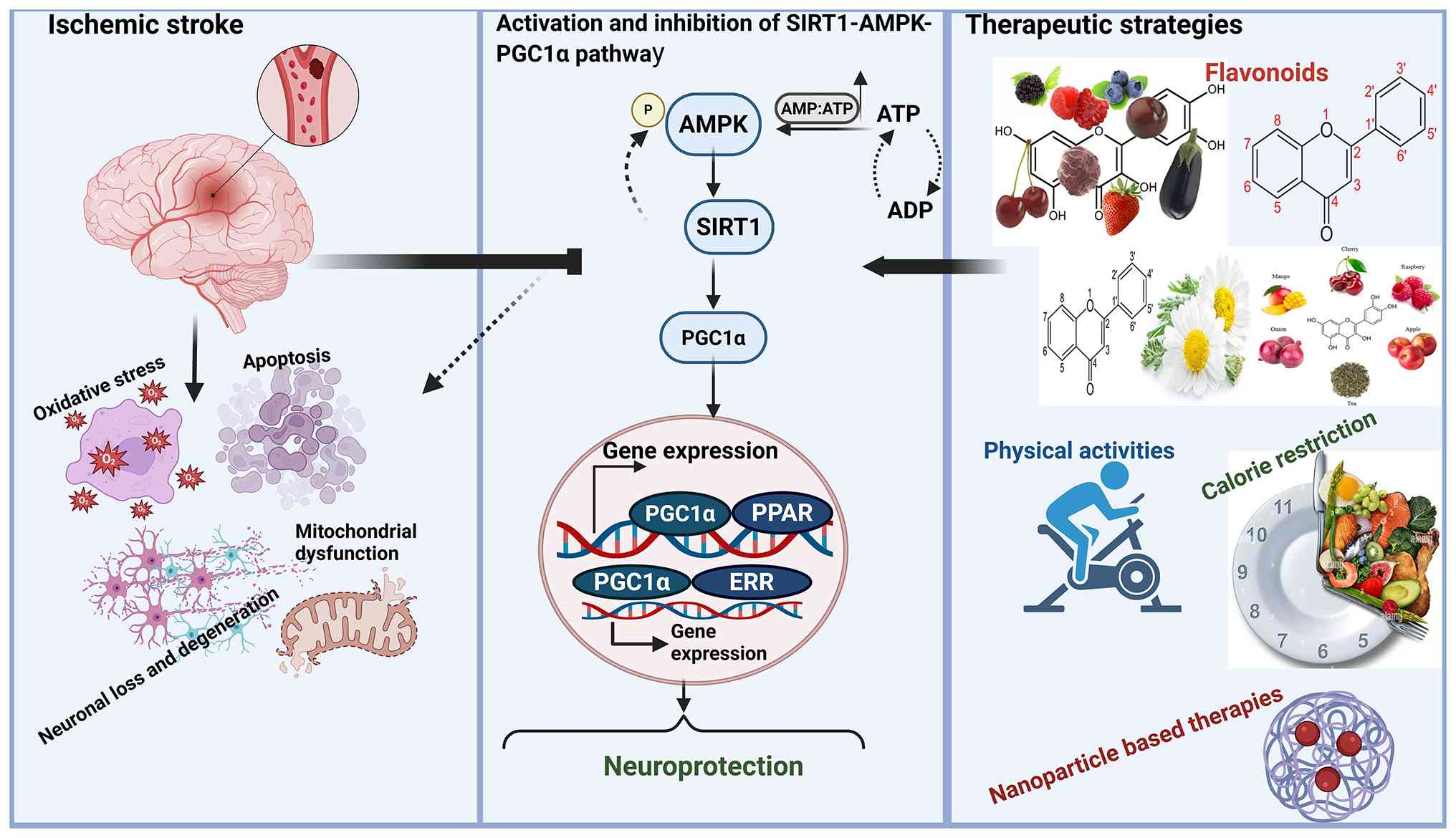
Introduction
Globally, stroke is the second largest cause of
death, and ischemic strokes make up >60% of all strokes
(1). Moreover, it is a main cause
of neurological problems and long-term disability in resource-poor
settings (1,2). Ischemia occurs when cerebral blood
flow to the brain is suddenly reduced or blocked, triggering
oxidative stress, damaged mitochondria and inflammation (3,4).
These events collectively lead to nerve cell injury and
degeneration causing sustained neurological deficits (5).
The silent information regulator 2 homolog (SIRT)
1/AMP-activated protein kinase (AMPK)/peroxisome
proliferator-activated receptor γ coactivator 1-α (PGC1α) pathway
has been recognized as a key controller of cell metabolism, new
mitochondria formation and neuroprotection (6,7).
Activation of this pathway reduces oxidative stress, enhances
mitochondrial function and decreases neuronal damage, highlighting
its therapeutic potential in the treatment of ischemic stroke
(8,9). However, whilst individual aspects of
this pathway have been assessed in numerous neurological
conditions, a systemic review providing an integrated view
specifically for ischemic stroke is lacking (10,11).
We hypothesize that the SIRT1/AMPK/PGC1α pathway
mediates endogenous neuroprotection in ischemic stroke, and that
its targeted regulation offers promise for therapeutic development.
The present review had three primary aims: i) To synthesize and
evaluate research on the role of this pathway in stroke pathology
and recovery; ii) to integrate findings on multi-targeted
therapeutic approaches (such as flavonoids, physical activity and
caloric restriction) that confer protection via this pathway; and
iii) to highlight emerging research directions, including
brain-specific PGC1α isoforms and novel drug carrier systems that
pass through the blood-brain barrier (BBB). Overall, the present
review aimed to elucidate the complex function of this pathway and
establish a foundation for innovative treatment strategies for
ischemic stroke. A conceptual model summarizing these aims is
presented in Fig. 1.
SIRT1, AMPK and PGC1α and their
interconnections
Sirtuins, particularly SIRT1, confer neuroprotective
effects during ischemic events by mitigating stroke-induced damage
(12,13). These proteins function to
deacetylate histones and transcription factors, thereby regulating
gene expression and the activity of metabolic enzymes in response
to ischemic stress. Mammals have seven types of sirtuins
(SIRT1-SIRT7), with SIRT1 being the most extensively investigated
when it comes to ischemia (14).
When SIRT1 is activated or upregulated, it mitigates ischemic brain
injury, reduces infarct size and enhances neurological outcomes
(15,16). SIRT1 deacetylates transcription
factors and coactivators, including PGC1α, thereby enhancing the
activation of genes that are crucial for brain recovery following
ischemic injury (17).
Moreover, lower AMPK activity within the brain and
spinal cord [central nervous system (CNS)] is also associated with
increased pathology in conditions such as Alzheimer's disease
(18,19). By contrast, when AMPK is activated,
it raises NAD+ levels and thereby activates SIRT1. As a result,
this promotes neuroprotection (20–22).
The co-activator PGC1α also has a notable role in
the production of new mitochondria and the scavenging of reactive
oxygen species (ROS) (23,24). Its expression is mainly restricted
to tissues with high energy demands (such as the brain) and is
regulated by metabolic stimuli such as caloric restriction,
physical exercise or hypoxia (25). Notably, PGC1α is markedly expressed
in certain areas of the brain (cortex and striatum), whereas it is
not found in the hypothalamus (26). Structural models suggest that PGC1α
promotes polymerase II recruitment with the cooperation of
transcription factors of the nuclear receptor family, such as
peroxisome proliferator activated receptors (PPARs), estrogen
receptor α and retinoid × receptor α (25,27).
This mechanism is mediated through acetyl and methyltransferase
protein groups along with helper proteins such as steroid receptor
coactivator 1, CRE-binding proteins (CBP/p300) (28). The distribution of PGC1α between
the nucleus and cytoplasm is modulated by energy-sensing molecules,
including SIRT1, AMPK and histone acetyltransferases (29). The acetylation of PGC1α, which is
directly mediated by the general control non-repressed 5 protein,
leads to a reduction in how well it can activate gene transcription
(23). Under conditions of energy
stress, the NAD+/NADH ratio is elevated, leading to the activation
of SIRT1. Subsequently, SIRT1 deacetylates PGC1α, thereby enhancing
its transcriptional activity and then increasing the generation of
antioxidant proteins, including glutathione peroxidase and
superoxide dismutase (24,30). AMPK additionally activates the
PGC1α/SIRT1-dependent antioxidant system by enhancing the
expression of antioxidant enzymes, thereby sustaining mitochondrial
equilibrium during disruptions in cellular energy (29,31).
The interaction among SIRT1, AMPK and PGC1α is of
notable importance, establishing a pathway with therapeutic
potential for diseases associated with aging, particularly those
impacting the nervous system (Table
I) (32). Specifically,
excessive activation of AMPK may adversely affect synaptic
plasticity by inhibiting MORC1 (31) and sustained overexpression of SIRT1
could enhance metabolic resilience in cancer cells (33). Additionally, an increase in PGC1α
has been associated with elevated oxidative stress in neurons
affected by Parkinson's disease (33). Furthermore, inflammatory signals
and metabolic stress may attenuate the benefit of this pathway
(34).
 | Table I.Potential benefits of
SIRT1/AMPK/PGC1α signaling in management of different
neurodegenerative disorders. |
Table I.
Potential benefits of
SIRT1/AMPK/PGC1α signaling in management of different
neurodegenerative disorders.
| Disease | Species | Drug | Doses | Results | (Refs.) |
|---|
| Aging | VSMC senescence,
mouse Na2 neuroblastoma cell line | Quercetin | 50 µm | Activation of AMPK
in VSMCs | (109) |
| Alzheimer's
disease | Institute of Cancer
Research mouse strain mice with Aβ25-35-induced Alzheimer's like
pathology | Astaxanthin | 10 mg/kg | Turned on
SIRT1/PGC1α pathway in hippocampus. Lowered hippocampal oxidative
stress | (54) |
| Vascular cognitive
impairment | Sprague Dawley rat
model of vascular dementia following bilateral carotid artery
ligation | Triptolide | 5 µg/kg/day | Upregulated SIRT1
in hippocampus CA1 area. Reduced serum S100B and neuron-specific
enolase levels. | (100) |
| Diabetic cognitive
impairment | Zucker diabetic
fatty rats | High-fat diet
combined with Zibu Piyin Recipe | Daily dose of 32.9
g/kg | Upregulated PGC1α
and Mfn2 in cortex/hippocampus. Enhanced mitochondrial structure
and function | (102) |
| Ischemic brain
injury | Transient MCAO (1.5
h) model in male and female Sprague-Dawley rats | Luteolin | 15, 30 and 60
mg/kg | Activated cerebral
SIRT3/AMPK/mTOR pathway. Enhanced SIRT3 transduction in rat
brain | (106) |
| Depression-like
behavior | Male Dawley rats
and BV2-SH-SY5Y | Baicalin | In vivo:
30–60 mg/kg; In vitro: 10-50-100 µM | Elevated
hippocampal PGC1α levels and reduced depression like behaviors.
PGC1α upregulation in SH-SY5Y cells | (37) |
| Parkinson's
disease | Rotenone or
MPP+-treated SH-SY5Y cells, PC12 cells and zebrafish | Resveratrol
Teaghrelin Panaxatriol saponins | 0–50 µm 1–100 µm
0–4 mg/ml | The activation of
AMPK/SIRT1 enhances autophagy and clears misfolded proteins and
damaged mitochondria | (109) |
| Multiple
sclerosis | Cuprizone-exposed
male C57Bl/6 mice (n=48) | Linagliptin | 10 mg/kg | AMPK/SIRT1
activation safeguarded neurons by lowering oxidative stress and
demyelination damage | (56) |
The present review distinctively emphasizes the
collaborative mechanisms by which these proteins regulate
mitochondrial function, oxidative stress and neuroinflammation,
thereby providing novel insights into multi-targeted treatment
approaches. Additionally, future studies should evaluate the role
of CNS-specific PGC1α isoforms, which may represent new therapeutic
targets for stroke recovery.
SIRT1/AMPK/PGC1α signaling and structural
features
SIRT1: Molecular structure and
biological functions
SIRT1 serves as a pivotal regulator of metabolic
processes, aging and cellular responses, encompassing apoptosis,
inflammation and oxidative stress (35,36).
SIRT1 is found in both the nucleus and cytoplasm, where it
regulates several cellular processes (36). This protein shows high levels
throughout human tissues, with a notably strong presence within the
neural tissues (33).
SIRT1 activation is modulated by factors such as
physical exercise and hypoxia. It primarily acts like a deacetylase
enzyme targeting histone along with other non-histone proteins
including forkhead box O (FOXO), P53 and NF-κB, and in doing so
modulates how cells react to oxidative stress, apoptosis and
inflammation (37–39). Moreover, SIRT1 serves a regulatory
function in regulating autophagy, new mitochondria formation and
cell longevity, which make it a promising treatment target for
aging and disease (40).
AMPK: Structural insights, activation
and physiological functions
AMPK is composed of α, β and γ subunits along with
numerous isoforms (α1/α2, β1/β2 and γ1/γ2/γ3), forming 12
configurations. The α1, β1 and γ1 subunits are ubiquitously
expressed, whereas the α2, β2 and γ2/γ3 are mainly expressed in
cardiac and skeletal muscles (41,42).
Activation occurs when Thr172 on the α-subunit is phosphorylated,
mediated by CaMKKβ and liver kinase B1 (LKB1) (43,44).
LKB1 activity is AMP/ATP ratio dependent and phosphatases help
stabilize Thr172 phosphorylation (44,45).
The γ-subunit binds AMP/ATP for allosteric regulation, and the
β-subunit contains a carbohydrate binding module influencing
activity (41,43).
Direct activators, such as the thiophene pyridine
derivative A-769662, bind the β1 subunit which enhances activity
and prevents dephosphorylation (45,46).
Other activators include M2958-7438, M5050-0116 (β1-specific) and
C2 (γ-subunit) (44). Indirect
activators, such as the flavonoid quercetin, act via LKB1-AMPK
signaling to improve cardiovascular health by enhancing endothelial
function, reducing oxidative stress and improving lipid metabolism
(47). AMPK regulates energy
balance through blocking anabolic pathways and encouraging
catabolic pathways (45). It
alleviates diabetic nephropathy via Akt and Nrf2 (48), promotes autophagy and reduces
inflammation by elevating HIF-1α. Additionally, it collaborates
with SIRT1 to prevent lipid accumulation and mitochondrial
dysfunction (34,49). Its activation also offers
neuroprotection through improvement of mitochondrial activity and
activation of autophagy via ULK1 phosphorylation and mTOR
inhibition (43). AMPK serves a
notable function in regulating energy homeostasis by boosting
mitochondrial performance and supporting neuronal survival through
activation of autophagy via the ULK1 pathway (50), inhibition of mTOR signaling and
modulation of the SIRT1/PGC1α signaling axis (31).
PGC1α: Structural insights and
physiological functions
The PGC1 family includes PGC1α, PGC1β and PRC. PGC1α
and 1β share a high sequence similarity in their N-terminal
activation and C-terminal RNA binding domains (15) and are found in metabolically active
organs including the brain, heart and brown adipose tissue
(23). PRC is more ubiquitous, but
its function is less understood (15). The N-terminal region contains LXXLL
motifs for recruiting transcriptional coactivators such as SRC-1
and CBP/p300 (23). Host cell
factor (HCF) contains a repression domain and RNA recognition
motifs that modulate transcription and splicing. HCF enhances
transcriptional activity during the cell cycle (15). Its C-terminal domain interacts with
transcription factors such as FOXO1 and YY1, thereby co-activating
PPARs, NRFs and ERRs, which regulate genes involved in
mitochondrial function, oxidative stress and metabolism (15,23).
PGC1α regulates mitochondrial biogenesis by
activating NRF1/2, which control genes such as TFAM, PLOG
and cytochrome c oxidase subunits involved in mitochondrial
DNA replication and the electron transport chain (51). This process is stimulated by
exercise, caloric restriction and hormones, such as adiponectin and
leptin, via AMPK/SIRT1 signaling, particularly after stroke
(52). In a photothrombotic stroke
model, mexidol (100 mg/kg) and semax (25 µg/kg) increased neurons
with high nuclear PGC1α immunoreactivity by ~3- and 2.5-fold,
respectively, at day 7, and increased total PGC1α expressing
neurons by 1.5- and 1.4-fold, respectively, at day 21.
PGC1α also promotes mitophagy and its dysregulation
is associated with diabetes and neurodegeneration (51). It supports antioxidant defense via
Nrf2, reduces Bax and increases Bcl2, thereby limiting oxidative
stress induced apoptosis (53). In
neurodegenerative models, increased expression or activation of
PGC1α, achieved through pharmacological treatment or overexpression
approaches, reduces mitochondrial dysfunction and neuronal damage
(25). Overall, PGC1α is essential
in managing energy metabolism, oxidative stress and cell survival
especially via the SIRT1/AMPK pathway, which holds therapeutic
promise for neurodegenerative diseases (54) (Fig.
2).
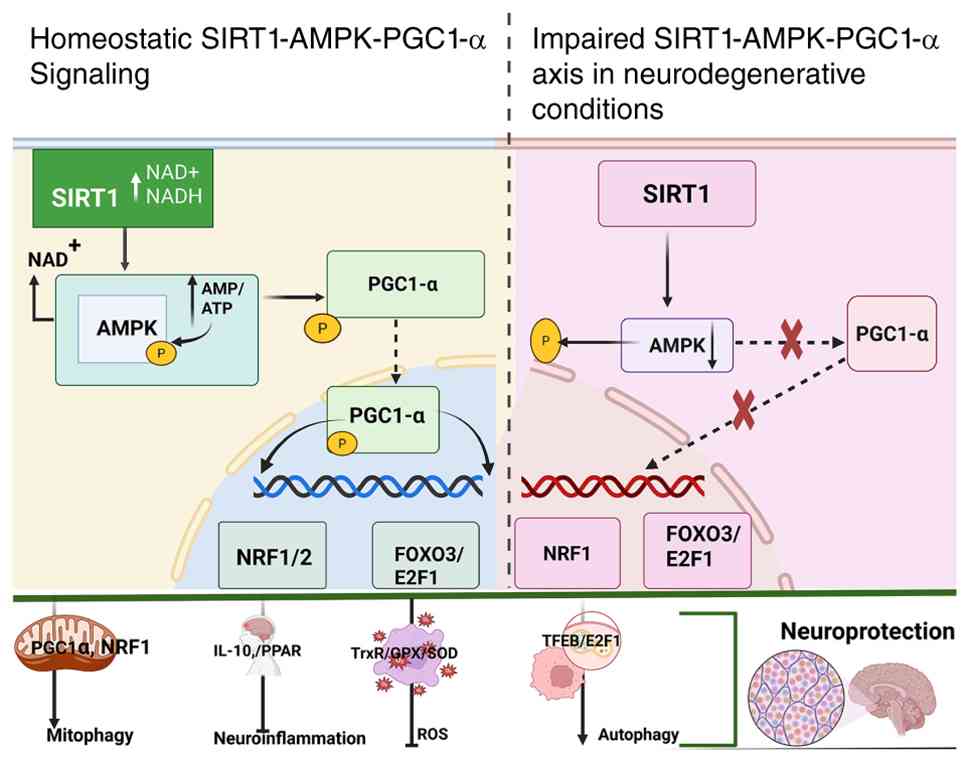 | Figure 2.The SIRT1-AMPK-PGC1α signaling
pathway serves an essential function in both physiological and
diseased states. In healthy states, SIRT1 and AMPK react in
response to reduced energy availability or diminished metabolic
breakdown rates, initiating the addition of phosphate groups and
removal acetyl groups from PGC1α, in that order. However, during a
stroke, reduced activation of AMPK and SIRT1 prevents PGC1α from
undergoing deacetylation and phosphorylation, hindering its
translocation to the nucleus. This distortion has been reported to
impact several vital processes, such as the management of ROS
levels, the regulation of Mitochondrial Homeostasis, the
encouragement of Autophagy, control over Neuroinflammation and
assistance in Synapse formation that consequently led to stroke.
Based on the theory by Rakshe et al (24). SIRT1, silent information regulator
2 homolog 1; AMPK, AMP-activated protein kinase; ROS, reactive
oxygen species; PGC1α, peroxisome proliferator-activated receptor γ
coactivator 1-α. |
Role of SIRT1/AMPK/PGC1α in cerebral
ischemia
The SIRT1/AMPK/PGC1α pathway is a notable protective
survival signal in neurons as it controls cellular metabolism,
energy homeostasis and neuroprotection. The upregulation of this
pathway provides a potential therapeutic target for ischemic stroke
(55). AMPK, an energy sensor,
serves as a primary controller of bioenergetic metabolism and
cellular growth. Its activation by phosphorylation of the α subunit
at Thr172 is anti-apoptotic and anti-neuroinflammatory leading to
increased cell survival (49).
AMPK activates SIRT1 by increasing cellular NAD+ levels, which acts
as an essential co-substrate for SIRT1 deacetylation of LKB1. This
reciprocal crosstalk modulates the PGC1α, FOXO1 and NF-κB signaling
axis, leading to reduced apoptosis and inflammation and enhanced
neuronal survival (25). The
decreased PGC1α is associated with elevated oxidative stress
levels, reduced mitochondrial number and neuronal loss. On the
other hand, the SIRT1/AMPK/PGC1α axis is beneficial for the
neuronal survival and mitochondrial function during ischemic stroke
(6). For example, the adipokine
CTRP3 activates this pathway, protects mitochondrial function and
controls mitochondrial dynamics such as fission and fusion. Within
hippocampal neurons subjected to oxygen-glucose deprivation
followed by reperfusion, CTRP3 enhanced viability, reduced
apoptosis and promoted mitochondrial biogenesis whereas PGC1α
silencing abolished these protective effects (7).
Previous research has underscored the
neuroprotective function of PGC1α. Mice deficient in PGC1α were
reported to have increased infarct sizes, notable motor and
cognitive impairments and elevated oxidative stress and
inflammation following a stroke (9,10).
These observations highlight the neuroprotective function of PGC1α
within the context of cerebral ischemia and suggest the possibility
of using it as a therapeutic candidate for managing stroke.
The SIRT1/AMPK/PGC1α pathway is integral to
neuroprotection following ischemic stroke, as it modulates
inflammation, oxidative stress and mitochondrial function (56). Notably, in vivo studies
provide data supporting their therapeutic utility. The knockout or
inhibition of SIRT1 (such as using EX527) negates (i.e., abolishes)
the neuroprotective effects of compounds such as resveratrol and
enhances HMGB1 acetylation and NLRP3 inflammasome activation,
thereby confirming that loss of SIRT1 activity is detrimental and
exacerbates ischemic injury (57).
In mice subjected to 1-h middle cerebral artery occlusion (MCAO)
followed by reperfusion for 24 h, resveratrol was associated with
reduced infarct size, brain edema and neurological impairments via
SIRT1 dependent autophagy, as these effects were blocked by
3-methyladenine or SIRT1 small interfering RNA. In permanent focal
ischemia, SIRT1 activation (activator 3, 10 mg/kg) was associated
with reduced infarct volume, whereas SIRT1 inhibition (sirtinol, 10
mg/kg) and SIRT1 deletion were associated with enlarged injury and
increased p53/NF-kB acetylation (6).
Similarly, AMPKα-deficient mice or those treated
with AMPK inhibitors (such as compound C) exhibit increased infarct
volume, enhanced neuronal death and worsened neurological deficits,
underscoring the essential role of AMPK in mitochondrial biogenesis
and neuronal survival (24).
Furthermore, PGC1α knockout or knockdown were associated with
compromised mitochondrial function and heightened neuronal
susceptibility, emphasizing its neuroprotective function (58). In a rodent photothrombotic stroke
model, intranasal mitochondrial administration (100 µg protein),
given at 30 min, 24 and 48 h, was associated with reduced infarct
volume and edema with increased p-AMPKα, PGC1α and restored SIRT1
(59).
Collectively, these findings substantiate the
involvement of SIRT1, AMPKα and PGC1α in stroke protection,
rendering them promising therapeutic targets. Despite the overall
neuroprotective role of this axis, the strength of evidence varies
across its components. SIRT1 activation consistently reduces
infarct volume and inflammation, whereas AMPKα exhibits
context-dependent effects; early post stroke activation reduces
neuronal death and promotes autophagy, while excessive or late
activation may exacerbate apoptosis under energy depleted
conditions (60). Similarly,
autophagy regulation via this axis shows duality, with both
pro-autophagic and anti-autophagic strategies reported as
protective. By contrast, the SIRT1/PGC1α arm demonstrate more
consistent benefits in mitochondrial preservation. Taken together,
these observations suggest that while the pathway is a promising
therapeutic target, its actions are notably context-sensitive and
require careful consideration of timing, magnitude and cell type
specificity (61). Furthermore,
the administration of quercetin has been reported to mitigate
oxidative stress and promote neuronal recovery by upregulating
phosphorylated AMPK, PGC1α, SIRT1, NRF1 and Tfam (53,62).
Furthermore, Icariin has been reported to facilitate mitochondrial
biogenesis and mitigates ROS through the activation of AMPK,
highlighting the notable role of mitochondrial regulation in the
recovery process following a stroke (63,64).
SIRT1 has also been reported to attenuate neuroinflammation through
inhibition of NF-κB and lowering oxidative stress. The elevated
levels of SIRT1 observed in human patients with stroke indicate its
potential as a biomarker for assessing stroke severity (65–67).
These findings collectively underscore how critical the
SIRT1/AMPK/PGC1α axis in ischemic stroke, elucidating its
connection to metabolic disturbances, neuroinflammation and
mitochondrial dysfunction. Therefore, modulating this axis offers a
promising strategy for mitigating neuronal damage and enhancing
recovery (Fig. 3).
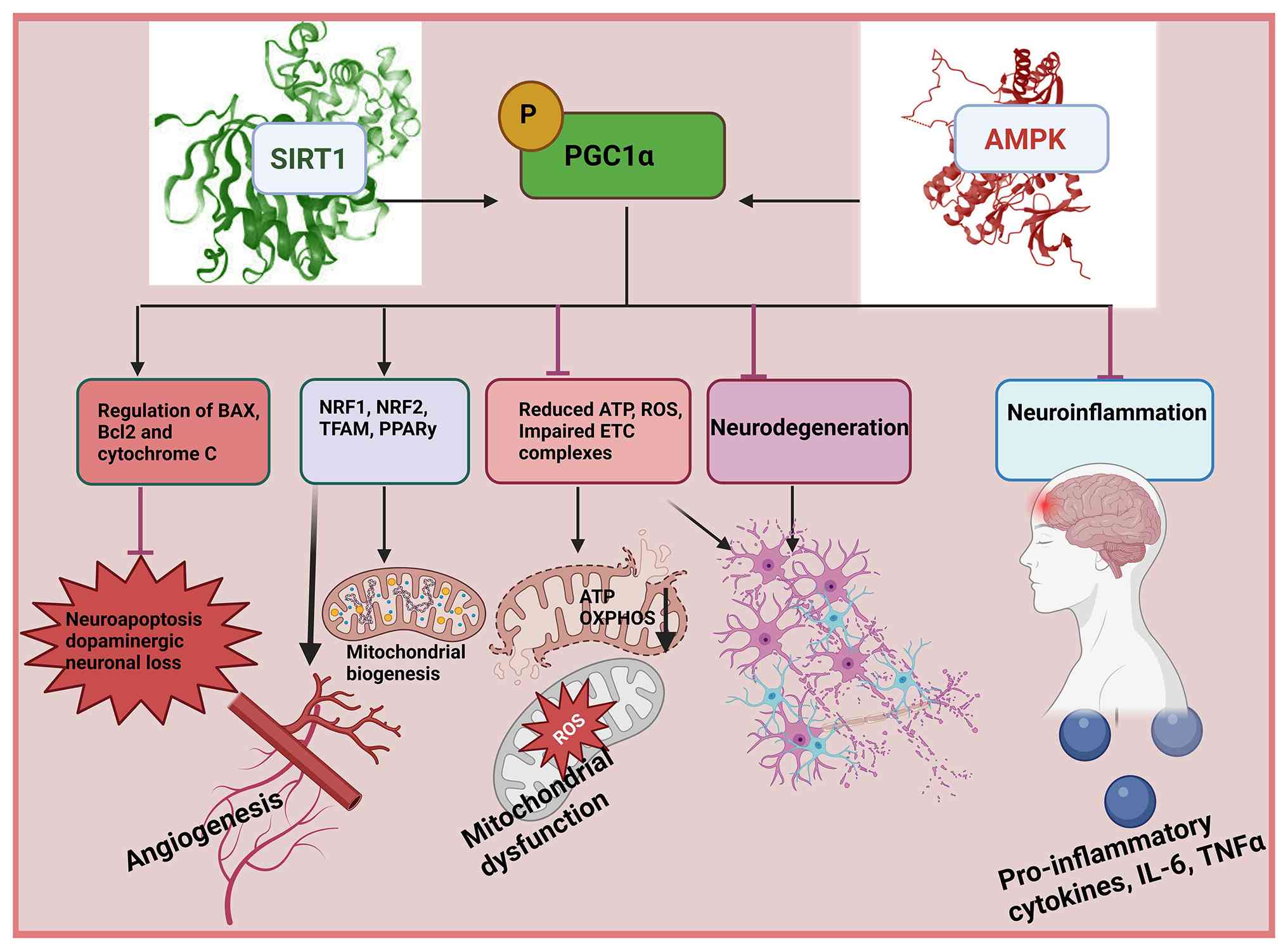 | Figure 3.In stroke, PGC1α has neuroprotective
properties by modulating multiple signaling cascades that
participate in the progression of the disease. These pathways
include impaired mitochondrial function, oxidative stress,
proteasome impairment, neuroinflammatory responses along with
autophagic and apoptotic processes. By targeting these pathogenic
processes, PGC1α greatly participates in the preservation of
neuronal cells in stroke (108).
Activation of PGC1α inhibits microglial activation by decreasing
cytokine generation and cell death through inhibition of Bax and
IL-1β and augmentation of Bcl-2. PGC1α enhances antioxidant,
mitochondria biogenesis, O2 consumption membrane
potential cellular recycling and the function of transcription
factors. Consequently, PGC1α activators show promise in controlling
gene expression supporting neuronal survival and offering
protective effects in neurons by addressing mitochondrial
malfunction oxidative damage proteasome impairment, autophagic
processes, neuroinflammatory responses and cell death (109). PGC1α, peroxisome
proliferator-activated receptor γ coactivator 1-α; ROS, reactive
oxygen species; AMPK, AMP-activated protein kinase. |
Therapeutic strategies targeting the
AMPK/SIRT1/PGC1α axis
Conceptual framework for axis
targeting
Targeting the SIRT1-AMPK pathway to elevate PGC1α
levels constitutes a viable treatment strategy for the management
of cerebral ischemia, offering potential advantages in
neuroprotection and functional recovery (34). Nonetheless, precise modulation of
this pathway is required, as both excessive activation and
inhibition may result in detrimental effects. Current research
increasingly emphasizes designing targeted treatment approaches for
cerebral stroke, integrating pharmacological interventions (such as
small molecules and flavonoids) with non-pharmacological methods
(such as exercise and caloric restriction) to enhance
neuroprotection and functional recovery (6).
Non-pharmacological approaches
Physical activity
Engagement in physical activity enhances the
adaptability of the brain, yielding marked benefits for both the
prevention and recovery of stroke. Regular exercise increases
resistance to oxidative stress, a notable factor in the pathology
of cerebral ischemia, in part through the activation of SIRT1
signaling cascades, which serve to mitigate cerebral damage
(68). Engaging in physical
activity triggers the release of brain-derived neurotrophic factor,
a notable mediator of neuroplasticity, along with insulin-like
growth factor 1. Both of these factors facilitate synapse
formation, neuronal growth and recovery following a stroke
(68,69). Additionally, physical exercise
enhances mitochondrial biogenesis, required for maintaining
neuronal energy homeostasis during and after ischemic events
(68,70). Both before and after a stroke,
physical activity reduces cerebral damage by influencing excitatory
amino acid transporters (EAATs), such as EAAT2, which clears
glutamate from synaptic cleft, and modulating the ERK1/2 signaling
pathways (70,71). The hippocampus, particularly the
CA1 region, is highly susceptible to stroke-induced damage
(63). Physical activity can
reactivate the SIRT1/AMPK/PGC1α pathway, protecting hippocampal
neurons, improving functional outcomes, and reducing the risk of
recurrent strokes (6,72,73).
Regular physical activity mitigates stroke-induced decreases in
AMPK activity and PGC1α expression, thereby restoring
neuroprotective mechanisms and facilitating recovery. These
advantageous effects occur via stimulation of the SIRT1/AMPK/PGC1α
axis, leading to improved mitochondrial function and diminishes
neuronal injury (63,69,73).
Dietary energy restrictions
The AMPK/SIRT1/PGC1α signaling axis serves an
essential role in cellular metabolism and mitochondrial biogenesis,
suggesting that dietary energy restriction could represent a
valuable treatment strategy against stroke. Caloric restriction,
when coupled with adequate nutrient intake, activates
PGC1α-mediated mitochondrial genesis, thereby enhancing neuronal
survival following a stroke (73).
SIRT1, a pivotal mediator of dietary energy restriction, enhances
neuroprotection in ischemic stroke by activating AMPK and
modulating several transcriptional regulators including FOXO1,
NF-κB and PGC1α. The upregulation of SIRT1 expression mitigates
neuronal damage and facilitates post-stroke recovery (63,66,74).
Dietary energy restriction induces the activation of SIRT1, which
subsequently upregulates PGC1α, thereby enhancing mitochondrial
biogenesis and cellular respiratory function. This mechanism
facilitates an increase in mitochondrial mass and promotes cell
survival, especially during cerebral ischemia (35,39,49).
The deficiency of PGC1α results in striatal degeneration,
underscoring its role in ischemic stroke. The upregulation of PGC1α
expression during cerebral reperfusion injury confers cellular
protection by modulating Nrf-2, preserving mitochondrial function,
and mitigating brain damage (75,76).
Exercise and caloric restriction engage the SIRT1/AMPK/PGC1α
pathway, offering neuroprotective effects. However, the detailed
mechanisms and clinical challenges such as patient compliance and
treatment protocol standardization limit their implementation in
stroke therapy.
Pharmacological approaches
Small molecule activators of the triad
Pharmacological interventions targeting the
AMPK/SIRT1/PGC1α pathway have emerged as a promising strategy for
the treatment of ischemic stroke, a condition characterized by
mitochondrial dysfunction, oxidative stress and neuroinflammation.
The AMPK/SIRT1/PGC1α axis serves a central role in regulating
mitochondrial biogenesis, cellular energy homeostasis and stress
responses, rendering it a notable therapeutic target. Numerous
pharmacological agents, including synthetic compounds and natural
phytochemicals, have been studied for their ability to regulate
this pathway and mitigate stroke-induced neuronal damage (77) (Table
II). Rosiglitazone and bezafibrate, for example, activate the
PPAR-PGC1α signaling cascade, leading to increased mitochondrial
mass and performance while mitigating mitochondrial impairment
under ischemic conditions (8).
Quercetin and resveratrol similarly activate SIRT1 and AMPK,
thereby promoting mitochondrial biogenesis and mitigating oxidative
stress (44). Flavonoids, a
category of polyphenolic compounds, have demonstrated marked
potential due to their diverse neuroprotective properties.
Tiliroside, a glycosylated flavonoid, exhibits
inflammation-reducing properties through suppression of microglial
activation and suppressing molecular cascades including p38MAPK,
NF-κB and Nrf2, which are associated with the AMPK/SIRT1/PGC1α axis
(24). Kaempferol is reported to
reduce the production of pro-inflammatory molecules, including
cyclooxygenase-2 (COX-2), TNF-α, prostaglandin E2, IL-6 and
nitrite, in activated BV-2 microglia. Concurrently, it upregulates
the levels of p-AMPK, Nrf2 and HO-1, indicating its potential to
mitigate inflammation associated with stroke (57,78).
Catechin, a flavonoid, attenuates neuroinflammation by decreasing
the expression of inducible nitric oxide synthase and COX-2. It
also reduces microglial generation of ROS and nitric oxide, while
suppressing the secretion of pro-inflammatory cytokines such as
IL-6 and TNF-α. Additionally, catechin enhances AMPK activity and
modulates signaling cascades involved in oxidative stress and
neuroinflammation (79,80). Digitoflavone confers protection to
PC12 cells against ischemia-induced oxidative stress by diminishing
ROS levels, preserving mitochondrial inner membrane integrity,
enhancing AMPK phosphorylation and promoting mitochondrial
biogenesis. Furthermore, it augments catalase activity and
glutathione levels, thereby contributing to neuroprotection
(81,82). Isoquercitrin, a flavonoid, enhances
neuronal resilience to stress under ischemic conditions,
underscoring its potential as a neuroprotective agent (83). Resveratrol, recognized for its
activation of SIRT1, AMPK and PGC1α, demonstrates potential in
mitigating stroke-related injury. Although its metabolic effects
are well-established in peripheral tissues, further investigation
is required to substantiate its direct neuroprotective efficacy in
ischemic stroke models (84).
| Table II.Pharmacokinetic parameters of
neuroprotective flavonoids. |
Table II.
Pharmacokinetic parameters of
neuroprotective flavonoids.
| Compound | Animal dose | Human dose | BBB
penetration | Safety profile,
mg/kg | (Refs.) |
|---|
| Quercetin | 25–100 mg/kg | 500–2,000
mg/kg | Competitive
inhibitor of GLUT1 (IC50 =8.5 µM) binds exofacial site | 0.4±0.07
(rodents) | (47) |
| EGCG | 10–15 mg/kg | 200–1,000
mg/kg | MCT1 (Km=156
µM) | 0.08±0.02 (low
bioavailability) | (105) |
| Luteolin | 5–20 mg/kg (oral
and ip) | 100–400 mg/day | Passive diffusion
due to lipophilicity, readily crosses BBB | 0.51±0.09 | (50) |
| Baicalein | 30–80 mg/kg
(oral) | 600–1,600
mg/kg | LAT1 transporter,
via intranasal solid lipid nanoparticles for enhanced BBB
delivery | 0.35±0.05 | Click or tap here
to enter text. (57) |
| Fisetin | 5–25 mg/kg
(oral) | 80–400 mg/kg | Passive diffusion,
high brain uptake partly p-glycoprotein substrate | 0.45±0.08 | (103) |
Flavonoids such as resveratrol and catechin face
challenges with bioavailability due to rapid metabolism and poor
absorption in the gastrointestinal tract. This leads to lower
bloodstream and brain concentrations, which diminishes their
clinical effectiveness despite promising preclinical results
(85). The challenge of enabling
these compounds to cross the BBB remains a notable hurdle. While
certain flavonoids pass through the BBB, their transport is often
unreliable. This hampers their potential for direct neuroprotection
in ischemic stroke where rapid delivery to the brain is vital.
Clinicals trials involving these compounds have yielded
inconsistent results, attributable to differences in dosing
schedules, formulations and diverse patient populations (53,86,87).
Achieving therapeutic levels in the brain requires high doses that
may not be practical or safe hindering translation from laboratory
research to clinical practice (88).
Although these compounds demonstrate promising
effects, a definitive treatment for ischemic stroke remains
elusive. Current research underscores the potential of combination
therapies to enhance neuroprotection. For instance, the
simultaneous application of flavonoids with interventions such as
exercise or caloric restriction may synergistically activate the
AMPK/SIRT1/PGC1α pathway; however, this remains a potential
strategy that requires further investigation. Furthermore, the
development of BBB-permeable formulations could substantially
improve the delivery and efficacy of these agents in stroke
treatment (89).
The AMPK/SIRT1/PGC1α signaling axis serves an
essential function in maintaining mitochondrial health and
alleviating oxidative stress, two processes that become compromised
in ischemic stroke. Age-associated declines in AMPK activity
further aggravate impairment of mitochondria and oxidative injury,
which in turn weaken stress responses and diminish autophagic
capacity (63). The activation of
AMPK enhances the activity of SIRT1, which subsequently activates
PGC1α, leading to enhanced generation of new mitochondria and
improved cellular resilience in stroke models (90–92).
Furthermore, emerging evidence underscores the beneficial effects
of flavonoids, including quercetin, luteolin and kaempferol, in the
context of ischemic stroke (93).
Quercetin functions as a proteasome inhibitor, mitigating
dysregulated inflammatory responses and enhancing recovery
following a stroke (94). Apigenin
and quercetin mitigate the adverse effects of 7-ketocholesterol in
neuronal cells by preserving mitochondrial function and modulating
the expression of AMPK, SIRT1 and PGC1α (95). Thus, both pharmacological and
non-pharmacological approaches targeting the AMPK/SIRT1/PGC1α
pathway holds promise for neuroprotection in ischemic stroke and
their efficacy may be further enhanced through combination
therapies and advanced delivery systems. Continued research is
essential to refine these strategies for clinical application.
Flavonoid pharmacokinetics and BBB
permeability
Preclinical investigations have consistently
demonstrated the neuroprotective properties of flavonoids (93); however, their clinical application
is hindered by marked pharmacokinetic challenges. When administered
orally, flavonoids typically exhibit an oral bioavailability of
<5%, primarily due to considerable presystemic metabolic
processes, such as glucuronidation, sulfation and methylation by
hepatic and intestinal enzymes (96). Their translocation across the BBB
is further constrained by their physicochemical properties, with
only those possessing a molecular weight of <500 kDa and
moderate lipophilicity (logP2-3) capable of limited passive
diffusion across the BBB (97).
Glycosylated flavonoids such as rutin and hesperidin, require
conversion to their active aglycone forms via enzymatic hydrolysis
by gut microbiota β-glucosidase. Certain flavonoids employ active
transport mechanisms. Catechins utilize monocarboxylate
transporters [Km, 156 µM for epigallocatechin-3-gallate (EGCG)],
quercetin and fisetin utilize GLUT1 (Km, 18.7 µM) and specific
flavonoid metal complexes are transported via transferrin receptors
(98). Nevertheless, efflux
systems, such as P-glycoprotein, breast cancer resistance protein
and multidrug resistance-associated proteins, actively limit their
cellular accumulation and retention within the central nervous
system (94). Advancements in drug
delivery have shown potential for overcoming these barriers.
Liposomal formulations such as quercetin polyethylene glycol
liposomes have enhanced bioavailability by 3- to 5-fold,
nanocrystals (such as baicalein-polyvinylpyrrolidone) improve
dissolution and reduce effective doses by 50–80%, and prodrug
strategies targeting LAT1 amino acid transporters have demonstrated
increased brain uptake (99).
Despite these advancements clinical applications necessitate
careful consideration of dosage and safety issues. For instance,
quercetin doses of 25–100 mg/kg in animals correspond to 500–2,000
mg/day in humans, achieving plasma concentrations of 5–10 µM with a
half-life of 3–5 h. However, doses exceeding 1 g (1,000 mg) may
induce headaches (Table II)
(100). Similarly, EGCG is
generally safe at human equivalent doses of 200–1,000 mg per day,
however, high doses such as those of >8,000 mg per day have been
associated with elevated liver enzymes indicating potential
hepatotoxicity (Table II)
(101). Other flavonoids, such as
baicalein and luteolin, possess more favorable safety profiles but
may cause gastrointestinal disturbances at higher doses than
quercetin (Table II). Notably,
most flavonoids exhibit U-shaped dose response curves and chronic
high doses may result in adverse effects or drug interactions due
to the modulation of CYP3A4 and P-glycoprotein. Nanoencapsulation
strategies not only enhance efficacy but also reduce the required
doses while maintaining brain exposure (90). Overall, these pharmacokinetics
insights underscore the need for optimized delivery systems and
rational dosing strategies to completely realize the therapeutic
promise of flavonoids in managing cerebral ischemia and other
neurological disorders.
Future insights
Despite encouraging preclinical results, key
knowledge gaps remain before the SIRT1/AMPK/PGC1α axis can be
translated into stroke therapy. Most studies use young rodent
models with short observation windows, whereas clinical stroke
occurs in older patients with comorbidities (such as hypertension
and diabetes) (102). Pathway
effects are cell and phase specific; AMPK and autophagy can be
protective or detrimental depending on context. Direct evidence
linking pathway activation to sustained functional recovery remains
limited, and human evidence is sparse. Serum SIRT1 levels in
patients with stroke show no association with clinical outcomes,
highlighting the gap between experimental promise and clinical
utility (61). Recent advances
have identified novel brain specific isoforms of PGC1α in human
neural tissue, regulated by CNS-specific promoters located ~500
kilobases upstream of the canonical promoter (24). Among these, a truncated 17 kDa
isoform has garnered attention for its potential role in
suppressing full length PGC1α, thereby contributing to stroke
pathology (25). However, while
these findings are promising, they remain at a preclinical and
mechanistic stage. Further research is required to elucidate the
physiological and pathological functions of these isoforms before
they can be translated into therapeutic targets for ischemic
stroke.
Epigenetic modulation of PGC1α via DNA methylation
and nucleosome positioning also presents a compelling therapeutic
concept (103). DNA
methyltransferases (3A and 3B) influence these epigenetic marks,
which are associated with mitochondrial dysfunction and oxidative
stress (104). Although these
mechanisms have been demonstrated in experimental settings,
clinical application remains exploratory. Targeting epigenetic
regulators could offer novel stroke therapies; however, such
approaches require validation in translational models.
By contrast, modulation of the SIRT1/AMPK/PGC1α
pathway through existing pharmacological agents offers more
immediate translational potential. Agents such as flavonoids
(quercetin) and metabolic regulators (resveratrol or metformin)
(105) have demonstrated
neuroprotective effects and are currently under investigation in
clinical and preclinical contexts (106). Their combination with
non-pharmacological strategies such as exercise and caloric
restriction presents a feasible, multi model approach to enhancing
mitochondrial biogenesis and neuroprotection in stroke recovery
(24).
Similarly, PPARγ agonists, known to activate PGC1α,
have demonstrated effectiveness in enhancing mitochondrial
performance and lowering oxidative damage (107). However, their clinical utility is
limited by challenges such as poor BBB penetration during stroke,
adverse side effects (for example, anemia and edema) and narrow
therapeutic windows. Therefore, while these agents are closer to
clinical use than isoform targeting or epigenetic therapies,
further optimization of BBB permeable formulations and dosing
strategies is critical.
Future research should prioritize strategies with
high translational potential such as enhancing PGC1α activation
through well characterized pharmacological and lifestyle
intervention. Parallel exploration of emerging but less clinically
validated areas such as isoform-specific regulation and epigenetic
targeting may open novel therapeutic avenues in the longer term.
Integrating these approaches can improve stroke outcomes and
broaden the therapeutic landscape for neurodegeneration.
Conclusion
The SIRT1/AMPK/PGC1α signaling pathway constitutes a
viable therapeutic target for cerebral ischemia. Nevertheless, the
intricate nature of stroke pathophysiology necessitates the
development of multi-targeted treatment strategies for effective
intervention. The synergism between phytochemicals and
non-pharmacological interventions such as exercise and caloric
restriction has shown promise in targeting the essential mechanisms
in stroke such as mitochondrial dysfunction, oxidative stress and
neuroinflammation (108). These
approaches may also reduce age related impairments in cognition,
the nervous system and memory, potentially through modulation of
the SIRT1/AMPK/PGC1α pathway. Advances in the base formulations and
drug delivery methods of flavonoids, including the nano
formulations able to traverse the BBB, offer new hopes for
neuroprotective interventions. In addition, the possible
synergistic effects combining phytochemicals with an exercise
modality, such as yoga, warrant further evaluation as they may
offer potential benefits in terms of reduced toxicity and treatment
costs, which could be explored for future prevention and
intervention strategies.
In summary, the present review provides insights
into the therapeutic benefits of activating the SIRT1/AMPK/PGC1α
signaling axis in ischemic stroke and emphasizes that
multi-targeted strategies, including pharmacological interventions
and non-pharmacological interventions, are an essential factor to
modulate mediators for neuroprotection. It also highlights the
potential therapeutic effect of brain PGC1α isoforms, BBB-permeable
delivery systems and epigenetic regulation as promising therapeutic
targets (109). Through targeting
several pathological mechanisms, these new development strategies
provide hopeful therapeutic avenues of improving patient outcome
and reducing the global burden of ischemic stroke. In order to
further enhance the application of stroke therapy new adjuvant drug
delivery systems capable of enhancing BBB penetration and flavonoid
bioavailability need to be developed. In addition, the
brain-specific PGC1α isoforms need to be addressed for therapeutic
precision. It would be necessary to validate efficacy by performing
clinical trials using standardized dosing regimens, formulations
and patient classification.
Acknowledgements
Not applicable.
Funding
The present review was funded by the National Natural Science
Foundation of China (grant nos. 82472164 and 82272163) and
Mechanism of Curcumin Regulating Mitophagy in the Brain of a Mouse
Model of Depression-General Project of Zhejiang Provincial Health
Commission (grant no. 2023KY1014).
Availability of data and materials
Not applicable.
Authors' contributions
INA made substantial contributions to the conception
and design of the review, drafted the initial manuscript,
critically revised it for important intellectual content, approved
the final version, and agrees to be accountable for all aspects of
the work. NA made substantial contributions to the acquisition,
analysis, and interpretation of literature data (including doses,
outcomes, and mechanisms), participated in drafting, editing, and
reviewing the manuscript, approved the final version, and agrees to
be accountable for the accuracy and integrity of the data
extracted. QX made substantial contributions to the critical
evaluation and interpretation of the literature, played a key role
in synthesizing and organizing the findings into a coherent
narrative, critically revised the manuscript for important
intellectual content, approved the final version, and agrees to be
accountable for the integrity of the literature synthesis. ABH made
substantial contributions to the acquisition of data by
systematically searching for relevant papers, collecting and
managing all references, participated in interpreting the collected
data, critically reviewed the manuscript, approved the final
version, and agrees to be accountable for the accuracy and
completeness of the reference management and data collection. FW
made substantial contributions to the visualization and
interpretation of data by preparing all figures and tables, which
required intellectual input to accurately represent the signaling
pathways; contributed to the conceptualization; helped revise
figure legends and the manuscript; approved the final version; and
agrees to be accountable for the accuracy and integrity of the
visual content. XY made substantial contributions to the
acquisition and screening of literature, determined which papers to
include based on predefined criteria, helped structure and arrange
the content into different sections, participated in the
interpretation of the selected studies, critically revised the
manuscript, approved the final version, and agrees to be
accountable for the integrity of the literature selection and
section organization. YY made substantial contributions to the
literature search and organization of references, participated in
writing and editing portions of the manuscript, critically reviewed
the content, approved the final version, and agrees to be
accountable for the accuracy of the references and contributed
sections. SY, MF and YJ made substantial contributions to the
conception and design of the review, provided project
administration and supervision, acquired funding, critically
revised the manuscript for important intellectual content, approved
the final version, and agree to be accountable for all aspects of
the work. Data authentication is not applicable. All authors have
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
González-Santos J, Rodríguez-Fernández P,
Pardo-Hernández R, González-Bernal JJ, Fernández-Solana J and
Santamaría-Peláez M: A cross-sectional study: Determining factors
of functional independence and quality of life of patients one
month after having suffered a stroke. Int J Environ Res Public
Health. 20:9952023. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pu L, Wang L, Zhang R, Zhao T, Jiang Y and
Han L: Projected global trends in ischemic stroke incidence, deaths
and disability-adjusted life years from 2020 to 2030. Stroke.
54:1330–1339. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Salaudeen MA, Bello N, Danraka RN and
Ammani ML: Understanding the pathophysiology of ischemic stroke:
The basis of current therapies and opportunity for new ones.
Biomolecules. 14:3052024. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen Y, Peng F, Yang C, Hou H, Xing Z,
Chen J, Liu L, Peng C and Li D: SIRT1 activation by
2,3,5,6-tetramethylpyrazine alleviates neuroinflammation via
inhibiting M1 microglia polarization. Front Immunol.
14:12065132023. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhao J, Deng H, Xun C, Chen C, Hu Z, Ge L
and Jiang Z: Therapeutic potential of stem cell extracellular
vesicles for ischemic stroke in preclinical rodent models: A
meta-analysis. Stem Cell Res Ther. 14:622023. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Salman M, Stayton AS, Parveen K, Parveen
A, Puchowicz MA, Parvez S, Bajwa A and Ishrat T: Intranasal
delivery of mitochondria attenuates brain injury by AMPK and
SIRT1/PGC-1α pathways in a murine model of photothrombotic stroke.
Mol Neurobiol. 61:2822–2838. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gao J, Qian T and Wang W: CTRP3 activates
the AMPK/SIRT1-PGC-1α pathway to protect mitochondrial biogenesis
and functions in cerebral ischemic stroke. Neurochem Res.
45:3045–3058. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhou Y, Wang S, Li Y, Yu S and Zhao Y:
SIRT1/PGC-1α signaling promotes mitochondrial functional recovery
and reduces apoptosis after intracerebral hemorrhage in rats. Front
Mol Neurosci. 10:4432018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Han B, Jiang W, Cui P, Zheng K, Dang C,
Wang J, Li H, Chen L, Zhang R, Wang QM, et al: Microglial PGC-1α
protects against ischemic brain injury by suppressing
neuroinflammation. Genome Med. 13:472021. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yuan Y, Tian Y, Jiang H, Cai L, Song J,
Peng R and Zhang X: Mechanism of PGC-1α-mediated mitochondrial
biogenesis in cerebral ischemia-reperfusion injury. Front Mol
Neurosci. 16:12249642023. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bai S, Geng X, Li F and Ding Y: Abstract
TP283: Histone lactylation and neuronal autophagy: Insights into
the penumbra-core transition and potential biomarker in ischemic
stroke. Stroke 55:. (Suppl 1):ATP2832024.
|
|
12
|
Bonomi RE, Riordan W and Gelovani JG: The
structures, functions, and roles of class III HDACs (Sirtuins) in
neuropsychiatric diseases. Cells. 13:16442024. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu QJ, Zhang TN, Chen HH, Yu XF, Lv JL,
Liu YY, Liu YS, Zheng G, Zhao JQ, Wei YF, et al: The sirtuin family
in health and disease. Signal Transduct Target Ther. 7:4022022.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sarvarian P, Samadi P, Gholipour E, Shams
Asenjan K, Hojjat-Farsangi M, Motavalli R, Motavalli Khiavi F and
Yousefi M: Application of emerging plant-derived nanoparticles as a
novel approach for nano-drug delivery systems. Immunol Invest.
51:1039–1059. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Qian L, Zhu Y, Deng C, Liang Z, Chen J,
Chen Y, Wang X, Liu Y, Tian Y and Yang Y: Peroxisome
proliferator-activated receptor gamma coactivator-1 (PGC-1) family
in physiological and pathophysiological process and diseases.
Signal Transduct Target Ther. 9:502024. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhao Y, Zhang Y, Zhang J, Zhang X and Yang
G: Molecular mechanism of autophagy: Its role in the therapy of
Alzheimer's disease. Curr Neuropharmacol. 18:720–739. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen M, Tan J, Jin Z, Jiang T, Wu J and Yu
X: Research progress on Sirtuins (SIRTs) family modulators. Biomed
Pharmacother. 174:1164812024. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Marin TL, Gongol B, Zhang F, Martin M,
Johnson DA, Xiao H, Wang Y, Subramaniam S, Chien S and Shyy JY:
AMPK promotes mitochondrial biogenesis and function by
phosphorylating the epigenetic factors DNMT1, RBBP7, and HAT1. Sci
Signal. 10:eaaf74782017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Herzig S and Shaw RJ: AMPK: Guardian of
metabolism and mitochondrial homeostasis. Nat Rev Mol Cell Biol.
19:121–135. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shin YJ, Evitts KM, Jin S, Howard C,
Sharp-Milgrom M, Schwarze-Taufiq T, Kinoshita C, Young JE and Zheng
Y: Amyloid beta peptides (Aβ) from Alzheimer's disease neuronal
secretome induce endothelial activation in a human cerebral
microvessel model. Neurobiol Dis. 181:1061252023. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Adhikary A, Mukherjee A, Banerjee R and
Nagotu S: DRP1: At the crossroads of dysregulated mitochondrial
dynamics and altered cell signaling in cancer cells. ACS Omega.
8:45208–45223. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mishra P, Mittal AK, Kalonia H, Madan S,
Ghosh S, Sinha JK and Rajput SK: SIRT1 promotes neuronal
fortification in neurodegenerative diseases through attenuation of
pathological hallmarks and enhancement of cellular lifespan. Curr
Neuropharmacol. 19:1019–1037. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Abu Shelbayeh O, Arroum T, Morris S and
Busch KB: PGC-1α is a master regulator of mitochondrial lifecycle
and ROS stress response. Antioxidants (Basel). 12:10752023.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rakshe PS, Dutta BJ, Chib S, Maurya N and
Singh S: Unveiling the interplay of AMPK/SIRT1/PGC-1α axis in brain
health: Promising targets against aging and NDDs. Ageing Res Rev.
96:1022552024. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Panes J, Wendt A, Ramirez-Molina O, Castro
P and Fuentealba J: Deciphering the role of PGC-1α in neurological
disorders: From mitochondrial dysfunction to synaptic failure.
Neural Regen Res. 17:237–245. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Guo J, Huang X, Dou L, Yan M, Shen T, Tang
W and Li J: Aging and aging-related diseases: from molecular
mechanisms to interventions and treatments. Signal Transduct Target
Ther. 7:3912022. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Soyal SM, Zara G, Ferger B, Felder TK,
Kwik M, Nofziger C, Dossena S, Schwienbacher C, Hicks AA,
Pramstaller PP, et al: The PPARGC1A locus and CNS-specific PGC-1α
isoforms are associated with Parkinson's disease. Neurobiol Dis.
121:34–46. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Luo X, Liao C, Quan J, Cheng C, Zhao X,
Bode AM and Cao Y: Posttranslational regulation of PGC-1α and its
implication in cancer metabolism. Int J Cancer. 145:1475–1483.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fang Y, Wang X, Yang D, Lu Y, Wei G, Yu W,
Liu X, Zheng Q, Ying J and Hua F: Relieving cellular energy stress
in aging, neurodegenerative, and metabolic diseases, SIRT1 as a
therapeutic and promising node. Front Aging Neurosci.
13:7386862021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xie N, Zhang L, Gao W, Huang C, Huber PE,
Zhou X, Li C, Shen G and Zou B: NAD+ metabolism:
Pathophysiologic mechanisms and therapeutic potential. Signal
Transduct Target Ther. 5:2272020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Guan G, Chen Y and Dong Y: Unraveling the
AMPK-SIRT1-FOXO pathway: The in-depth analysis and breakthrough
prospects of oxidative stress-induced diseases. Antioxidants
(Basel). 14:702025. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sharma A, Anand SK, Singh N, Dwarkanath A,
Dwivedi UN and Kakkar P: Berbamine induced activation of the
SIRT1/LKB1/AMPK signaling axis attenuates the development of
hepatic steatosis in high-fat diet-induced NAFLD rats. Food Funct.
12:892–909. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang X, Lu Y, Tuo Z, Zhou H, Zhang Y, Cao
Z, Peng L, Yu D and Bi L: Role of SIRT1/AMPK signaling in the
proliferation, migration and invasion of renal cell carcinoma
cells. Oncol Rep. 45:1092021. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mosaoa RM, Al-Rabia MW, Asfour HZ,
Alhakamy NA, Mansouri RA, El-Agamy DS, Abdulaal WH, Mohamed GA,
Ibrahim SRM and Elshal M: Targeting SIRT1/AMPK/Nrf2/NF-кB by
sitagliptin protects against oxidative stress-mediated ER stress
and inflammation during ANIT-induced cholestatic liver injury.
Toxicology. 507:1538892024. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jiao F and Gong Z: The beneficial roles of
SIRT1 in neuroinflammation-related diseases. Oxid Med Cell Longev.
2020:67828722020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cetrullo S, D'Adamo S, Tantini B, Borzi RM
and Flamigni F: mTOR, AMPK, and Sirt1: Key players in metabolic
stress management. Crit Rev Eukaryot Gene Expr. 25:59–75. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jin X, Zhu L, Lu S, Li C, Bai M, Xu E,
Shen J and Li Y: Baicalin ameliorates CUMS-induced depression-like
behaviors through activating AMPK/PGC-1α pathway and enhancing
NIX-mediated mitophagy in mice. Eur J Pharmacol. 938:1754352023.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang Y, Wu Y, Meng X, Wang Z, Younis M,
Liu Y, Wang P and Huang X: SARS-CoV-2 membrane protein causes the
mitochondrial apoptosis and pulmonary edema via targeting BOK. Cell
Death Differ. 29:1395–1408. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lu C, Zhao H, Liu Y, Yang Z, Yao H, Liu T,
Gou T, Wang L, Zhang J, Tian Y, et al: Novel role of the SIRT1 in
endocrine and metabolic diseases. Int J Biol Sci. 19:484–501. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Peng Z, Ru D, Leng G, Peng J, Zhang M and
Cai B: Apelin-13 enhances neurofunctional recovery and suppresses
neuroinflammation via the SIRT1/NF-κB axis in ischemic stroke. Cell
Immunol. 413:1049582025. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang BB, Zhou G and Li C: AMPK: An
emerging drug target for diabetes and the metabolic syndrome. Cell
Metab. 9:407–416. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Olivier S, Foretz M and Viollet B: Promise
and challenges for direct small molecule AMPK activators. Biochem
Pharmacol. 153:147–158. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jin T, Zhang Y, Botchway BOA, Huang M, Lu
Q and Liu X: Quercetin activates the Sestrin2/AMPK/SIRT1 axis to
improve amyotrophic lateral sclerosis. Biomed Pharmacother.
161:1145152023. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Steinberg GR and Carling D: AMP-activated
protein kinase: The current landscape for drug development. Nat Rev
Drug Discov. 18:527–551. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Szewczuk M, Boguszewska K,
Kaźmierczak-Barańska J and Karwowski BT: The role of AMPK in
metabolism and its influence on DNA damage repair. Mol Biol Rep.
47:9075–9086. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bhutta M, Gallo E and Borenstein R:
Multifaceted role of AMPK in viral infections. Cells. 10:11182021.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kim SG, Kim JR and Choi HC:
Quercetin-induced AMP-activated protein kinase activation
attenuates vasoconstriction through LKB1-AMPK signaling pathway. J
Med Food. 21:146–153. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Li Z, Guo H, Li J, Ma T, Zhou S, Zhang Z,
Miao L and Cai L: Sulforaphane prevents type 2 diabetes-induced
nephropathy via AMPK-mediated activation of lipid metabolic
pathways and Nrf2 antioxidative function. Clin Sci (Lond).
134:2469–2487. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Muraleedharan R and Dasgupta B: AMPK in
the brain: Its roles in glucose and neural metabolism. FEBS J.
289:2247–2262. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Singh NK, Bhushan B, Singh P and Sahu KK:
Therapeutic expedition of luteolin against brain-related disorders:
An updated review. Comb Chem High Throughput Screen. 28:371–391.
2025. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Chen L, Qin Y, Liu B, Gao M, Li A, Li X
and Gong G: PGC-1 α-mediated mitochondrial quality control:
Molecular mechanisms and implications for heart failure. Front Cell
Dev Biol. 10:8713572022. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Lambona C, Zwergel C, Valente S and Mai A:
SIRT3 activation a promise in drug development? New insights into
SIRT3 biology and its implications on the drug discovery process. J
Med Chem. 67:1662–1689. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Choi HI, Kim HJ, Park JS, Kim IJ, Bae EH,
Ma SK and Kim SW: PGC-1α attenuates hydrogen peroxide-induced
apoptotic cell death by upregulating Nrf-2 via GSK3β inactivation
mediated by activated p38 in HK-2 cells. Sci Rep. 7:43192017.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Liu N, Lyu X, Zhang X, Zhang F, Chen Y and
Li G: Astaxanthin attenuates cognitive deficits in Alzheimer's
disease models by reducing oxidative stress via the SIRT1/PGC-1α
signaling pathway. Cell Biosci. 13:1732023. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Rahmani S, Roohbakhsh A, Pourbarkhordar V
and Karimi G: The cardiovascular protective function of natural
compounds through AMPK/SIRT1/PGC-1α signaling pathway. Food Sci
Nutr. 12:9998–10009. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Elbaz EM, Senousy MA, El-Tanbouly DM and
Sayed RH: Neuroprotective effect of linagliptin against
cuprizone-induced demyelination and behavioural dysfunction in
mice: A pivotal role of AMPK/SIRT1 and JAK2/STAT3/NF-κB signalling
pathway modulation. Toxicol Appl Pharmacol. 352:153–161. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Cheng X, Yang YL, Yang H, Wang YH and Du
GH: Kaempferol alleviates LPS-induced neuroinflammation and BBB
dysfunction in mice via inhibiting HMGB1 release and
down-regulating TLR4/MyD88 pathway. Int Immunopharmacol. 56:29–35.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Peng J, Yang Z, Li H, Hao B, Cui D, Shang
R, Lv Y, Liu Y, Pu W, Zhang H, et al: Quercetin reprograms
immunometabolism of macrophages via the SIRT1/PGC-1α signaling
pathway to ameliorate lipopolysaccharide-induced oxidative damage.
Int J Mol Sci. 24:55422023. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Shakova FM, Kirova YI, Silachev DN,
Romanova GA and Morozov SG: Protective effects of PGC-1α activators
on ischemic stroke in a rat model of photochemically induced
thrombosis. Brain Sci. 11:3252021. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yang L, Qian J, Yang B, He Q, Wang J and
Weng Q: Challenges and improvements of novel therapies for ischemic
stroke. Front Pharmacol. 12:7211562021. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Hernández-Jiménez M, Hurtado O, Cuartero
MI, Ballesteros I, Moraga A, Pradillo JM, McBurney MW, Lizasoain I
and Moro MA: Silent information regulator 1 protects the brain
against cerebral ischemic damage. Stroke. 44:2333–2337. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Zhao B, Zhang Q, Liang X, Xie J and Sun Q:
Quercetin reduces inflammation in a rat model of diabetic
peripheral neuropathy by regulating the TLR4/MyD88/NF-κB signalling
pathway. Eur J Pharmacol. 912:1746072021. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Cantó C and Auwerx J: PGC-1alpha, SIRT1
and AMPK, an energy sensing network that controls energy
expenditure. Curr Opin Lipidol. 20:98–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Zhou Z, Li W, Ni L, Wang T, Huang Y, Yu Y,
Hu M, Liu Y, Wang J, Huang X and Wang Y: Icariin improves oxidative
stress injury during ischemic stroke via inhibiting mPTP opening.
Mol Med. 30:772024. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Cheng M, Yuan C, Ju Y, Liu Y, Shi B, Yang
Y, Jin S, He X, Zhang L and Min D: Quercetin attenuates oxidative
stress and apoptosis in brain tissue of APP/PS1 double transgenic
AD mice by regulating Keap1/Nrf2/HO-1 pathway to improve cognitive
impairment. Behav Neurol. 2024:56981192024. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
de Gregorio E, Colell A, Morales A and
Marí M: Relevance of SIRT1-NF-κB axis as therapeutic target to
ameliorate inflammation in liver disease. Int J Mol Sci.
21:38582020. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Uzdensky A and Demyanenko S: Histone
acetylation and deacetylation in ischemic stroke. Neural Regen Res.
16:1529–1530. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Liu Y, Zhu C, Guo J, Chen Y and Meng C:
The neuroprotective effect of irisin in ischemic stroke. Front
Aging Neurosci. 12:5889582020. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Wang X, Zhang M, Feng R, Li WB, Ren SQ and
Zhang F: Exercise pre-conditioning alleviates brain damage via
excitatory amino acid transporter 2 and extracellular
signal-regulated kinase 1/2 following ischemic stroke in rats. Mol
Med Rep. 11:1523–1527. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Zhang H, Xie Q and Hu J: Neuroprotective
effect of physical activity in ischemic stroke: Focus on the
neurovascular unit. Front Cell Neurosci. 16:8605732022. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Li F, Geng X, Lee H, Wills M and Ding Y:
Neuroprotective effects of exercise postconditioning after stroke
via SIRT1-mediated suppression of endoplasmic reticulum (ER)
stress. Front Cell Neurosci. 15:5982302021. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Buvarp D, Viktorisson A, Axelsson F, Lehto
E, Lindgren L, Lundström E and Sunnerhagen KS: Physical activity
trajectories and functional recovery after acute stroke among
adults in Sweden. JAMA Netw Open. 6:e23109192023. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Rashad S, Saigusa D, Yamazaki T, Matsumoto
Y, Tomioka Y, Saito R, Uruno A, Niizuma K, Yamamoto M and Tominaga
T: Metabolic basis of neuronal vulnerability to ischemia; an in
vivo untargeted metabolomics approach. Sci Rep. 10:65072020.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Chen M, Liu J, Wu W, Guo T, Yuan J, Wu Z,
Zheng Z, Zhao Z, Lin Q, Liu N and Chen H: SIRT1 restores
mitochondrial structure and function in rats by activating SIRT3
after cerebral ischemia/reperfusion injury. Cell Biol Toxicol.
40:312024. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Rius-Pérez S, Torres-Cuevas I, Millán I,
Ortega ÁL and Pérez S: PGC-1 α, inflammation, and oxidative stress:
An integrative view in metabolism. Oxid Med Cell Longev.
2020:14526962020. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Wen Y, Hu Z, Tang X and Hu K:
Overexpression of PGC-1α reduces inflammation and protects against
focal cerebral ischaemia/reperfusion injury. Arch Med Sci.
16:2020.
|
|
77
|
Chen J, Liu B, Yao X, Yang X, Sun J, Yi J,
Xue F, Zhang J, Shen Y, Chen B and Sun H: AMPK/SIRT1/PGC-1α
signaling pathway: Molecular mechanisms and targeted strategies
from energy homeostasis regulation to disease therapy. CNS Neurosci
Ther. 31:e706572025. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Li WH, Cheng X, Yang YL, Liu M, Zhang SS,
Wang YH and Du GH: Kaempferol attenuates neuroinflammation and
blood brain barrier dysfunction to improve neurological deficits in
cerebral ischemia/reperfusion rats. Brain Res. 1722:1463612019.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Kim J, Yang G, Kim Y, Kim J and Ha J: AMPK
activators: Mechanisms of action and physiological activities. Exp
Mol Med. 48:e2242016. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Li S, Wang Z, Liu G and Chen M:
Neurodegenerative diseases and catechins:
(−)-epigallocatechin-3-gallate is a modulator of chronic
neuroinflammation and oxidative stress. Front Nutr. 11:14258392024.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Zhang J, Jiang H, Wu F, Chi X, Pang Y, Jin
H, Sun Y and Zhang S: Neuroprotective effects of hesperetin in
regulating microglia polarization after ischemic stroke by
inhibiting TLR4/NF-κB pathway. J Healthc Eng. 2021:99388742021.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Mu Q, Zhang Y, Cheng Q, Huang H, Huang C
and Tang L: Research progress on the mechanism of action of
hesperetin in cerebral ischemia: A narrative review. Ann Transl
Med. 10:8062022. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Shi Y, Chen X, Liu J, Fan X, Jin Y, Gu J,
Liang J, Liang X and Wang C: Isoquercetin improves inflammatory
response in rats following ischemic stroke. Front Neurosci.
15:5555432021. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Fan L, Lin X, Hong L, Li L, Lin R, Ren T,
Tian J and Chen M: Simultaneous antioxidant and neuroprotective
effects of two-dimensional (2D) MXene-loaded isoquercetin for
ischemic stroke treatment. J Mater Chem B. 12:2795–2806. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Dong H, Gao X, Li H, Gao J and Zhang L:
Protective effects of flavonoids against intracerebral and
subarachnoid hemorrhage (review). Exp Ther Med. 28:3502024.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Lan F, Weikel K, Cacicedo J and Ido Y:
Resveratrol-induced AMP-activated protein kinase activation is
cell-type dependent: Lessons from basic research for clinical
application. Nutrients. 9:7512017. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Morand C: How to better consider and
understand interindividual variability in response to polyphenols
in clinical trials. Front Nutr. 11:15225162024. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Baqer SH, Al-Shawi SG and Al-Younis ZK:
Quercetin, the potential powerful flavonoid for human and food: A
review. Front Biosci (Elite Ed). 16:302024. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Guo K and Lu Y: Acupuncture modulates the
AMPK/PGC-1 signaling pathway to facilitate mitochondrial biogenesis
and neural recovery in ischemic stroke rats. Front Mol Neurosci.
17:13887592024. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Dong-Chen X, Yong C, Yang X, Chen-Yu S and
Li-Hua P: Signaling pathways in Parkinson's disease: Molecular
mechanisms and therapeutic interventions. Signal Transduct Target
Ther. 8:732023. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Jiang S, Li T, Ji T, Yi W, Yang Z, Wang S,
Yang Y and Gu C: AMPK: Potential therapeutic target for ischemic
stroke. Theranostics. 8:4535–4551. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Giovarelli M, Zecchini S, Casati SR,
Lociuro L, Gjana O, Mollica L, Pisanu E, Mbissam HD, Cappellari O,
De Santis C, et al: The SIRT1 activator SRT2104 exerts exercise
mimetic effects and promotes Duchenne muscular dystrophy recovery.
Cell Death Dis. 16:2592025. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Alexander C, Parsaee A and Vasefi M:
Polyherbal and multimodal treatments: Kaempferol- and
quercetin-rich herbs alleviate symptoms of Alzheimer's disease.
Biology (Basel). 12:14532023.PubMed/NCBI
|
|
94
|
Guo C, Wang WJ, Liao YC, Zhao C, Yin Y,
Yao MN, Ding Y and Wang JW: Effect and mechanisms of quercetin for
experimental focal cerebral ischemia: A systematic review and
meta-analysis. Oxid Med Cell Longev. 2022:97494612022. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Ardianto C, Lestari D, Primadani LH,
Puspitasari DR, Sumartha INB, Nisak K, Budiatin AS, Shinta DW,
Andarsari MR, Ifadotunnikmah F, et al: Quercetin exerts a
protective effect on ischemic stroke-induced memory deficits in
mice. J Pharmacol Pharmacother. 14:133–141. 2023. View Article : Google Scholar
|
|
96
|
Zhang N, Zhang S and Dong X: Plant-derived
bioactive compounds and their novel role in central nervous system
disorder treatment via ATF4 targeting: A systematic literature
review. Biomed Pharmacother. 176:1168112024. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Kadry H, Noorani B and Cucullo L: A
blood-brain barrier overview on structure, function, impairment,
and biomarkers of integrity. Fluids Barriers CNS. 17:692020.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Pan L, Ye H, Pi X, Liu W, Wang Z, Zhang Y
and Zheng J: Effects of several flavonoids on human gut microbiota
and its metabolism by in vitro simulated fermentation. Front
Microbiol. 14:10927292023. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Goebel J, Chmielewski J and Hrycyna CA:
The roles of the human ATP-binding cassette transporters
P-glycoprotein and ABCG2 in multidrug resistance in cancer and at
endogenous sites: Future opportunities for structure-based drug
design of inhibitors. Cancer Drug Resist. 4:784–804.
2021.PubMed/NCBI
|
|
100
|
Yao P, Li Y, Yang Y, Yu S and Chen Y:
Triptolide improves cognitive dysfunction in rats with vascular
dementia by activating the SIRT1/PGC-1α signaling pathway.
Neurochem Res. 44:1977–1985. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Han MK, Barreto TA, Martinez FJ, Comstock
AT and Sajjan US: Randomised clinical trial to determine the safety
of quercetin supplementation in patients with chronic obstructive
pulmonary disease. BMJ Open Respir Res. 7:e0003922020. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Sui H, Zhu L, Zhan L, Bi T and Zhang B:
ZiBuPiYin recipe ameliorates diabetes-associated cognitive decline
by improving neuronal mitochondrial function in chronic
psychologically stressed zucker diabetic fatty rats. J
Ethnopharmacol. 302:1159472023. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Yang H, Hong Y, Gong M, Cai S, Yuan Z,
Feng S, Chen Q, Liu X and Mei Z: Fisetin exerts neuroprotective
effects in vivo and in vitro by inhibiting ferroptosis and
oxidative stress after traumatic brain injury. Front Pharmacol.
15:14803452024. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Yang X, Xu S, Qian Y, He X, Chen S and
Xiao Q: Hypermethylation of the gene coding for PGC-1α in
peripheral blood leukocytes of patients with Parkinson's disease.
Front Neurosci. 14:972020. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Zhou F, Deng S, Luo Y, Liu Z and Liu C:
Research progress on the protective effect of green tea polyphenol
(−)-epigallocatechin-3-gallate (EGCG) on the liver. Nutrients.
17:11012025. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Liu S, Su Y, Sun B, Hao R, Pan S, Gao X,
Dong X, Ismail AM and Han B: Luteolin protects against CIRI,
potentially via regulation of the SIRT3/AMPK/mTOR signaling
pathway. Neurochem Res. 45:2499–2515. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Behl T, Madaan P, Sehgal A, Singh S,
Sharma N, Bhatia S, Al-Harrasi A, Chigurupati S, Alrashdi I and
Bungau SG: Elucidating the neuroprotective role of PPARs in
Parkinson's disease: A neoteric and prospective target. Int J Mol
Sci. 22:101612021. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Kim SG, Sung JY, Kim JR and Choi HC:
Quercetin-induced apoptosis ameliorates vascular smooth muscle cell
senescence through AMP-activated protein kinase signaling pathway.
Korean J Physiol Pharmacol. 24:69–79. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Zhang C, Li C, Chen S, Li Z, Ma L, Jia X,
Wang K, Bao J, Liang Y, Chen M, et al: Hormetic effect of
panaxatriol saponins confers neuroprotection in PC12 cells and
zebrafish through PI3K/AKT/mTOR and AMPK/SIRT1/FOXO3 pathways. Sci
Rep. 7:410822017. View Article : Google Scholar : PubMed/NCBI
|