
circRNAs. Specifically, the METTL3-METTL14 complex may mediate
m6A modification of circRNAs without affecting their
stability. Translation of circRNAs requires eukaryotic translation
initiation factor 4 γ-2 (eIF4G2), which is key for the initiation
of eukaryotic circRNA translation; deficiency of eIF4G2 decreases
circRNA translation. YTHDF3 serves as a key recognition factor,
recruiting eIF4G2 to m6A-modified circRNAs to initiate
translation, while FTO serves as a negative regulator in this
process (94). Subsequent research
has shown that m6A modification may not only facilitate
circRNA translation but also serve a key role in circRNA
degradation, innate immune responses and tumor development
(95).
BMSCs are multipotent stromal cells capable of
differentiating into osteoblasts, chondrocytes and adipocytes
(112). Their lineage allocation
is key for skeletal development and homeostasis, and a shift toward
adipogenesis is closely associated with increased marrow adiposity
and bone loss (113). METTL3 is a
key epitranscriptomic regulator of BMSC fate through multiple
downstream pathways (114,115).
In vivo, conditional deletion of Mettl3 in mesenchymal stem
cells impairs bone formation, reduces osteogenic differentiation
potential, and increases marrow adiposity, whereas Mettl3
overexpression in MSCs protects mice from estrogen
deficiency-induced OP (114).
Mechanistically, METTL3 promotes the translation of Pth1r and
maintains the PTH/PTH1R signaling axis in MSCs; it also suppresses
adipogenic differentiation through the
m6A-YTHDF2-JAK1/STAT5/C/EBPβ pathway, and enhances
osteogenic differentiation by regulating m6A
modification of RUNX2 and precursor miR-320 (102,113–116). These findings identify METTL3 as
a central regulator of the osteogenic-adipogenic balance in BMSCs
(113–115). The parathyroid hormone (PTH)/PTH
receptor-1 (PTH1R) signaling pathway is downstream of METTL3 action
and reduced METTL3 expression impairs PTH1R translation efficiency
(114). In addition, METTL3
expression is markedly downregulated in ovariectomy (OVX)-induced
OP models, with in vitro overexpression of METTL3 restoring
the osteogenic potential of BMSCs (117). Tian et al (118) demonstrated that METTL3
downregulation decreases both early and late stages of osteoblast
differentiation in BMSCs, accompanied by decreased alkaline
phosphatase (ALP) activity and mineralized nodule formation. This
suggests METTL3-mediated m6A modification is pivotal in
osteoblast differentiation. Downstream targets of m6A, including
osteogenesis-associated genes such as RUNX2 and osterix, exhibit
decreased expression upon METTL3 downregulation (114,118).
METTL14 exhibits similar regulatory functions. A
recent study showed that METTL14 alleviates H2O2-induced impairment
of osteoblast differentiation in MC3T3-E1 murine calvaria-derived
clonal preosteoblastic/osteoblast-like cell line) (125). Mechanistically, GLUT3 was
identified as an m6A-modified target of METTL14, and
YTHDF1 participated in promoting GLUT3 expression, thereby
enhancing osteogenesis under oxidative stress conditions (125,126). The demethylase FTO serves an
important role in normal bone development. Zhang et al
(127) demonstrated that FTO
expression is key in bone formation: FTO knockout mice exhibit
reduced trabecular bone volume and number, resulting in bone
formation defects. FTO is key in osteoblast differentiation; its
deficiency increases osteoblast apoptosis and renders cells more
susceptible to physical and chemical stressors such as ultraviolet
radiation and H2O2, partly via the NF-κB signaling
pathway. These findings underscore the importance of FTO in
maintaining normal bone formation (127).
Osteoclasts are multinucleated cells derived from
hematopoietic SCs and differentiate from osteoclast precursors upon
stimulation by macrophage colony-stimulating factor and receptor
activator of NF-κB ligand (RANKL) (103). In the skeletal system,
osteoblasts synthesize and secrete RANKL and osteoprotegerin (OPG).
Osteoclast precursors express RANK, which binds RANKL, promoting
differentiation into osteoclasts and enhancing bone resorption. OPG
serves as a decoy receptor by competing with RANKL, thereby
delaying osteoclast precursor differentiation and inhibiting bone
resorption, maintaining the balance between osteoblast and
osteoclast activity (128).
METTL3 knockout results in enlarged osteoclasts
with decreased resorptive capacity. METTL3 deficiency suppresses
the expression of osteoclast-specific genes, including nuclear
factor of activated T cells 1 (NFATC1), c-Fos, cathepsin K, acid
phosphatase 5 and dendrocyte-expressed seven transmembrane protein,
while upregulating the cell fusion-specific gene ATP6V0D2, METTL3
knockout enhances ATP6V0D2 mRNA stability, thereby inhibiting
osteoclast differentiation and bone resorption activity (129). Similarly, during LPS-induced
osteoclastogenesis, both total m6A content and METTL3
expression decrease. METTL3 knockdown decreases osteoclast numbers,
the expression of osteoclast-related genes and bone resorption
area, while increasing osteoclast apoptosis and expression of
pro-apoptotic proteins. Mechanistically, METTL3 deficiency
stabilizes nitric oxide synthase 2 mRNA, thereby inhibiting
osteoclast differentiation and promoting apoptosis (130). However, as aforementioned, METTL3
deficiency also impairs osteoblast differentiation and maturation.
The mechanism by which METTL3 coordinates the maturation of
osteoblasts and osteoclasts to regulate bone formation remains
unclear. m6A modification within the 1916–1992 bp region
of osteoblast-derived exosomal circ_0008542, particularly at the
A1956 site, promotes osteoclast differentiation and bone
resorption, and that these effects are attenuated by METTL3
inhibition or ALKBH5 overexpression, highlighting an
m6A-dependent mechanism of osteoblast-osteoclast
crosstalk m6A m6A (131).
Disruption of regulatory mechanisms that govern
skeletal development typically predisposes tissue to degenerative
and inflammatory disease later in life. Given the key role of
m6A modification in skeletal development and cell
homeostasis, dysregulation of m6A regulators is
increasingly implicated in the pathogenesis of skeletal disorder.
Aberrant m6A regulation is associated with notable
skeletal diseases, including OP, OA, RA and intervertebral disc
degeneration (IVDD) (97,132–134).
OP is a skeletal disorder characterized primarily
by decreased bone mass, deterioration of bone microarchitecture,
decreased bone strength and an increased risk of fractures
(135). A decrease in bone
mineral density ≥2.5 SD compared with age- and sex-matched adults
is diagnostic for OP (136). With
advancing age, OP becomes more common, and its prevalence is
notably higher in women than in men. Based on US NHANES 2017–2018
data cited in the 2025 USPSTF Recommendation Statement, the
age-adjusted prevalence of OP among adults aged 50 years or older
was 12.6%, including 19.6% in females and 4.4% in men; among those
aged 65 years or older, the prevalence increased to 27.1% in women
and 5.7% in men (137). OP
markedly increases fracture risk, imposing a health burden on
patients (138). A key cause of
OP is an imbalance between osteoblast and osteoclast activity.
Therefore, strategies that promote osteoblast differentiation while
inhibiting osteoclast activity are key in the prevention and
treatment of OP (139). Daily
supplementation with calcium and vitamin D improves bone health,
while representative pharmacological treatments include
bisphosphonates (140),
calcitonin (141) and strontium
(142).
Changes in bone density serve as the primary
diagnostic criterion for OP. Advances in epigenetic research have
revealed additional molecular mechanisms underlying OP, among which
m6A-mediated regulation serves a key role, offering
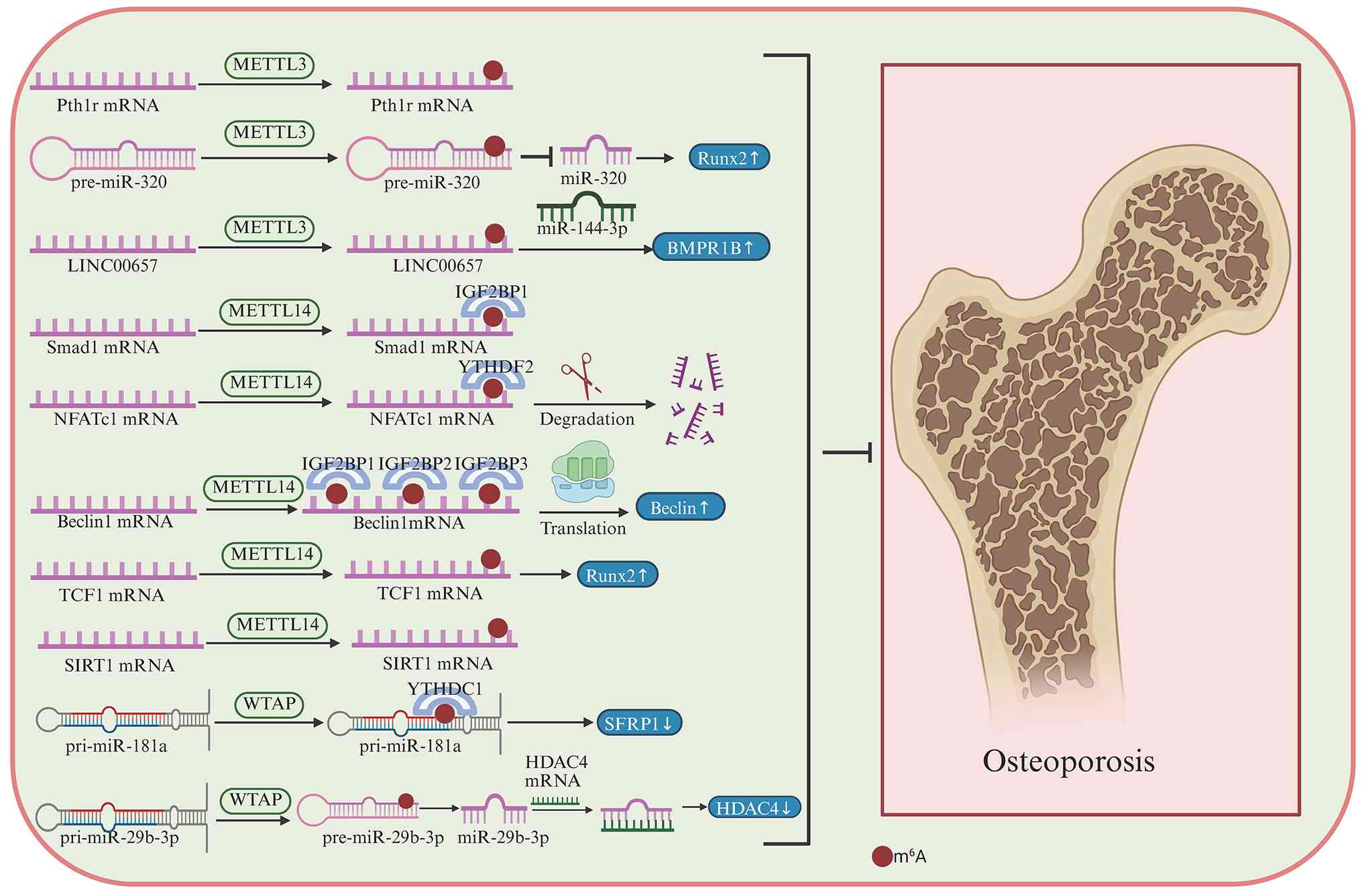
novel insights for diagnosis and therapeutic intervention (132,143). Mesenchymal stem cells (MSCs) are
multipotent progenitors whose lineage allocation is key for
skeletal homeostasis. A shift in MSC fate from osteogenesis toward
adipogenesis disrupts bone homeostasis and contributes to
osteoporosis. In this context, METTL3 is a key regulator of the
osteogenic-adipogenic balance in bone marrow MSCs. Conditional loss
of Mettl3 in MSCs impairs bone formation, decreases bone mass, and
increases marrow adiposity, whereas Mettl3 overexpression protects
mice from ovariectomy-induced osteoporosis. Mechanistically, METTL3
promotes translation of Pth1r mRNA and maintains PTH/PTH1R
signaling, thereby favoring osteogenic commitment over adipogenic
differentiation (114). RUNX2, a
member of the RUNT-related transcription factor family, serves a
key role in osteoblast differentiation and is regulated by numerous
miRNAs (144). Studies have
reported that METTL3 decreases the abundance of miRNA-320 by
enhancing m6A modification of pre-miRNA-320, thereby
increasing RUNX2 expression and promoting osteogenesis, exerting an
anti-osteoporotic effect (145,146). In addition, METTL3 mediates
m6A modification of long intergenic non-protein coding
(LINC)-00657, promoting bone formation by serving as a competing
endogenous RNA to upregulate bone morphogenetic protein receptor
type 1B by sponging miRNA-144-3p (147).
OA is a degenerative joint disease characterized by
joint pain, swelling, stiffness and restricted mobility. OA affects
~15% of individuals aged 30 years or older worldwide, and the
number of people living with OA is projected to approach 1 billion
by 2050, driven primarily by population ageing, population growth,
and increasing obesity (163). OA
pathogenesis primarily involves joint inflammation, cartilage
degradation and deformation and osteophyte formation (164). Therapeutic strategies for
alleviating OA pain primarily rely on cyclooxygenase-2 selective
inhibitors, such as celecoxib (165) and meloxicam (166), as well as traditional
non-steroidal anti-inflammatory drugs including diclofenac
(167). However, these treatments
are symptomatic, providing pain relief without slowing disease
progression or promoting cartilage repair and fail to improve the
long-term quality of life for patients. Consequently, identifying
molecular targets that inhibit chondrocyte apoptosis, improve the
inflammatory microenvironment and promote cartilage regeneration
has become a key research focus (168,169). m6A RNA modification
represents a promising avenue in this regard. Studies investigating
the role of m6A in OA pathogenesis have primarily
focused on autophagy, fibrosis, oxidative stress and associated
processes (170,171).
Aberrant activation of fibroblast-like synoviocytes
(FLSs) is a key inflammatory driver in OA. Under physiological
conditions, FLSs reside in the synovial intimal lining and
contribute to joint homeostasis. Following activation by
inflammatory stimuli, FLSs acquire an aggressive phenotype
characterized by enhanced proliferation, migration and invasion
(172). In this state, FLSs
participate in joint destruction by promoting synovial hyperplasia
and pannus formation, and by directly invading adjacent cartilage
and bone (173). Mechanistically,
the DDR2/annexin A2/MMP-13 loop promotes FLS migration and
invasion, whereas RasGRP4 contributes to pathological FLS
proliferation, driving persistent synovitis and structural joint
damage (174,175). Impaired FLS homeostasis and
defective autophagy are associated with OA development. Reduced
autophagy has been documented in OA tissues and patient-derived
cells, and in surgically induced OA, particularly in articular
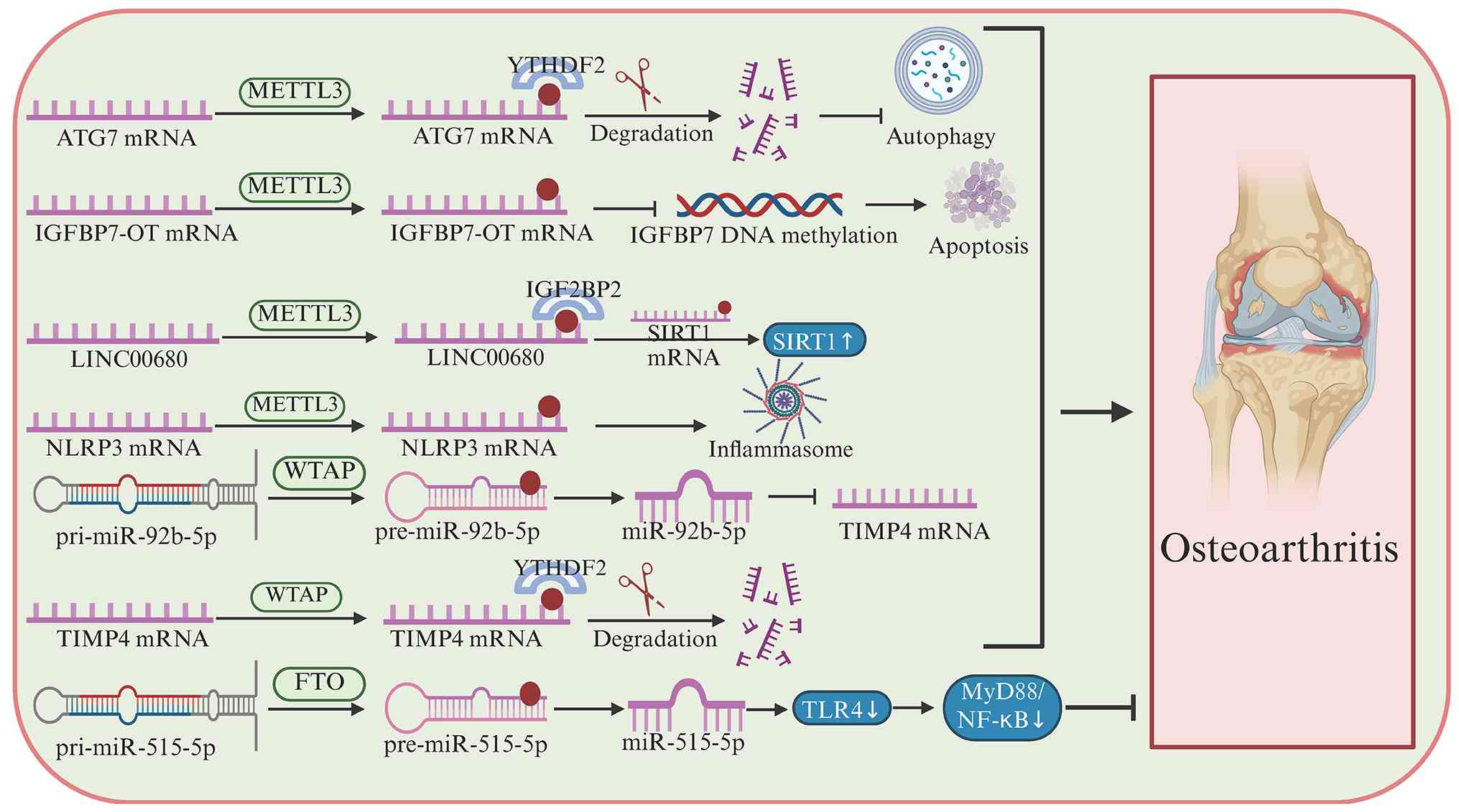
cartilage (171). In OA-FLS,
METTL3-mediated m6A modification of autophagy-related
gene 7 (ATG7) promotes cellular senescence (176). Mechanistically, YTHDF2 recognizes
m6A-modified ATG7 mRNA and decreases its RNA stability,
thereby reducing ATG7 protein expression, impairing autophagic
flux, and accelerating OA progression (171,176,177). In osteoarthritis, m6A
modification directly regulates chondrocyte survival and
extracellular matrix (ECM) homeostasis by modulating the stability,
maturation, or translation of key RNAs that control apoptotic,
inflammatory-catabolic, and anabolic pathways. For example,
increased METTL3-mediated m6A promotes NF-κB activation
in chondrocytes, enhances apoptosis and inflammatory responses, and
shifts ECM metabolism toward degradation, as reflected by increased
MMP13 and collagen X and reduced aggrecan and collagen II. By
contrast, FTO-mediated demethylation stabilizes SMAD2 mRNA, thereby
preserving anabolic signaling and restraining cartilage catabolism.
In addition, WTAP-dependent m6A regulation aggravates
chondrocyte injury either by enhancing CA12 mRNA stability or by
promoting pri-miR-92b maturation and YTHDF2-dependent TIMP4
downregulation, leading to reduced chondrocyte viability, increased
apoptosis, and ECM degradation (178–180).
Unlike miRNAs, lncRNAs do not encode proteins but
regulate gene expression at multiple levels. In OA, lncRNA
IGFBP7-OT is upregulated in osteoarthritic cartilage and is
positively correlated with its sense gene, IGFBP7. Functionally,
IGFBP7-OT overexpression inhibits chondrocyte viability, promotes
apoptosis, and reduces the expression of extracellular matrix
components, including collagen II and aggrecan, whereas its
silencing exerts the opposite effects. Mechanistically, the
upregulation of IGFBP7-OT is partially controlled by
METTL3-mediated m6A modification. Increased IGFBP7-OT,
in turn, suppresses the occupancy of DNMT1 and DNMT3a on the IGFBP7
promoter, reduces promoter methylation, and thereby upregulates
IGFBP7 expression, ultimately promoting OA progression (181). Similarly, in IL-1β-stimulated
chondrocytes, METTL3 increases the m6A modification and
stability of LINC00680. LINC00680 interacts with the
m6A-containing 3′-UTR of SIRT1 mRNA and enhances its
stability. Functionally, silencing LINC00680 partially rescues
chondrocyte proliferation and attenuates ECM degradation under
inflammatory conditions (182).
m6A regulation alters OA progression through
miRNA-dependent control of macrophage NLRP3 inflammasome signaling.
Activation of the NLRP3 inflammasome is a key source of IL-1β- and
IL-18-mediated inflammatory responses in OA. EVs derived from MSCs
inhibit m6A modification of NLRP3 mRNA by decreasing
METTL3 expression, with miR-1208 serving as a central upstream
regulator. This indicates that miRNAs modulate m6A
modification from an upstream position (183).
WTAP is upregulated in OA and promotes chondrocyte
apoptosis while impairing ECM homeostasis by inhibiting ECM
synthesis and accelerating ECM degradation. Mechanistically,
WTAP-mediated m6A modification enhances the processing
of pri-miR-92b into mature miR-92b-5p, which directly suppresses
TIMP4; in addition, WTAP facilitates YTHDF2-dependent degradation
of m6A-modified TIMP4 mRNA, leading to markedly reduced
TIMP4 expression in OA chondrocytes (184). As a demethylase, FTO inhibits OA
progression. By reducing the m6A level of
pri-miR-515-5p, FTO suppresses the toll-like receptor 4/myeloid
differentiation primary response 88/NF-κB signaling pathway,
thereby exerting anti-inflammatory effects in OA (185).
RA is a chronic autoimmune-inflammatory disease
that primarily affects the joints and may involve numerous organs
and tissue (186). Its hallmark
pathological features include persistent synovitis with progressive
cartilage destruction and bone erosion. In RA, protein
citrullination generates neoepitopes that are recognized by ACPAs,
thereby triggering autoimmune inflammation (187). This response promotes synovial
fibroblasts and macrophages to produce pro-inflammatory mediators,
particularly TNF-α, IL-1β, IL-6, IL-8, IL-17, and GM-CSF, which
drive joint inflammation and subsequent cartilage and bone damage
(188). Sustained autoimmune and
vascular inflammation contributes to systemic complications,
including small vessel vasculitis, interstitial lung disease,
pleuritis/pericarditis, cardiovascular disease, secondary
amyloidosis, and lymphoma (189).
In RA, FLSs serve as major effector cells in synovial hyperplasia
and joint destruction. Activated RA-FLSs exhibit a hyperplastic and
aggressive phenotype characterized by increased proliferation,
migration, and invasion, and they produce pro-inflammatory
mediators, including TNF-α, IL-1β, IL-6, and IL-8. Through these
pathogenic properties, RA-FLSs contribute to pannus formation and
promote cartilage and bone destruction (190,191). METTL3 expression is elevated in
RA synovial tissue and RA-FLSs (192). METTL3 silencing decreases IL-6
production, downregulates MMP-3 and MMP-9, and suppresses FLS
proliferation, migration, and invasion, whereas METTL3
overexpression exerts the opposite effects. Mechanistically, METTL3
may regulate FLS activation and inflammatory responses via the
NF-κB signaling pathway, thereby contributing to RA progression
m6A (192).
LIM and SH3 domain protein 1 (LASP1) is a key
epigenetic regulator in RA. LASP1 expression is markedly elevated
in cartilage tissue and FLSs of patients with RA (197,198). Loss of LASP1 impairs the
invasiveness of FLSs, stabilizes cell-cell contacts and weakens the
ability of FLSs to form zipper-like adhesions with the cadherin-11
complex, thereby decreasing bone destruction in RA mouse models
(198,199). METTL14 is markedly upregulated in
RA rats and its silencing suppresses TNF-α-induced LASP1 expression
as well as Src/AKT signaling pathway activation in FLSs, suggesting
that METTL14 may promote RA progression through the LASP1/Src/AKT
axis (196). In addition, METTL14
may affect the NF-κB pathway to suppress inflammatory responses in
RA. Downregulation of METTL14 decreases m6A levels in
TNF-α-induced protein-3 mRNA, resulting in activation of NF-κB
signaling and subsequent elevation of IL-6 and IL-17 levels
(200).
IVDD is a notable contributor to spinal pain,
particularly lower back pain, and is also implicated in neck pain
(206,207). Globally, low back pain affected
619 million people in 2020; this is projected to increase to 843
million by 2050, while neck pain affected 203 million people in
2020 and is projected to rise to 269 million by 2050 (208,209). IVDD is associated with
inflammation (210), autophagy
(211) and oxidative
stress-induced damage (212,213). Global dysregulation of
m6A machinery has been observed during IVDD progression
(214). In a bipedal
standing-induced mouse model of IVDD, the expression of
m6A writers, including METTL3, METTL14 and WTAP, is
elevated in nucleus pulposus (NP) tissue compared with controls,
indicating a positive association between enhanced m6A
modification and disc degeneration (214).
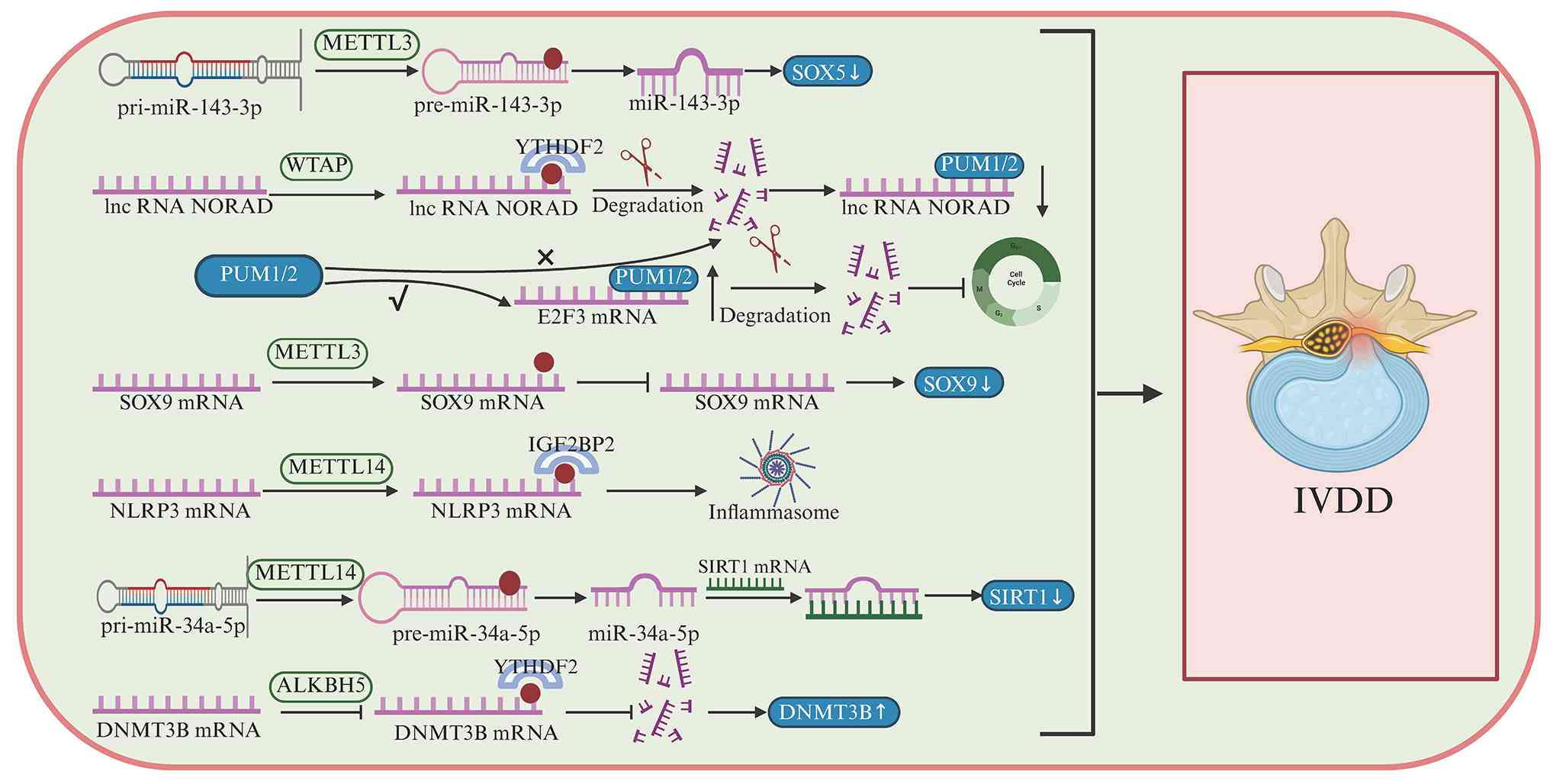
Endplate cartilage is a key disc component and
apoptosis of endplate chondrocytes is a key driver of IVDD
(222), with iron
overload-mediated oxidative stress contributing to this process
(223). Consistent with a
mechanotransduction-driven m6A-transcription factor axis
in endplate degeneration, mechanical loading increases both METTL3
expression and global m6A levels in endplate
chondrocytes (223). SOX9 is an
m6A target and METTL3 overexpression enhances
m6A modification of SOX9 precursor mRNA, decreases SOX9
RNA abundance and promotes IVDD, whereas METTL3 inhibition
alleviates disease severity (224). Collectively, these findings
indicate that SOX5- and SOX9-associated observations in IVDD
reflect distinct, context-dependent m6A regulatory
programs, namely an inflammatory NP-associated pathway impacting
SOX5 and lncRNA stability, as well as a mechanically driven
endplate pathway suppressing SOX9, both converging on ECM
dysregulation and disc degeneration.
SIRT1 expression is decreased in degenerated IVD
tissue. METTL14 functions as an upstream regulator of SIRT1,
mediating the m6A modification of miRNA-34a-5p, which
suppresses SIRT1 expression and induces senescence in NP cells
(225–228). ALKBH5 expression is increased in
aging IVD tissues, and that its silencing partially alleviates
age-related degeneration (229,230). Mechanistically, reduced H3K9me3
enrichment at the ALKBH5 promoter contributes to its upregulation,
whereas ALKBH5 promotes IVD cell senescence by reducing m6A
modification of its downstream target DNMT3B m6A
(231). m6AFTO and
YAP1 are downregulated in degenerative nucleus pulposus tissues
from patients with IVDD and in rat models, and this is associated
with increased m6A modification of YAP1 transcripts; however, the
reader proteins mediating this effect remain unclear and may be
related to mRNA stability or degradation pathways (232,233).
Although high-throughput sequencing technology
rapidly generates data on thousands of m6A sites,
sequencing biases and technical limitations may still result in
false positives or negatives. Therefore, methods capable of
detecting m6A at single-gene resolution are key. The
single-base extension and ligation-based quantitative PCR
amplification method leverages m6A inhibition of DNA
polymerase and ligase activity, enabling rapid, convenient
detection and quantification of site-specific m6A
modification (247).
Not applicable.
The present study was supported by the Natural Science
Foundation of Heilongjiang Province (grant no. LH2021H092) and the
Heilongjiang Provincial Traditional Chinese Medicine Research
Project (grant no. ZHY2025-141).
Not applicable.
XX, CL, HTZ, XG and GG analyzed the data and
constructed the figures. XG conceived and designed the study and
wrote and reviewed the manuscript. Data authentication is not
applicable. All authors have read and approved the final
manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Luft FC: Epigenetic ‘Transgenerational’
Inheritance. Circulation. 146:1096–1098. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Agudelo Garcia PA and Berger SL: Genetics
meets epigenetics in treg cells and autoimmunity. Immunity.
52:897–899. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Greally JM: A user's guide to the
ambiguous word 'epigenetics'. Nat Rev Mol Cell Biol. 19:207–208.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Waddington CH: Genetic assimilation of the
bithorax phenotype. Evolution. 10:1–13. 1956. View Article : Google Scholar
|
|
5
|
Huo S, Tang X, Chen W, Gan D, Guo H, Yao
Q, Liao R, Huang T, Wu J, Yang J, et al: Epigenetic regulations of
cellular senescence in osteoporosis. Ageing Res Rev. 99:1022352024.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mu S, Wang W, Liu Q, Ke N, Li H, Sun F,
Zhang J and Zhu Z: Autoimmune disease: A view of epigenetics and
therapeutic targeting. Front Immunol. 15:14827282024. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nadeu F, Diaz-Navarro A, Delgado J, Puente
XS and Campo E: Genomic and epigenomic alterations in chronic
lymphocytic leukemia. Annu Rev Pathol. 15:149–177. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xiong X, James BT, Boix CA, Park YP,
Galani K, Victor MB, Sun N, Hou L, Ho LL, Mantero J, et al:
Epigenomic dissection of Alzheimer's disease pinpoints causal
variants and reveals epigenome erosion. Cell. 186:4422–4437.e21.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sarode P, Pullamsetti SS and Savai R: New
insights into the epigenomic landscape of small-cell lung cancer: A
game changer? Am J Respir Crit Care Med. 206:1441–1443. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu C, Huang KK, Law JH, Chua JS, Sheng T,
Flores NM, Pizzi MP, Okabe A, Tan ALK, Zhu F, et al: Comprehensive
molecular phenotyping of ARID1A-deficient gastric cancer reveals
pervasive epigenomic reprogramming and therapeutic opportunities.
Gut. 72:1651–1663. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
van der Harst P, de Windt LJ and Chambers
JC: Translational perspective on epigenetics in cardiovascular
disease. J Am Coll Cardiol. 70:590–606. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lu C, Jain SU, Hoelper D, Bechet D, Molden
RC, Ran L, Murphy D, Venneti S, Hameed M, Pawel BR, et al: Histone
H3K36 mutations promote sarcomagenesis through altered histone
methylation landscape. Science. 352:844–849. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Roundtree IA, Evans ME, Pan T and He C:
Dynamic RNA modifications in gene expression regulation. Cell.
169:1187–1200. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cappannini A, Ray A, Purta E, Mukherjee S,
Boccaletto P, Moafinejad SN, Lechner A, Barchet C, Klaholz BP,
Stefaniak F and Bujnicki JM: MODOMICS: A database of RNA
modifications and related information. 2023 update. Nucleic Acids
Res. 52:D239–D244. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Galloway A and Cowling VH: mRNA cap
regulation in mammalian cell function and fate. Biochim Biophys
Acta Gene Regul Mech. 1862:270–279. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Raettig R, Kersten H, Weissenbach J and
Dirheimer G: Methylation of an adenosine in the D-loop of specific
transfer RNAs from yeast by a procaryotic tRNA (adenine-1)
methyltransferase. Nucleic Acids Res. 4:1769–1782. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Desrosiers R, Friderici K and Rottman F:
Identification of methylated nucleosides in messenger RNA from
Novikoff hepatoma cells. Proc Natl Acad Sci USA. 71:3971–3975.
1974. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Perry RP and Kelley DE: Kinetics of
formation of 5′terminal caps in mRNA. Cell. 8:433–442. 1976.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wyatt GR: Occurrence of 5-methylcytosine
in nucleic acids. Nature. 166:237–238. 1950. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu N, Dai Q, Zheng G, He C, Parisien M
and Pan T: N(6)-methyladenosine-dependent RNA structural switches
regulate RNA-protein interactions. Nature. 518:560–564. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rodan GA and Martin TJ: Therapeutic
approaches to bone diseases. Science. 289:1508–1514. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
GBD 2023 Disease and Injury Risk Factor
Collaborators, . Burden of 375 diseases and injuries,
risk-attributable burden of 88 risk factors, and healthy life
expectancy in 204 countries and territories, including 660
subnational locations, 1990–2023: A systematic analysis for the
Global Burden of Disease Study 2023. Lancet. 406:1873–1922. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen YS, Lian WS, Kuo CW, Ke HJ, Wang SY,
Kuo PC, Jahr H and Wang FS: Epigenetic regulation of skeletal
tissue integrity and osteoporosis development. Int J Mol Sci.
21:49232020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang Z, Wang M, Xie D, Huang Z, Zhang L,
Yang Y, Ma D, Li W, Zhou Q, Yang YG and Wang XJ: METTL3-mediated
N6-methyladenosine mRNA modification enhances long-term
memory consolidation. Cell Res. 28:1050–1061. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Han Z, Niu T, Chang J, Lei X, Zhao M, Wang
Q, Cheng W, Wang J, Feng Y and Chai J: Crystal structure of the FTO
protein reveals basis for its substrate specificity. Nature.
464:1205–1209. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Theler D, Dominguez C, Blatter M, Boudet J
and Allain FHT: Solution structure of the YTH domain in complex
with N6-methyladenosine RNA: A reader of methylated RNA. Nucleic
Acids Res. 42:13911–13919. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu J, Yue Y, Han D, Wang X, Fu Y, Zhang
L, Jia G, Yu M, Lu Z, Deng X, et al: A METTL3-METTL14 complex
mediates mammalian nuclear RNA N6-adenosine methylation. Nat Chem
Biol. 10:93–95. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ping XL, Sun BF, Wang L, Xiao W, Yang X,
Wang WJ, Adhikari S, Shi Y, Lv Y, Chen YS, et al: Mammalian WTAP is
a regulatory subunit of the RNA N6-methyladenosine
methyltransferase. Cell Res. 24:177–189. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang X, Zhao BS, Roundtree IA, Lu Z, Han
D, Ma H, Weng X, Chen K, Shi H and He C: N(6)-methyladenosine
modulates messenger RNA translation efficiency. Cell.
161:1388–1399. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shi H, Wang X, Lu Z, Zhao BS, Ma H, Hsu
PJ, Liu C and He C: YTHDF3 facilitates translation and decay of
N6-methyladenosine-modified RNA. Cell Res. 27:315–328.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hsu PJ, Zhu Y, Ma H, Guo Y, Shi X, Liu Y,
Qi M, Lu Z, Shi H, Wang J, et al: Ythdc2 is an
N6-methyladenosine binding protein that regulates
mammalian spermatogenesis. Cell Res. 27:1115–1127. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jia G, Fu Y, Zhao X, Dai Q, Zheng G, Yang
Y, Yi C, Lindahl T, Pan T, Yang YG and He C: N6-methyladenosine in
nuclear RNA is a major substrate of the obesity-associated FTO. Nat
Chem Biol. 7:885–887. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang Y, Hsu PJ, Chen YS and Yang YG:
Dynamic transcriptomic m(6)A decoration: Writers, erasers, readers
and functions in RNA metabolism. Cell Res. 28:616–624. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zheng G, Dahl JA, Niu Y, Fedorcsak P,
Huang CM, Li CJ, Vågbø CB, Shi Y, Wang WL, Song SH, et al: ALKBH5
is a mammalian RNA demethylase that impacts RNA metabolism and
mouse fertility. Molecular Cell. 49:18–29. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Boulias K and Greer EL: Biological roles
of adenine methylation in RNA. Nat Rev Genet. 24:143–160. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gao Z, Zha X, Li M, Xia X and Wang S:
Insights into the m6A demethylases FTO and ALKBH5: Structural,
biological function, and inhibitor development. Cell Biosci.
14:1082024. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bokar JA, Shambaugh ME, Polayes D, Matera
AG and Rottman FM: Purification and cDNA cloning of the
AdoMet-binding subunit of the human mRNA
(N6-adenosine)-methyltransferase. RNA. 3:1233–1247. 1997.PubMed/NCBI
|
|
38
|
Wang X, Feng J, Xue Y, Guan Z, Zhang D,
Liu Z, Gong Z, Wang Q, Huang J, Tang C, et al: Corrigendum:
Structural basis of N6-adenosine methylation by the
METTL3-METTL14 complex. Nature. 542:2602017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Huang J, Dong X, Gong Z, Qin LY, Yang S,
Zhu YL, Wang X, Zhang D, Zou T, Yin P and Tang C: Solution
structure of the RNA recognition domain of METTL3-METTL14
N6-methyladenosine methyltransferase. Protein Cell.
10:272–284. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Su S, Li S, Deng T, Gao M, Yin Y, Wu B,
Peng C, Liu J, Ma J and Zhang K: Cryo-EM structures of human
m6A writer complexes. Cell Res. 32:982–994. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang P, Doxtader KA and Nam Y: Structural
basis for cooperative function of Mettl3 and Mettl14
Methyltransferases. Mol Cell. 63:306–317. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Horiuchi K, Umetani M, Minami T, Okayama
H, Takada S, Yamamoto M, Aburatani H, Reid PC, Housman DE, Hamakubo
T and Kodama T: Wilms' tumor 1-associating protein regulates G2/M
transition through stabilization of cyclin A2 mRNA. Proc Natl Acad
Sci USA. 103:17278–17283. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tang J, Wang F, Cheng G, Si S, Sun X, Han
J, Yu H, Zhang W, Lv Q, Wei JF and Yang H: Wilms' tumor
1-associating protein promotes renal cell carcinoma proliferation
by regulating CDK2 mRNA stability. J Exp Clin Cancer Res.
37:402018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Huang Q, Mo J, Liao Z, Chen X and Zhang B:
The RNA m6A writer WTAP in diseases: Structure, roles, and
mechanisms. Cell Death Dis. 13:8522022. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sorci M, Ianniello Z, Cruciani S, Larivera
S, Ginistrelli LC, Capuano E, Marchioni M, Fazi F and Fatica A:
METTL3 regulates WTAP protein homeostasis. Cell Death Dis.
9:7962018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Schöller E, Weichmann F, Treiber T, Ringle
S, Treiber N, Flatley A, Feederle R, Bruckmann A and Meister G:
Interactions, localization, and phosphorylation of the m6A
generating METTL3-METTL14-WTAP complex. RNA. 24:499–512. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Spector DL and Lamond AI: Nuclear
speckles. Cold Spring Harb Perspect Biol. 3:a0006462011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lamond AI and Spector DL: Nuclear
speckles: A model for nuclear organelles. Nat Rev Mol Cell Biol.
4:605–612. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Bhat P, Chow A, Emert B, Ettlin O,
Quinodoz SA, Strehle M, Takei Y, Burr A, Goronzy IN, Chen AW, et
al: Genome organization around nuclear speckles drives mRNA
splicing efficiency. Nature. 629:1165–1173. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Faber GP, Nadav-Eliyahu S and Shav-Tal Y:
Nuclear speckles-a driving force in gene expression. J Cell Sci.
135:jcs2595942022. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Meyer KD, Saletore Y, Zumbo P, Elemento O,
Mason CE and Jaffrey SR: Comprehensive analysis of mRNA methylation
reveals enrichment in 3′UTRs and near stop codons. Cell.
149:1635–1646. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Dominissini D, Moshitch-Moshkovitz S,
Schwartz S, Salmon-Divon M, Ungar L, Osenberg S, Cesarkas K,
Jacob-Hirsch J, Amariglio N, Kupiec M, et al: Topology of the human
and mouse m6A RNA methylomes revealed by m6A-seq. Nature.
485:201–206. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ke S, Alemu EA, Mertens C, Gantman EC, Fak
JJ, Mele A, Haripal B, Zucker-Scharff I, Moore MJ, Park CY, et al:
A majority of m6A residues are in the last exons, allowing the
potential for 3′UTR regulation. Genes Dev. 29:2037–2053. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhang H, Shi X, Huang T, Zhao X, Chen W,
Gu N and Zhang R: Dynamic landscape and evolution of m6A
methylation in human. Nucleic Acids Res. 48:6251–6264. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Meyer KD, Patil DP, Zhou J, Zinoviev A,
Skabkin MA, Elemento O, Pestova TV, Qian SB and Jaffrey SR: 5′UTR
m(6)A promotes cap-independent translation. Cell. 163:999–1010.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhou Y, Ćorović M, Hoch-Kraft P, Meiser N,
Mesitov M, Körtel N, Back H, Naarmann-de Vries IS, Katti K, Obrdlík
A, et al: m6A sites in the coding region trigger
translation-dependent mRNA decay. Mol Cell. 84:4576–4593.e12. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Ćorović M, Hoch-Kraft P, Zhou Y, Hallstein
S, König J and Zarnack K: m6A in the coding sequence: Linking
deposition, translation, and decay. Trends Genet. 41:963–973. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Xu C, Wang X, Liu K, Roundtree IA, Tempel
W, Li Y, Lu Z, He C and Min J: Structural basis for selective
binding of m6A RNA by the YTHDC1 YTH domain. Nat Chem Biol.
10:927–929. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Luo S and Tong L: Molecular basis for the
recognition of methylated adenines in RNA by the eukaryotic YTH
domain. Proc Natl Acad Sci USA. 111:13834–13839. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Liu T, Wei Q, Jin J, Luo Q, Liu Y, Yang Y,
Cheng C, Li L, Pi J, Si Y, et al: The m6A reader YTHDF1 promotes
ovarian cancer progression via augmenting EIF3C translation.
Nucleic Acids Res. 48:3816–3831. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zong X, Xiao X, Shen B, Jiang Q, Wang H,
Lu Z, Wang F, Jin M, Min J, Wang F, et al: The N6-methyladenosine
RNA-binding protein YTHDF1 modulates the translation of TRAF6 to
mediate the intestinal immune response. Nucleic Acids Res.
49:5537–5552. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wang X, Lu Z, Gomez A, Hon GC, Yue Y, Han
D, Fu Y, Parisien M, Dai Q, Jia G, et al:
N6-methyladenosine-dependent regulation of messenger RNA stability.
Nature. 505:117–120. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Li A, Chen YS, Ping XL, Yang X, Xiao W,
Yang Y, Sun HY, Zhu Q, Baidya P, Wang X, et al: Cytoplasmic m6A
reader YTHDF3 promotes mRNA translation. Cell Res. 27:444–447.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Chen Y, Wan R, Zou Z, Lao L, Shao G, Zheng
Y, Tang L, Yuan Y, Ge Y, He C and Lin S: O-GlcNAcylation determines
the translational regulation and phase separation of YTHDF
proteins. Nat Cell Biol. 25:1676–1690. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Mannino MP and Hart GW: The Beginner's
guide to O-GlcNAc: From nutrient sensitive pathway regulation to
its impact on the immune system. Front Immunol. 13:8286482022.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Zou Z, Sepich-Poore C, Zhou X, Wei J and
He C: The mechanism underlying redundant functions of the YTHDF
proteins. Genome Biol. 24:172023. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Du H, Zhao Y, He J, Zhang Y, Xi H, Liu M,
Ma J and Wu L: YTHDF2 destabilizes m(6)A-containing RNA through
direct recruitment of the CCR4-NOT deadenylase complex. Nat Commun.
7:126262016. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zou Z and He C: The YTHDF proteins display
distinct cellular functions on m6A-modified RNA. Trends
Biochem Sci. 49:611–621. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Roundtree IA, Luo GZ, Zhang Z, Wang X,
Zhou T, Cui Y, Sha J, Huang X, Guerrero L, Xie P, et al: YTHDC1
mediates nuclear export of N6-methyladenosine methylated
mRNAs. Elife. 6:e313112017. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Abby E, Tourpin S, Ribeiro J, Daniel K,
Messiaen S, Moison D, Guerquin J, Gaillard JC, Armengaud J, Langa
F, et al: Implementation of meiosis prophase I programme requires a
conserved retinoid-independent stabilizer of meiotic transcripts.
Nat Commun. 7:103242016. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Wojtas MN, Pandey RR, Mendel M, Homolka D,
Sachidanandam R and Pillai RS: Regulation of m6A
Transcripts by the 3′→5′ RNA helicase YTHDC2 is essential for a
successful meiotic program in the mammalian germline. Mol Cell.
68:374–387.e12. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Gerken T, Girard CA, Tung YC, Webby CJ,
Saudek V, Hewitson KS, Yeo GS, McDonough MA, Cunliffe S, McNeill
LA, et al: The obesity-associated FTO gene encodes a
2-oxoglutarate-dependent nucleic acid demethylase. Science.
318:1469–1472. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Jia G, Fu Y and He C: Reversible RNA
adenosine methylation in biological regulation. Trends Genet.
29:108–115. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Fu Y, Jia G, Pang X, Wang RN, Wang X, Li
CJ, Smemo S, Dai Q, Bailey KA, Nobrega MA, et al: FTO-mediated
formation of N6-hydroxymethyladenosine and N6-formyladenosine in
mammalian RNA. Nat Commun. 4:17982013. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Toh JDW, Crossley SWM, Bruemmer KJ, Ge EJ,
He D, Iovan DA and Chang CJ: Distinct RNA N-demethylation pathways
catalyzed by nonheme iron ALKBH5 and FTO enzymes enable regulation
of formaldehyde release rates. Proc Natl Acad Sci USA.
117:25284–25292. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Wei J, Liu F, Lu Z, Fei Q, Ai Y, He PC,
Shi H, Cui X, Su R, Klungland A, et al: Differential
m6A, m6Am, and m1A
Demethylation mediated by FTO in the cell nucleus and cytoplasm.
Mol Cell. 71:973–985.e5. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Yu F, Zhu AC, Liu S, Gao B, Wang Y,
Khudaverdyan N, Yu C, Wu Q, Jiang Y, Song J, et al: RBM33 is a
unique m6A RNA-binding protein that regulates ALKBH5
demethylase activity and substrate selectivity. Mol Cell.
83:2003–2019.e6. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Bai L, Xiang Y, Tang M, Liu S, Chen Q,
Chen Q, Zhang M, Wan S, Sang Y, Li Q, et al: ALKBH5 controls the
meiosis-coupled mRNA clearance in oocytes by removing the N
6-methyladenosine methylation. Nat Commun. 14:65322023. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Shah AM and Giacca M: Small non-coding RNA
therapeutics for cardiovascular disease. Eur Heart J. 43:4548–4561.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Yates LA, Norbury CJ and Gilbert RJC: The
long and short of microRNA. Cell. 153:516–519. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Alarcón CR, Lee H, Goodarzi H, Halberg N
and Tavazoie SF: N6-methyladenosine marks primary microRNAs for
processing. Nature. 519:482–485. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Alarcón CR, Goodarzi H, Lee H, Liu X,
Tavazoie S and Tavazoie SF: HNRNPA2B1 is a mediator of
m(6)A-dependent nuclear RNA processing events. Cell. 162:1299–1308.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Chen Z, Chen X, Lei T, Gu Y, Gu J, Huang
J, Lu B, Yuan L, Sun M and Wang Z: Integrative analysis of NSCLC
identifies LINC01234 as an oncogenic lncRNA that interacts with
HNRNPA2B1 and regulates miR-106b biogenesis. Mol Ther.
28:1479–1493. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Sun M, Shen Y, Jia G, Deng Z, Shi F, Jing
Y and Xia S: Activation of the HNRNPA2B1/miR-93-5p/FRMD6 axis
facilitates prostate cancer progression in an m6A-dependent manner.
J Cancer. 14:1242–1256. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Chen T, Hao YJ, Zhang Y, Li MM, Wang M,
Han W, Wu Y, Lv Y, Hao J, Wang L, et al: m(6)A RNA methylation is
regulated by microRNAs and promotes reprogramming to pluripotency.
Cell Stem Cell. 16:289–301. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Hwang HW, Wentzel EA and Mendell JT: A
hexanucleotide element directs microRNA nuclear import. Science.
315:97–100. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Zhang Z and Wang XJ:
N6-methyladenosine mRNA modification: From modification
site selectivity to neurological functions. Acc Chem Res.
56:2992–2999. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Zhang Z, Zhou K, Han L, Small A, Xue J,
Huang H, Weng H, Su R, Tan B, Shen C, et al: RNA m6A
reader YTHDF2 facilitates precursor miR-126 maturation to promote
acute myeloid leukemia progression. Genes Dis. 11:382–396. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Kopp F and Mendell JT: Functional
classification and experimental dissection of long noncoding RNAs.
Cell. 172:393–407. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Patil DP, Chen CK, Pickering BF, Chow A,
Jackson C, Guttman M and Jaffrey SR: m(6)A RNA methylation promotes
XIST-mediated transcriptional repression. Nature. 537:369–373.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Yang X, Zhang S, He C, Xue P, Zhang L, He
Z, Zang L, Feng B, Sun J and Zheng M: METTL14 suppresses
proliferation and metastasis of colorectal cancer by
down-regulating oncogenic long non-coding RNA XIST. Mol Cancer.
19:462020. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Shi B, Liu WW, Yang K, Jiang GM and Wang
H: The role, mechanism, and application of RNA methyltransferase
METTL14 in gastrointestinal cancer. Mol Cancer. 21:1632022.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
He AT, Liu J, Li F and Yang BB: Targeting
circular RNAs as a therapeutic approach: Current strategies and
challenges. Signal Transduct Target Ther. 6:1852021. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Yang Y, Fan X, Mao M, Song X, Wu P, Zhang
Y, Jin Y, Yang Y, Chen LL, Wang Y, et al: Extensive translation of
circular RNAs driven by N6-methyladenosine. Cell Res.
27:626–641. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Zhang L, Hou C, Chen C, Guo Y, Yuan W, Yin
D, Liu J and Sun Z: The role of N6-methyladenosine
(m6A) modification in the regulation of circRNAs. Mol
Cancer. 19:1052020. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Ahi EP: Regulation of Skeletogenic
Pathways by m6A RNA Modification: A comprehensive review. Calcif
Tissue Int. 116:582025. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Gu Y, Song Y, Pan Y and Liu J: The
essential roles of m6A modification in osteogenesis and common bone
diseases. Genes Dis. 11:335–345. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Su N, Yang J, Xie Y, Du X, Chen H, Zhou H
and Chen L: Bone function, dysfunction and its role in diseases
including critical illness. Int J Biol Sci. 15:776–787. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Galea GL, Zein MR, Allen S and
Francis-West P: Making and shaping endochondral and intramembranous
bones. Dev Dyn. 250:414–449. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Long F and Ornitz DM: Development of the
endochondral skeleton. Cold Spring Harb Perspect Biol.
5:a0083342013. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Kim JM, Lin C, Stavre Z, Greenblatt MB and
Shim JH: Osteoblast-osteoclast communication and bone homeostasis.
Cells. 9:20732020. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Long F: Building strong bones: Molecular
regulation of the osteoblast lineage. Nat Rev Mol Cell Biol.
13:27–38. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Salhotra A, Shah HN, Levi B and Longaker
MT: Mechanisms of bone development and repair. Nat Rev Mol Cell
Biol. 21:696–711. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Nakashima K and de Crombrugghe B:
Transcriptional mechanisms in osteoblast differentiation and bone
formation. Trends Genet. 19:458–466. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Ducy P, Zhang R, Geoffroy V, Ridall AL and
Karsenty G: Osf2/Cbfa1: A transcriptional activator of osteoblast
differentiation. Cell. 89:747–754. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Komori T, Yagi H, Nomura S, Yamaguchi A,
Sasaki K, Deguchi K, Shimizu Y, Bronson RT, Gao YH and Inada M:
Targeted disruption of Cbfa1 results in a complete lack of bone
formation owing to maturational arrest of osteoblasts. Cell.
89:755–764. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Choi KY, Lee SW, Park MH, Bae YC, Shin HI,
Nam S, Kim YJ, Kim HJ and Ryoo HM: Spatio-temporal expression
patterns of Runx2 isoforms in early skeletogenesis. Exp Mol Med.
34:426–433. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Yoshida CA, Furuichi T, Fujita T, Fukuyama
R, Kanatani N, Kobayashi S, Satake M, Takada K and Komori T:
Core-binding factor beta interacts with Runx2 and is required for
skeletal development. Nat Genet. 32:633–638. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Chen W, Ma J, Zhu G, Jules J, Wu M,
McConnell M, Tian F, Paulson C, Zhou X, Wang L and Li YP: Cbfβ
deletion in mice recapitulates cleidocranial dysplasia and reveals
multiple functions of Cbfβ required for skeletal development. Proc
Natl Acad Sci USA. 111:8482–8487. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Kanatani N, Fujita T, Fukuyama R, Liu W,
Yoshida CA, Moriishi T, Yamana K, Miyazaki T, Toyosawa S and Komori
T: Cbfβ regulates Runx2 function isoform-dependently in postnatal
bone development. Dev Biol. 296:48–61. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Qin X, Jiang Q, Matsuo Y, Kawane T, Komori
H, Moriishi T, Taniuchi I, Ito K, Kawai Y, Rokutanda S, et al: Cbfb
regulates bone development by stabilizing Runx family proteins. J
Bone Miner Res. 30:706–714. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Yang L, Ren Z, Yan S, Zhao L, Liu J, Zhao
L, Li Z, Ye S, Liu A, Li X, et al: Nsun4 and Mettl3 mediated
translational reprogramming of Sox9 promotes BMSC chondrogenic
differentiation. Commun Biol. 5:4952022. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Yao Y, Bi Z, Wu R, Zhao Y, Liu Y, Liu Q,
Wang Y and Wang X: METTL3 inhibits BMSC adipogenic differentiation
by targeting the JAK1/STAT5/C/EBPβ pathway via an
m6A-YTHDF2-dependent manner. FASEB J. 33:7529–7544. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Wu Y, Xie L, Wang M, Xiong Q, Guo Y, Liang
Y, Li J, Sheng R, Deng P, Wang Y, et al: Mettl3-mediated m6A RNA
methylation regulates the fate of bone marrow mesenchymal stem
cells and osteoporosis. Nat Commun. 9:47722018. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Yan G, Yuan Y, He M, Gong R, Lei H, Zhou
H, Wang W, Du W, Ma T, Liu S, et al: m6A Methylation of
Precursor-miR-320/RUNX2 controls osteogenic potential of bone
marrow-derived mesenchymal stem cells. Mol Ther Nucleic Acids.
19:421–436. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Sebo ZL, Rendina-Ruedy E, Ables GP,
Lindskog DM, Rodeheffer MS, Fazeli PK and Horowitz MC: Bone marrow
adiposity: Basic and clinical implications. Endocr Rev.
40:1187–1206. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Wu T, Tang H, Yang J, Yao Z, Bai L, Xie Y,
Li Q and Xiao J: METTL3-m6 A methylase regulates the
osteogenic potential of bone marrow mesenchymal stem cells in
osteoporotic rats via the Wnt signalling pathway. Cell Prolif.
55:e132342022. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Tian C, Huang Y, Li Q, Feng Z and Xu Q:
Mettl3 Regulates osteogenic differentiation and alternative
splicing of vegfa in bone marrow mesenchymal stem cells. Int J Mol
Sci. 20:5512019. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Cai GP, Liu YL, Luo LP, Xiao Y, Jiang TJ,
Yuan J and Wang M: Alkbh1-mediated DNA N6-methyladenine
modification regulates bone marrow mesenchymal stem cell fate
during skeletal aging. Cell Prolif. 55:e131782022. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Kong Y, Zhang Y, Cai Y, Li D, Yi B and Xu
Q: METTL3 mediates osteoblast apoptosis by regulating endoplasmic
reticulum stress during LPS-induced inflammation. Cell Signal.
95:1103352022. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Wang Y, Chen Y, Xiao H, Liu Z, Liu X, Feng
Z, Sheng X, Peng B, Ren X, Xu L, et al: METTL3-mediated m6A
modification increases Hspa1a stability to inhibit osteoblast
aging. Cell Death Discov. 10:1552024. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Chen Y, Wang Y, Xiao H, Teng F, Yang A,
Liu J, Liu Z, Sheng X, Zhang C, Zhang S, et al: METTL3-mediated
modification of SIRT1 m6A methylation protects osteoblasts from
TBHP-induced senescence and promotes osteoblast proliferation. Chem
Biol Interact. 420:1116722025. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Zhou D, Ran Y, Yu R, Liu G, Ran D and Liu
Z: SIRT1 regulates osteoblast senescence through SOD2 acetylation
and mitochondrial dysfunction in the progression of Osteoporosis
caused by Cadmium exposure. Chem Biol Interact. 382:1106322023.
View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Zhang Y, Gu X, Li D, Cai L and Xu Q:
METTL3 regulates osteoblast differentiation and inflammatory
response via smad signaling and MAPK signaling. Int J Mol Sci.
21:1992019. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Wang Y, Yu X, Sun F, Fu Y, Hu T, Shi Q and
Man Q: METTL14 Mediates Glut3 m6A methylation to improve
osteogenesis under oxidative stress condition. Redox Rep.
30:24352412024. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Wang P, Zhou W, Chen F, Zhang X, Zhang Q,
Chen Y and Zhang N: METTL14-mediated methylation of SLC25A3
mitigates mitochondrial damage in osteoblasts, leading to the
improvement of osteoporosis. Exp Gerontol. 194:1124962024.
View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Zhang Q, Riddle RC, Yang Q, Rosen CR,
Guttridge DC, Dirckx N, Faugere MC, Farber CR and Clemens TL: The
RNA demethylase FTO is required for maintenance of bone mass and
functions to protect osteoblasts from genotoxic damage. Proc Natl
Acad Sci USA. 116:17980–17989. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Okamoto K, Nakashima T, Shinohara M,
Negishi-Koga T, Komatsu N, Terashima A, Sawa S, Nitta T and
Takayanagi H: Osteoimmunology: The conceptual framework unifying
the immune and skeletal systems. Physiol Rev. 97:1295–1349. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Li D, Cai L, Meng R, Feng Z and Xu Q:
METTL3 modulates osteoclast differentiation and function by
controlling RNA stability and nuclear export. Int J Mol Sci.
21:16602020. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Li D, He J, Fang C, Zhang Y, He M, Zhang
Z, Hou J and Xu Q: METTL3 regulates osteoclast biological behaviors
via iNOS/NO-mediated mitochondrial dysfunction in inflammatory
conditions. Int J Mol Sci. 24:14032023. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Wang W, Qiao SC, Wu XB, Sun B, Yang JG, Li
X, Zhang X, Qian SJ, Gu YX and Lai HC: Circ_0008542 in osteoblast
exosomes promotes osteoclast-induced bone resorption through m6A
methylation. Cell Death Dis. 12:6282021. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Liang J, Yi Q, Liu Y, Li J, Yang Z and Sun
W and Sun W: Recent advances of m6A methylation in skeletal system
disease. J Transl Med. 22:1532024. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Wu S, Li XF, Wu YY, Yin SQ, Huang C and Li
J: N6-Methyladenosine and rheumatoid arthritis: A comprehensive
review. Front Immunol. 12:7318422021. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Liu B, Song G, Wang Y, Song C, Cao Y, Tong
J, Wang Y, Fan X, Shi N, Zhao H and Fan D: N6-methyladenosine and
intervertebral disc degeneration: Advances in detection and
pathological insights. J Orthop Translat. 53:38–51. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Clynes MA, Harvey NC, Curtis EM, Fuggle
NR, Dennison EM and Cooper C: The epidemiology of osteoporosis. Br
Med Bull. 133:105–117. 2020.PubMed/NCBI
|
|
136
|
Okolo S, Ginsburg J and Hardiman P:
Prevention and treatment of osteoporosis. Lancet. 341:13491993.
View Article : Google Scholar : PubMed/NCBI
|
|
137
|
US Preventive Services Task Force, .
Nicholson WK, Silverstein M, Wong JB, Chelmow D, Coker TR, Davis
EM, Jaén CR, Krousel-Wood M, Lee S, et al: Screening for
osteoporosis to prevent fractures: US preventive services task
force recommendation statement. JAMA. 333:498–508. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Li J, Chen X, Lu L and Yu X: The
relationship between bone marrow adipose tissue and bone metabolism
in postmenopausal osteoporosis. Cytokine Growth Factor Rev.
52:88–98. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Cotts KG and Cifu AS: Treatment of
osteoporosis. JAMA. 319:1040–1041. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Vis M, Dijkmans BAC and Lems WF:
Bisphosphonates for osteoporosis. N Engl J Med. 364:2011.PubMed/NCBI
|
|
141
|
Khosla S and Hofbauer LC: Osteoporosis
treatment: Recent developments and ongoing challenges. Lancet
Diabetes Endocrinol. 5:898–907. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Meunier PJ, Roux C, Seeman E, Ortolani S,
Badurski JE, Spector TD, Cannata J, Balogh A, Lemmel EM and
Pors-Nielsen S: The effects of strontium ranelate on the risk of
vertebral fracture in women with postmenopausal osteoporosis. New
Engl J Med. 350:459–468. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Chen X, Hua W, Huang X, Chen Y, Zhang J
and Li G: Regulatory Role of RNA N6-Methyladenosine modification in
bone biology and osteoporosis. Front Endocrinol (Lausanne).
10:9112020. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Vimalraj S, Arumugam B, Miranda PJ and
Selvamurugan N: Runx2: Structure, function, and phosphorylation in
osteoblast differentiation. Int J Biol Macromol. 78:202–208. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Yan G, Yuan Y, He M, Gong R, Lei H, Zhou
H, Wang W, Du W, Ma T, Liu S, et al: m6A methylation of
Precursor-miR-320/RUNX2 controls osteogenic potential of bone
Marrow-derived mesenchymal stem cells. Mol Ther Nucleic Acids.
19:421–436. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Hamam D, Ali D, Vishnubalaji R, Hamam R,
Al-Nbaheen M, Chen L, Kassem M, Aldahmash A and Alajez NM:
microRNA-320/RUNX2 axis regulates adipocytic differentiation of
human mesenchymal (skeletal) stem cells. Cell Death Dis.
5:e14992014. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Peng J, Zhan Y and Zong Y: METTL3-mediated
LINC00657 promotes osteogenic differentiation of mesenchymal stem
cells via miR-144-3p/BMPR1B axis. Cell Tissue Res. 388:301–312.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Huang C and Wang Y: Downregulation of
METTL14 improves postmenopausal osteoporosis via IGF2BP1 dependent
posttranscriptional silencing of SMAD1. Cell Death Dis. 13:9192022.
View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Théry C, Witwer KW, Aikawa E, Alcaraz MJ,
Anderson JD, Andriantsitohaina R, Antoniou A, Arab T, Archer F and
Atkin-Smith GK: Minimal information for studies of extracellular
vesicles 2018 (MISEV2018): A position statement of the
international society for extracellular vesicles and update of the
MISEV2014 guidelines. J Extracell Vesicles. 7:15357502018.
View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Kalluri R and LeBleu VS: The biology,
function, and biomedical applications of exosomes. Science.
367:eaau69772020. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Yang JG, Sun B, Wang Z, Li X, Gao JH, Qian
JJ, Li J, Wei WJ, Zhang P and Wang W: Exosome-targeted delivery of
METTL14 regulates NFATc1 m6A methylation levels to correct
osteoclast-induced bone resorption. Cell Death Dis. 14:7382023.
View Article : Google Scholar : PubMed/NCBI
|
|
152
|
He M, Lei H, He X, Liu Y, Wang A, Ren Z,
Liu X, Yan G, Wang W, Wang Y, et al: METTL14 regulates osteogenesis
of bone marrow mesenchymal stem cells via inducing autophagy
through m6A/IGF2BPs/Beclin-1 signal axis. Stem Cells Transl Med.
11:987–1001. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Fang C, He M, Li D and Xu Q: YTHDF2
mediates LPS-induced osteoclastogenesis and inflammatory response
via the NF-κB and MAPK signaling pathways. Cell Signal.
85:1100602021. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Mizushima N and Levine B: Autophagy in
human diseases. N Engl J Med. 383:1564–1576. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Yin X, Zhou C, Li J, Liu R, Shi B, Yuan Q
and Zou S: Autophagy in bone homeostasis and the onset of
osteoporosis. Bone Res. 7:282019. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Wang CG, Hu YH, Su SL and Zhong D: LncRNA
DANCR and miR-320a suppressed osteogenic differentiation in
osteoporosis by directly inhibiting the Wnt/β-catenin signaling
pathway. Exp Mol Med. 52:1310–1325. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Wang X, Zou C, Li M, Hou C, Jiang W, Bian
Z and Zhu L: METTL14 upregulates TCF1 through m6A mRNA methylation
to stimulate osteogenic activity in osteoporosis. Hum Cell.
36:178–194. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Sun W, Qiao W, Zhou B, Hu Z, Yan Q, Wu J,
Wang R, Zhang Q and Miao D: Overexpression of Sirt1 in mesenchymal
stem cells protects against bone loss in mice by FOXO3a
deacetylation and oxidative stress inhibition. Metabolism.
88:61–71. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Wang C, Chen R, Zhu X, Zhang X and Lian N:
METTL14 alleviates the development of osteoporosis in
ovariectomized mice by upregulating m6A level of SIRT1 mRNA. Bone.
168:1166522023. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
You Y, Liu J, Zhang L, Li X, Sun Z, Dai Z,
Ma J, Jiao G and Chen Y: WTAP-mediated m6A modification modulates
bone marrow mesenchymal stem cells differentiation potential and
osteoporosis. Cell Death Dis. 14:332023. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Liu J, You Y, Sun Z, Zhang L, Li X, Dai Z,
Ma J, Chen Y and Jiao G: WTAP-Mediated m6A RNA methylation
regulates the differentiation of bone marrow mesenchymal stem cells
via the miR-29b-3p/HDAC4 Axis. Stem Cells Transl Med. 12:307–321.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Zhang P, Chen H, Xie B, Zhao W, Shang Q,
He J, Shen G, Yu X, Zhang Z, Zhu G, et al: Bioinformatics
identification and experimental validation of m6A-related
diagnostic biomarkers in the subtype classification of blood
monocytes from postmenopausal osteoporosis patients. Front
Endocrinol (Lausanne). 14:9900782023. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
GBD 2021 Osteoarthritis Collaborators, .
Global, regional, and national burden of osteoarthritis, 1990–2020
and projections to 2050: A systematic analysis for the Global
Burden of Disease Study 2021. Lancet Rheumatol. 5:e508–e522. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Wang Z, Yan K, Ge G, Zhang D, Bai J, Guo
X, Zhou J, Xu T, Xu M, Long X, et al: Exosomes derived from
miR-155-5p-overexpressing synovial mesenchymal stem cells prevent
osteoarthritis via enhancing proliferation and migration,
attenuating apoptosis, and modulating extracellular matrix
secretion in chondrocytes. Cell Biol Toxicol. 37:85–96. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Arendt-Nielsen L, Egsgaard LL and Petersen
KK: Evidence for a central mode of action for etoricoxib (COX-2
inhibitor) in patients with painful knee osteoarthritis. Pain.
157:1634–1644. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Zhong Y, Zhou Y, Ding R, Zou L, Zhang H,
Wei X and He D: Intra-articular treatment of temporomandibular
joint osteoarthritis by injecting actively-loaded meloxicam
liposomes with dual-functions of anti-inflammation and lubrication.
Mater Today Bio. 19:1005732023. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
da Costa BR, Pereira TV, Saadat P,
Rudnicki M, Iskander SM, Bodmer NS, Bobos P, Gao L, Kiyomoto HD and
Montezuma T: Effectiveness and safety of non-steroidal
anti-inflammatory drugs and opioid treatment for knee and hip
osteoarthritis: Network meta-analysis. BMJ. 375:n23212021.
View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Tang Sa, Zhang C, Oo WM, Fu K, Risberg MA,
Bierma-Zeinstra SM, Neogi T, Atukorala I, Malfait AM, Ding C and
Hunter DJ: Osteoarthritis. Nat Rev Dis Primers. 11:102025.
View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Lee YT, Mohd Yunus MH, Yazid MD and
Ugusman A: Unraveling the path to osteoarthritis management:
Targeting chondrocyte apoptosis for therapeutic intervention. Front
Cell Dev Biol. 12:13471262024. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Radbakhsh S, Najar M, Merimi M, Benderdour
M, Fernandes JC, Martel-Pelletier J, Pelletier JP and Fahmi H: RNA
Modifications in osteoarthritis: Epitranscriptomic insights into
pathogenesis and therapeutic targets. Int J Mol Sci. 26:49552025.
View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Chen X, Gong W, Shao X, Shi T, Zhang L,
Dong J, Shi Y, Shen S, Qin J, Jiang Q and Guo B: METTL3-mediated
m6A modification of ATG7 regulates autophagy-GATA4 axis
to promote cellular senescence and osteoarthritis progression. Ann
Rheum Dis. 81:87–99. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Damerau A, Rosenow E, Alkhoury D,
Buttgereit F and Gaber T: Fibrotic pathways and fibroblast-like
synoviocyte phenotypes in osteoarthritis. Front Immunol.
15:13850062024. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Shibakawa A, Aoki H, Masuko-Hongo K, Kato
T, Tanaka M, Nishioka K and Nakamura H: Presence of Pannus-like
tissue on osteoarthritic cartilage and its histological character.
Osteoarthritis Cartilage. 11:133–140. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Zhao W, Zhang C, Shi M, Zhang J, Li M, Xue
X, Zhang Z, Shu Z, Zhu J, Mu N, et al: The discoidin domain
receptor 2/annexin A2/matrix metalloproteinase 13 loop promotes
joint destruction in arthritis through promoting migration and
invasion of fibroblast-like synoviocytes. Arthritis Rheumatol.
66:2355–2367. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Kono M, Yasuda S, Stevens RL, Koide H,
Kurita T, Shimizu Y, Kanetsuka Y, Oku K, Bohgaki T, Amengual O, et
al: Ras guanine nucleotide-releasing protein 4 is aberrantly
expressed in the fibroblast-like synoviocytes of patients with
rheumatoid arthritis and controls their proliferation. Arthritis
Rheumatol. 67:396–407. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Lotz MK and Caramés B: Autophagy and
cartilage homeostasis mechanisms in joint health, aging and OA. Nat
Rev Rheumatol. 7:579–587. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Zhang Y, Vasheghani F, Li YH, Blati M,
Simeone K, Fahmi H, Lussier B, Roughley P, Lagares D, Pelletier JP,
et al: Cartilage-specific deletion of mTOR upregulates autophagy
and protects mice from osteoarthritis. Ann Rheum Dis. 74:1432–1440.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Liu Q, Li M, Jiang L, Jiang R and Fu B:
METTL3 promotes experimental osteoarthritis development by
regulating inflammatory response and apoptosis in chondrocyte.
Biochem Biophys Res Commun. 516:22–27. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Zhou H, Xie Z, Qian Y, Ni W, Cui L, Fang
X, Wan S, Zhao X, Qin A, Fan S and Wu Y: FTO-mediated SMAD2 m6A
modification protects cartilage against Osteoarthritis. Exp Mol
Med. 56:2283–2295. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Deng G, Xu Y, Li Z and Zeng G: WTAP
mediates IL-1β-induced chondrocyte injury by enhancing CA12 mRNA
stability depending on m6A modification. J Orthop Surg Res.
19:8262024. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Tang Y, Hong F, Ding S, Yang J, Zhang M,
Ma Y, Zheng Q, Yang D, Jin Y and Ma C: METTL3-mediated
m6A modification of IGFBP7-OT promotes osteoarthritis
progression by regulating the DNMT1/DNMT3a-IGFBP7 axis. Cell Rep.
42:1125892023. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Ren J, Li Y, Wuermanbieke S, Hu S and
Huang G: N6-methyladenosine (m6A) methyltransferase METTL3-mediated
LINC00680 accelerates osteoarthritis through m6A/SIRT1 manner. Cell
Death Discov. 8:2402022. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Zhou H, Shen X, Yan C, Xiong W, Ma Z, Tan
Z, Wang J, Li Y, Liu J, Duan A and Liu F: Extracellular vesicles
derived from human umbilical cord mesenchymal stem cells alleviate
osteoarthritis of the knee in mice model by interacting with METTL3
to reduce m6A of NLRP3 in macrophage. Stem Cell Res Ther.
13:3222022. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Lin Z, Jiang T, Zheng W, Zhang J, Li A, Lu
C and Liu W: N6-methyladenosine (m6A) methyltransferase
WTAP-mediated miR-92b-5p accelerates osteoarthritis progression.
Cell Commun Signal. 21:1992023. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Cai D, Zhang J, Yang J, Lv Q and Zhong C:
Overexpression of FTO alleviates osteoarthritis by regulating the
processing of miR-515-5p and the TLR4/MyD88/NF-κB axis. Int
Immunopharmacol. 114:1095242022. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Smolen JS, Aletaha D and McInnes IB:
Rheumatoid arthritis. Lancet. 388:2023–2038. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Curran AM, Girgis AA, Jang Y, Crawford JD,
Thomas MA, Kawalerski R, Coller J, Bingham CO III, Na CH and Darrah
E: Citrullination modulates antigen processing and presentation by
revealing cryptic epitopes in rheumatoid arthritis. Nat Commun.
14:10612023. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
McInnes IB and Schett G: The pathogenesis
of rheumatoid arthritis. N Engl J Med. 365:2205–2219. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Rheumatoid arthritis. Nat Rev Dis Primers.
4:180022018. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Zhao D, Jiang Z, Wang Z and Gao J:
Retinoid interferon-induced mortality19 (GRIM19) inhibits
proliferation and invasion in rheumatoid arthritis fibroblast-like
synoviocytes. Biomed Pharmacother. 98:719–725. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Liu F, Feng XX, Zhu SL, Huang HY, Chen YD,
Pan YF, June RR, Zheng SG and Huang JL: Sonic hedgehog signaling
pathway mediates proliferation and migration of fibroblast-like
synoviocytes in rheumatoid arthritis via MAPK/ERK signaling
pathway. Front Immunol. 9:28472018. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Shi W, Zheng Y, Luo S, Li X, Zhang Y, Meng
X, Huang C and Li J: METTL3 promotes activation and inflammation of
FLSs through the NF-κB signaling pathway in rheumatoid arthritis.
Front Med (Lausanne). 8:6075852021. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Chen J, Lin X, He J, Liu D, He L, Zhang M,
Luan H, Hu Y, Tao C and Wang Q: Artemisitene suppresses rheumatoid
arthritis progression via modulating METTL3-mediated
N6-methyladenosine modification of ICAM2 mRNA in fibroblast-like
synoviocytes. Clin Transl Med. 12:e11482022. View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Wang Q, Chen C, Ding Q, Zhao Y, Wang Z,
Chen J, Jiang Z, Zhang Y, Xu G, Zhang J, et al: METTL3-mediated
m6A modification of HDGF mRNA promotes gastric cancer
progression and has prognostic significance. Gut. 69:1193–1205.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
196
|
Li X, Xu X, Zhang Q, Ling M, Li X and Tan
X: METTL14 promotes fibroblast-like synoviocytes activation via the
LASP1/SRC/AKT axis in rheumatoid arthritis. Am J Physiol Cell
Physiol. 324:C1089–C1100. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Ai R, Laragione T, Hammaker D, Boyle DL,
Wildberg A, Maeshima K, Palescandolo E, Krishna V, Pocalyko D,
Whitaker JW, et al: Comprehensive epigenetic landscape of
rheumatoid arthritis fibroblast-like synoviocytes. Nat Commun.
9:19212018. View Article : Google Scholar : PubMed/NCBI
|
|
198
|
Beckmann D, Römer-Hillmann A, Krause A,
Hansen U, Wehmeyer C, Intemann J, de Gorter DJJ, Dankbar B, Hillen
J, Heitzmann M, et al: Lasp1 regulates adherens junction dynamics
and fibroblast transformation in destructive arthritis. Nat Commun.
12:36242021. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Lee DM, Kiener HP, Agarwal SK, Noss EH,
Watts GF, Chisaka O, Takeichi M and Brenner MB: Cadherin-11 in
synovial lining formation and pathology in arthritis. Science.
315:1006–1010. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
200
|
Tang J, Yu Z, Xia J, Jiang R, Chen S, Ye
D, Sheng H and Lin J: METTL14-mediated m6A modification of TNFAIP3
involved in inflammation in patients with active rheumatoid
arthritis. Arthritis Rheumatol. 75:2116–2129. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
201
|
Li R, Kuang Y, Niu Y, Zhang S, Chen S, Su
F, Wang J, Lin S, Liu D, Shen C, et al: FTO-mediated RNA
m6A methylation regulates synovial aggression and
inflammation in rheumatoid arthritis. Biochim Biophy Acta Mol Basis
Dis. 1870:1673412024. View Article : Google Scholar : PubMed/NCBI
|
|
202
|
Kuang Y, Li R, Wang J, Xu S, Qiu Q, Lin S,
Liu D, Shen C, Liu Y, Xu M, et al: ALKBH5-nediated RNA
m6A methylation regulates the migration, invasion, and
proliferation of rheumatoid fibroblast-like synoviocytes. Arthritis
Rheumatol. 76:192–205. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
203
|
Jin L, Chen Q, Hu K, Fan D, Zhang H, Deng
J, Qi W and Yu Q: The FTO-CMPK2 pathway in fibroblast-like
synoviocytes modulates rheumatoid arthritis synovial inflammation
and cartilage homeostasis via mtDNA regulation. Int J Biol Sci.
20:1617–1633. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
204
|
Huang Y, Xu P, Liao F, Ca H, Wang X, Wang
X, Chang J and Miao C: Fat mass and obesity-associated protein
inhibit the pathology of rheumatoid arthritis through the
NSUN2/SFRP1/Wnt/β-catenin signal axis. J Pharm Pharmacol.
76:283–294. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
205
|
Luo Q, Lan M, Wu Z, Wang S, Fu P, Xiao Q,
Fu B, Guo Y, Qing C, Huang Z and Li J: Decreased ALKBH5 in
neutrophil correlates with disease activity in rheumatoid arthritis
and ALKBH5 modulates neutrophil autophagy. Sci Rep. 15:378802025.
View Article : Google Scholar : PubMed/NCBI
|
|
206
|
Lyu FJ, Cui H, Pan H, Mc Cheung K, Cao X,
Iatridis JC and Zheng Z: Painful intervertebral disc degeneration
and inflammation: From laboratory evidence to clinical
interventions. Bone Res. 9:72021. View Article : Google Scholar : PubMed/NCBI
|
|
207
|
Peng B and DePalma MJ: Cervical disc
degeneration and neck pain. J Pain Res. 2853–2857. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
208
|
GBD 2021 Low Back Pain Collaborators, .
Global, regional, and national burden of low back pain, 1990–2020,
its attributable risk factors, and projections to 2050: A
systematic analysis of the Global Burden of Disease Study 2021.
Lancet Rheumatol. 5:e316–e329. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
209
|
GBD 2021 Neck Pain Collaborators, .
Global, regional, and national burden of neck pain, 1990–2020, and
projections to 2050: A systematic analysis of the Global Burden of
disease study 2021. Lancet Rheumatol. 6:e142–e155. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
210
|
Li XC, Luo SJ, Fan W, Zhou TL, Tan DQ, Tan
RX, Xian QZ, Li J, Huang CM and Wang MS: Macrophage polarization
regulates intervertebral disc degeneration by modulating cell
proliferation, inflammation mediator secretion, and extracellular
matrix metabolism. Front Immunol. 13:9221732022. View Article : Google Scholar : PubMed/NCBI
|
|
211
|
Wu T, Jia X, Zhu Z, Guo K, Wang Q, Gao Z,
Li X, Huang Y and Wu D: Inhibition of miR-130b-3p restores
autophagy and attenuates intervertebral disc degeneration through
mediating ATG14 and PRKAA1. Apoptosis. 27:409–425. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
212
|
Shi Y, Li H, Chu D, Lin W, Wang X, Wu Y,
Li K, Wang H, Li D and Xu Z: Rescuing nucleus pulposus cells from
senescence via Dual-functional greigite nanozyme to alleviate
intervertebral disc degeneration. Adv Sci (Weinh). 10:e23009882023.
View Article : Google Scholar : PubMed/NCBI
|
|
213
|
Wang J, Xia D, Li Y, Xu W, Wu Y, Chen J,
Chu J, Shen P, Weng S, Wang X, et al: Oxidative stress-induced
circKIF18A downregulation impairs MCM7-mediated anti-senescence in
intervertebral disc degeneration. Exp Mol Med. 54:285–297. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
214
|
Zhu B, Chen HX, Li S, Tan JH, Xie Y, Zou
MX, Wang C, Xue JB, Li XL, Cao Y and Yan YG: Comprehensive analysis
of N6-methyladenosine (m6A) modification during the degeneration of
lumbar intervertebral disc in mice. J Orthop Translat. 31:126–138.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
215
|
Smits P, Li P, Mandel J, Zhang Z, Deng JM,
Behringer RR, de Crombrugghe B and Lefebvre V: The transcription
factors L-Sox5 and Sox6 are essential for cartilage formation. Dev
Cell. 1:277–290. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
216
|
Wang Y, Li N and Wu X: Circular
RNA_0003800 exacerbates IL-1β-induced chondrocyte injury via
miR-197-3p/SOX5 axis. International Immunopharmacology.
115:1096432023. View Article : Google Scholar : PubMed/NCBI
|
|
217
|
Nakamichi R and Asahara H: The
transcription factors regulating intervertebral disc development.
JOR Spine. 3:e10812020. View Article : Google Scholar : PubMed/NCBI
|
|
218
|
Tsingas M, Ottone OK, Haseeb A, Barve RA,
Shapiro IM, Lefebvre V and Risbud MV: Sox9 deletion causes severe
intervertebral disc degeneration characterized by apoptosis, matrix
remodeling, and compartment-specific transcriptomic changes. Matrix
Biol. 94:110–133. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
219
|
Wang H, He P, Pan H, Long J, Wang J, Li Z,
Liu H, Jiang W and Zheng Z: Circular RNA circ-4099 is induced by
TNF-α and regulates ECM synthesis by blocking miR-616-5p inhibition
of Sox9 in intervertebral disc degeneration. Exp Mol Med. 50:1–14.
2018. View Article : Google Scholar
|
|
220
|
Gao D, Hu B, Ding B, Zhao Q, Zhang Y and
Xiao L: N6-Methyladenosine-induced miR-143-3p promotes
intervertebral disc degeneration by regulating SOX5. Bone.
163:1165032022. View Article : Google Scholar : PubMed/NCBI
|
|
221
|
Li G, Ma L, He S, Luo R, Wang B, Zhang W,
Song Y, Liao Z, Ke W, Xiang Q, et al: WTAP-mediated m6A
modification of lncRNA NORAD promotes intervertebral disc
degeneration. Nat Commun. 13:14692022. View Article : Google Scholar : PubMed/NCBI
|
|
222
|
Ariga K, Miyamoto S, Nakase T, Okuda S,
Meng W, Yonenobu K and Yoshikawa H: The relationship between
apoptosis of endplate chondrocytes and aging and degeneration of
the intervertebral disc. Spine (Phila Pa 1976). 26:2414–2420. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
223
|
Wang W, Jing X, Du T, Ren J, Liu X, Chen
F, Shao Y, Sun S, Yang G and Cui X: Iron overload promotes
intervertebral disc degeneration via inducing oxidative stress and
ferroptosis in endplate chondrocytes. Free Radic Biol Med.
190:234–246. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
224
|
Xiao L, Hu B, Ding B, Zhao Q, Liu C, Öner
FC and Xu H: N(6)-methyladenosine RNA methyltransferase like 3
inhibits extracellular matrix synthesis of endplate chondrocytes by
downregulating sex-determining region Y-Box transcription factor 9
expression under tension. Osteoarthritis Cartilage. 30:613–625.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
225
|
Yuan X, Li T, Shi L, Miao J, Guo Y and
Chen Y: Human umbilical cord mesenchymal stem cells deliver
exogenous miR-26a-5p via exosomes to inhibit nucleus pulposus cell
pyroptosis through METTL14/NLRP3. Mol Med. 27:912021. View Article : Google Scholar : PubMed/NCBI
|
|
226
|
Song Y, Wang Z, Liu L, Zhang S, Zhang H
and Qian Y: 1,4-Dihydropyridine (DHP) suppresses against oxidative
stress in nucleus pulposus via activating sirtuin-1. Biomed
Pharmacother. 121:1095922020. View Article : Google Scholar : PubMed/NCBI
|
|
227
|
Xiang Q, Kang L, Wang J, Liao Z, Song Y,
Zhao K, Wang K, Yang C and Zhang Y: CircRNA-CIDN mitigated
compression loading-induced damage in human nucleus pulposus cells
via miR-34a-5p/SIRT1 axis. EBioMedicine. 53:1026792020. View Article : Google Scholar : PubMed/NCBI
|
|
228
|
Zhu H, Sun B, Zhu L, Zou G and Shen Q:
N6-methyladenosine induced miR-34a-5p promotes TNF-α-induced
nucleus pulposus cell senescence by targeting SIRT1. Front Cell Dev
Biol. 9:6424372021. View Article : Google Scholar : PubMed/NCBI
|
|
229
|
Lei Y, Zhan E, Chen C, Hu Y, Lv Z, He Q,
Wang X, Li X and Zhang F: ALKBH5-mediated m6A
demethylation of Runx2 mRNA promotes extracellular matrix
degradation and intervertebral disc degeneration. Cell Biosci.
14:792024. View Article : Google Scholar : PubMed/NCBI
|
|
230
|
Gao X, Liang X, Liu B, Hong Y, He H, Shen
Y, Chen J, Huang X, Hu B, Li W, et al: Downregulation of ALKBH5
rejuvenates aged human mesenchymal stem cells and enhances their
therapeutic efficacy in myocardial infarction. FASEB J.
37:e232942023. View Article : Google Scholar : PubMed/NCBI
|
|
231
|
Li G, Luo R, Zhang W, He S, Wang B, Liang
H, Song Y, Ke W, Shi Y, Feng X, et al: m6A hypomethylation of
DNMT3B regulated by ALKBH5 promotes intervertebral disc
degeneration via E4F1 deficiency. Clin Transl Med. 12:e7652022.
View Article : Google Scholar : PubMed/NCBI
|
|
232
|
Zheng-Wei S, Yuan T, Chao-Shuai F, Lei Z,
Zong-Rang S, Tuan-Jiang L and Ding-Jun H: Roles of Hippo-YAP/TAZ
signalling in intervertebral disc degeneration. Biomed
Pharmacother. 159:1140992023. View Article : Google Scholar : PubMed/NCBI
|
|
233
|
Sun R, Wu XT, Shi H, Wang F, Gao JW, Wang
PY, Xu ZY, Gan WW, Wang YT and Zhang C: Mechanism of FTO-mediated
m6A demethylation regulation of YAP1 in nucleus pulposus cell
senescence. Mech Ageing Dev. 227:1121012025. View Article : Google Scholar : PubMed/NCBI
|
|
234
|
Cerneckis J, Ming GL, Song H, He C and Shi
Y: The rise of epitranscriptomics: Recent developments and future
directions. Trends Pharmacol Sci. 45:24–38. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
235
|
Han J, Wang C, Yang H, Luo J, Zhang X and
Zhang XA: Novel Insights into the Links between N6-Methyladenosine
and regulated cell death in musculoskeletal diseases. Biomolecules.
14:5142024. View Article : Google Scholar : PubMed/NCBI
|
|
236
|
Li P, Zhang C, Yin W, Tao M, Niu Z, Cui Y,
Wu D and Gao F: From bone marrow mesenchymal stem cells to
diseases: The crucial role of m6A methylation in
orthopedics. Stem Cell Res Ther. 16:2282025. View Article : Google Scholar : PubMed/NCBI
|
|
237
|
Moshitch-Moshkovitz S, Sevilla-Sharon M,
Ashwal-Fluss R, Glick-Saar E, Rechavi G and Dominissini D: mRNA m6A
detection. Nat Rev Methods Primers. 4:872024. View Article : Google Scholar
|
|
238
|
Grozhik AV and Jaffrey SR: Distinguishing
RNA modifications from noise in epitranscriptome maps. Nat Chem
Biol. 14:215–225. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
239
|
Yang Y, Lu Y, Wang Y, Wen X, Qi C, Piao W
and Jin H: Current progress in strategies to profile transcriptomic
m6A modifications. Front Cell Dev Biol. 12:13921592024.
View Article : Google Scholar : PubMed/NCBI
|
|
240
|
Garcia-Campos MA, Edelheit S, Toth U,
Safra M, Shachar R, Viukov S, Winkler R, Nir R, Lasman L, Brandis
A, et al: Deciphering the ‘m6A code’ via
antibody-independent quantitative profiling. Cell. 178:731–747.e16.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
241
|
Cun Y, Guo W, Ma B, Okuno Y and Wang J:
Decoding the specificity of m6A RNA methylation and its implication
in cancer therapy. Mol Ther. 32:2461–2469. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
242
|
Dominissini D, Moshitch-Moshkovitz S,
Salmon-Divon M, Amariglio N and Rechavi G: Transcriptome-wide
mapping of N(6)-methyladenosine by m(6)A-seq based on
immunocapturing and massively parallel sequencing. Nat Protoc.
8:176–189. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
243
|
Linder B, Grozhik AV, Olarerin-George AO,
Meydan C, Mason CE and Jaffrey SR: Single-nucleotide-resolution
mapping of m6A and m6Am throughout the transcriptome. Nat Methods.
12:767–772. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
244
|
McIntyre ABR, Gokhale NS, Cerchietti L,
Jaffrey SR, Horner SM and Mason CE: Limits in the detection of m6A
changes using MeRIP/m6A-seq. Sci Rep. 10:65902020. View Article : Google Scholar : PubMed/NCBI
|
|
245
|
Wang Y, Xiao Y, Dong S, Yu Q and Jia G:
Antibody-free enzyme-assisted chemical approach for detection of
N6-methyladenosine. Nat Chem Biol. 16:896–903. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
246
|
Ge R, Ye C, Peng Y, Dai Q, Zhao Y, Liu S,
Wang P, Hu L and He C: m6A-SAC-seq for quantitative whole
transcriptome m6A profiling. Nat Protoc. 18:626–657. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
247
|
Xiao Y, Wang Y, Tang Q, Wei L, Zhang X and
Jia G: An Elongation- and ligation-based qPCR amplification method
for the Radiolabeling-free detection of Locus-specific
N6-methyladenosine modification. Angew Chem Int Ed Engl.
57:15995–16000. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
248
|
Gilbert WV and Nachtergaele S: mRNA
regulation by RNA modifications. Annu Rev Biochem. 92:175–198.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
249
|
Moser JC, Papadopoulos KP, Rodon Ahnert J,
Ofir-Rosenfeld Y and Holz JB; STC15-22101 Study Team, : Phase 1
dose escalation and cohort expansion study evaluating safety, PK,
PD and clinical activity of STC-15, a METTL-3 inhibitor, in
patients with advanced malignancies. J Clin Oncol. 42:25862024.
View Article : Google Scholar
|
|
250
|
Yankova E, Blackaby W, Albertella M, Rak
J, De Braekeleer E, Tsagkogeorga G, Pilka ES, Aspris D, Leggate D,
Hendrick AG, et al: Small-molecule inhibition of METTL3 as a
strategy against myeloid leukaemia. Nature. 593:597–601. 2021.
View Article : Google Scholar : PubMed/NCBI
|