Introduction
Heart failure (HF) with preserved ejection fraction
(HFpEF) is currently the predominant form of HF and represents a
major public health challenge (1).
The incidence and prevalence of HFpEF continue to rise with
increasing age and the increasing burden of obesity and sedentary
and cardiometabolic disease (2).
The diagnosis of HFpEF involves the assessment of cardiac
dysfunction based on imaging techniques and assessment of increased
left ventricular filling pressure (3). Although HFpEF accounts for >50% of
all HF cases, the underlying mechanisms driving its pathogenesis
remain unclear, thereby impeding the development and translation of
mechanism-based therapies (4).
Current treatments for HFpEF are largely restricted to alleviating
congestion through diuretics, encouraging a healthy active
lifestyle and managing comorbidities, as there are few effective
treatments (5). Therefore, further
exploration of the pathogenic mechanisms underlying HFpEF and the
development of novel, effective therapeutic approaches are key.
Pyroptosis is an inflammatory form of programmed
cell death mediated by Caspase-1, which differs markedly from
classical apoptosis in both morphological and functional
characteristics (6). Pathogen
infection can trigger macrophages to undergo Caspase-1-mediated
cell lysis, releasing pro-inflammatory cytokines (such as IL-1β)
and thereby initiating the host immune response. Shao (7) first identified Gasdermin D (GSDMD) as
a substrate of inflammatory Caspases (Caspase-1/4/5/11) and the
Gasdermin D N-terminal fragment (GSDMD-N) forms pores in the cell
membrane, thereby triggering pyroptosis. Pyroptosis drives
inflammatory amplification, increases the size of myocardial
infarction and accelerates the occurrence of cardiovascular
diseases, while it also represents a promising therapeutic target
for cardiovascular disorders via the modulation of cardiomyocyte
pyroptosis (8). The absent in
melanoma 2 (AIM2) inflammasome is an intracellular protein platform
with extensive pro-inflammatory and pro-pyroptotic properties
(9). Hornung et al first
identified AIM2 as a sensor for cytosolic double-stranded DNA
(dsDNA), demonstrating that AIM2 binds to dsDNA via its HIN domain
and recruits apoptosis-associated Speck-like protein containing a
CARD and caspase-1 to form the AIM2 inflammasome complex (10). This study also showed that the
activation of the AIM2 inflammasome can trigger Caspase-1-dependent
pyroptosis and the release of IL-1β/IL-18, contributing to host
defense against intracellular pathogens such as DNA viruses and
bacteria. Jin et al (11)
further elucidated that AIM2 binds to dsDNA via its HIN domain,
activates inflammasome assembly and promotes Caspase-1-dependent
IL-1β secretion and pyroptosis. Mechanistically, the AIM2
inflammasome activates the innate immune response by cleaving
pro-Caspase-1 and processing IL-1β and IL-18 into mature forms,
while simultaneously promoting pyroptosis by generating the GSDMD-N
fragment from GSDMD (12). AIM2
expression is elevated in patients with HF irrespective of the
cause (that is, ischemic or dilated cardiomyopathy) (13,14).
Therefore, the AIM2 inflammasome may serve as a novel biomarker and
therapeutic target for HF. In addition, in murine HFpEF models,
myocardial levels of the pro-inflammatory cytokines TNF-α and IL-1β
are found to be elevated, accompanied by increased inflammatory
cell infiltration in cardiac tissue. Moreover, the
inflammasome-mediated pyroptosis pathway is activated in epicardial
adipose tissue and inhibition of GSDMD has been shown to reduce
cardiomyocyte inflammation and autophagy (15). However, despite these findings, the
mechanism of the AIM2/Caspase-1/GSDMD axis in HFpEF remains
unclear.
Dapagliflozin is an inhibitor of sodium-glucose
cotransporter type 2 (SGLT2) (16). Dapagliflozin can alleviate renal
pyroptosis by regulating the HO-1/NOD-like receptor family pyrin
domain containing 3 axis (17).
Additionally, dapagliflozin has been shown to suppress pyroptosis
mediated by the NLRP3/Caspase-1 pathway in vascular smooth muscle
cells by repressing CTSB (18). In
patients with HFpEF, 12 weeks of treatment with dapagliflozin was
shown to improve symptoms, physical limitations and motor function
in patients with HFpEF and was well tolerated in patients with
chronic HFpEF (19). However, the
molecular mechanism underlying the interplay between dapagliflozin
and pyroptosis in HFpEF remains largely unclear. The present study
hypothesized that dapagliflozin could alleviate pyroptosis and
improve HFpEF by inhibiting the AIM2/Caspase-1/GSDMD pathway. To
investigate this hypothesis, an HFpEF model was constructed and
dapagliflozin administered simultaneously to examine its effects on
cardiac function and inflammatory responses, thereby evaluating the
intervention effect of dapagliflozin during the development of
HFpEF. Furthermore, the present study assessed pyroptosis in mouse
cardiomyocytes by establishing a cell model combined with the
interference of AIM2 to explore the potential mechanism through
which dapagliflozin alleviates HFpEF.
Materials and methods
Animals
A total of 60 SPF-grade male C57 mice (8 weeks old,
weight, 20–24 g) were obtained from Hunan SJA Laboratory Animal
Co., Ltd. The animals were housed under controlled conditions
(temperature, 22±2°C; humidity, 50±10%; 12-h light/dark cycle) with
free access to standard chow and water. The animals underwent one
week of adaptive feeding before formal experimental procedures were
initiated. Prior to the start of the experiment, all mice were
randomly assigned to different experimental groups. There were 5
mice in each group. During the experiment, the researchers were
blinded to the group allocation information, ensuring that they
were unaware of the specific group assignments during experimental
operations and data collection.
After the experiment was completed, another
researcher who did not participate in the experimental operations
unblinded and analyzed the data, thereby ensuring the objectivity
and accuracy of data processing.
Preparation of lentivirus
A third-generation lentiviral packaging system was
used. 293T cells (cat. no. AW-CNH086; Changsha Abiwei Biotechnology
Co., Ltd.) were co-transfected with the transfer plasmid (oe-AIM2
or oe-NC), the packaging plasmid psPAX2, and the envelope plasmid
pMD2.G. In total, 5 µg transfer plasmid, 3.75 µg of psPAX2, and
1.25 µg of pMD2.G (mass ratio 4:3:1) were mixed and transfected for
48 h at room temperature using 25 µl Lipofectamine 2000 (cat. no.
11668019; Invitrogen). Six h post-transfection, the medium was
replaced with fresh complete medium containing 5% fetal bovine
serum (cat. no. 10099141, Gibco, Thermo Fisher Scientific) and 1%
penicillin/streptomycin. At 48 h after transfection, the viral
supernatant was collected. The supernatant was centrifuged at 4,000
× g for 10 min at 4°C to remove cell debris, followed by filtration
through a 0.45 µm filter. The filtrate was subjected to
ultracentrifugation at 4,450 × g for 2 h at 4°C. After carefully
discarding the supernatant, the viral pellet was resuspended in 1
ml PBS or cell culture medium, then centrifuged at 1,780 × g for 5
min at 4°C. The final titer was adjusted to 1×107 TU/ml.
Furthermore, successful lentiviral transduction in vivo was
confirmed by western blot analysis of protein expression in mouse
myocardial tissues at the end of the experiment. The lentivirus was
injected into the myocardium, and 7 days after injection, model
treatment and simultaneous intervention with dapagliflozin for 6
weeks were performed.
Group allocation and animal
treatments
Experiment 1 was divided into four groups: Sham,
Sham + Dapagliflozin, HFpEF and HFpEF + Dapagliflozin, with five
animals in each group. After the pH of N-omega-nitro-L-arginine
methyl ester (L-NAME; 0.5 g/l; cat. no. BD3941-25 g; Bide
Pharmatech Ltd.) was adjusted to 7.4, it was added to the drinking
water, followed by the start of high-fat feeding and the HFpEF
animal model was induced for 6 weeks (20).
Following the 6-week induction period and before the
intervention with dapagliflozin, the successful establishment of
the HFpEF model in mice treated with L-NAME combined with a
high-fat diet was confirmed, in strict accordance with the
diagnostic criteria for HFpEF recommended by the 2022 ACC/AHA/HFSA
Heart Failure Guideline (21) and
classic published studies in this field (22,23).
Conventional echocardiography and Doppler imaging were performed to
obtain representative left ventricular M-mode echocardiographs. The
key echocardiographic parameters were measured and calculated,
including left ventricular ejection fraction (LVEF), global
longitudinal strain (GLS) and the ratio of mitral E wave to E' wave
(E/E').
Heart weight to tibia length (HW/TL) was measured as
an auxiliary evaluation index. In addition, systolic blood pressure
(SBP) and diastolic blood pressure (DBP) of conscious mice were
measured by the noninvasive tail-cuff method (BP-98A) (24). The mice in the HFpEF group met the
diagnostic criteria for HFpEF, with preserved LVEF, typical
diastolic dysfunction (increased E/E' ratio, decreased GLS) and
abnormal blood pressure (elevated SBP and DBP), as well as obvious
myocardial hypertrophy and myocardial fibrosis. Mice in the HFpEF +
Dapagliflozin group received L-NAME and were intragastrically
administered dapagliflozin (1 mg/kg; 1 ml/100 g; cat. no.
BD161909-1g; Bide Pharmatech Ltd.) daily for 6 weeks (25). Dapagliflozin was administered
concurrently with the induction of the HFpEF model. Sham group
received an equal amount of normal saline and had unrestricted
access to food (normal feed) and water.
Experiment 2 was divided into the following groups:
Sham, oe-NC and oe-AIM2. The oe-NC and AIM2-overexpressing
lentiviruses (oe-AIM2) were injected into the mouse myocardium at a
viral concentration of 1×107 TU/ml with an injection
volume of 30 µl. Lentiviral transduction performed for 7 days
following intramyocardial injection (25). Experiment 3 was divided into the
following groups: HFpEF, HFpEF + dapagliflozin; HFpEF +
dapagliflozin + oe-NC and HFpEF + dapagliflozin + oe-AIM2. In the
HFpEF + dapagliflozin + oe-NC and HFpEF + dapagliflozin + oe-AIM2
groups, oe-NC and oe-AIM2 lentiviruses were injected into the mouse
myocardium 7 days prior to modeling and dapagliflozin treatment,
using the same viral concentration and an injection volume of 30 µl
(26). On the 8th day, HFpEF
induction was started and dapagliflozin treatment was initiated
simultaneously and continued for 6 weeks to evaluate the
interventional efficacy of dapagliflozin during HFpEF progression.
During the experimental period, the body weight of all mice were
measured weekly and the changes recorded. After the experiment, the
mouse myocardial tissues were collected for pathological
analysis.
Anesthesia was performed when mice required invasive
operations, including intramyocardial lentivirus injection,
echocardiographic detection (for more accurate parameter
measurement) and myocardial tissue collection. The anesthetic used
was 1% pentobarbital sodium (CAS: 57-33-0; cat. no. BD081201-10g;
Bide Pharmatech Ltd.) and the administration method and dose were
strictly standardized to ensure anesthesia safety and
effectiveness. Mice were anesthetized by intraperitoneal injection
at a dose of 50 mg/kg body weight (injection volume: 0.1 ml/10 g
body weight). After injection, the mice were placed in a warm
environment (25±1°C) and observed continuously until the righting
reflex disappeared (5–8 min), indicating that sufficient anesthesia
was achieved. During the entire invasive operation, the mouse's
respiratory rate and skin color were monitored to maintain the
depth of anesthesia; if the mouse showed signs of waking (such as
limb twitching, increased respiratory rate), supplementary
anesthesia was performed at 1/3 of the initial dose to ensure the
smooth progress of the operation.
Following the completion of all experimental
interventions (6 weeks of intervention), all mice were subjected to
humane sacrifice to collect myocardial tissues for subsequent
experimental detection. The process was divided into two steps to
ensure the mice suffered no pain: i) Pre-anesthesia: The same
anesthetic as used in invasive operations was adopted, namely 1%
pentobarbital sodium (CAS: 57-33-0; cat. no. BD081201-10g; Bide
Pharmatech Ltd.). Mice were anesthetized by intraperitoneal
injection at a dose of 100 mg/kg body weight (injection volume: 0.2
ml/10 g body weight), which was twice the dose used for surgical
anesthesia. This high dose was designed to ensure deep anesthesia,
so that the mice would not feel pain during the subsequent
euthanasia process. ii) Sacrifice: After confirming that the mice
had achieved deep anesthesia (loss of righting reflex and pain
response), cervical dislocation was performed to ensure rapid and
painless death of the mice. This method is a standard humane method
for mice, which can cause immediate cardiac and respiratory arrest
without causing unnecessary pain to the animals. To avoid false
death and ensure the rigor of the experiment, three key indicators
were comprehensively used to confirm the death of the mice after
euthanasia and only when all three indicators were met was the
mouse considered dead: i) Respiratory arrest: The chest movement of
the mice observed continuously for >30 sec; no obvious chest
undulation was observed, indicating that respiratory arrest has
occurred. ii) Cardiac arrest: The chest of the mice was gently
pressed with fingers for more than 1 min; no obvious heartbeat was
felt, confirming cardiac arrest. iii) Pupil response: The pupils of
the mice were observed under light stimulation; the pupils were
dilated and unresponsive to light, indicating that the central
nervous system has stopped functioning. All experimental protocols
were approved by the Animal Ethics Committee of Changsha Hospital
of Traditional Chinese Medicine (approval no. 2022111003).
Conventional echocardiography and
doppler imaging
A previous study (26) conducted transthoracic
echocardiography using a Vevo 2100 system equipped with an MS400
transducer (VisualSonics, Inc.). LVEF and other systolic function
indices were obtained from short-axis M-mode scans at the
mid-ventricular level in conscious mice that were gently
restrained, with the papillary muscles visible in the imaging
plane. Diastolic function measurements were collected from apical
4-chamber views of anesthetized mice using pulsed-wave Doppler and
tissue Doppler techniques at the level of the mitral valve.
Representative left ventricular M-mode echocardiographic images
were obtained. The percentage of LVEF and GLS, the ratio of the
mitral E/E' and the HW/TL were quantified. Each parameter was
measured ≥3 times.
Exercise exhaustion test
After 3 days of treadmill exercise, the mice were
subjected to an exhaustion test. The mice were initially set to run
uphill on a treadmill at a 20° incline, starting with a warm-up
speed of 5 m/min for 4 min. The speed was subsequently increased to
14 m/min and the mice continued to run at this speed for an
additional 2 min. Subsequently, the speed was incrementally
increased by 2 m/min every 2 min until the mice reached exhaustion.
Exhaustion was defined as the animal's failure to resume running
within 10 sec after making direct contact with the electrically
stimulated grid. Running duration was recorded and running distance
was further calculated (26).
Tail cuff blood pressure
recordings
SBP and DBP were assessed in conscious mice using
the noninvasive tail-cuff method (BP-98A) (24). Each mouse was placed in an
individual holder on a heated platform maintained at 37°C.
Measurements were performed once the mice reached a stable state.
Prior to the actual measurements, the mice underwent a training
period to acclimate them to brief periods of restraint.
Intraperitoneal glucose tolerance test
(ipGTT)
ipGTT was conducted by administering glucose (2 g/kg
in saline) following a 6-h fast (24,27).
Tail vein blood glucose concentrations were quantified using a
glucometer at baseline (0 min) and at 15, 30, 60, 90 and 120 min
following glucose administration.
Flow cytometry
The degree of pyroptosis in myocardial tissue was
assessed by flow cytometry. Myocardial tissues were cut into ~0.5
mm pieces using scissors and then homogenized. The resulting cell
suspension was gently dispersed by repeated aspiration with a
syringe. The cell suspension was filtered through a 100 µm filter
and the filtrate was collected and centrifuged at 178 × g for 5 min
at room temperature. For each sample, 300 µl of washing buffer was
mixed with 2 µl of FAM-YVAD-FMK (cat. no. ab219935; Abcam) and the
solution was protected from light. Each sample was incubated with
300 µl of the prepared dye solution at 37°C for 1 h in the dark for
1 h and then centrifuged at 178 × g for 5 min at room temperature.
Subsequently, the samples were washed twice with 200 µl of washing
buffer. The samples were then stained with the PI working solution
at 37°C for 10 min. Finally, pyroptosis was quantified as the
percentage of cells that were double-positive for active caspase-1
(FAM-YVAD-FMK+) and PI+ using a flow
cytometer (A00-1-1102, Beckman). Data acquisition and analysis were
performed using FlowJo™ v10.8 Software (BD Life Sciences).
For cellular pyroptosis detection, the dye solution
was prepared in the same manner. The number of samples was
calculated first and a concentrated dye solution was prepared. For
each sample, 300 µl of washing buffer was mixed with 2 µl of
FAM-YVAD-FMK and the solution was protected from light. The
remaining experimental steps were consistent with those for
myocardial tissue detection. A detailed gating strategy is
presented in Fig. S1.
Western blotting
Western blotting was performed to determine the
levels of pro-Caspase-1, Caspase-1, GSDMD-N, GSDMD, pro-IL-1β,
IL-1β, pro-IL-18, IL-18 and AIM2 in myocardial tissue or cells.
Initially, the tissues or cell samples were lysed in RIPA buffer
(cat. no. AWB0136; Changsha Abiwei Biotechnology Co., Ltd.) to
extract total protein. Protein concentration was determined using a
bicinchoninic acid assay. Equal amounts of protein (30 µg/lane)
were separated by 12% SDS-PAGE) and transferred onto nitrocellulose
membranes. Membranes were blocked with 5% skimmed milk in
Tris-buffered saline with 0.1% Tween-20 (TBST) for 1.5 h at room
temperature and subsequently incubated with primary antibodies
overnight at 4°C. The primary antibodies used were anti-Caspase-1
(cat. no. ab207802, 1:1000, Abcam), anti-GSDMD (cat. no.
20770-1-AP; 1:5,000; Proteintech Group, Inc.), anti-GSDMD-N (cat.
no. ab215203; 1:1,000; Abcam), anti-NLRP3 (cat. no. ab263899;
1:1,000; Abcam), anti-ASC (cat. no. 10500-1-AP; 1:15,000;
Proteintech Group, Inc.), anti-pro-IL-1β (16806-1-AP; 1:5,000;
Proteintech Group, Inc.), anti-pro-IL-18 (cat. no. 10663-1-AP;
1:10,000; Proteintech Group, Inc.), anti-IL-1β (cat. no. ab254360;
1:1,000; Abcam), anti-IL-18 (cat. no. ab191860; 1:1,000; Abcam),
anti-AIM2 (cat. no. AWA00390; 1:1,000; Changsha Abiwei
Biotechnology Co., Ltd.) and anti-GAPDH (cat. no. 10494-1-AP;
1:5,000; Proteintech Group, Inc.) antibodies. The membrane was then
incubated with an HRP-conjugated goat anti-mouse secondary antibody
(cat. no. SA00001-1, 1:5,000) or anti-rabbit secondary antibody
(cat. no. SA00001-2, 1:6,000; both Proteintech Group, Inc.) for 1.5
h at room temperature. After incubation, the membrane was treated
with enhanced chemiluminescence reagent (cat. no. AWB0005; Changsha
Abiwei Biotechnology Co., Ltd.) for 1 min and imaged using an
imaging system. GAPDH was used as the internal reference protein
and protein band intensities were quantified using Quantity One
4.6.6 (Bio-Rad Laboratories, Inc.).
For subcellular fractionation analysis, membrane and
cytoplasmic fractions were isolated separately prior to Western
blot. Western blot analysis was then performed to detect the levels
of GSDMD-N in the membrane and cytoplasmic fractions. The blots
were probed with anti-sodium/potassium-transporting ATPase subunit
α1 (ATP1a; cat. no. ab76020, 1:10,0000, Abcam) and anti-GAPDH
antibodies. ATP1a was used as the membrane marker and GAPDH was
used as the cytoplasmic marker and loading control, respectively.
Band quantification was conducted as aforementioned.
Biochemical detection
The concentrations of lactate dehydrogenase (LDH),
IL-1β and IL-18 in mouse myocardial tissues or cell supernatants
were quantified using biochemical assays. Specifically, commercial
kits for LDH (cat. no. A020-1-2; Nanjing Jiancheng Bioengineering
Institute), IL-1β (cat. no. CSB-E08054m; Cusabio Technology, LLC)
and IL-18 (cat. no. CSB-E04609m; Cusabio Technology, LLC) were used
following the manufacturers' instructions to quantify the
respective levels of these biomarkers.
Immunofluorescence (IF) staining
The levels of GSDMD and Caspase-1 in mouse
myocardial tissue were assessed via IF staining. Tissue sections
were first incubated at 60°C for 12 h. The sections were then
deparaffinized and rehydrated, followed by heat-induced antigen
retrieval. Sections were fixed with 4% paraformaldehyde for 30 min
at room temperature, followed by washing with PBS. Sections were
blocked with 5% BSA for 60 min at 37°C, followed by three washes
with PBS. Primary antibody against GSDMD (cat. no. 20770-1-AP;
1:100; Proteintech Group, Inc.) was added and the sections were
incubated overnight at 4°C, followed by three washes with PBS. The
sections were incubated with HRP-conjugated secondary antibody from
the immunohistochemistry secondary antibody kit (Mouse/Rabbit
Ultra-Sensitive Polymer Detection System, cat. no. AWI0629;
Changsha Abiwei Biotechnology Co., Ltd.; ready-to-use working
solution without further dilution, for 30 min at 37°C, followed by
three washes with PBS and then incubated with TYP-520 fluorescent
dye at 37°C for 5–10 min in the dark, followed by rinsing with PBS
three times for 5 min each. Heat-induced antigen retrieval was then
performed again. The sections were blocked with 0.3% hydrogen
peroxide for 15 min and rinsed with PBS three times for 5 min each.
The sections were blocked with 10% normal goat serum (cat. no.
005-000-121; Jackson ImmunoResearch Laboratories) or 5% BSA for 60
min, followed by three 3-min washes with PBS. Appropriately diluted
primary antibody against Caspase-1 (cat. no. AWA44822; 1:100;
Changsha Abiwei Biotechnology Co., Ltd.) was added dropwise and the
sections were incubated overnight at 4°C. After rinsing with PBS
three times for 5 min each, the sections were incubated with an
HRP-conjugated secondary antibody (cat. no. AWI0629; Changsha
Abiwei Biotechnology Co., Ltd.) at 37°C for 31 min, followed by
three washes with PBS. The sections were incubated with TYP-570
fluorescent dye at 37°C for 5 min in the dark, followed by rinsing
with PBS three times for 5 min each. Heat-induced antigen retrieval
was performed again and the sections were blocked with 0.3%
hydrogen peroxide for 15 min before rinsing with PBS three times
for 5 min each. The sections were blocked with 10% normal serum or
5% BSA for 60 min, followed by three 3-min washes with PBS.
Appropriately diluted primary antibody against cTnT (cat. no.
15513-1-AP; 1:100; Proteintech Group, Inc.) was added dropwise and
the sections were incubated overnight at 4°C. The sections were
rinsed with PBS three times for 5 min each, incubated with an
HRP-conjugated secondary antibody (cat. no. AWI0629; Changsha
Abiwei Biotechnology Co., Ltd.) at 37°C for 31 min and rinsed again
with PBS three times for 5 min each. The sections were incubated
with TYP-620 fluorescent dye at 37°C for 5 min in the dark,
followed by rinsing with PBS three times for 5 min each. DAPI (cat.
no. AWC0293a; Changsha Abiwei Biotechnology Co., Ltd.) was applied
to stain the nuclei at 37°C for 10 min. Finally, the sections were
mounted with buffered glycerin and stored in a light-protected
environment or examined under a fluorescence microscope.
The levels of Caspase-1 in mouse cardiomyocytes were
assessed via IF staining. The coverslips were removed, washed with
PBS 2–3 times and then in 4% paraformaldehyde for 30 min at room
temperature, followed by six washes with PBS. Subsequently, the
cells were permeabilized with 0.3% Triton X-100 for 30 min at 37°C.
After three 3-min washes with PBS, the samples were blocked with 5%
BSA for 60 min at 37°C, followed by three 3-min washes with PBS.
Appropriate dilutions of the primary antibody against Caspase-1
(cat. no. 22915-1-AP; 1:50; Proteintech Group, Inc.) were added and
the samples were incubated at 4°C overnight, followed by three
5-min washes with PBS. The secondary antibody,
CoraLite488-conjugated Goat Anti-Rabbit IgG (H + L) (cat. no.
SA00013-2; 1:200; Proteintech Group, Inc.), was added and incubated
for 90 min at 37°C, followed by three 5-min washes with PBS. For
nuclear staining, the samples were incubated with DAPI for 10 min
at 37°C. The samples were mounted with buffered glycerin and
protected from light or viewed under a fluorescence microscope.
Wheat germ agglutinin (WGA)
staining
Morphological changes in the myocardial tissue cell
membrane were evaluated using WGA staining. Briefly, the tissue
sections were incubated at 60°C for 12 h, then dewaxed to water and
the antigens underwent heat-induced retrieval. The sections were
placed in sodium borohydride solution for 30 min and then
transferred to 75% ethanol solution for 5 min at room temperature.
The WGA staining solution was prepared as follows: Particulate
matter was diluted with 0.01 M PBS (pH 7.2–7.6) at a ratio of 1
mg/ml. The prepared WGA working solution was applied to the
sections, which were then incubated in a humidified incubator at
37°C for 30 min under strict protection from light. For nuclear
staining, the sections were stained with DAPI for 10 min at 37°C.
The sections were then sealed with buffered glycerin and stored in
a light-protected environment or examined under a fluorescence
microscope.
Hematoxylin-eosin (H&E)
staining
Pathological alterations in myocardial tissue were
assessed using H&E staining. The sections were heated at
60°C for 12 h. Sections were placed in xylene (cat. no. 10023418;
Shanghai Sinopharm Group Co., Ltd.) and rehydrated in descending
ethanol. The sections were stained with hematoxylin (cat. no.
AWI0001a; Changsha Abiwei Biotechnology Co., Ltd.) for 1–10 min,
rinsed with distilled water and then counterstained using blue with
PBS. Subsequently, the sections were dyed with eosin (cat. no.
AWI0029a; Changsha Abiwei Biotechnology Co., Ltd.) for 1–5 min and
rinsed with distilled water. The sections were dehydrated in a
graded series of alcohol solutions (95–100%) for 5 min at each step
or until they were completely dry. Finally, the sections were
transferred to xylene (cat. no. 10023418; Shanghai Sinopharm Group
Co., Ltd.) for 10 min, mounted with neutral gum (cat. no. AWI0238a;
Changsha Abiwei Biotechnology Co., Ltd.) and examined under a
microscope.
Masson staining
A Masson staining kit (cat. no. AWI0253a; Changsha
Abiwei Biotechnology Co., Ltd.) was employed to evaluate collagen
fiber deposition in the myocardial tissue of the mice. The tissue
sections were incubated at 60°C for >12 h. Sections were placed
in xylene and rehydrated in descending ethanol. A total of 2 ml
nuclear dye was gently applied for 10 sec at room temperature. The
staining solution was then rinsed thoroughly with tap water,
followed by immersion of the sections in distilled water. The
sections were placed in a weak alkaline solution, such as PBS (pH
7.2–7.6) or ammonia, for 5 min to ensure that the nuclei returned
to a blue color. Subsequently, an appropriate volume of staining
solution was added to cover the entire tissue section and allowed
to stain for 5 min at room temperature. The sections were then
thoroughly washed with the rinsing solution to remove excess stain.
Subsequently, the sections were treated with a color separation
solution for ~30 sec. Thereafter, an appropriate volume of
restaining solution was added to completely cover the tissue
sections, which were stained for ~1 min at room temperature and
then rinsed with anhydrous ethanol. The sections were dried with
cold air from a hair dryer, cleared with xylene and finally
sealed.
Cell culture and treatment
Mouse cardiomyocytes were obtained from Changsha
Abiwei Biotechnology Co., Ltd. (cat. no. AW-CNM080) and cultured in
MEM supplemented with 10% high-quality fetal bovine serum (cat. no.
10099141, Gibco, Thermo Fisher Scientific, Inc.) and 1%
penicillin/streptomycin. The mouse cardiomyocytes are immortalized
cardiomyocytes (HL-1 cell line). Dapagliflozin (0, 0.25, 2.5, 5,
10, 20, 50, 100 and 200 µmol/l, 461432-26-8, MCE) was used to treat
mouse cardiomyocytes for 24 h and the maximum concentration that
did not affect cell viability was identified (25).
To investigate the role of AIM2 in pyroptosis in
HFpEF, AIM2 was knocked down and the following groups were
established: Control, short hairpin RNA-negative control (sh-NC),
sh-AIM2)#1, sh-AIM2#2 and sh-AIM2#3. After identifying the most
effective interference fragment, the following groups were
established: Control (mouse cardiomyocytes cultured normally for 72
h), Model group (mouse cardiomyocytes cultured normally for 48 h
and then treated with 1 µg/ml tunicamycin (11089-65-9, MCE) for 24
h), Model + sh-NC group (mouse cardiomyocytes transfected with
sh-NC for 48 h, then treated with 1 µg/ml tunicamycin for 24 h) and
Model + sh-AIM2 group (mouse cardiomyocytes transfected with
sh-AIM2 for 48 h and then treated with 1 µg/ml tunicamycin for 24
h). In addition, to explore the role of Caspase-1 in pyroptosis in
HFpEF, Caspase-1 was overexpressed and the following groups were
established: Control, overexpression-negative control (oe-NC) and
oe-Caspase-1. The cardiomyocytes were subsequently grouped as
follows: Control group (mouse cardiomyocytes cultured normally for
72 h), Model group (mouse cardiomyocytes cultured normally for 48 h
and treated with 1 µg/ml tunicamycin for 24 h), sh-NC + oe-NC group
(mouse cardiomyocytes transfected with sh-NC and oe-NC for 48 h and
then treated with 1 µg/ml tunicamycin for 24 h), sh-AIM2 + oe-NC
group (mouse cardiomyocytes co-transfected with sh-AIM2 and oe-NC
for 48 h and then treated with 1 µg/ml tunicamycin for 24 h) and
sh-AIM2 + oe-Caspase-1 group (mouse cardiomyocytes transfected with
sh-AIM2 and oe-Caspase-1 for 48 h and then treated with 1 µg/ml
tunicamycin for 24 h).
In addition, the cells were divided into three
groups: Control group (untreated normal control), oe-NC group
(transfected with empty oe-NC vector) and oe-AIM2 group
(transfected with AIM2 overexpression vector oe-AIM2). To
investigate the mechanism of dapagliflozin and AIM2 in pyroptosis
in HFpEF, the cells were grouped as follows: Control group (mouse
cardiomyocytes cultured normally for 72 h), Model group (mouse
cardiomyocytes cultured normally for 48 h and then treated with 1
µg/ml tunicamycin for 24 h), Model + Dapagliflozin group (mouse
cardiomyocytes cultured normally for 48 h and then treated
simultaneously with dapagliflozin and 1 µg/ml tunicamycin for 24
h), Model + dapagliflozin + oe-NC group (mouse cardiomyocytes
transfected with oe-NC for 48 h and then treated simultaneously
with dapagliflozin and 1 µg/ml tunicamycin for 24 h) and Model +
Dapagliflozin + oe-AIM2 group (mouse cardiomyocytes transfected
with oe-AIM2 for 48 h and treated simultaneously with dapagliflozin
and 1 µg/ml tunicamycin for 24 h).
Transfection was performed using Lipofectamine 3000
(cat. no. L3000015, Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. For knockdown, sh-AIM2 fragments (cat.
no. HG-Mi1013779) or sh-NC (both HonorGene) were used at a final
concentration of 100 nM. For overexpression, oe-AIM2 (HG-MO1013779,
HonorGene), oe-Caspase-1 (HG-MO009807; both HonorGene) or oe-NC
were used at 3 µg/well. Cells were incubated with the transfection
mixture for 48 h at 37°C. The time interval between transfection
and subsequent experimentation was 48 h. Untransfected control
cells were maintained under identical culture conditions. The
sequences used were as follows: sh-NC: 5′-TGCGGGGCAAGTCCACCACCA-3′;
sh-AIM2#1: 5′-GACCACATCACGGAGGAAGAACTGAA-3′; sh-AIM2#2:
5′-GCCCATTCTGTGAATAATACAAA-3′; and sh-AIM2#3:
5′-GAAACTCCCAGGATTAGTAAACTGAAG-3′.
Reverse transcription-quantitative
(RT-q) PCR
The mRNA levels of AIM2 and Caspase-1 were
quantified via RT-qPCR. First, mouse cardiomyocytes were seeded at
a density of 5×105 cells/well in a 6-well plate and
cultured until 80–90% confluence. Total RNA was isolated from
samples using a TRIzol® total RNA extraction kit (cat.
no. 15596026, Thermo Fisher Scientific, Inc.) and its concentration
and purity were determined. The extracted total RNA was then
reverse-transcribed into complementary DNA using an mRNA reverse
transcription kit (cat. no. CW2569; CWBio) according to the
manufacturer's protocols. Subsequently, relative gene expression
levels were analyzed with Ultra SYBR Mixture (cat. no. CW2601;
CWBio) on an ABI 7900 Real-Time PCR System (Applied Biosystems).
The thermocycling conditions were as follows: initial denaturation
at 95°C for 10 min, followed by 40 cycles of 95°C for 15 s
(denaturation), 60°C for 30 s (annealing), and 72°C for 30 sec. A
melt curve analysis was performed to confirm product specificity.
Gene expression levels were calculated with the 2−ΔΔCq
method (28), with GAPDH as the
internal reference gene. All experiments were performed in
triplicate. The primers used were as follows: AIM2-F:
5′-CTGTCTGCCGCCATGCTTC-3′; AIM2-R: 5′-CATGAATATACCAGCAGTCCAGT-3′;
Caspase-1-F: 5′-ACAAGGCACGGGACCTATG-3′; Caspase-1-R:
5′-TCCCAGTCAGTCCTGGAAATG-3′; GAPDH-F: 5′-GCGACTTCAACAGCAACTCCC-3′;
and GAPDH-R: 5′-CACCCTGTTGCTGTAGCCGTA-3′.
Cell Counting Kit 8 (CCK-8) assay
Cell viability was evaluated via a CCK-8 assay.
Cells were trypsinized and seeded into a 24-well plate at a density
of 5×103 cells per well, with 300 µl of culture medium
added to each well. Triplicate wells were established for each
group. After cell adhesion, 10 µl/well of CCK-8 reagent (cat. no.
NU679; Dojindo Laboratories, Inc.) was added at the corresponding
time points following treatment as aforementioned. After incubation
at 37°C in a humidified atmosphere containing 5% CO2 for
4 h, the absorbance was measured at a wavelength of 450 nm using a
microplate reader.
Transmission electron microscopy
(TEM)
TEM experiment was only employed to evaluate
mitochondrial damage, not to assess plasma membrane integrity or
GSDMD pore formation. Briefly, the cells were fixed with 2.5%
glutaraldehyde at 4°C for 6–12 h. After removal of the fixative,
the cells were rinsed with PBS buffer. Then, 1% osmium tetroxide
was added and the cells were fixed for 1–2 h. The cells were then
dehydrated, embedded in pure epoxy resin and cured in an oven at
40°C for 12 h. The samples were further heated at 60°C for 48 h.
After the embedded blocks were removed and trimmed, ultrathin
sections (70–90 nm in thickness) were cut, mounted on copper grids
and stained with electron-dense dyes (lead and uranium stains) at
room temperature. Finally, the sections were examined using a
transmission electron microscope (JEM 1400; JEOL, Ltd.).
Hoechst 33342/PI fluorescence
staining
Pyroptosis was evaluated using Hoechst 33342/PI
fluorescence staining. The excess medium was aspirated and 2,000 µl
of dye working solution was added (1 mg/ml Hoechst storage solution
prepared with DMSO and 1 mg/ml PI stock solution prepared with
DDH2O). The storage solution was diluted with preheated
serum-free culture medium or PBS to obtain final concentrations of
10 µg/ml Hoechst working solution and 20 µg/ml propidium iodide
working solution. The dye working solution was gently mixed with
the cells to ensure complete coverage, followed by incubation for 5
min at room temperature. Subsequently, the dye working solution was
removed and the cells were washed twice with serum-free culture
medium for 5 min each time before observation under a fluorescence
microscope.
Molecular docking
Molecular docking was performed to investigate the
binding interaction between dapagliflozin and AIM2. VINA 1.1.2
software (vina.scripps.edu/) was used to perform docking
simulations between compounds and proteins, employing semiempirical
free energy fields to predict receptor-ligand binding energies.
After docking simulations were completed, PyMOL software (version
2.6.2; pymol.org/) was used for structural visualization and
chemical analysis to evaluate the stable binding conformation of
dapagliflozin within the protein binding pocket and its
intermolecular interactions with adjacent amino acid residues.
Drug affinity responsive target
stability (DARTS) assay
DARTS was performed as previously described
(29). The recombinant AIM2
protein (0.5 µg/ml) was incubated with different concentrations of
dapagliflozin (0–50 µM) at room temperature for 1 h with gentle
shaking. Pronase (5 µg/ml) was added and the mixture was incubated
for 15 min at room temperature. The reaction was then terminated by
adding loading buffer. AIM2 protein levels were subsequently
detected by western blotting.
Cellular thermal shift assay
(CETSA)
For the living cell CETSA, cells were treated with
or without dapagliflozin for 3 h. The cells were then collected and
equally divided into 12 portions in 0.2% NP40-PBS solution, heated
at specific temperatures (37–72°C) and subsequently cooled for 3
min. The samples were lysed by three freeze-thaw cycles using
liquid nitrogen. After ultracentrifugation at 20,000 × g for 20 min
at 4°C, the supernatant was collected and subjected to Western blot
analysis (30). For
concentration-dependent CETSA, the samples were treated with
vehicle control or dapagliflozin (2, 5, 10, 20, 50, 100, 200, 300
or 500 µM), followed by heating for 3 min and cooling for 3 min.
High-speed centrifugation at 20,000 × g for 20 min at 4°C was
performed and the supernatant was collected for western blot
analysis.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 8.0 software (Dotmatics). Measurement data are presented as
the mean ± standard deviation. The normality of data distribution
was assessed using the Shapiro-Wilk test (31). For comparisons between two groups,
the F-test associated with unpaired Student's t-test was used to
assess homogeneity of variance (31). The Brown-Forsythe test was applied
to examine homogeneity of variance across multiple groups (32). If both normality and homogeneity of
variance were confirmed, differences between two groups were
analyzed using an unpaired Student's t-test (31). When normality was confirmed but
homogeneity of variance was not satisfied, differences between two
groups were analyzed using the Mann-Whitney U test (33,34).
For multiple-group comparisons of normally distributed data,
one-way analysis of variance (ANOVA) was applied (35), followed by Tukey-Kramer post hoc
tests when the ANOVA results were significant (36). In the Shapiro-Wilk test, P>0.1
indicated that the data followed a normal distribution. For the
F-test and Brown-Forsythe test, P>0.05 indicated homogeneity of
variance. For the Mann-Whitney U test, unpaired Student's t-test or
one-way ANOVA, P<0.05 was considered to indicate a statistically
significant difference. No data were excluded and all analyses were
conducted in a blinded manner. For post-hoc power analysis, G*Power
3.1.9.2 software (Heinrich Heine University Düsseldorf) was
employed with parameters including the actual effect size (Cohen's
d), α=0.05, n=5 per group and the corresponding number of
experimental groups. The results demonstrated that the power values
of all primary endpoint indicators were greater than 0.8,
confirming that a sample size of n=5 per group was sufficient to
meet the detection requirements. Pearson correlation analysis was
used to evaluate the linear relationship between two continuous
variables (37). Before performing
Pearson correlation analysis, normality tests were conducted to
confirm that the data followed a normal distribution. The
correlations between AIM2 protein levels and pyroptosis-related
indices (IL-18, IL-1β, GSDMD-N and Caspase-1), as well as
myocardial function indicators (LVEF, GLS and E/E'), were
determined using Pearson correlation coefficients.
Results
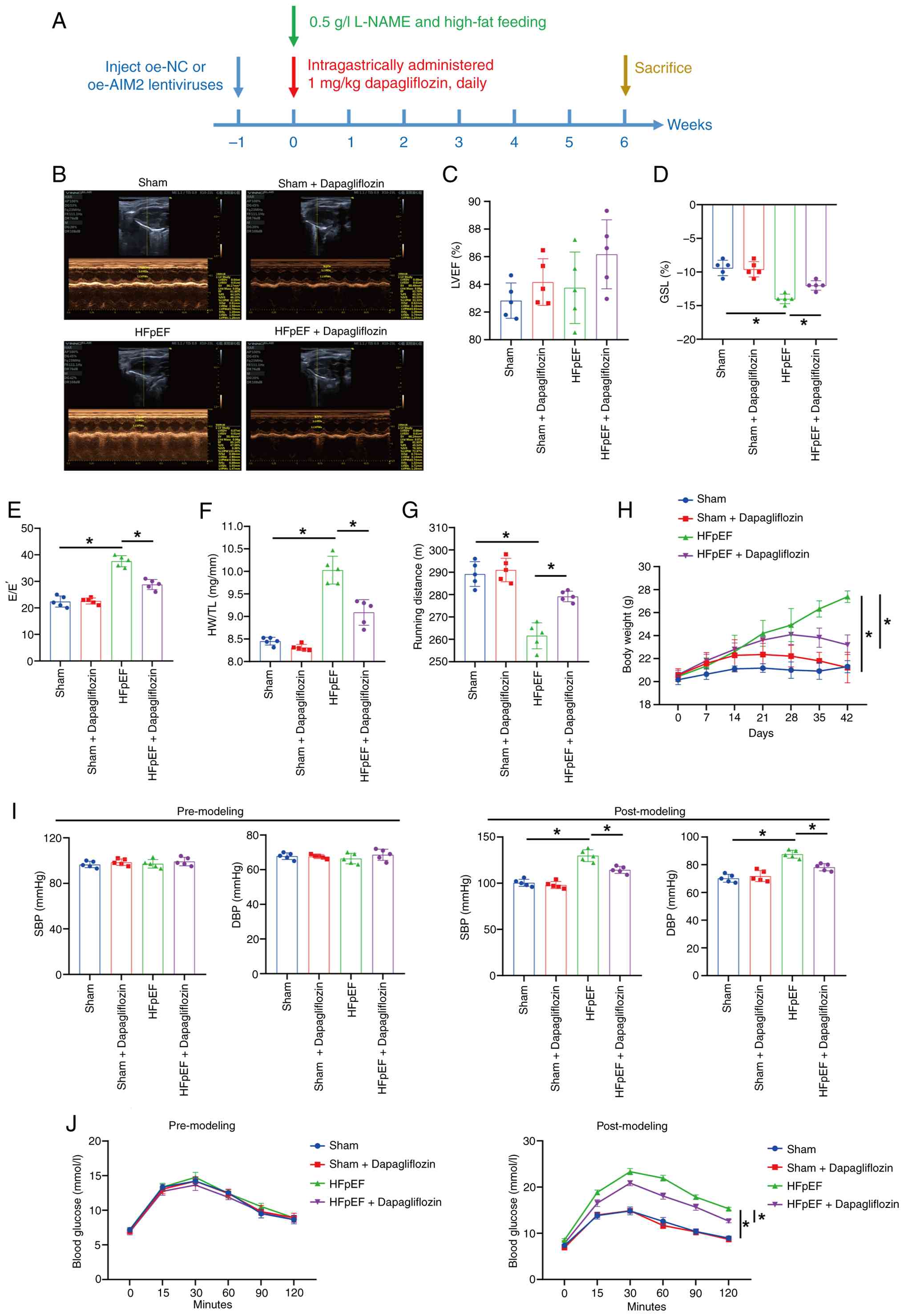
Dapagliflozin improves HFpEF symptoms
in mice
The present study constructed an HFpEF model and
administered dapagliflozin simultaneously to evaluate the
interventional effect of dapagliflozin during the development of
HFpEF. Fig. 1A illustrates the
overall experimental workflow from lentiviral injection to the end
of the study. Briefly, mice first received intramyocardial
lentiviral injection for 7 days. On day 8 following lentiviral
injection, HFpEF model induction was initiated and dapagliflozin
intervention was started concurrently and continued for 6 weeks.
Fig. 1B shows representative left
ventricular M-mode echocardiographic images. The percentage of LVEF
was essentially comparable among all groups (Fig. 1C). By contrast, compared with the
Sham group, the percentage of GLS was lower in the HFpEF group,
whereas dapagliflozin treatment was associated with an increase in
GLS (Fig. 1D). Furthermore,
compared with the Sham group, the E/E' ratio and heart weight to
tibia length (HW/TL) ratio were higher in the HFpEF group, while
the running distance in the exercise exhaustion test was reduced.
Following dapagliflozin treatment, the E/E' and HW/TL ratios
decreased, whereas the running distance increased (Fig. 1E-G). The present study also
monitored body weight, blood pressure and blood glucose in mice.
Over the 6-week intervention, mice in the HFpEF group exhibited
increased body weight compared with the Sham group, whereas
dapagliflozin treatment was associated with reduced body weight
(Fig. 1H). Following model
induction, SBP and DBP were markedly increased in the HFpEF group
compared with the Sham group and blood glucose levels were also
elevated. Following dapagliflozin administration, both SBP and DBP
were markedly reduced, accompanied by a significant decrease in
blood glucose (Fig. 1I and J).
Collectively, these results demonstrated that dapagliflozin
alleviated HFpEF-related phenotypes in mice. Given that
dapagliflozin had no effect on the cardiac function of Sham mice,
further exploration was conducted on the remaining three groups
(Sham, HFpEF and HFpEF + Dapagliflozin groups).
Dapagliflozin reduces pyroptosis in
the myocardial tissue of HFpEF mice
Next, the effect of dapagliflozin on the pyroptosis
levels in the myocardial tissue of HFpEF mice were investigated.
Compared with the Sham group, myocardial pyroptosis was markedly
elevated in the HFpEF group, whereas this increase was markedly
suppressed following dapagliflozin treatment (Fig. 2A). Additionally, the levels of
pro-Caspase-1, Caspase-1, GSDMD, GSDMD-N, AIM2, NLRP3, ASC,
pro-IL-1β, pro-IL-18, IL-1β, IL-18 and LDH in myocardial tissue
were increased in the HFpEF group compared with the Sham group;
however, these elevations were reduced after dapagliflozin
administration (Fig. 2B-E). WGA
staining revealed that myocardial hypertrophy was more pronounced
in the HFpEF group than in the Sham group and dapagliflozin
treatment reduced myocardial hypertrophy to some extent (Fig. 2F). Moreover, compared with the Sham
group, the HFpEF group exhibited increased myocardial inflammatory
infiltration and fibrosis. After dapagliflozin treatment,
inflammatory infiltration in myocardial tissue was reduced and
fibrosis was alleviated (Fig. 2G and
H). Together, these results indicate that dapagliflozin
inhibited pyroptosis in the myocardial tissue of HFpEF mice.
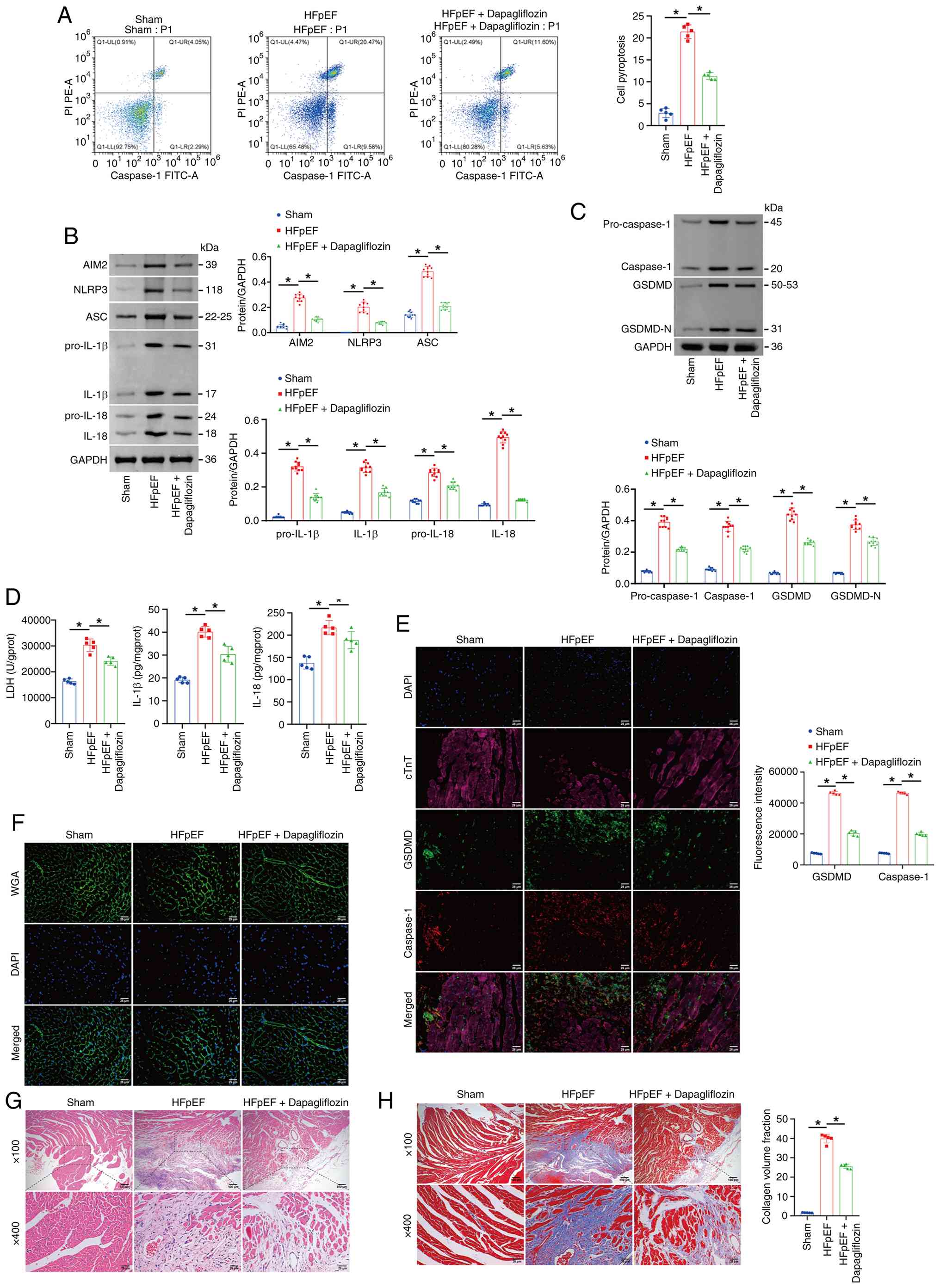 | Figure 2.Dapagliflozin reduces pyroptosis in
the myocardial tissue of HFpEF mice. (A) Flow cytometry analysis of
myocardial tissue pyroptosis (Caspase-1). (B and C) Western blot
analysis of pro-Caspase-1, Caspase-1, GSDMD, GSDMD-N, AIM2, NLRP3,
ASC, pro-IL-1β, pro-IL-18, IL-1β and IL-18 levels in myocardial
tissue. (D) LDH, IL-1β and IL-18 levels in myocardial tissue were
determined using commercial kits. (E) IF staining of GSDMD and
Caspase-1. (F) WGA staining of morphological alterations in the
myocardial tissue cell membrane in mice. (G) Pathological changes
in myocardial tissues assessed via H&E staining. (H) Masson
staining of collagen fiber deposition in the myocardial tissue of
the mice. Scale bars, 100 µm, magnification, ×100; scale bars, 25
µm, magnification, ×400. The blot was probed with anti-GAPDH
antibody. GAPDH was used as a loading control. *P<0.05. HFpEF,
heart failure with preserved ejection fraction; GSDMD, Gasdermin D;
GSDMD-N, Gasdermin D N-terminal fragment; AIM2, absent in melanoma
2; NLRP3, NOD-like receptor family pyrin domain containing 3; ASC,
apoptosis-associated Speck-like protein containing a CARD; LDH,
lactate dehydrogenase; IF, immunofluorescence; WGA, wheat germ
agglutinin; H&E, hematoxylin-eosin. |
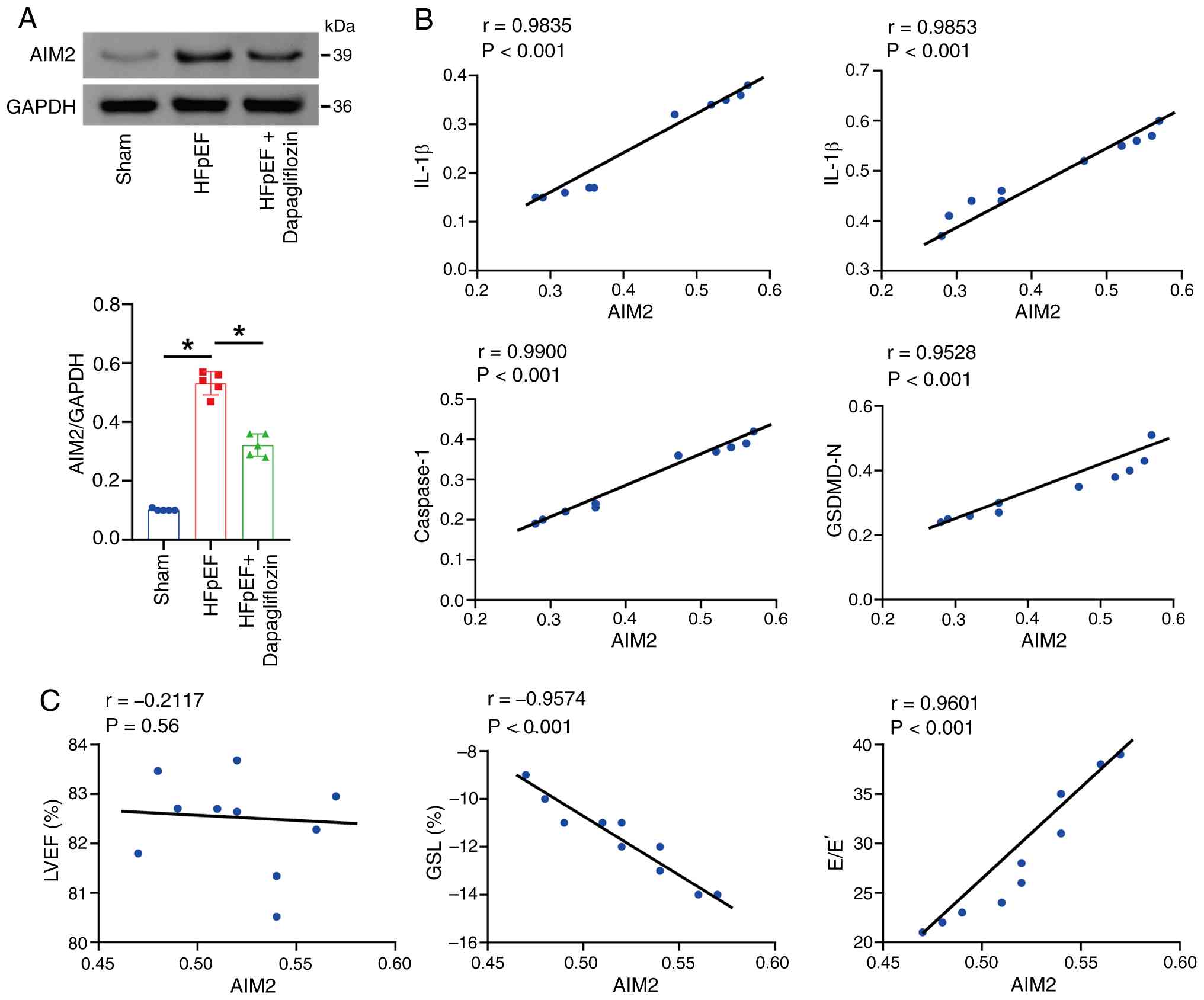
Changes in pyroptosis levels in HFpEF
mice are accompanied by fluctuations in AIM2 protein levels
AIM2 is elevated in human HF regardless of the cause
(ischemic or dilated cardiomyopathy) (13) and may represent a novel biomarker
and therapeutic target for HF. Therefore, the role of AIM2 in HFpEF
was next explored. The present study first assessed AIM2 protein
levels in mouse myocardial tissue and found that, compared with the
Sham group, AIM2 expression was increased in the HFpEF group,
whereas AIM2 expression decreased after dapagliflozin treatment
(Fig. 3A). Pearson correlation
analysis further showed that AIM2 levels were positively associated
with IL-1β, IL-18, Caspase-1 and GSDMD-N (Fig. 3B). In addition, AIM2 levels were
not markedly associated with LVEF; however, AIM2 levels were
markedly negatively associated with GLS and markedly positively
associated with E/E' (Fig. 3C),
which further supports a potential role for AIM2 in HFpEF.
To determine whether AIM2 overexpression alone was
sufficient to activate the Caspase-1/GSDMD axis, promote pyroptosis
and exacerbate inflammatory injury, AIM2 expression was upregulated
in mouse myocardial tissue and in cultured cardiomyocytes to
investigate its functional role in HFpEF pathogenesis. In
vivo, AIM2 protein expression was markedly higher in the
oe-AIM2 group compared with the oe-NC group, confirming successful
AIM2 overexpression in mouse myocardial tissue (Fig. 4A). Moreover, the protein levels of
NLRP3, ASC, pro-Caspase-1, Caspase-1 (p20), GSDMD, GSDMD-N,
pro-IL-1β, pro-IL-18, IL-1β and IL-18 were markedly elevated in the
oe-AIM2 group compared with the oe-NC group, indicating that AIM2
overexpression robustly activated the Caspase-1/GSDMD pathway and
promoted inflammatory factor release (Fig. 4B). Consistently, the levels of LDH,
IL-1β and IL-18 were markedly higher in the oe-AIM2 group than in
the oe-NC group, suggesting that AIM2 overexpression exacerbated
tissue injury and inflammatory responses (Fig. 4C and D). In vitro, cell
viability was markedly lower in the oe-AIM2 group than in the oe-NC
group, indicating that AIM2 overexpression reduced cardiomyocyte
viability (Fig. 4E). Flow
cytometric analysis further revealed that the proportion of
pyroptotic (Caspase-1 positive) cells was markedly higher in the
oe-AIM2 group than in the oe-NC group, suggesting that AIM2
overexpression markedly promoted cardiomyocyte pyroptosis (Fig. 4F). In line with these findings, the
expression levels of AIM2, NLRP3, ASC, pro-Caspase-1, Caspase-1
(p20), GSDMD, GSDMD-N, pro-IL-1β and pro-IL-18 were markedly
increased in the oe-AIM2 group relative to the oe-NC group, further
confirming activation of the Caspase-1/GSDMD pathway and enhanced
inflammatory factor release following AIM2 overexpression (Fig. 4G). Additionally, LDH, IL-1β and
IL-18 levels in the cell supernatant were markedly higher in the
oe-AIM2 group than in the oe-NC group, indicating aggravated
cellular injury and inflammation upon AIM2 overexpression (Fig. 4H). To investigate whether AIM2
overexpression affects the subcellular localization of GSDMD-N,
western blot analysis was performed on membrane and cytoplasmic
fractions. As shown in the Figure
4I, compared with the oe-NC group, AIM2 overexpression markedly
increased GSDMD-N expression in the membrane fraction (relative to
the membrane marker ATP1a), while cytoplasmic GSDMD-N levels were
also markedly elevated (relative to the cytoplasmic marker GAPDH).
These results suggested that AIM2 may enhance pyroptosis execution
by promoting GSDMD cleavage activation and its translocation to the
cell membrane. Collectively, these results indicate that changes in
pyroptosis levels in HFpEF mice were accompanied by corresponding
fluctuations in AIM2 protein levels.
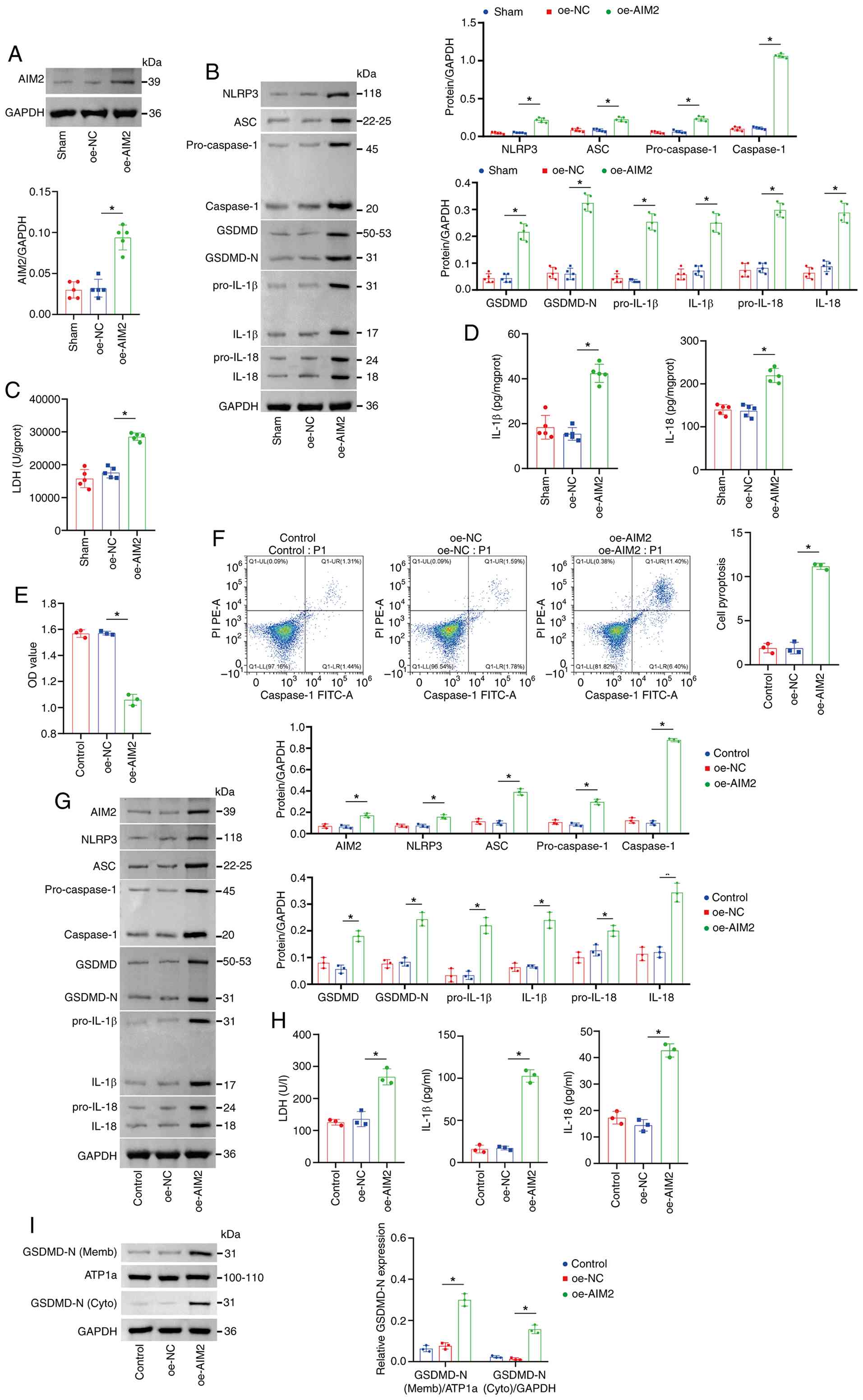 | Figure 4.AIM2 promoted pyroptosis in mouse
myocardial tissue and cardiomyocytes. (A) Western blot analysis of
AIM2 protein expression in mouse myocardial tissue. The blot was
probed with anti-GAPDH antibody. GAPDH was used as a loading
control. (B) Western blot analysis of the expression of NLRP3, ASC,
pro-Caspase-1, Caspase-1 (p20), GSDMD, GSDMD-N, pro-IL-1β,
pro-IL-18, IL-1β and IL-18 in myocardial tissue. The blot was
probed with anti-GAPDH antibody. GAPDH was used as a loading
control. (C) Measurement of myocardial LDH and (D) Measurement of
myocardial IL-1β and IL-18 levels. (E) Cell viability detected by
CCK-8 assay. (F) Cardiomyocyte pyroptosis rate (Caspase-1) measured
by flow cytometry. (G) Western blot analysis of AIM2, NLRP3, ASC,
pro-Caspase-1, Caspase-1 (p20), GSDMD, GSDMD-N, pro-IL-1β,
pro-IL-18, IL-1β and IL-18 levels in cell lysates. GAPDH was used
as a loading control. (H) Measurement of LDH, IL-1β and IL-18
levels in cell supernatant using commercial assay kits. (I) Western
blot analysis of the levels of GSDMD-N in the membrane and
cytoplasm. The blot was probed with anti-ATP1a/GAPDH antibody.
ATP1a (membrane marker) and GAPDH (cytoplasmic marker) were used as
a loading control. *P<0.05. AIM2, absent in melanoma 2; NLRP3,
NOD-like receptor family pyrin domain containing 3; ASC,
apoptosis-associated Speck-like protein containing a CARD; GSDMD,
Gasdermin D; GSDMD-N, Gasdermin D N-terminal fragment; LDH, lactate
dehydrogenase; oe, overexpressed; NC, negative control. |
AIM2 knockdown alleviates pyroptosis
in mouse cardiomyocytes
To investigate the role of AIM2 in mouse
cardiomyocyte pyroptosis, the present study first knocked down
AIM2. Compared with the sh-NC group, AIM2 expression was markedly
downregulated in the sh-AIM2#1, sh-AIM2#2 and sh-AIM2#3 groups,
with the most pronounced decrease in the sh-AIM2#3 group.
Therefore, sh-AIM2#3 was selected for subsequent experiments
(Fig. 5A). Next, mouse
cardiomyocytes were treated with 1 µg/ml tunicamycin to construct a
cellular model. Compared with the Control group, cardiomyocyte
viability was decreased, pyroptosis was increased and mitochondrial
damage was aggravated in the Model group. Following AIM2 knockdown,
the viability of mouse cardiomyocytes increased, pyroptosis
decreased and mitochondrial damage was alleviated (Fig. 5B-D). Furthermore, the levels of
AIM2, pro-Caspase-1, Caspase-1, GSDMD, GSDMD-N, NLRP3, ASC,
pro-IL-1β, pro-IL-18, IL-1β, IL-18 and LDH were markedly increased
in the Model group relative to the Control group. Following AIM2
knockdown, the levels of these factors were decreased in mouse
cardiomyocytes (Fig. 5E and F).
Moreover, compared with the Control group, the percentage of
PI-positive cardiomyocytes in the Model group was greater, whereas
AIM2 interference reduced the proportion of PI-positive mouse
cardiomyocytes (Fig. 5G). To
investigate whether AIM2 regulates the subcellular localization of
GSDMD-N, western blot analysis was performed on membrane and
cytoplasmic fractions. As shown in the Fig. 5H, compared with the Control group,
the Model group exhibited markedly elevated GSDMD-N protein
expression in the membrane fraction, along with markedly increased
GSDMD-N levels in the cytoplasmic fraction, indicating substantial
GSDMD activation and membrane translocation in the model group.
Transfection with sh-AIM2 markedly suppressed GSDMD-N protein
levels in both the cell membrane and cytoplasm. ATP1a (membrane
internal reference) and GAPDH (cytoplasmic internal reference)
showed stable expression across all groups, confirming the
reliability of subcellular fractionation and equal loading. These
results demonstrated that knockdown of AIM2 could alleviate
pyroptosis in mouse cardiomyocytes.
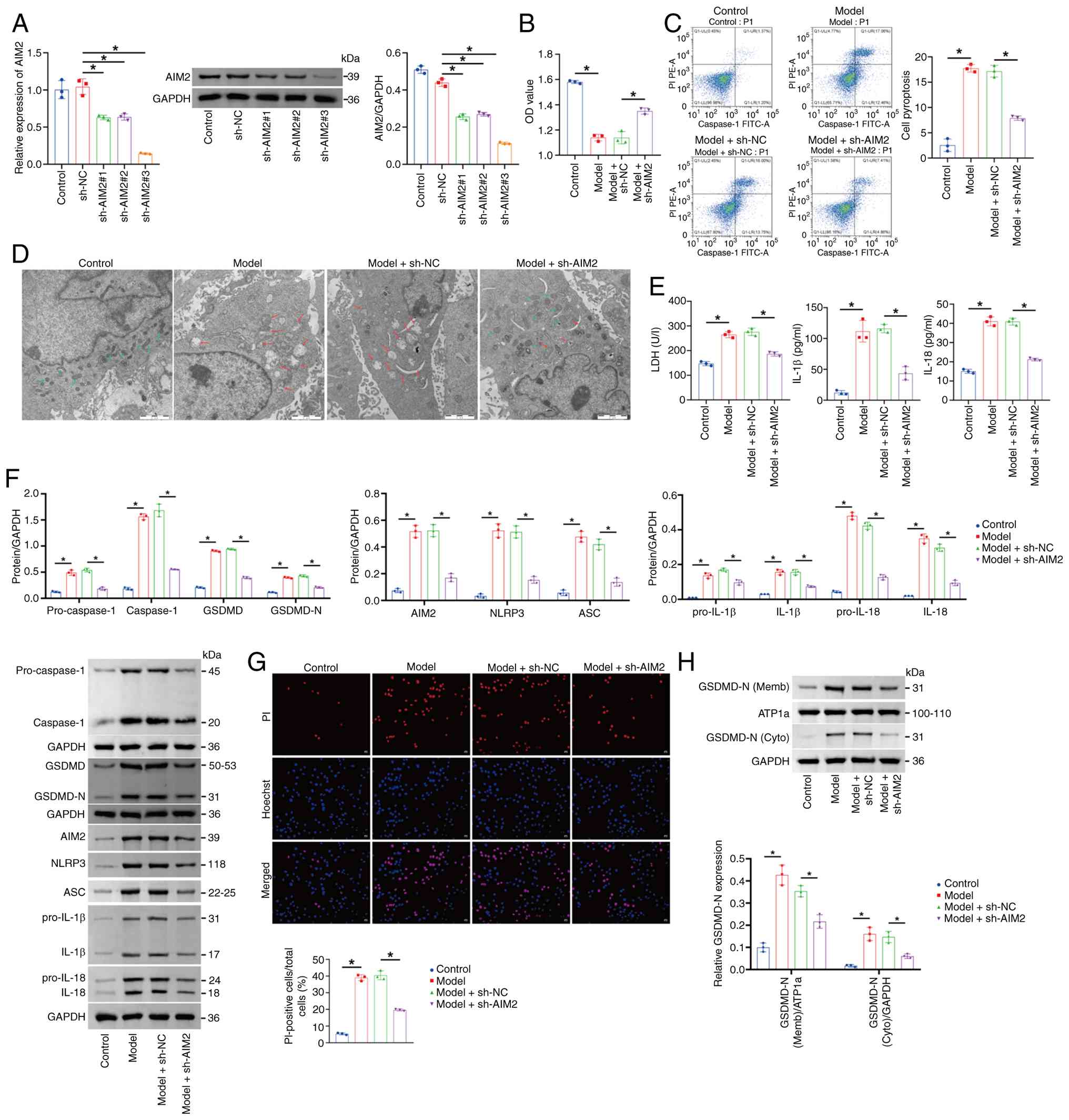 | Figure 5.AIM2 knockdown alleviates pyroptosis
in mouse cardiomyocytes. (A) AIM2 levels assessed by reverse
transcription-quantitative PCR and western blotting. The blot was
probed with anti-GAPDH antibody. GAPDH was used as a loading
control. (B) CCK-8 assay measurement of cell viability. (C) Flow
cytometry analysis of pyroptosis (Caspase-1). (D) Transmission
electron microscopy analysis of mitochondrial damage. Scale bar, 2
µm. (E) LDH, IL-1β and IL-18 levels were determined. (F) Western
blot analysis of AIM2, Pro-Caspase-1, Caspase-1, GSDMD, GSDMD-N,
NLRP3, ASC, pro-IL-1β, pro-IL-18, IL-1β and IL-18 levels in the
cell lysate. The blot was probed with anti-GAPDH antibody. GAPDH
was used as a loading control. (G) Hoechst 33342/PI fluorescence
staining showing the percentage of PI-positive mouse
cardiomyocytes. Scale bar, 50 µm, magnification, ×200. (H) Western
blot analysis of the levels of GSDMD-N in the membrane and
cytoplasm. The blot was probed with anti-ATP1a/GAPDH antibody.
ATP1a (membrane marker) and GAPDH (cytoplasmic marker) were used as
a loading control. *P<0.05. AIM2, absent in melanoma 2; GSDMD,
Gasdermin D; GSDMD-N, Gasdermin D N-terminal fragment; NLRP3,
NOD-like receptor family pyrin domain containing 3; ASC,
apoptosis-associated Speck-like protein containing a CARD; LDH,
lactate dehydrogenase; sh, short hairpin; NC, negative control. |
AIM2 regulated pyroptosis in mouse
cardiomyocytes via the Caspase-1/GSDMD axis
The AIM2/Caspase-1/GSDMD signaling pathway has been
reported to mediate renal cell pyroptosis in chronic kidney disease
(38). However, the mechanism
through which the AIM2/Caspase-1/GSDMD axis functions in HFpEF
remains unclear. Therefore, Caspase-1 was overexpressed in mouse
cardiomyocytes to explore the role of Caspase-1 in HFpEF-related
pyroptosis. Compared with the oe-NC group, the levels of
pro-Caspase-1 and Caspase-1 were markedly increased in the
oe-Caspase-1 group (Fig. 6A and
B). Compared with the Control group, cardiomyocyte viability
was decreased, pyroptosis was increased and mitochondrial damage
was aggravated in the Model group. Caspase-1 overexpression
reversed the increased viability, reduced pyroptosis and alleviated
mitochondrial damage in mouse cardiomyocytes caused by AIM2
knockdown (Fig. 6C-F).
Furthermore, compared with the Control group, the levels of LDH,
pro-Caspase-1, Caspase-1, GSDMD, GSDMD-N, AIM2, NLRP3, ASC,
pro-IL-1β, pro-IL-18, IL-1β and IL-18 levels in the Model group
were increased. However, the overexpression of Caspase-1 could
reverse the decreased expression of LDH, pro-Caspase-1, Caspase-1,
GSDMD, GSDMD-N, AIM2, NLRP3, ASC, pro-IL-1β, pro-IL-18, IL-1β and
IL-18 in mouse cardiomyocytes caused by the knockdown of AIM2
(Fig. 6G-J). Furthermore, compared
with the Control group, the percentage of PI-positive
cardiomyocytes in the Model group was greater. Caspase-1
overexpression reversed the decrease in PI-positive mouse
cardiomyocytes caused by AIM2 knockdown (Fig. 6K and L). To investigate whether
AIM2 regulates GSDMD-N membrane translocation through Caspase-1,
western blot analysis was performed on membrane and cytoplasmic
fractions. As shown in the Fig.
6M, compared with the Control group, the Model group displayed
markedly increased relative expression levels of GSDMD-N protein in
the cell membrane fraction, while GSDMD-N protein levels in the
cytoplasmic fraction were also markedly upregulated, suggesting
significant GSDMD activation and membrane translocation in the
Model group. Compared with the sh-NC + oe-NC group, membrane
GSDMD-N expression in the sh-AIM2 + oe-NC group was markedly
decreased, with cytoplasmic GSDMD-N levels also decreasing
synchronously; however, upon overexpressing Caspase-1 on the basis
of sh-AIM2 (sh-AIM2 + oe-Caspase-1 group), GSDMD-N expression in
both the cell membrane and cytoplasm was markedly rescued,
indicating that Caspase-1 overexpression can reverse the inhibitory
effect of AIM2 knockdown on GSDMD activation. ATP1a (membrane
internal reference) and GAPDH (cytoplasmic internal reference)
maintained stable expression across all groups, validating loading
uniformity. Collectively, these results indicated that AIM2
regulates pyroptosis in mouse cardiomyocytes through the
Caspase-1/GSDMD axis.
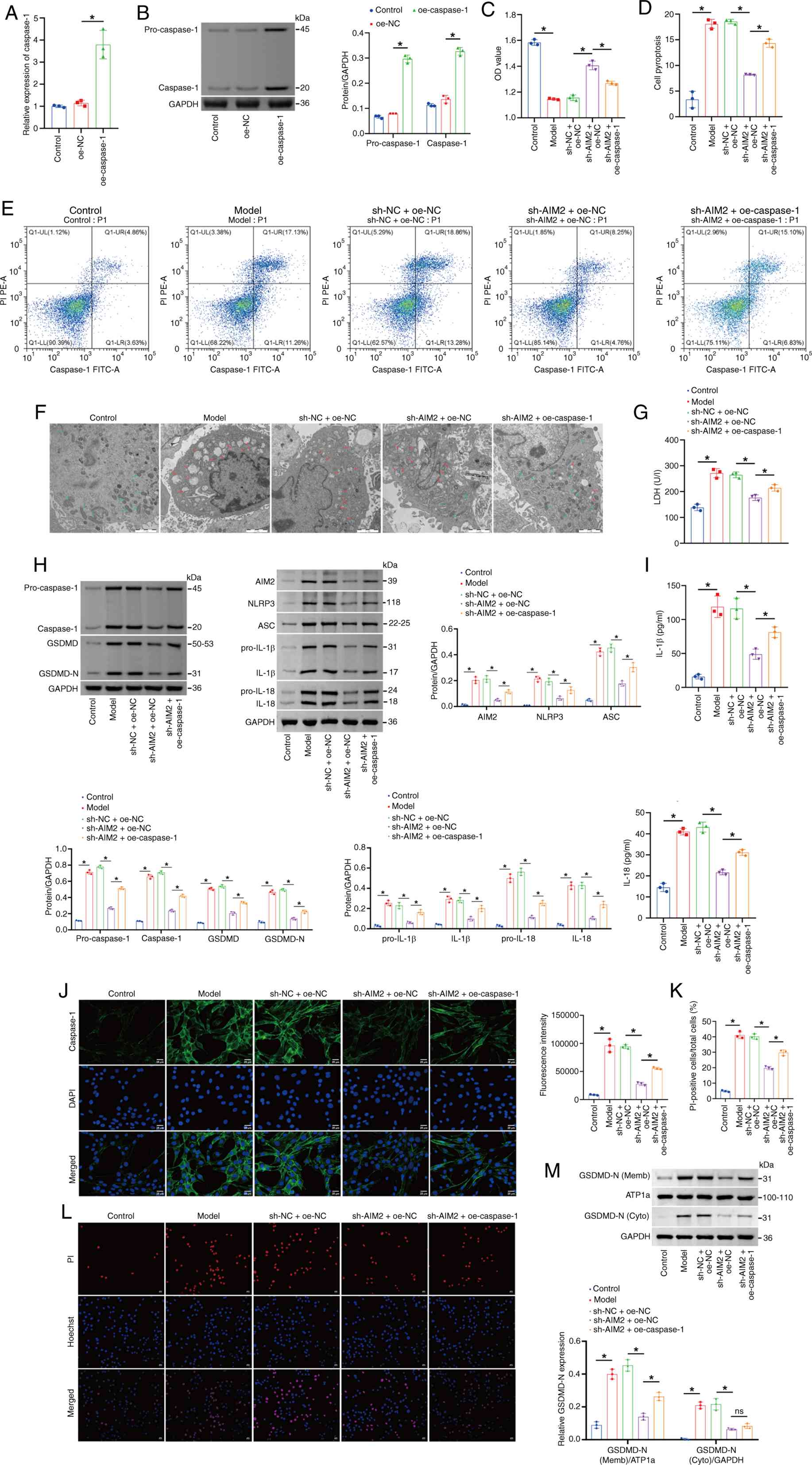 | Figure 6.AIM2 regulates pyroptosis in
cardiomyocytes via the Caspase-1/GSDMD axis. (A) Caspase-1 levels
evaluated by reverse transcription-quantitative PCR. (B) Western
blot analysis of pro-Caspase-1 and Caspase-1 levels. The blot was
probed with anti-GAPDH antibody. GAPDH was used as a loading
control. (C) CCK-8 assay of cell viability. (D and E) Flow
cytometry analysis of pyroptosis (Caspase-1). (F) TEM analysis of
mitochondrial damage. Scale bar, 2 µm. (G) LDH content in the cell
supernatant. (H) Western blot assessment of pro-Caspase-1,
Caspase-1, GSDMD, GSDMD-N, AIM2, NLRP3, ASC, pro-IL-1β, pro-IL-18,
IL-1β and IL-18 levels in the cell lysate. The blot was probed with
anti-GAPDH antibody. GAPDH was used as a loading control. (I) IL-1β
and IL-18 levels in the cell supernatant. (J) IF staining of
Caspase-1 in cells. (K and L) Hoechst 33342/PI fluorescence
staining showing the percentage of PI-positive mouse
cardiomyocytes. Scale bar, 50 µm, magnification, ×200; scale bar,
25 µm, magnification, ×400. (M) Western blot analysis of the levels
of GSDMD-N in the membrane and cytoplasm. The blot was probed with
anti-ATP1a/GAPDH antibody. ATP1a (membrane marker) and GAPDH
(cytoplasmic marker) were used as a loading control. *P<0.05.
AIM2, absent in melanoma 2; GSDMD, Gasdermin D; TEM, transmission
electron microscopy; LDH, lactate dehydrogenase; GSDMD-N, Gasdermin
D N-terminal fragment; NLRP3, NOD-like receptor family pyrin domain
containing 3; ASC, apoptosis-associated Speck-like protein
containing a CARD; IF, immunofluorescence; ATP1a,
sodium/potassium-transporting ATPase subunit α1; oe, overexpressed;
NC, negative control; sh, short hairpin. |
Dapagliflozin inhibits the
AIM2/Caspase-1/GSDMD axis to alleviate pyroptosis in mouse
cardiomyocytes
Dapagliflozin has been reported to improve symptoms,
physical limitations and motor function in patients with HFpEF
(19). However, the mechanism by
which dapagliflozin and AIM2 regulate pyroptosis in HFpEF remains
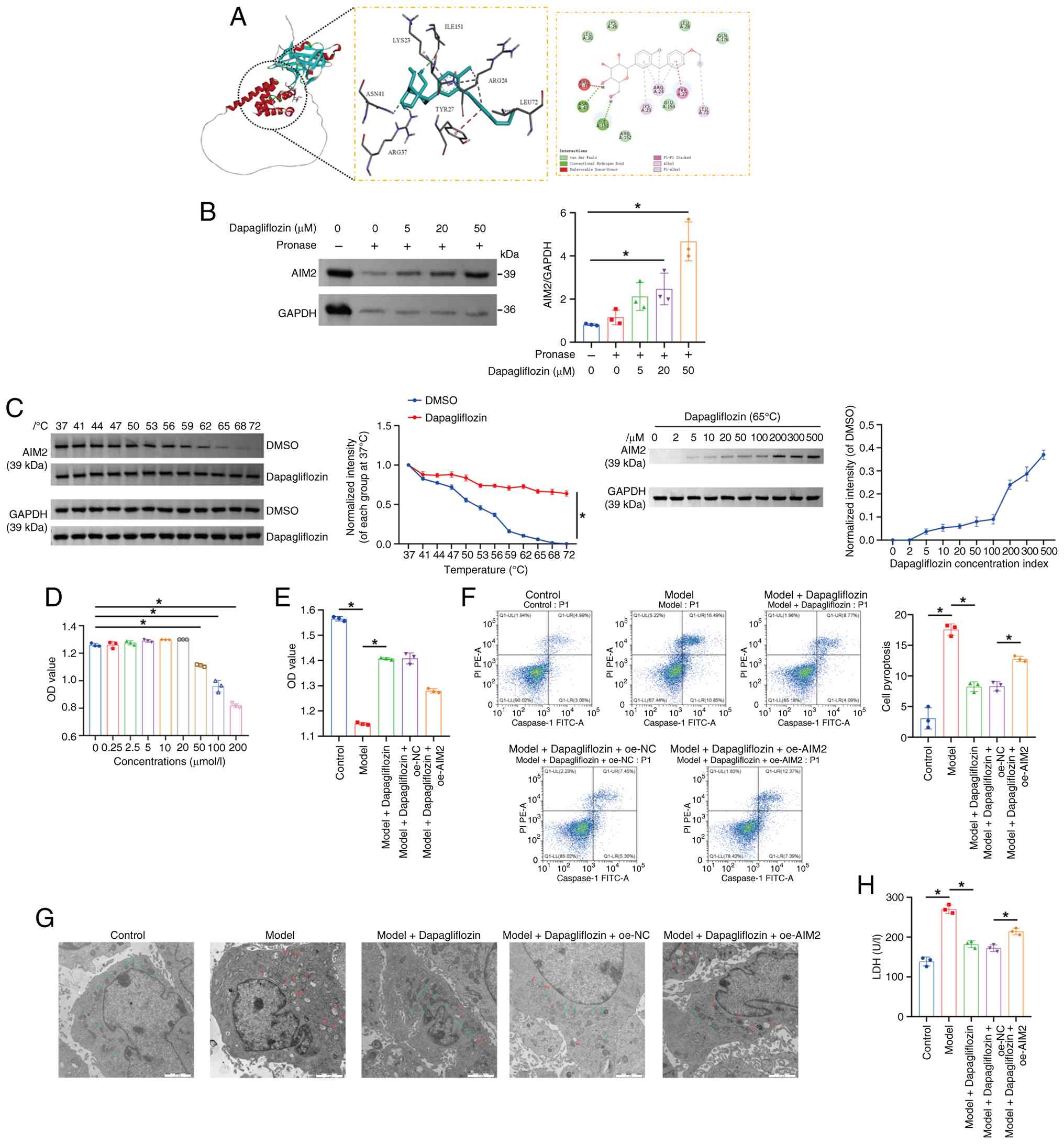
unclear. To explore whether dapagliflozin could bind to AIM2, the
present study first performed molecular docking analysis. The
molecular docking results showed that the binding energy between
dapagliflozin and AIM2 was −5.6 kcal/mol. As the binding energy was
lower than −5 kcal/mol, this suggested that the compound could form
relatively stable and favorable interactions with the protein
(Fig. 7A). The results of the
DARTS assay showed that dapagliflozin protected the AIM2 protein
from pronase digestion in a dose-dependent manner, indicating a
targeting association between dapagliflozin and the AIM2 protein
(Fig. 7B). CETSA results revealed
that specific binding of dapagliflozin to its target protein AIM2
markedly altered the thermal stability of AIM2 compared with the
free protein, as reflected by a marked downward shift in its
melting temperature. In living cells, dapagliflozin nevertheless
enhanced AIM2 stability. At a fixed temperature of 65°C, the amount
of soluble AIM2 increased in a concentration-dependent manner with
increasing concentrations of dapagliflozin (Fig. 7C). Next, mouse cardiomyocytes were
treated with different concentrations of dapagliflozin (0, 0.25,
2.5, 5, 10, 20, 50, 100 and 200 µmol/l) to determine the maximum
concentration that had no effect on cell viability. The results
showed that treatment with 50 µmol/l dapagliflozin had no
significant effect on cardiomyocyte viability (Fig. 7D). Therefore, this concentration
was selected for subsequent experiments. Compared with the Control
group, cardiomyocyte viability was lower, pyroptosis was greater
and mitochondrial damage was more pronounced in the Model group.
Following dapagliflozin treatment, cardiomyocyte viability
increased, while pyroptosis and mitochondrial damage were
decreased. However, overexpression of AIM2 reversed these effects
(Fig. 7E-G). Moreover, compared
with the Control group, LDH, pro-Caspase-1, Caspase-1, GSDMD,
GSDMD-N, AIM2, NLRP3, ASC, pro-IL-1β, pro-IL-18, IL-1β and IL-18
levels in the Model group were elevated. After dapagliflozin
treatment, LDH, pro-Caspase-1, Caspase-1, GSDMD, GSDMD-N, AIM2,
NLRP3, ASC, pro-IL-1β, pro-IL-18, IL-1β and IL-18 levels in mouse
cardiomyocytes decreased, while the overexpression of AIM2 reversed
this phenomenon (Figs. 7H and
8A-C). Furthermore, compared with
the Control group, the percentage of PI-positive cardiomyocytes was
greater in the Model group. After dapagliflozin treatment, the
percentage of PI-positive mouse cardiomyocytes decreased, whereas
AIM2 overexpression increased the proportion of PI-positive
cardiomyocytes (Fig. 8D). To
investigate whether dapagliflozin regulates GSDMD-N membrane
translocation through AIM2, western blot analysis was performed on
membrane and cytoplasmic fractions. As shown in the Fig. 8E, compared with the Control group,
the Model group exhibited markedly elevated relative expression
levels of GSDMD-N protein in the cell membrane fraction, while
cytoplasmic GSDMD-N levels were also markedly upregulated,
indicating that the model successfully induced GSDMD cleavage
activation and membrane translocation. Compared with the Model
group, both membrane and cytoplasmic GSDMD-N expression were
markedly suppressed in the Model + Dapagliflozin group; however,
upon overexpressing AIM2 on the basis of dapagliflozin intervention
(Model + Dapagliflozin + oe-AIM2 group), GSDMD-N membrane
translocation and cytoplasmic expression levels were markedly
rescued, demonstrating that AIM2 overexpression can reverse the
inhibitory effect of dapagliflozin on GSDMD activation. ATP1a
(membrane internal reference) and GAPDH (cytoplasmic internal
reference) showed stable expression across all groups, validating
the uniformity and reliability of subcellular fractionation and
loading. Taken together, these results suggested that dapagliflozin
inhibited the AIM2/Caspase-1/GSDMD axis and thereby alleviated
pyroptosis in mouse cardiomyocytes.
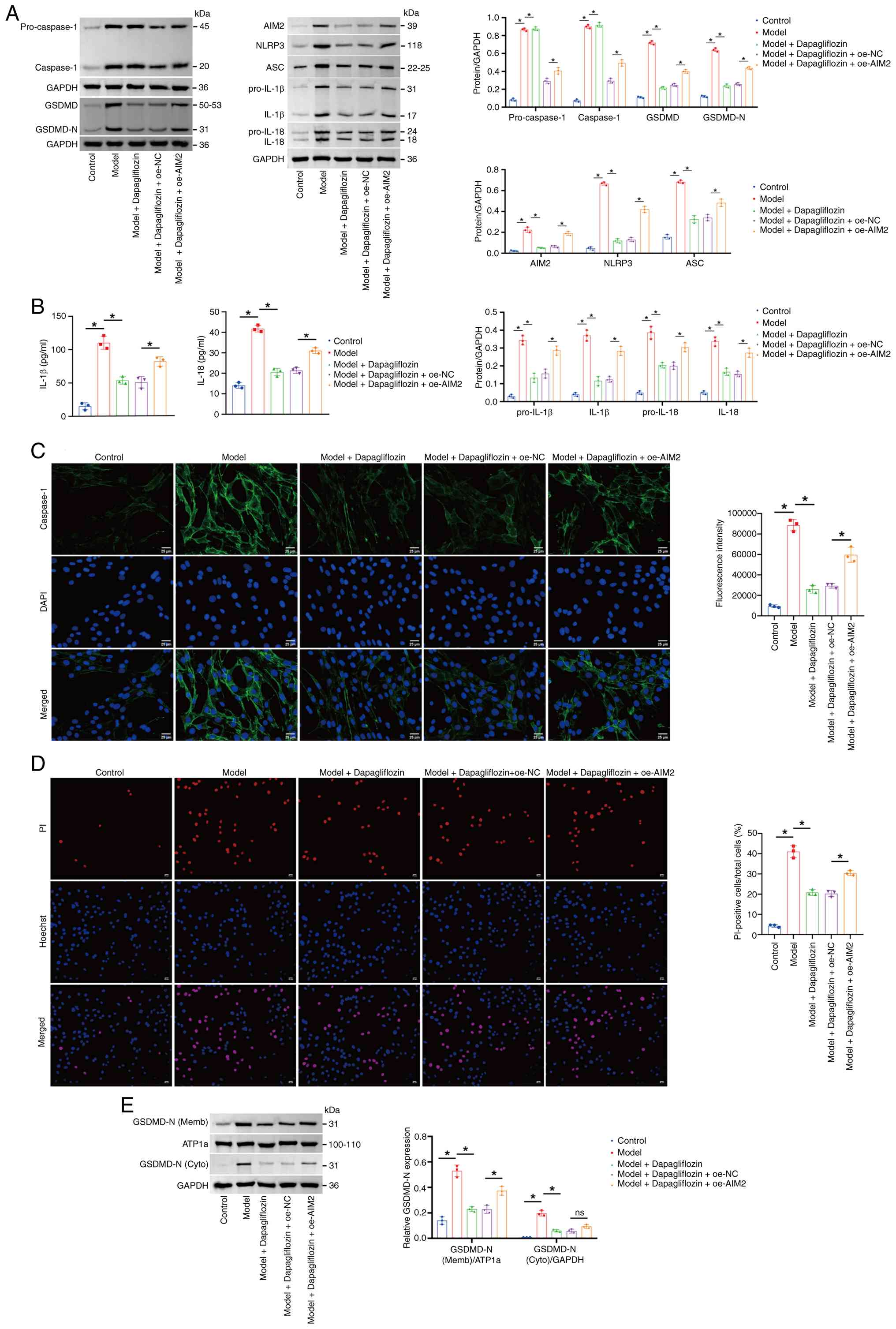 | Figure 8.Dapagliflozin inhibits the
AIM2/Caspase-1/GSDMD axis to alleviate pyroptosis in mouse
cardiomyocytes. (A) Western blotting of pro-Caspase-1, Caspase-1,
GSDMD, GSDMD-N, AIM2, NLRP3, ASC, pro-IL-1β, pro-IL-18, IL-1β and
IL-18 levels in the cell lysate. The blot was probed with
anti-GAPDH antibody. GAPDH was used as a loading control. (B) IL-1β
and IL-18 levels in the cell supernatant. (C) Immunofluorescence
staining of Caspase-1 in cells. (D) Hoechst 33342/PI fluorescence
staining showing the percentage of PI-positive mouse
cardiomyocytes. Scale bar, 50 µm, magnification, ×200; scale bar,
25 µm, magnification, ×400. (E) Western blotting of the levels of
GSDMD-N in the membrane and cytoplasm. The blot was probed with
anti-ATP1a/GAPDH antibody. ATP1a (membrane marker) and GAPDH
(cytoplasmic marker) were used as a loading control. *P<0.05.
AIM2, absent in melanoma 2; GSDMD, Gasdermin D; GSDMD-N, Gasdermin
D N-terminal fragment; NLRP3, NOD-, LRR- and pyrin
domain-containing protein 3; ASC, apoptosis-associated Speck-like
protein containing a CARD; ATP1a, sodium/potassium-transporting
ATPase subunit α1; oe, overexpressed; NC, negative control; sh,
short hairpin. |
Dapagliflozin inhibits the
AIM2-Caspase-1/GSDMD axis to improve HFpEF symptoms in mice
Finally, AIM2 was overexpressed in combination with
dapagliflozin treatment to explore further the underlying
mechanisms at the animal level. Fig.
9A shows representative left ventricular M-mode
echocardiographic images. The LVEF percentage remained essentially
consistent across all groups (Fig.
9B). Compared with the HFpEF group, the GLS percentage was
higher in the HFpEF + dapagliflozin group, whereas AIM2
overexpression reduced the GLS percentage (Fig. 9C). Compared with the HFpEF group,
the HFpEF + dapagliflozin group exhibited lower E/E' and HW/TL
ratios and a longer running distance in the treadmill exhaustion
test. Notably, AIM2 overexpression abolished these beneficial
effects, leading to increased E/E' and HW/TL ratios and a decreased
running distance (Fig. 9D-F).
Additionally, myocardial hypertrophy was reduced in the HFpEF +
dapagliflozin group compared with the HFpEF group, whereas AIM2
overexpression resulted in more severe myocardial hypertrophy
(Fig. 9G). Moreover, compared with
the HFpEF group, the levels of Caspase-1, GSDMD, GSDMD-N, AIM2,
NLRP3, ASC, pro-IL-1β, pro-IL-18, IL-1β and IL-18 in myocardial
tissue were decreased in the HFpEF + dapagliflozin group. After
AIM2 overexpression, the levels of pro-Caspase-1, Caspase-1, GSDMD,
GSDMD-N, AIM2, NLRP3, ASC, pro-IL-1β, pro-IL-18, IL-1β and IL-18
increased in myocardial tissue (Fig.
9H and I). Collectively, these results suggested that
dapagliflozin inhibited the AIM2-Caspase-1/GSDMD axis and thereby
improved HFpEF-related phenotypes in mice.
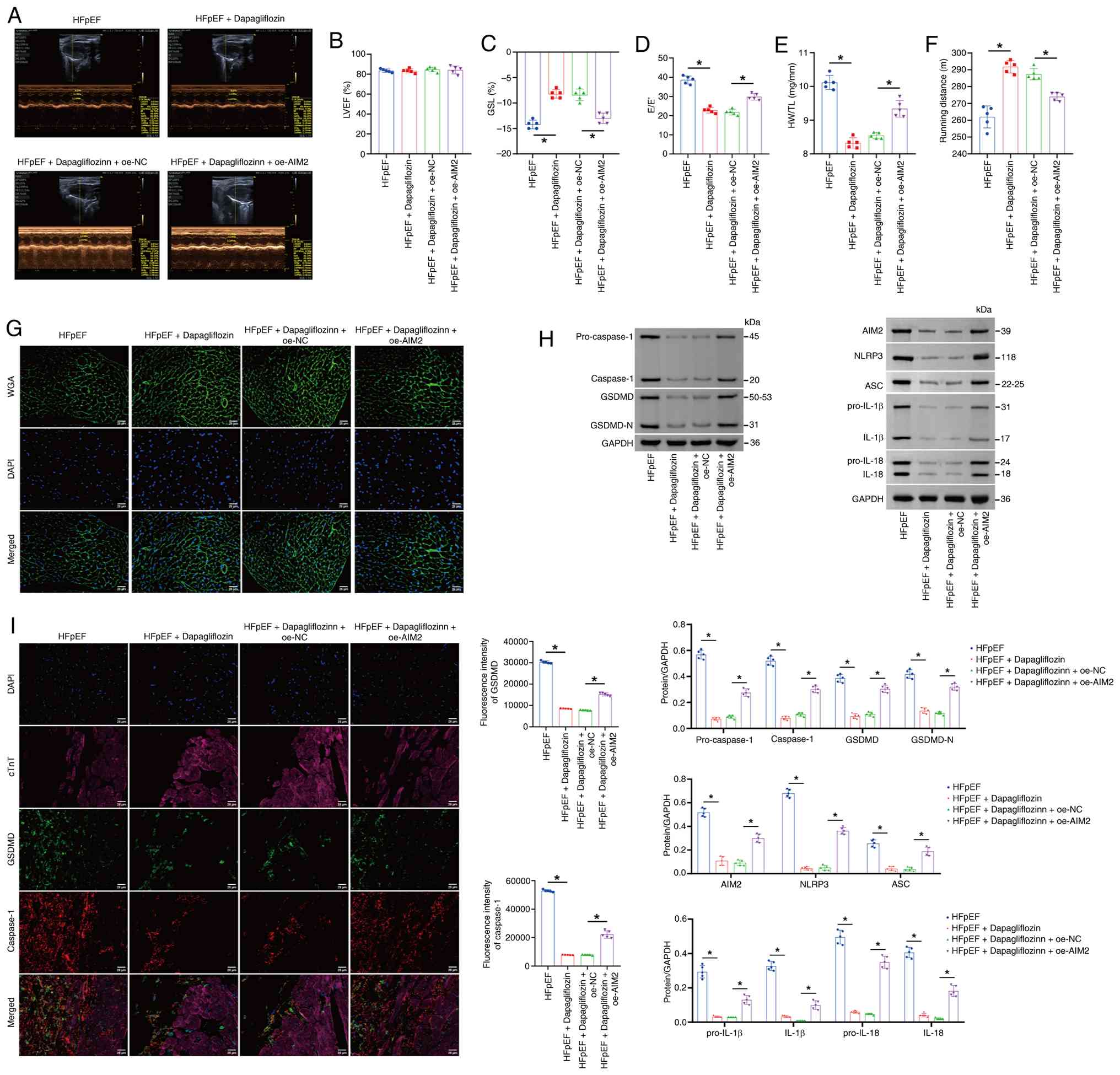 | Figure 9.Dapagliflozin inhibits the
AIM2-Caspase-1/GSDMD axis to improve HFpEF symptoms in mice. (A)
Representative left ventricular M-mode echocardiograph. (B) The
percentage of the LVEF. (C) The percentage of GLS. (D) The E/E'.
(E) HW/TL. (F) Exercise exhaustion test based on running distance.
(G) WGA staining of the morphological alterations in the myocardial
tissue cell membrane in the mice. (H) Western blot analysis of
pro-Caspase-1, Caspase-1, GSDMD, GSDMD-N, AIM2, NLRP3, ASC,
pro-IL-1β, pro-IL-18, IL-1β and IL-18 levels in myocardial tissue.
(I) Immunofluorescence staining showing the expression of GSDMD and
Caspase-1. Scale bar, 25 µm; magnification, ×400. The blot was
probed with anti-GAPDH antibody. GAPDH was used as a loading
control. *P<0.05. AIM2, absent in melanoma 2; GSDMD, Gasdermin
D; HFpEF, heart failure with preserved ejection fraction; LVEF,
left ventricular ejection fraction; GLS, global longitudinal
strain; E/E', ratio of the mitral E wave to the E' wave; HW/TL,
ratio of heart weight to tibia length; WGA, wheat germ agglutinin;
GSDMD-N, Gasdermin D N-terminal fragment; NLRP3, NOD-, LRR- and
pyrin domain-containing protein 3; ASC, apoptosis-associated
Speck-like protein containing a CARD; oe, overexpressed; NC,
negative control; sh, short hairpin. |
Discussion
With an aging population and the increasing
prevalence of obesity, diabetes and hypertension, HFpEF has become
the main form of HF worldwide (39). However, effective treatments for
HFpEF remain limited. Therefore, elucidating the mechanisms
underlying HFpEF may facilitate the identification of new
therapeutic approaches. The present study investigated the
mechanism by which dapagliflozin treats HFpEF, with a focus on
pyroptosis, primarily at the animal and cellular levels. It was
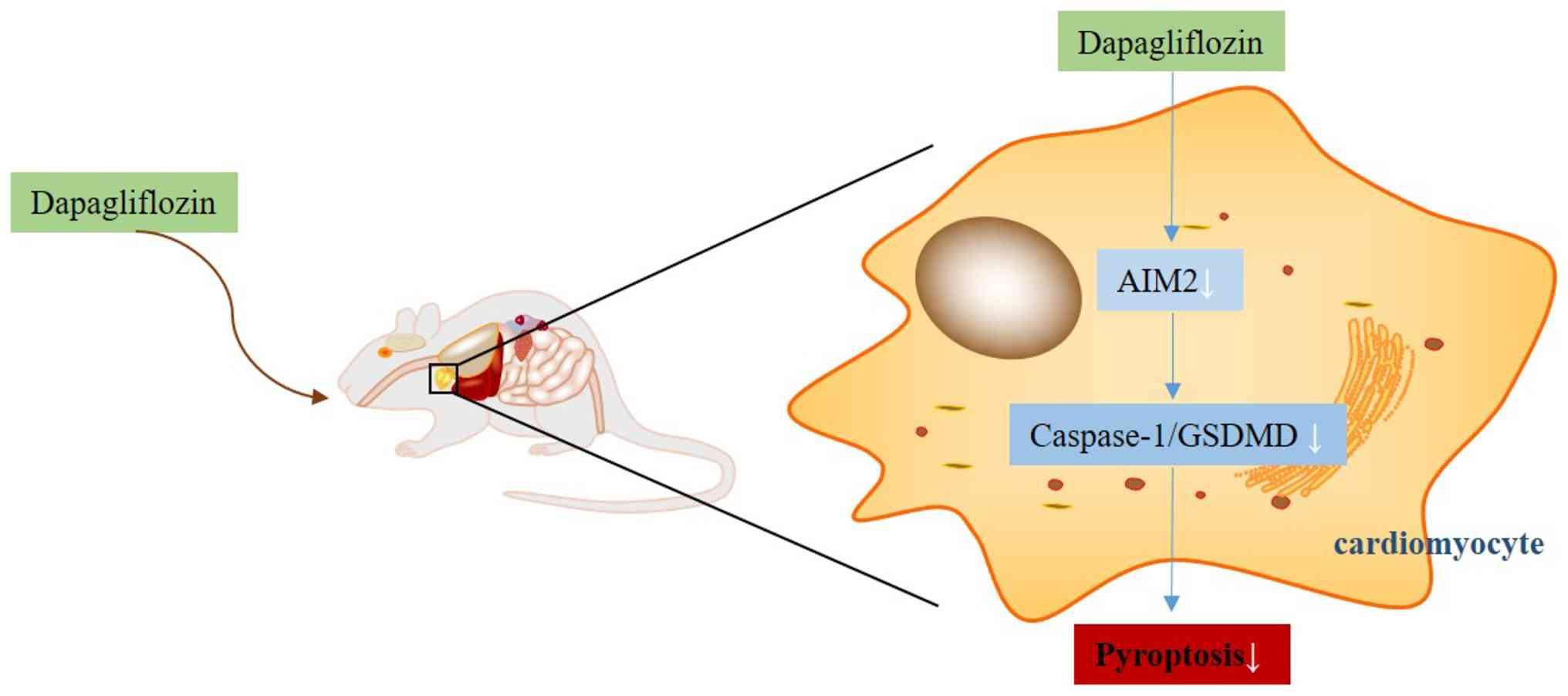
found that dapagliflozin may alleviate HFpEF through mechanisms
involving the AIM2/Caspase-1/GSDMD axis to attenuate pyroptosis
(Fig. 10). To the best of the
authors' knowledge, this is the first study to link dapagliflozin
to the regulation of the AIM2/Caspase-1/GSDMD axis in HFpEF.
SGLT2 inhibitors are used in the treatment of HF
(40). SGLT2 inhibitor therapy can
effectively alleviate symptoms and decrease overall hospitalization
rates in patients with HF (41).
Previous research has demonstrated that dapagliflozin, an SGLT2
inhibitor, can improve renal outcomes in patients with HF with
reduced ejection fraction and reduce all-cause and cardiovascular
mortality (42). In the
EMPEROR-Preserved trial, dapagliflozin decreased the risk of
cardiovascular death or hospitalization for HF and SGLT2 inhibitors
may become a new standard of care for patients with HFpEF (43). In addition, dapagliflozin has been
reported to prevent renal pyroptosis via regulation of the
miR-155-5p/HO-1/NLRP3 axis (44).
However, the mechanism by which dapagliflozin modulates pyroptosis
in HFpEF remains poorly understood. The present study not only
evaluated the effects of dapagliflozin on the pathological features
of HFpEF but also comprehensively monitored changes in body weight,
blood pressure and blood glucose levels. Mice receiving
dapagliflozin exhibited significant improvements in body weight,
blood pressure and blood glucose control. These findings suggested
that, in addition to its direct myocardial effects, dapagliflozin
may alleviate HFpEF-related pathological features by reducing body
weight, improving glucose metabolism and lowering blood pressure.
Importantly, these systemic effects may act in concert to
contribute to HFpEF improvement rather than reflecting a single
isolated mechanism. Furthermore, the present study demonstrated
that dapagliflozin alleviated HFpEF-related phenotypes and
suppressed myocardial pyroptosis in HFpEF mice. Accordingly, the
mechanism by which dapagliflozin regulates pyroptosis in HFpEF was
further explored.
In recent years, pyroptosis, as a form of programmed
cell death, has increasingly attracted attention because of its
involvement in inflammatory responses and cardiovascular diseases.
Pyroptosis can contribute to cardiac fibrosis, myocardial
hypertrophy, cardiomyocyte death, myocardial dysfunction, excessive
inflammation and cardiac remodeling (45,46).
Accordingly, targeting pyroptosis has been considered a promising
therapeutic strategy for HF treatment (47). The inflammasome is a multiprotein
complex that plays a critical role in the innate immune response by
promoting the cleavage and activation of IL-1β and IL-18 (48). Among these complexes, the AIM2
inflammasome functions as a multiprotein platform that detects
abnormal cytoplasmic dsDNA, thereby promoting cytokine maturation
and release and ultimately triggering pyroptosis (49). Moreover, the AIM2 inflammasome is
an important mediator of inflammatory signaling, as its activation
induces the production of the proinflammatory cytokines IL-1β and
IL-18 and contributes to the initiation of the pyroptotic response
involved in regulating excessive cell proliferation (50). In the present study, AIM2 protein
levels in myocardial tissue were markedly elevated in the HFpEF
group compared with the Sham group, whereas this increase was
reversed following dapagliflozin treatment. Notably, changes in
myocardial pyroptosis in HFpEF mice were accompanied by
corresponding fluctuations in AIM2 protein levels. Pearson
correlation analysis further showed that AIM2 levels were markedly
negatively correlated with GLS and markedly positively associated
with E/E', which supports a potential role for AIM2 in HFpEF.
Collectively, these findings suggested that AIM2 may exert
substantial effects on myocardial pyroptosis in HFpEF. From a
mechanistic perspective, inhibition of AIM2 activity may help
reduce myocardial injury and improve cardiac function; therefore,
further investigations were conducted to clarify the mechanisms
linking AIM2 activation to pyroptosis in this context.
Pioneering research has clarified key molecular
mechanisms of pyroptosis, particularly the central role of the
Caspase-1/GSDMD pathway in this form of inflammatory cell death
(7). AIM2, acting as a
pattern-recognition receptor, can activate Caspase-1 by sensing
cytosolic dsDNA, thereby initiating pyroptosis (10). Through its downstream effector
caspase, Caspase-1, the AIM2 inflammasome exerts important
biological functions, highlighting its potential involvement in
cardio-cerebrovascular diseases and related metabolic disorders
(51). In addition, activation of
GSDMD, a prototypical member of the gasdermin family, is closely
linked to inflammasome-dependent innate immune surveillance
(52). Upon activation by
inflammatory caspases such as Caspase-1 and Caspase-11, GSDMD is
cleaved to generate the pore-forming N-terminal fragment, which
disrupts plasma membrane integrity and ultimately drives pyroptosis
(53). Research has indicated
that, in chronic kidney disease, the AIM2-Caspase-1/GSDMD pathway
is involved in mediating renal cell pyroptosis (38) and inhibition of AIM2 inflammasome
activation, achieved by downregulating AIM2 and Caspase-1, has been
shown to alleviate pyroptosis in early brain injury following
subarachnoid hemorrhage induced by GSDMD (54). However, the functional mechanism of
the AIM2/Caspase-1/GSDMD axis in HFpEF remains largely elusive. In
the present study, the findings revealed that downregulation of
AIM2 alleviated pyroptosis in mouse cardiomyocytes at the cellular
level. Moreover, the data indicated that AIM2 regulated
cardiomyocyte pyroptosis through the Caspase-1/GSDMD axis.
Consistently, these results supported a crucial role for the
AIM2/Caspase-1/GSDMD axis in mediating myocardial pyroptosis in
HFpEF.
AIM2-associated pyroptosis has also been implicated
in other disease settings. For example, sevoflurane has been shown
to induce pyroptosis in mouse models of Alzheimer's disease by
activating Caspase-1 through the NLRP3 and AIM2 pathways, while
also facilitating GSDMD cleavage and the downstream production of
IL-1β and IL-18 (55).
Additionally, Talaromyces marneffei has been reported to
activate the AIM2-Caspase-1/-4/GSDMD pathway and thereby induce
pyroptosis in hepatocytes (56).
However, the mechanism by which dapagliflozin influences the
AIM2/Caspase-1/GSDMD axis and pyroptosis in HFpEF remains unclear.
The identification of AIM2 as a key regulator of pyroptosis in
HFpEF therefore represents an important finding of the present
study. In the context of HFpEF, upregulation of AIM2 may be
associated with exacerbated inflammatory responses that contribute
to cardiomyocyte injury and dysfunction. Mechanistically, AIM2
upregulation likely activates the Caspase-1/GSDMD axis to enhance
proinflammatory cytokine release, which impairs the contractile and
relaxational functions of cardiomyocytes. Moreover, AIM2-mediated
pyroptosis may promote cardiomyocyte loss, further aggravating the
pathological progression of HFpEF. Finally, the present study
demonstrated both in vitro and in vivo that
dapagliflozin-mediated inhibition of the AIM2/Caspase-1/GSDMD
pathway attenuated myocardial pyroptosis and improved HFpEF-related
phenotypes in mice.
AMPK plays an important role in energy metabolism
and cardiovascular protection (57). SGLT2 inhibitors have been shown to
activate AMPK, thereby improving cardiomyocyte energy metabolism
and function (58). Accordingly,
AMPK activation may alleviate energy stress and injury in
cardiomyocytes by promoting fatty acid oxidation and improving
mitochondrial function. Mitochondrial dysfunction is an important
pathophysiological characteristic of HFpEF (59) and SGLT2 inhibitors may enhance
energy metabolism and antioxidant capacity in cardiomyocytes, at
least in part, through improvements in mitochondrial function
(60). In the present study,
assessment of mitochondrial membrane potential showed that
dapagliflozin treatment markedly increased mitochondrial membrane
potential, indicating restoration of mitochondrial function. As
improved mitochondrial function can enhance cellular energy
metabolism and antioxidant capacity, it may consequently reduce
cardiomyocyte injury and inflammatory responses (61). However, further studies are still
required to clarify the specific mechanisms by which restored
mitochondrial function contributes to dapagliflozin-mediated
alleviation of HFpEF. Oxidative stress is also an important
pathophysiological factor in HfpEF (62). Therefore, reducing oxidative stress
may improve myocardial function by alleviating cardiomyocyte injury
and limiting inflammatory responses (63). Nevertheless, future studies are
needed to further define the contribution of oxidative stress
reduction to the amelioration of HFpEF by dapagliflozin.
However, the present study has some limitations.
While molecular docking, DARTS and CETSA analyses provide valuable
information regarding the potential binding sites and interaction
patterns between dapagliflozin and AIM2, they do not by themselves
directly demonstrate the actual physical binding of the two
molecules under physiological conditions. Further studies using
techniques such as surface plasmon resonance or isothermal
titration calorimetry will be required to definitively confirm
direct binding and characterize the binding affinity. Although the
HFpEF model induced by L-NAME plus a high-fat diet has been widely
used to investigate the pathophysiological mechanisms of HFpEF
(22,23) and successfully recapitulated key
features of HFpEF in the present study, such as diastolic
dysfunction and inflammatory responses, this model cannot fully
recapitulate the complex pathophysiological processes of human
HFpEF. Human HFpEF is a multifactorial disease whose pathogenesis
involves multiple comorbidities (such as hypertension, diabetes,
obesity and coronary heart disease) and the aging process (64). By contrast, the model induced by
L-NAME and a high-fat diet mainly mimics the adverse effects of
hypertension and metabolic disorders on the heart, but fails to
fully replicate all the pathophysiological features of human HFpEF.
Aging is one of the important risk factors for HFpEF and its effect
on cardiac structure and function plays a critical role in human
disease (65). However, the L-NAME
and high-fat diet-induced model is usually established in young
mice, which cannot fully mimic the cumulative aging-related damage
to the heart. Aging-related changes, such as increased
cardiomyocyte apoptosis, extracellular matrix remodeling and
mitochondrial dysfunction, are not adequately represented in this
model. Due to the aforementioned limitations of the L-NAME and
high-fat diet-induced model, the translational relevance of the
present study may be restricted to a certain extent. Although this
model provides valuable insights into the pathophysiological
mechanisms of HFpEF, it does not fully accurately mimic the human
disease. Therefore, caution is needed when translating these
findings into clinical applications. Future studies are required to
further explore animal models that more closely resemble the
pathophysiological characteristics of human HFpEF, so as to improve
the translational relevance of the research results. In addition,
the main limitation of the present study is that all experiments
were based on mouse models and in vitro cell lines, without
validation using human myocardial samples. Although mouse models
and cell lines provide valuable experimental platforms, they cannot
fully simulate the complex pathophysiological processes of human
HFpEF. Owing to the lack of validation in human samples, the
translational potential of the present findings into clinical
practice may be limited. While the results from mouse models and
cell lines offer important insights into the mechanism of action of
dapagliflozin, these findings need to be further validated in human
samples to ensure their clinical relevance and reproducibility.
Future studies should include more human myocardial samples to
evaluate the potential role of dapagliflozin in the treatment of
HFpEF. The present study focused on the therapeutic effects of
6-week dapagliflozin intervention in the HFpEF model. Although this
short-term intervention has revealed the significant beneficial
effects of dapagliflozin in improving cardiac function, inhibiting
inflammatory responses and pyroptosis, considering that HFpEF is a
chronic disease, the 6-week intervention period may be insufficient
to comprehensively evaluate the long-term effects of dapagliflozin.
Long-term intervention studies are essential for assessing the
effects of dapagliflozin on cardiac remodeling, survival rate and
sustained suppression of pyroptosis. Given the relatively short
intervention period in the present study, it is acknowledged that
this may limit the comprehensive evaluation of the long-term
effects of dapagliflozin. Future studies need to design longer-term
intervention experiments for improved understanding of the
long-term mechanisms and clinical potential of dapagliflozin in the
treatment of HFpEF. Due to equipment and experimental conditions,
SEM images showing membrane blebbing or pore structures have not
yet been obtained. Although the supplementary western blotting
results confirmed GSDMD-N translocation to the plasma membrane
(indirect evidence of pore formation), direct SEM evidence of
plasma membrane pore structures is still lacking. SEM experiments
will be further supplemented in future studies to provide more
direct evidence for GSDMD-mediated plasma membrane damage.
Furthermore, the present study did not use
pharmacological AIM2 inflammasome activators or inhibitors to
verify the role of this pathway in mediating the therapeutic effect
of dapagliflozin. This was mainly due to the lack of highly
specific drugs targeting the AIM2 inflammasome and the constraints
of experimental conditions. The existing non-specific inflammasome
inhibitors (such as Shikonin) mostly act by interfering with the
assembly process of the AIM2 inflammasome complex and directly
inhibiting Caspase-1 activity, without changing the expression
level of AIM2 protein itself. They cannot accurately simulate the
protein level-dependent regulatory effect achieved by AIM2
knockdown/overexpression in the present study, making it difficult
to accurately verify the core role of AIM2 protein level in the
therapeutic mechanism of dapagliflozin (66,67).
At the same time, the screening of appropriate drug concentrations
and administration methods requires a large number of preliminary
pre-experiments, which cannot be completed in time for this
revision due to the limitations of the current experimental cycle
and resource conditions. Although the rescue experiments (AIM2
overexpression/knockdown) have provided indirect evidence for the
involvement of the AIM2 pathway, there is still a lack of direct
pharmacological verification and the potential interference of
other inflammasome subtypes (such as NLRP3 and NLRC4) cannot be
completely excluded, which also brings certain limitations to the
conclusions of the present study. Moreover, the present study only
focused on the regulatory role of the AIM2/Caspase-1/GSDMD axis and
did not further explore the upstream molecular mechanism by which
dapagliflozin regulates AIM2 expression, so it is impossible to
clarify whether dapagliflozin indirectly regulates AIM2 level
through other signaling pathways, which is also a direction that
needs further improvement in future studies. Although the rescue
experiments (AIM2 overexpression/knockdown) provided indirect
evidence for the involvement of the AIM2 pathway, there is still a
lack of direct pharmacological verification. Future studies will
actively seek specific AIM2 inflammasome activators and inhibitors
and clarify the upstream mechanism of dapagliflozin regulating AIM2
expression and perform relevant experiments to further confirm the
causal relationship between the AIM2 inflammasome pathway and the
therapeutic effect of dapagliflozin. At the same time, the sample
size will be expanded, the experimental design optimized, potential
interference factors reduced and the reliability and universality
of the research conclusions improved.
In conclusion, the findings of the present study
indicated that dapagliflozin may be beneficial for improving HFpEF.
The present study established both in vitro and in
vivo HFpEF models and administered dapagliflozin, thereby
demonstrating that dapagliflozin may alleviate HFpEF through
mechanisms involving the AIM2/Caspase-1/GSDMD axis to attenuate
pyroptosis. Further clinical trials and experimental studies are
needed to deepen our understanding of the regulatory mechanisms of
dapagliflozin and to facilitate its broader clinical application in
the management of HFpEF.
Supplementary Material
Supporting Data
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by Natural Science Foundation of
Hunan (grant no. 2023JJ60408).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
LX and XZ contributed to the study conception and
design. Material preparation, data collection and analysis were
performed by SD, HY, TL and YL. The first draft of the manuscript
was written by SD. LX and XZ confirm the authenticity of all the
raw data. All authors commented on previous versions of the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All experimental protocols were approved by the
Animal Ethics Committee of Changsha Hospital of Traditional Chinese
Medicine (approval no. 2022111003). The present study is in
accordance with the ARRIVE guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Daou D, Gillette TG and Hill JA:
Inflammatory mechanisms in heart failure with preserved ejection
fraction. Physiology (Bethesda). 38:02023.PubMed/NCBI
|
|
2
|
Borlaug BA, Borlaug BA, Sharma K, Shah SJ
and Ho JE: Heart failure with preserved ejection fraction: JACC
scientific statement. J Am Coll Cardiol. 81:1810–1834. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
von Haehling S, Assmus B, Bekfani T,
Dworatzek E, Edelmann F, Hashemi D, Hellenkamp K, Kempf T, Raake P,
Schütt KA, et al: Heart failure with preserved ejection fraction:
Diagnosis, risk assessment and treatment. Clin Res Cardiol.
113:1287–1305. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gao S, Liu XP, Li TT, Chen L, Feng YP,
Wang YK, Yin YJ, Little PJ, Wu XQ, Xu SW and Jiang XD: Animal
models of heart failure with preserved ejection fraction (HFpEF):
from metabolic pathobiology to drug discovery. Acta Pharmacol Sin.
45:23–35. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Omote K, Verbrugge FH and Borlaug BA:
Heart failure with preserved ejection fraction: Mechanisms and
treatment strategies. Annu Rev Med. 73:321–337. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cookson BT and Brennan MA:
Pro-inflammatory programmed cell death. Trends Microbiol.
9:113–114. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shao F: Gasdermins: Making pores for
pyroptosis. Nat Rev Immunol. 21:620–621. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen X, Tian PC and Wang K, Wang M and
Wang K: Pyroptosis: Role and mechanisms in cardiovascular disease.
Front Cardiovasc Med. 9:8978152022. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhao ZZ, Zheng XL and Jiang ZS: Emerging
roles of absent in melanoma 2 in cardiovascular diseases. Clin Chim
Acta. 511:14–23. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hornung V, Ablasser A, Charrel-Dennis M,
Bauernfeind F, Horvath G, Caffrey DR, Latz E and Fitzgerald KA:
AIM2 recognizes cytosolic dsDNA and forms a caspase-1-activating
inflammasome with ASC. Nature. 458:514–518. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jin T, Perry A, Jiang J, Smith P, Curry
JA, Unterholzner L, Jiang Z, Horvath G, Rathinam VA, Johnstone RW,
et al: Structures of the HIN domain:DNA complexes reveal ligand
binding and activation mechanisms of the AIM2 inflammasome and
IFI16 receptor. Immunity. 36:561–571. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Du L, Wang X, Chen S and Guo X: The AIM2
inflammasome: A novel biomarker and target in cardiovascular
disease. Pharmacol Res. 186:1065332022. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Onódi Z, Ruppert M, Kucsera D, Sayour AA,
Tóth VE, Koncsos G, Novák J, Brenner GB, Makkos A, Baranyai T, et
al: AIM2-driven inflammasome activation in heart failure.
Cardiovasc Res. 117:2639–2651. 2021.PubMed/NCBI
|
|
14
|
Liu H, Liu X, Zhou J and Li T:
Mitochondrial DNA is a vital driving force in ischemia-reperfusion
injury in cardiovascular diseases. Oxid Med Cell Longev.
2022:62357472022. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xia YY, Shi Y, Li Z, Li H, Wu LD, Zhou WY,
Gu Y, Ling ZY, Zhang JX and Chen SL: Involvement of pyroptosis
pathway in epicardial adipose tissue-myocardium axis in
experimental heart failure with preserved ejection fraction.
Biochem Biophys Res Commun. 636((Pt 2)): 62–70. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
El-Rous MA, Saber S, Raafat EM and Ahmed
AAE: Dapagliflozin, an SGLT2 inhibitor, ameliorates acetic
acid-induced colitis in rats by targeting NFκB/AMPK/NLRP3 axis.
Inflammopharmacology. 29:1169–1185. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang Z, Ni P, Tang M, Song Y, Liu C and
Zhao B: Dapagliflozin alleviates renal podocyte pyroptosis via
regulation of the HO-1/NLRP3 axis. Mol Med Rep. 28:2002023.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li H, Zhao Q, Liu D, Zhou B, Gong C and
Zhao G: Dapagliflozin attenuates NLRP3/Caspase-1 signaling
pathway-mediated pyroptosis of vascular smooth muscle cells by
downregulating CTSB. Altern Ther Health Med. 29:384–392.
2023.PubMed/NCBI
|
|
19
|
Nassif ME, Windsor SL, Borlaug BA, Kitzman
DW, Shah SJ, Tang F, Khariton Y, Malik AO, Khumri T, Umpierrez G,
et al: The SGLT2 inhibitor dapagliflozin in heart failure with
preserved ejection fraction: A multicenter randomized trial. Nat
Med. 27:1954–1960. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li Y, Xiang Y, Zhang S, Wang Y, Yang J,
Liu W and Xue F: Intramyocardial injection of thioredoxin
2-expressing lentivirus alleviates myocardial ischemia-reperfusion
injury in rats. Am J Transl Res. 9:4428–4439. 2017.PubMed/NCBI
|
|
21
|
Heidenreich PA, Bozkurt B, Aguilar D,
Allen LA, Byun JJ, Colvin MM, Deswal A, Drazner MH, Dunlay SM,
Evers LR, et al: 2022 AHA/ACC/HFSA guideline for the management of
heart failure: A report of the American College of
Cardiology/American Heart association joint committee on clinical
practice guidelines. J Am Coll Cardiol. 79:e263–e421. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tong D, Schiattarella GG, Jiang N,
Altamirano F, Szweda PA, Elnwasany A, Lee DI, Yoo H, Kass DA,
Szweda LI, et al: NAD(+) Repletion reverses heart failure with
preserved ejection fraction. Circ Res. 128:1629–1641. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Matsiukevich D, Kovacs A, Li T,
Kokkonen-Simon K, Matkovich SJ, Oladipupo SS and Ornitz DM:
Characterization of a robust mouse model of heart failure with
preserved ejection fraction. Am J Physiol Heart Circ Physiol.
325:H203–H231. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dawuti A, Sun S, Wang R, Gong D, Liu R,
Kong D, Yuan T, Zhou J, Lu Y, Wang S, et al: Salvianolic acid A
alleviates heart failure with preserved ejection fraction via
regulating TLR/Myd88/TRAF/NF-κB and p38MAPK/CREB signaling
pathways. Biomed Pharmacother. 168:1158372023. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wei R, Cui X, Feng J, Gu L, Lang S, Wei T,
Yang J, Liu J, Le Y, Wang H, et al: Dapagliflozin promotes beta
cell regeneration by inducing pancreatic endocrine cell phenotype
conversion in type 2 diabetic mice. Metabolism. 111:1543242020.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Schiattarella GG, Altamirano F, Tong D,
French KM, Villalobos E, Kim SY, Luo X, Jiang N, May HI, Wang ZV,
et al: Nitrosative stress drives heart failure with preserved
ejection fraction. Nature. 568:351–356. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sun Q, Güven B, Wagg CS, Almeida de
Oliveira A, Silver H, Zhang L, Chen B, Wei K, Ketema EB, Karwi QG,
et al: Mitochondrial fatty acid oxidation is the major source of
cardiac adenosine triphosphate production in heart failure with
preserved ejection fraction. Cardiovasc Res. 120:360–371. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhuo FF, Li L, Liu TT, Liang XM, Yang Z,
Zheng YZ, Luo QW, Lu JH, Liu D, Zeng KW and Tu PF: Lycorine
promotes IDH1 acetylation to induce mitochondrial dynamics
imbalance in colorectal cancer cells. Cancer Lett. 573:2163642023.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li F, Zhang Z, Shi Q, Wang R, Ji M, Chen
X, Li Y, Liu Y and Yu S: Thermal proteome profiling (TPP) reveals
NAMPT as the anti-glioma target of phenanthroindolizidine alkaloid
PF403. Acta Pharm Sin B. 15:2008–2023. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yudovich D, Bäckström A, Schmiderer L,
Žemaitis K, Subramaniam A and Larsson J: Combined lentiviral- and
RNA-mediated CRISPR/Cas9 delivery for efficient and traceable gene
editing in human hematopoietic stem and progenitor cells. Sci Rep.
10:223932020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Venâncio C, Félix L, Almeida V, Coutinho
J, Antunes L, Peixoto F and Summavielle T: Acute ketamine impairs
mitochondrial function and promotes superoxide dismutase activity
in the rat brain. Anesth Analg. 120:320–328. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fay MP and Proschan A:
Wilcoxon-Mann-Whitney or t-test? On assumptions for hypothesis
tests and multiple interpretations of decision rules. Stat Surv.
4:1–39. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zimmerman DW and Zumbo BD: Parametric
alternatives to the student T test under violation of normality and
homogeneity of variance. Perceptual and Motor Skills. 74:835–844.
1992. View Article : Google Scholar
|
|
35
|
Wang X, Wang Z and Tang D: Aerobic
exercise improves LPS-induced sepsis via regulating the Warburg
effect in mice. Sci Rep. 11:177722021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Meah VL, Backx K, Cockcroft JR, Shave RE
and Stöhr EJ: Left ventricular mechanics in late second trimester
of healthy pregnancy. Ultrasound Obstet Gynecol. 54:350–358. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rovetta A: Raiders of the lost
correlation: A guide on using pearson and spearman coefficients to
detect hidden correlations in medical sciences. Cureus.
12:e117942020.PubMed/NCBI
|
|
38
|
Ding B, Ma G, Wang Z, Liang W and Gao W:
Mechanisms of kidney cell pyroptosis in chronic kidney disease and
the effects of traditional Chinese medicine. Evid Based Complement
Alternat Med. 2021:11733242021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Borlaug BA: Evaluation and management of
heart failure with preserved ejection fraction. Nat Rev Cardiol.
17:559–573. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pabel S, Hamdani N, Luedde M and Sossalla
S: SGLT2 inhibitors and their mode of action in heart failure-has
the mystery been unravelled? Curr Heart Fail Rep. 18:315–328. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Starr JA and Pinner NA: The impact of
SGLT2 inhibitors on cardiovascular outcomes in patients with heart
failure with preserved ejection fraction. Ann Pharmacother.
58:506–513. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zannad F, Ferreira JP, Pocock SJ, Anker
SD, Butler J, Filippatos G, Brueckmann M, Ofstad AP, Pfarr E, Jamal
W and Packer M: SGLT2 inhibitors in patients with heart failure
with reduced ejection fraction: A meta-analysis of the
EMPEROR-Reduced and DAPA-HF trials. Lancet. 396:819–829. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jhund PS: SGLT2 inhibitors and heart
failure with preserved ejection fraction. Heart Fail Clin.
18:579–586. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhang ZW, Tang MQ, Liu W, Song Y, Gao MJ,
Ni P, Zhang DD, Mo QG and Zhao BQ: Dapagliflozin prevents kidney
podocytes pyroptosis via miR-155-5p/HO-1/NLRP3 axis modulation. Int
Immunopharmacol. 131:1117852024. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chai R, Xue W, Shi S, Zhou Y, Du Y, Li Y,
Song Q, Wu H and Hu Y: Cardiac remodeling in heart failure: Role of
pyroptosis and its therapeutic implications. Front Cardiovasc Med.
9:8709242022. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wu D, Deng D and Tang B: Programmed cell
death in heart failure: Mechanisms, impacts, and therapeutic
prospects. Rev Cardiovasc Med. 26:384072025. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Qin J, Yang Q, Wang Y, Shi M, Zhao X and
Zhou Y: The role of pyroptosis in heart failure and related
traditional chinese medicine treatments. Front Pharmacol.
15:13773592024. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wu J, Dong E, Zhang Y and Xiao H: The role
of the inflammasome in heart failure. Front Physiol. 12:7097032021.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang B and Yin Q: AIM2 inflammasome
activation and regulation: A structural perspective. J Struct Biol.
200:279–282. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Uresti-Rivera EE and García-Hernández MH:
AIM2-inflammasome role in systemic lupus erythematous and
rheumatoid arthritis. Autoimmunity. 55:443–454. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Lin J, Wang J, Fang J, Li M, Xu S, Little
PJ, Zhang D and Liu Z: The cytoplasmic sensor, the AIM2
inflammasome: A precise therapeutic target in vascular and
metabolic diseases. Br J Pharmacol. 181:1695–1719. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Vasudevan SO, Behl B and Rathinam VA:
Pyroptosis-induced inflammation and tissue damage. Semin Immunol.
69:1017812023. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhu Q, Zheng M, Balakrishnan A, Karki R
and Kanneganti TD: Gasdermin D promotes AIM2 inflammasome
activation and is required for host protection against Francisella
novicida. J Immunol. 201:3662–3668. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yuan B, Zhou XM, You ZQ, Xu WD, Fan JM,
Chen SJ, Han YL, Wu Q and Zhang X: Inhibition of AIM2 inflammasome
activation alleviates GSDMD-induced pyroptosis in early brain
injury after subarachnoid haemorrhage. Cell Death Dis. 11:762020.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Tian D, Xing Y, Gao W, Zhang H, Song Y,
Tian Y and Dai Z: Sevoflurane aggravates the progress of
Alzheimer's disease through NLRP3/Caspase-1/Gasdermin D pathway.
Front Cell Dev Biol. 9:8014222022. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wang G, Wei W, Jiang Z, Jiang J, Han J,
Zhang H, Hu J, Zhang P, Li X, Chen T, et al: Talaromyces marneffei
activates the AIM2-caspase-1/-4-GSDMD axis to induce pyroptosis in
hepatocytes. Virulence. 13:963–979. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zaha VG and Young LH: AMP-activated
protein kinase regulation and biological actions in the heart. Circ
Res. 111:800–814. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Yesilyurt-Dirican ZE, Qi C, Wang YC, Simm
A, Deelen L, Hafiz Abbas Gasim A, Lewis-McDougall F and
Ellison-Hughes GM: SGLT2 inhibitors as a novel senotherapeutic
approach. NPJ Aging. 11:352025. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Schauer A, Adams V, Kämmerer S, Langner E,
Augstein A, Barthel P, Männel A, Fabig G, Alves PKN, Günscht M, et
al: Empagliflozin improves diastolic function in HFpEF by
restabilizing the mitochondrial respiratory chain. Circ Heart Fail.
17:e0111072024. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Maejima Y: SGLT2 inhibitors play a
salutary role in heart failure via modulation of the mitochondrial
function. Front Cardiovasc Med. 6:1862020. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Chu Q, He D, Xie W, Li S, Dong Z and Fu X:
Gelated microvesicle-mediated delivery of mesenchymal stem cell
mitochondria for the treatment of myocardial infarction. Proc Natl
Acad Sci USA. 122:e24245291222025. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Tsutsui H, Kinugawa S and Matsushima S:
Oxidative stress and heart failure. Am J Physiol Heart Circ
Physiol. 301:H2181–H2190. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Han YL, Yan TT, Li HX, Chen SS, Zhang ZZ,
Wang MY, Chen MJ, Chen YL, Yang XX, Wei LL, et al: Geniposide
alleviates heart failure with preserved ejection fraction in mice
by regulating cardiac oxidative stress via MMP2/SIRT1/GSK3β
pathway. Acta Pharmacol Sin. 45:2567–2578. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Peikert A, Vacca A, Norata GD,
Schiattarella GG and Osto E: cellular interactions and
immunometabolic mechanisms in heart failure with preserved ejection
fraction: From molecular mechanisms to clinical evidence. Circ
Heart Fail. 19:e0126742026. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Ono R and Menezes Falcão L: Primary heart
failure with preserved ejection fraction: The intertwined
pathophysiology and treatment of some of the most relevant
phenotypes. Rev Port Cardiol. 44:445–456. 2025.(In English,
Portuguese). View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Zorman J, Sušjan P and Hafner-Bratkovič I:
Shikonin suppresses NLRP3 and AIM2 inflammasomes by direct
inhibition of caspase-1. PLoS One. 11:e01598262016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Xie M, Yu Y, Kang R, Zhu S, Yang L, Zeng
L, Sun X, Yang M, Billiar TR, Wang H, et al: PKM2-dependent
glycolysis promotes NLRP3 and AIM2 inflammasome activation. Nat
Commun. 7:132802016. View Article : Google Scholar : PubMed/NCBI
|