Introduction
Among all cancer types, lung cancer has the highest
incidence and is the most common cause of cancer-related deaths
worldwide (1). Non-small cell lung
cancer (NSCLC) constitutes >85% of lung cancer cases, with lung
adenocarcinoma (LUAD) representing the most prevalent pathological
subtype (2). Despite notable
advances in targeted therapy and immunotherapy in recent years, the
survival rate for patients with LUAD remains <20% (3). Furthermore, LUAD is a highly
heterogeneous tumor (4) and
different patients with the same subtype may exhibit marked
variations in response to the same treatment (5,6).
Therefore, exploring the molecular mechanisms underlying LUAD
progression and identifying reliable biomarkers are essential for
developing individualized treatment and improving patient survival
outcomes.
Long non-coding RNAs (lncRNAs) are a class of RNA
transcripts >200 nucleotides in length that lack protein-coding
capacity (7). Accumulating
evidence has confirmed that lncRNAs participate in diverse
biological processes through mechanisms such as transcriptional
regulation, post-transcriptional modulation and epigenetic
modification (8–12). Extensive research has also shown
that lncRNAs modulate critical processes such as cell
proliferation, metastasis and drug resistance, thereby either
promoting or suppressing tumor development (13–16).
Notably, certain lncRNAs have shown promise as diagnostic or
prognostic biomarkers, and have the potential to serve as
therapeutic targets (17–19). Therefore, investigating the role of
lncRNAs in tumors is critical for elucidating pathogenesis and
advancing personalized therapeutic strategies.
Long intergenic non-protein coding RNA 1117
(LINC01117), located at chromosome 2q31.1, is a relatively novel
lncRNA that has only been investigated in a limited number of
studies thus far. One study has suggested that LINC01117 may be
included in a prognostic gene signature for breast cancer (20). A recent study reported that, in
postmenopausal patients with uterine fibroids, LINC01117 was
markedly downregulated in fibroid tissues compared with in the
adjacent myometrium, whereas no notable change was observed between
these tissues in premenopausal patients with uterine fibroids
(21). Another study has
demonstrated that LINC01117 may promote the migration of LUAD cells
(22). However, the molecular
mechanisms by which LINC01117 may promote malignant proliferation
in LUAD remains unclear.
lncRNAs are closely associated with protein-coding
genes (PCGs) and regulate various biological processes through
these interactions (23). Homeobox
D8 (HOXD8), a transcription factor located on chromosome 2q31.1, is
positioned upstream of LINC01117. The function of HOXD8 in
tumorigenesis remains controversial; while some studies have
proposed that it acts as a tumor suppressor and inhibits cancer
progression (24–26), others have indicated that it may
function as an oncogene. For example, HOXD8 hypermethylation has
been identified as a highly sensitive and specific biomarker for
biliary tract cancer (27) and one
study has demonstrated its elevated expression in NSCLC tissues,
where it may promote proliferation, cancer stemness and migratory
abilities (28). Given their
genomic proximity, further investigation into the functional
relationship between LINC01117 and HOXD8 is warranted. Therefore,
the present study aimed to investigate the functional roles of
LINC01117 in LUAD and explore its regulatory mechanism involving
HOXD8.
Materials and methods
Data acquisition from The Cancer
Genome Atlas (TCGA) database
In the present study, RNA sequencing (RNA-seq) data
and the related clinical characteristics for 535 LUAD and 502 lung
squamous cell carcinoma (LUSC) tumor tissues, along with 59
non-tumor control samples from patients in TCGA-LUAD cohort and 49
non-tumor control samples from patients in TCGA-LUSC cohort, were
acquired from TCGA database (https://portal.gdc.cancer.gov/). Part of the
bioinformatics analyses was performed using R software (version
4.4.2; http://www.r-project.org/) (29). The R package ‘edgeR’ (version
4.4.2) was used to perform differential expression analysis on
these RNA-seq data (30).
Differential expression was assessed based on log2 fold change and
false discovery rate. Patients with an overall survival (OS) time
of <60 days were excluded to minimize bias from
non-cancer-related deaths or perioperative mortality, which may not
accurately represent the true disease prognosis. After excluding
patients with an OS of <60 days or incomplete clinical
characteristics, 448 patients with LUAD and 444 patients with LUSC
were retained for subsequent analysis. The evaluated clinical
parameters included survival status, sex, age, tumor size (T)
stage, lymph node metastasis (N) stage and tumor-node-metastasis
(TNM) stage. The clinical characteristics of patients from TCGA
cohort are listed in Table I.
 | Table I.Clinical characteristics of patients
in The Cancer Genome Atlas cohort. |
Table I.
Clinical characteristics of patients
in The Cancer Genome Atlas cohort.
|
| Patients with LUAD
(n=448) | Patients with LUSC
(n=444) |
|---|
|
|
|
|
|---|
| Clinical
factor | L-group
(n=224) | H-group
(n=224) | L-group
(n=222) | H-group
(n=222) |
|---|
| Vital status |
|
|
|
|
|
Alive | 165 (73.7) | 134 (59.8) | 138 (62.2) | 133 (59.9) |
|
Dead | 59 (26.3) | 90 (40.2) | 84 (37.8) | 89 (40.1) |
| Age, years |
|
|
|
|
|
≤65 | 104 (46.4) | 109 (48.7) | 92 (41.4) | 81 (36.5) |
|
>65 | 120 (53.6) | 115 (51.3) | 130 (58.5) | 141 (63.5) |
| Sex |
|
|
|
|
|
Female | 116 (51.8) | 124 (55.4) | 58 (26.1) | 57 (25.7) |
|
Male | 108 (48.2) | 100 (44.6) | 164 (73.9) | 165 (74.3) |
| Stage |
|
|
|
|
| 1 | 124 (55.4) | 117 (52.2) | 106 (47.7) | 106 (47.7) |
| 2 | 52 (23.2) | 59 (26.3) | 79 (35.6) | 70 (31.5) |
|
3/4 | 48 (21.4) | 48 (21.4) | 37 (16.7) | 46 (20.7) |
| T stage |
|
|
|
|
| 1 | 80 (35.7) | 71 (31.7) | 47 (21.2) | 53 (23.9) |
| 2 | 121 (54.0) | 120 (53.6) | 134 (60.4) | 126 (56.7) |
|
3/4 | 23 (10.3) | 33 (14.7) | 41 (18.5) | 43 (19.4) |
| N stage |
|
|
|
|
| 0 | 151 (67.4) | 143 (63.8) | 142 (64.0) | 141 (63.5) |
| 1 | 38 (17.0) | 49 (21.9) | 62 (27.9) | 58 (26.1) |
| 2 | 35 (15.6) | 32 (14.3) | 18 (8.1) | 23 (10.4) |
Survival analyses
Survival analyses were performed using the
Kaplan-Meier (KM) method and Cox proportional hazards regression.
Patients with LUAD from TCGA were stratified into high- and
low-expression groups using the median LINC01117 expression as the
cutoff. The median expression level was 3.44 in LUAD and 12.59 in
LUSC. OS differences were assessed using the two-sided log-rank
test. KM curves were fitted using the ‘survival’ package (version
3.7.0; http://CRAN.R-project.org/package=survival) (31) and visualized using ‘survminer’
(version 0.5.1; http://CRAN.R-project.org/package=survminer) (32) in R. Univariate Cox regression was
conducted to evaluate the association between LINC01117 expression
and OS using the ‘survival’ package (version 3.7.0). Multivariate
Cox regression was further performed to determine whether LINC01117
was an independent prognostic factor, adjusting for age, sex, T
stage, N stage and TNM stage. Hazard ratios and 95% confidence
intervals were calculated.
Correlation analysis
Raw RNA-seq count data from TCGA-LUAD cohort were
normalized for library size and composition bias using the trimmed
mean of M-values method in the edgeR package. Based on the
edgeR-normalized RNA-seq expression matrix of patients with LUAD,
the correlations between LINC01117 and 17,109 PCGs from TCGA-LUAD
cohort were evaluated in R (version 4.4.2). Gene expression values
were log2-transformed [log2(x + 1)] prior to correlation analysis.
The ‘limma’ package (version 3.62.2; http://bioconductor.org/packages/limma) (33) was used for preprocessing of the
expression matrix, and Pearson correlation coefficients and
corresponding P-values were calculated using the cor.test function
from the stats package (version 4.4.2), which is included in the
base R distribution. Genes with r>0.4 and P<0.05 were
considered significantly correlated, and scatter plots with fitted
regression lines were generated for visualization.
Correlation analysis using gene
expression profiling interactive analysis (GEPIA)
To further investigate the correlation between
LINC01117 and HOXD8, the GEPIA online database (http://gepia.cancer-pku.cn/) was used. The TCGA-LUAD
tumor cohort was selected, and the correlation between the two
genes was assessed based on GEPIA-provided log2TPM expression
values. Pearson's correlation coefficient was calculated and the
corresponding scatter plot was downloaded.
Tissue collection
Fresh-frozen paired tumor and adjacent non-tumor
tissues (21 LUAD and 9 LUSC cases) were used to verify the
expression of LINC01117. The median age of patients with LUAD was
60 years (range, 40–76 years), including 11 women and 10 men. The
median age of patients with LUSC was 61 years (range, 45–73 years),
including 3 women and 6 men. These tissue specimens were collected
from The Second Affiliated Hospital of Xi'an Jiaotong University
(Xi'an, China) between May 10, 2016 and July 25, 2019. All patients
were newly diagnosed with tumors, and had not undergone
radiotherapy or chemotherapy prior to sample collection. The
patient tissue samples used in the current study were originally
collected with approval from the Ethics Committee of The Second
Affiliated Hospital of Xi'an Jiaotong University (approval no.
2016036). The present study and all experimental procedures
involving these samples were separately approved by the same
committee (approval no. 2022185). The present study adhered to The
Declaration of Helsinki and all specimens were obtained only after
securing written informed consent from each patient prior to tissue
collection.
Cell line culture and
transfection
In total, four LUAD cell lines (A549, H1299, PC-9
and H1975) and one normal bronchial epithelial cell line (BEAS-2B)
were used in the present study. All cell lines were obtained from
The Cell Bank of Type Culture Collection of The Chinese Academy of
Sciences and were cultured in RPMI 1640 medium (Gibco; Thermo
Fisher Scientific, Inc.) supplemented with 10% fetal bovine serum
(Biological Industries; Sartorius AG) at 37°C in a 5%
CO2 humidified atmosphere. The full-length LINC01117
sequence was cloned into the pEX-3 (pGCMV/MCS/Neo) vector to
generate the LINC01117 overexpression plasmid; the corresponding
empty pEX-3 (pGCMV/MCS/Neo) vector was used as the negative control
(NC) for overexpression experiments. Vectors were purchased from
Shanghai GenePharma Co., Ltd. For knockdown experiments,
gene-specific small interfering RNAs (siRNAs) were designed and a
non-targeting scrambled siRNA was used as the NC (siNC). All
plasmids and siRNAs were synthesized by Shanghai GenePharma Co.,
Ltd. PC-9 cells at 70–80% confluence were transfected with the
LINC01117 overexpression plasmid or the corresponding NC plasmid,
whereas A549, PC-9 and NCI-H1299 cells at 60–70% confluence were
transfected with siRNAs or the corresponding siNC. Transient
transfection was performed using Lipofectamine® 2000
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions, and all procedures were carried
out at room temperature. siRNAs were transfected at a final
concentration of 50 nM, and plasmids were transfected at 2 µg/well.
Cells were transfected with siRNAs or plasmids for 6 h in a
humidified atmosphere at 37°C with 5% CO2, after which
the transfection medium was replaced with fresh complete medium.
Total RNA was isolated 24 h post-transfection to confirm
overexpression efficiency or evaluate knockdown efficiency. For
subsequent experiments, transfected cells were reseeded at 24 h
post-transfection. Cells were harvested 48 h post-transfection for
protein extraction, and total RNA was also isolated at 48 h
post-transfection to assess the expression of other genes.
Following transfection, the cells were further incubated in a
humidified atmosphere at 37°C with 5% CO2. The siRNA
sequences targeting the relevant genes are listed in Table II.
| Table II.siRNA sequences for target genes. |
Table II.
siRNA sequences for target genes.
| siRNA | Sequence,
5′-3′ |
|---|
|
LINC01117-siRNA1 |
GCGAUACGUGAUCAUUUAATT |
|
LINC01117-siRNA2 |
GGUUAAUUGCUUGCCCAAATT |
|
LINC01117-siRNA3 |
GCAAACGAAGAGAUGGUAATT |
| HOXD8-siRNA1 |
GCCUGACAAAUUAACUUCUTT |
| HOXD8-siRNA2 |
GGUUCCAGAACAGGAGAAUTT |
| HOXD8-siRNA3 |
GUCGCUUCCAAACUCUAGATT |
| siNC |
UUCUCCGAACGUGUCACGUTT |
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Cytoplasmic and nuclear RNA was isolated from A549,
NCI-H1299, PC-9 and H1975 cells using the Cytoplasmic & Nuclear
RNA Purification Kit (Norgen Biotek Corp.). Total tissue RNA and
cellular RNA were extracted using the FAST1000 and FAST200 kits
(Shaanxi Xianfeng Biotechnology Co., Ltd.), respectively. Total RNA
was reverse transcribed into cDNA using the PrimeScript™ RT Reagent
Kit (Takara Bio, Inc.) under the following conditions: 37°C for 15
min and 85°C for 5 sec, followed by maintenance at 4°C.
Quantitative amplification was then performed with TB
Green® Premix Ex Taq™ II (Takara Bio, Inc.) under the
following cycling conditions: 95°C for 30 sec for initial
denaturation, followed by 40 cycles at 95°C for 5 sec and 60°C for
30 sec. For nucleo-cytoplasmic fractionation, U6 was used as a
nuclear fractionation control to verify enrichment of the nuclear
RNA fraction, whereas GAPDH was used as a cytoplasmic fractionation
control (34,35). The target gene expression levels
were quantified via the 2−ΔΔCq method (36). All primers were synthesized by
Sangon Biotech Co., Ltd., the sequences of which are listed in
Table III.
| Table III.Primer sequences. |
Table III.
Primer sequences.
| Gene | Sequence,
5′-3′ |
|---|
| GAPDH | F:
CTCCTCCACCTTTGACGCTG |
|
| R:
TCCTCTTGTGCTCTTGCTGG |
| LINC01117 | F:
ACCTCCTGACCCTGAAAGCA |
|
| R:
TGTTGGGGTCTGTCCCATCT |
| U6 | F:
CTCGCTTCGGCAGCACA |
|
| R:
AACGCTTCACGAATTTGCGT |
| HOXD8 | F:
AGCAGCTCCTGGTAGACGAA |
|
| GAG |
|
| R:
AGGGCTAGGGCGTGGGAAAC |
Colony formation assays
Following transfection, cells were seeded into
6-well plates (200 cells/well) and incubated in a humidified
incubator with 5% CO2 at 37°C for 7 days. Subsequently,
at room temperature, colonies were fixed with 4% formaldehyde for
20 min, stained with 0.5% crystal violet for 20 min and images were
captured. Colonies containing >50 cells were manually counted
for analysis.
Live cell imaging to evaluate cell
proliferation
Transfected cells were seeded into 24-well culture
plates at a density of 2×104 cells/well. After adhesion,
the plates were placed into the live cell imaging system
(Cytation5; BioTek; Agilent Technologies, Inc.) for real-time
monitoring, with images captured every 2 h. Cell proliferation was
then evaluated by quantifying the cells; the cell proliferation
rate (%) was calculated as the relative increase in cell number per
unit area compared that at 0 h using the formula:
[(Nt/A)-(N0/A)]/(N0/A) ×100, where Nt is the cell number
at the indicated time point, N0 is the cell number at 0
h, and A is the imaging area.
Cell Counting Kit-8 (CCK-8) assay
Transfected cells were seeded into 96-well plates at
a density of 2×103 cells/well. After 24, 48 and 72 h of
incubation, 10 µl CCK-8 reagent (Shanghai Qihai Futai Biotechnology
Co., Ltd.) was added to the medium and the plates were incubated
for 40 min. The absorbance at 450 nm was then measured using a
microplate reader (BioTek; Agilent Technologies, Inc.).
5-ethynyl-2′-deoxyuridine (EdU)
assay
The EdU kit was purchased from Guangzhou RiboBio
Co., Ltd. Post-transfection, 8×103 cells/well were
seeded into 96-well plates and incubated at 37°C for 48 h in a cell
incubator. The medium was then replaced with 50 µM EdU-containing
medium for an additional 2-h incubation. At room temperature,
following fixation with 4% paraformaldehyde for 30 min, the cells
were stained with Apollo solution for 30 min on a shaker in the
dark, and nuclear DNA was counterstained with Hoechst reagent for
30 min on a shaker in the dark, according to the manufacturer's
protocol. Fluorescence images were captured using a Nikon Ti-S
inverted fluorescence microscope (Nikon Corporation). The
EdU-positive rate (%) was determined as: (Apollo-stained
cells/Hoechst-stained nuclei) ×100.
Western blotting
Proteins were extracted from NCI-H1299 and PC-9
cells using RIPA lysis buffer (Beyotime Biotechnology). Protein
concentration was determined using a BCA Protein Assay Kit (cat.
no. P0012S; Beyotime Biotechnology). Equal amounts of protein (20
µg/lane) were separated by 10% Tris-glycine/SDS-PAGE. Following
electrophoretic transfer onto PVDF membranes, the membranes were
blocked with 5% non-fat milk at 37°C for 1 h. The membranes were
then incubated overnight at 4°C with primary antibodies against
HOXD8 (cat. no. sc-515357; dilution 1:200; Santa Cruz
Biotechnology, Inc.) and GAPDH (cat. no. 2118; dilution 1:1,000;
Cell Signaling Technology, Inc.), with the latter serving as the
internal control. The membranes probed for GAPDH and HOXD8 were
incubated with HRP-conjugated goat anti-rabbit IgG secondary
antibody (cat. no. 7074; dilution, 1:1,000; Cell Signaling
Technology, Inc.) and HRP-conjugated goat anti-mouse IgG secondary
antibody (cat. no. 7076; dilution, 1:1,000; Cell Signaling
Technology, Inc.), respectively, at 37°C for 1 h. The signals were
visualized using an HRP chemiluminescent detection kit (cat. no.
WBKLS0050; MilliporeSigma) and captured with an imaging system
(model no. 5200; Tanon Science and Technology Co., Ltd.). Band
intensities were semi-quantified by densitometric analysis using
ImageJ software (version 1.54g; National Institutes of Health).
Actinomycin D (ActD) treatment
ActD was purchased from TargetMol (cat. no. TN9827)
and dissolved in dimethyl sulfoxide. In PC-9 cells, ActD was added
to the LINC01117 overexpression and NC groups to achieve a final
concentration of 10 µg/ml, and the cells were incubated at 37°C for
0, 2, 4, 6 and 8 h after treatment. Cells were then harvested at
room temperature for total RNA extraction. HOXD8 mRNA levels at
each time point were quantified by RT-qPCR as aforementioned and
normalized to the 0-h time point to assess degradation
kinetics.
Statistical analysis
All statistical analyses were performed using
GraphPad Prism 9.0 (Dotmatics). The normality of data distribution
was assessed using the Shapiro-Wilk test. For normally distributed
data, comparisons between two groups were performed using the
Student's t-test, with paired t-tests used for tissue samples and
unpaired t-tests used for other two-group comparisons, whereas the
Mann-Whitney U test was used for non-normally distributed data. For
comparisons involving multiple groups, one-way ANOVA was applied,
followed by Tukey's post hoc test. Data following a normal
distribution are presented as the mean ± standard deviation,
whereas non-normally distributed data are presented as the median
(interquartile range). Pearson correlation analysis was used to
assess the correlation between gene expression levels. For tissue
samples, each sample represented an independent biological
replicate. RNA expression levels were measured with at least three
technical replicates per sample, and the mean value was used for
statistical analysis. All other in vitro experiments were
performed with at least three independent biological replicates.
For bioinformatics analyses based on TCGA data, the sample size was
determined by all eligible patients meeting the inclusion criteria
in the public database. Due to the retrospective nature of the
present study, no a priori power calculation was performed.
Two-sided P<0.05 was considered to indicate a statistically
significant difference.
Results
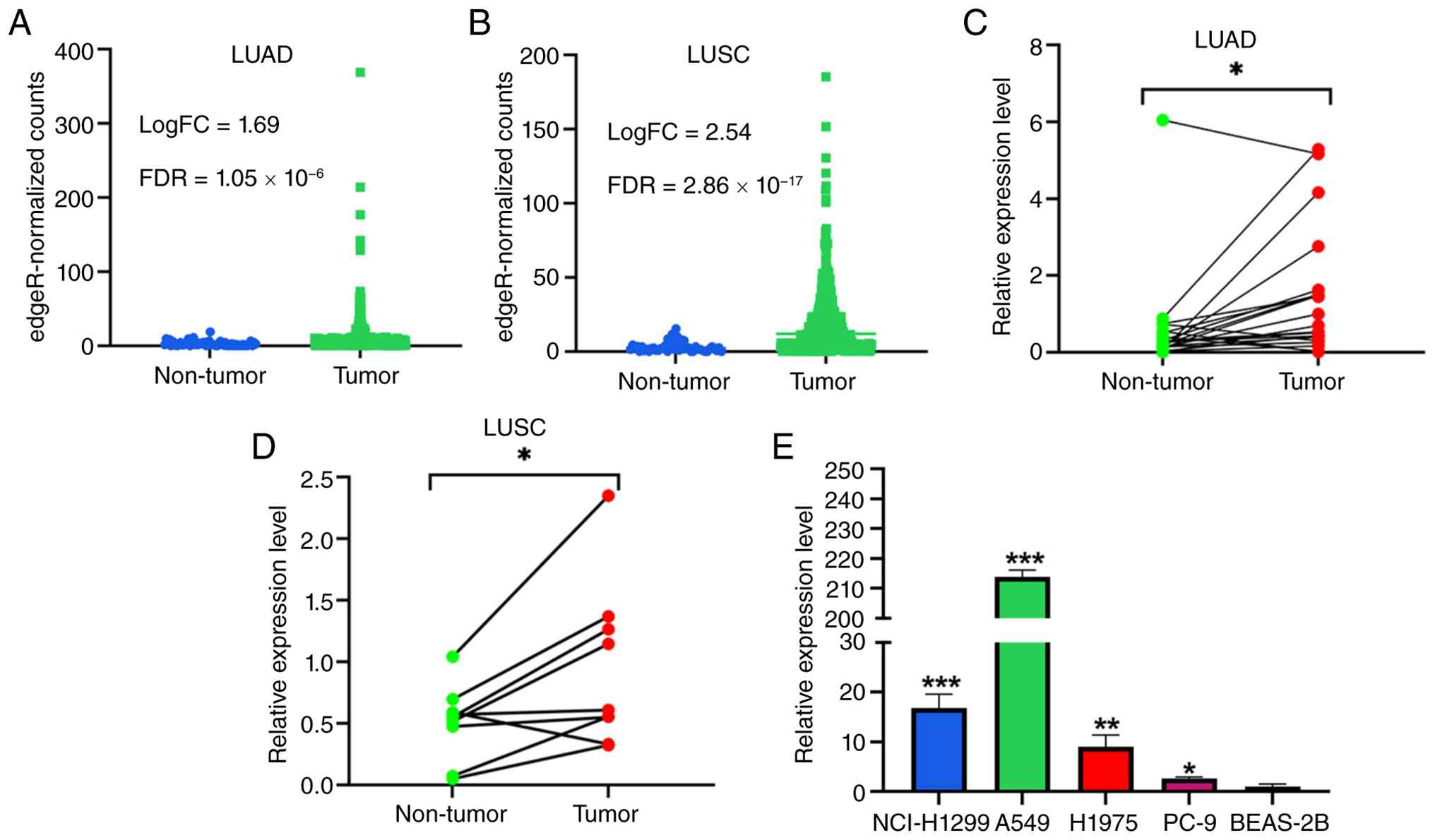
LINC01117 is upregulated in LUAD and
LUSC
Differential expression analysis of TCGA data
revealed that LINC01117 was upregulated in LUAD (Fig. 1A) and LUSC (Fig. 1B) tumor tissues compared with in
non-tumor tissues. Consistent with this, analysis of the collected
clinical tissue samples showed that LINC01117 was significantly
upregulated in the tumor tissues from 21 LUAD (Fig. 1C) and nine LUSC (Fig. 1D) samples relative to the matched
adjacent non-tumor tissues. Furthermore, LINC01117 expression was
elevated in LUAD cell lines compared with in the normal bronchial
epithelial BEAS-2B cell line (Fig.
1E). Based on these findings, the A549 and H1299 cell lines
were selected for subsequent LINC01117 knockdown experiments,
whereas the PC-9 cell line was used for overexpression
experiments.
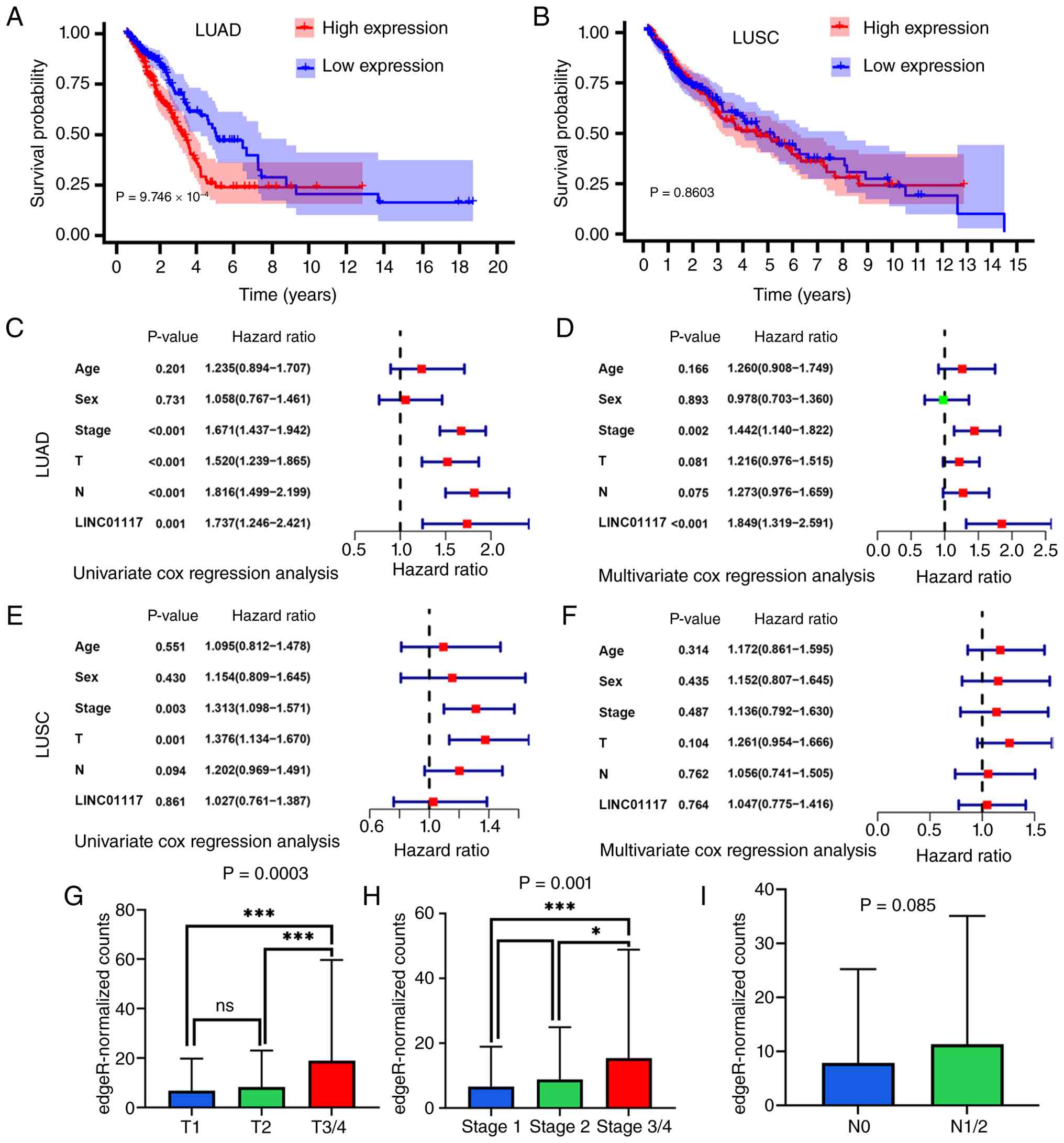
LINC01117 is positively associated
with poor prognosis in patients with LUAD
KM analysis of TCGA data revealed that elevated
LINC01117 expression was associated with poorer outcomes in LUAD
(Fig. 2A) but not in LUSC
(Fig. 2B). Among the patients with
LUAD, the median survival time was significantly shorter in the
high-expression group (3.14 years) than the low-expression group
(4.93 years). The 3- and 5-year survival rates were 50.5 and 24.0%
in the high-expression group vs. 68.3 and 47.0% in the
low-expression group, respectively. Univariate Cox regression
analysis confirmed a significant association between LINC01117 and
OS (Fig. 2C). Multivariate Cox
regression analysis further demonstrated that LINC01117 remained
independently associated with LUAD prognosis after adjusting for
clinical characteristics (Fig.
2D). These results suggested that LINC01117 may be an
independent prognostic biomarker for LUAD. By contrast, neither the
univariate nor multivariate Cox regression analyses showed a
significant prognostic association between LINC01117 and LUSC
(Fig. 2E and F). Therefore, the
present study focused primarily on investigating the functional
mechanisms of LINC01117 in LUAD.
To further evaluate the prognostic value of
LINC01117 in LUAD, its association with clinical stage was
analyzed. One-way ANOVA revealed a significant difference in
expression among the T1, T2 and T3/4 groups; subsequent Tukey's
multiple-comparisons test showed that expression levels in both the
T1 and T2 groups were significantly lower than those in the T3/4
group, whereas no significant difference was observed between the
T1 and T2 groups (Fig. 2G).
One-way ANOVA revealed a significant difference in expression among
the stage 1, stage 2 and stage 3/4 groups; subsequent Tukey's
multiple-comparisons test showed that expression levels in both the
stage 1 and stage 2 groups were significantly lower than those in
the stage 3/4 group, whereas no significant difference was observed
between the stage 1 and stage 2 groups (Fig. 2H). Although no significant
differences were observed across N stages, the LINC01117 expression
levels showed an increasing trend with advancing N stage (Fig. 2I). These findings collectively
suggested that elevated LINC01117 expression may predict poor
prognosis in patients with LUAD.
Knockdown of LINC01117 inhibits LUAD
cell proliferation
To further understand the role of LINC01117 in LUAD,
cytoplasmic and nuclear RNA were extracted from A549, NCI-H1299,
PC-9 and H1975cells, and RT-qPCR analysis of the fractionated RNA
showed enrichment of U6 in the nuclear fraction and GAPDH in the
cytoplasmic fraction, supporting successful nucleo-cytoplasmic
separation. LINC01117 expression was then detected using RT-qPCR,
and the results indicated that LINC01117 was mainly localized in
the cytoplasm (Fig. 3A).
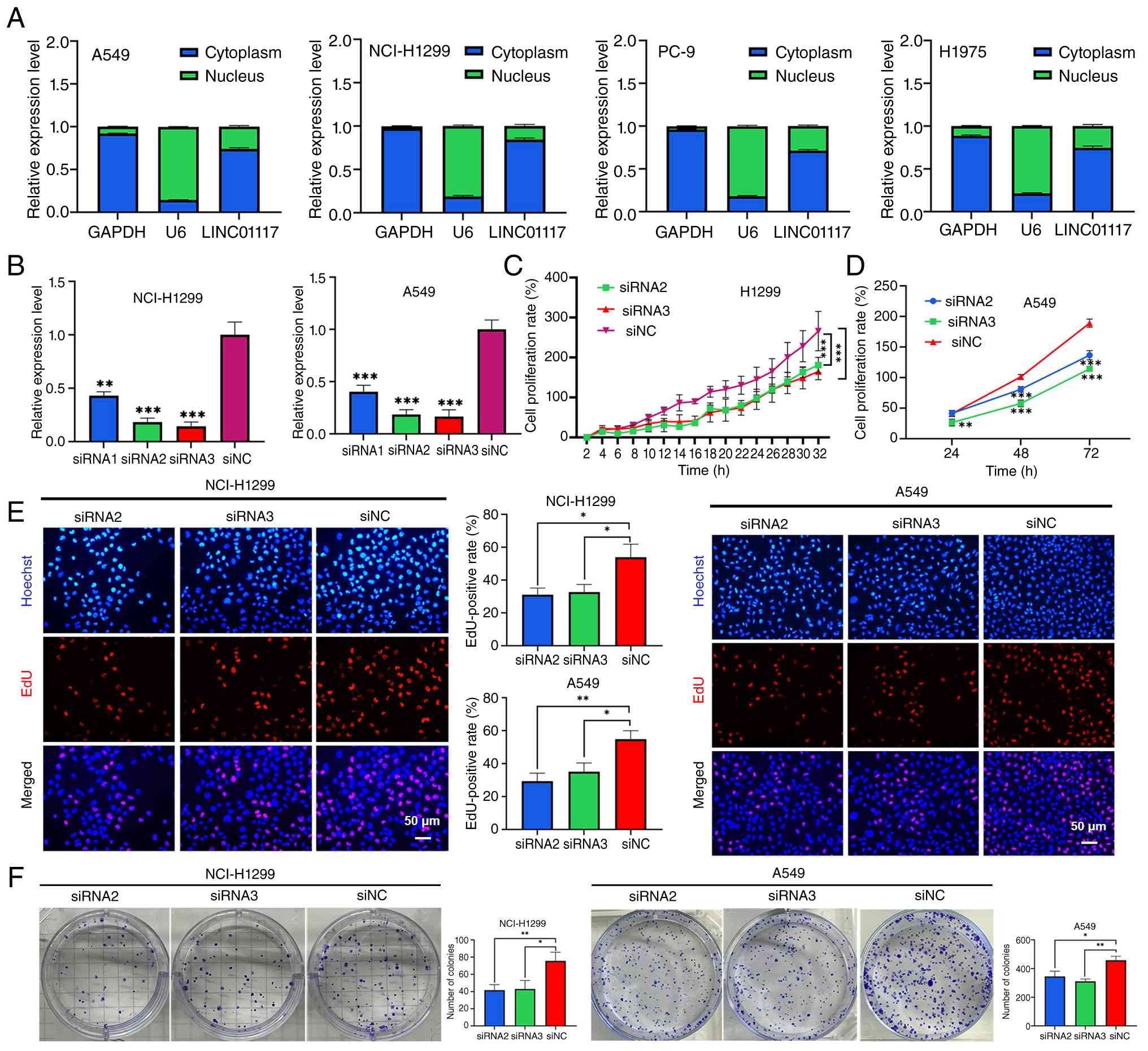 | Figure 3.Knockdown of LINC01117 inhibits LUAD
cell proliferation. (A) Subcellular distribution of LINC01117 in
LUAD cells. (B) Knockdown efficiency of LINC01117 using siRNAs. (C)
In NCI-H1299 cells, live-cell imaging assay measured cell
proliferation following LINC01117 knockdown. (D) In A549 cells,
Cell Counting Kit-8 assay measured cell proliferation following
LINC01117 knockdown. In NCI-H1299 and A549 cells, (E) EdU and (F)
colony formation assays assessed cell proliferation following
LINC01117 knockdown. *P<0.05, **P<0.01, ***P<0.001 vs.
siNC or as indicated. EdU, 5-ethynyl-2′-deoxyuridine; LINC01117,
long intergenic non-protein coding RNA 1117; LUAD, lung
adenocarcinoma; NC, negative control; si, small interfering. |
Subsequently, A549 and NCI-H1299cells were
transfected with siRNAs targeting LINC01117 to achieve knockdown of
expression. RT-qPCR analysis revealed that siRNA2 and siRNA3
exhibited higher knockdown efficiencies compared with siRNA1
(Fig. 3B) and were therefore
selected for further experiments. Live cell imaging demonstrated
significantly lower NCI-H1299 cell survival rates in the siRNA2 and
siRNA3 groups compared with those in the siNC group (Fig. 3C). To improve the reliability of
the findings, the CCK-8 assay was used to assess cell proliferation
in the A549 cell line, and the results showed that LINC01117
knockdown inhibited cell proliferation compared with in the siNC
group (Fig. 3D). In NCI-H1299 and
A549 cells, the EdU assay revealed markedly lower EdU-positive
rates in the LINC01117 knockdown groups than in the NC group
(Fig. 3E). Similarly, the colony
formation assay showed a significant reduction in colony numbers
following LINC01117 knockdown compared with the NC in NCI-H1299 and
A549 cells (Fig. 3F).
Collectively, these results demonstrated that LINC01117 knockdown
inhibited LUAD cell proliferation.
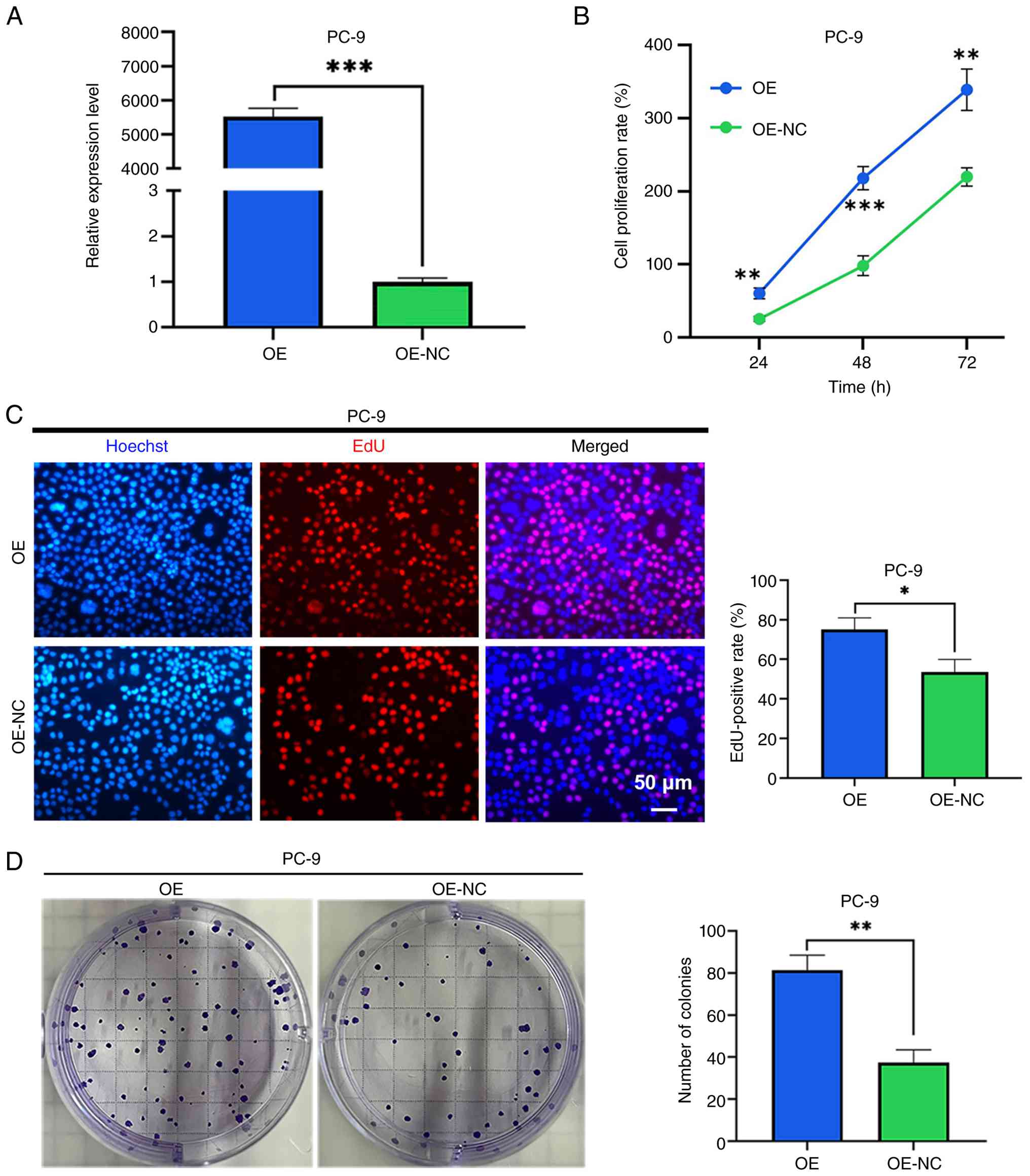
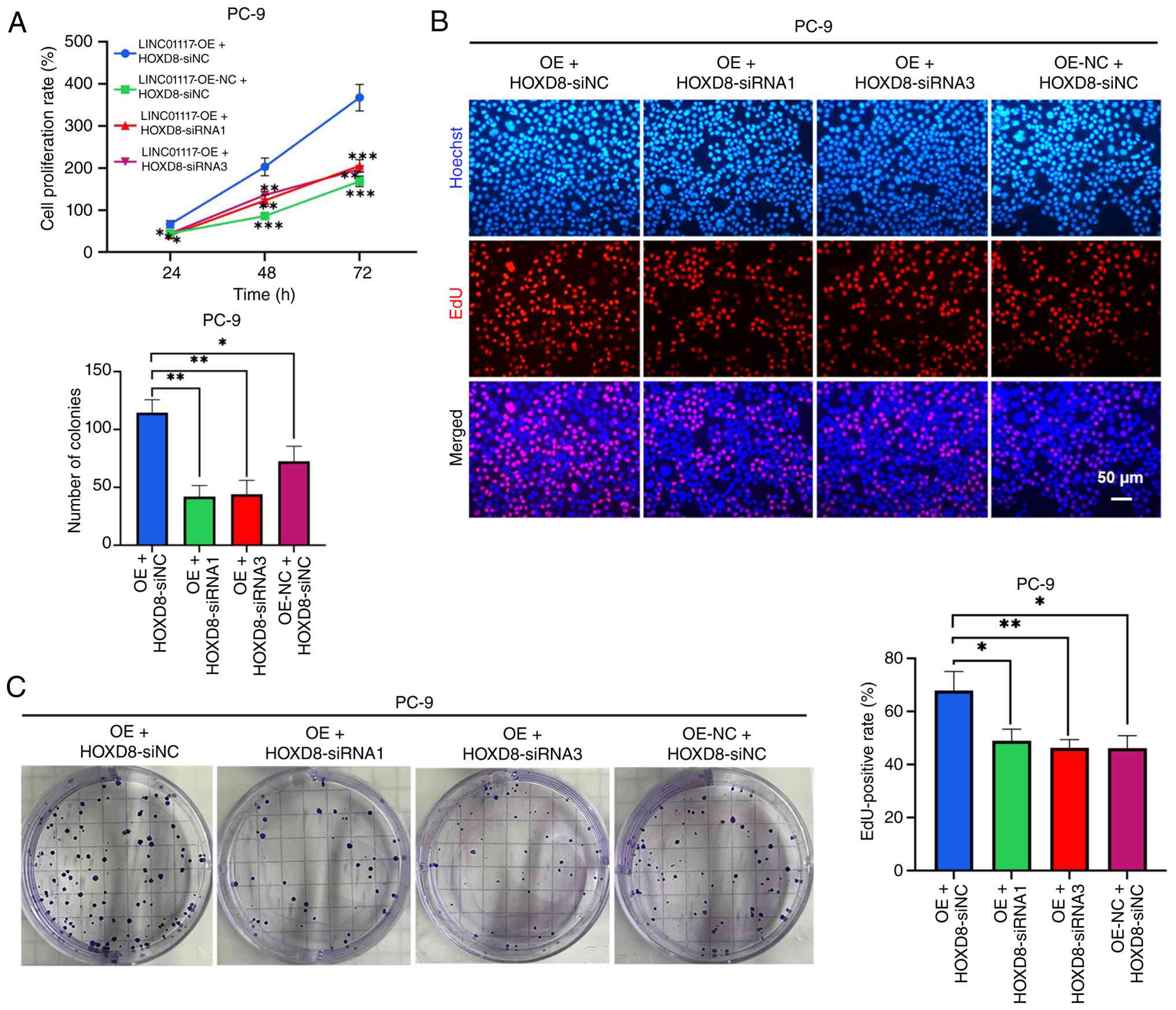
Overexpression of LINC01117 promotes
LUAD cell proliferation
LINC01117 was successfully overexpressed in PC-9
cells using a plasmid vector (Fig.
4A). The CCK-8 assay results demonstrated that the LINC01117
overexpression group exhibited a significantly higher cell
proliferation rate compared with that in the control group
(Fig. 4B). Similarly, the EdU
assay revealed a markedly higher EdU-positive rate in the LINC01117
overexpression group than in the NC group (Fig. 4C). Furthermore, the colony
formation assay showed a significantly increased number of colonies
in the LINC01117 overexpression group relative to the control group
(Fig. 4D). Collectively, these
results indicated that LINC01117 overexpression promoted LUAD cell
proliferation.
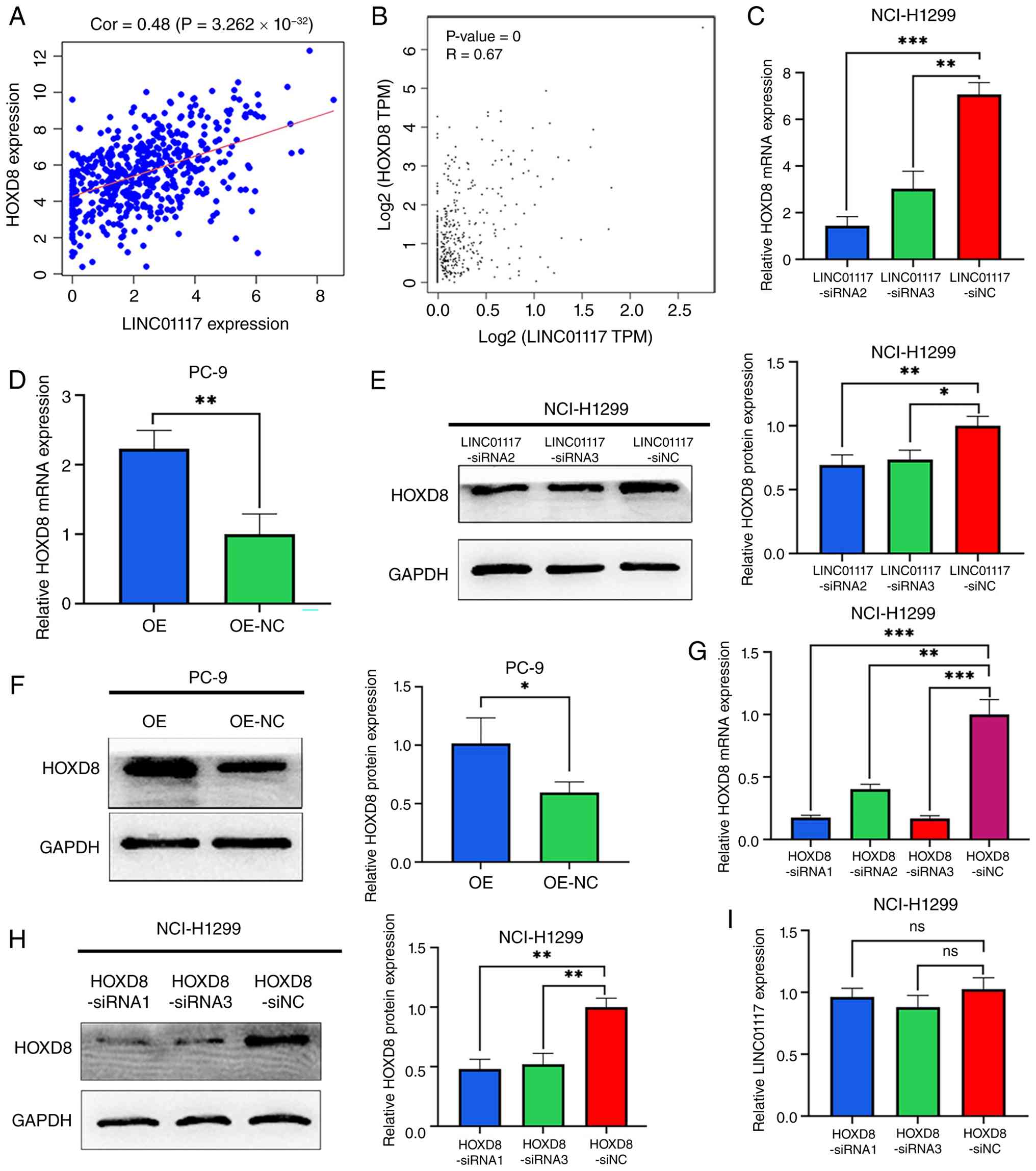
HOXD8 is a downstream target gene of
LINC01117
To investigate the mechanisms by which LINC01117 may
promote LUAD progression, Pearson correlation analyses between
LINC01117 expression and the expression levels of all PCGs from
TCGA-LUAD cohort were performed. The results revealed that
LINC01117 was most significantly co-expressed with HOXD8 (Fig. 5A). This co-expression pattern was
validated using data from the GEPIA database (Fig. 5B).
The regulatory relationship between LINC01117 and
HOXD8 was then examined. Knockdown of LINC01117 significantly
reduced the mRNA and protein levels of HOXD8 (Fig. 5C and E), whereas LINC01117
overexpression increased the mRNA and protein levels of HOXD8
(Fig. 5D and F). To determine
whether HOXD8 reciprocally regulated LINC01117, NCI-H1299 cells
were transfected with siRNAs to knockdown HOXD8 expression. RT-qPCR
analysis revealed that, among the three HOXD8 siRNAs, siRNA1 and
siRNA3 exhibited higher knockdown efficiencies than siRNA2 and were
therefore selected for further experiments (Fig. 5G). The knockdown efficiencies were
also successfully validated at the protein level (Fig. 5H). Notably, HOXD8 knockdown did not
alter LINC01117 expression (Fig.
5I). These results demonstrated that HOXD8 may be a downstream
target gene of LINC01117.
LINC01117 promotes LUAD cell
proliferation by targeting HOXD8
Having established that LINC01117 promotes LUAD cell
proliferation and that HOXD8 may be a downstream target gene of
LINC01117, the present study aimed to determine whether HOXD8
itself is involved in NCI-H1299 cell proliferation. CCK-8 assays
showed that knockdown of HOXD8 significantly suppressed the cell
proliferation rate (Fig. 6A). EdU
assays showed that the EdU-positive rate in the HOXD8 knockdown
groups was significantly lower than that in the NC group (Fig. 6B). Additionally, colony formation
assays showed that the number of colonies in the HOXD8 knockdown
groups was significantly lower than that in the NC group (Fig. 6C). These results indicated that
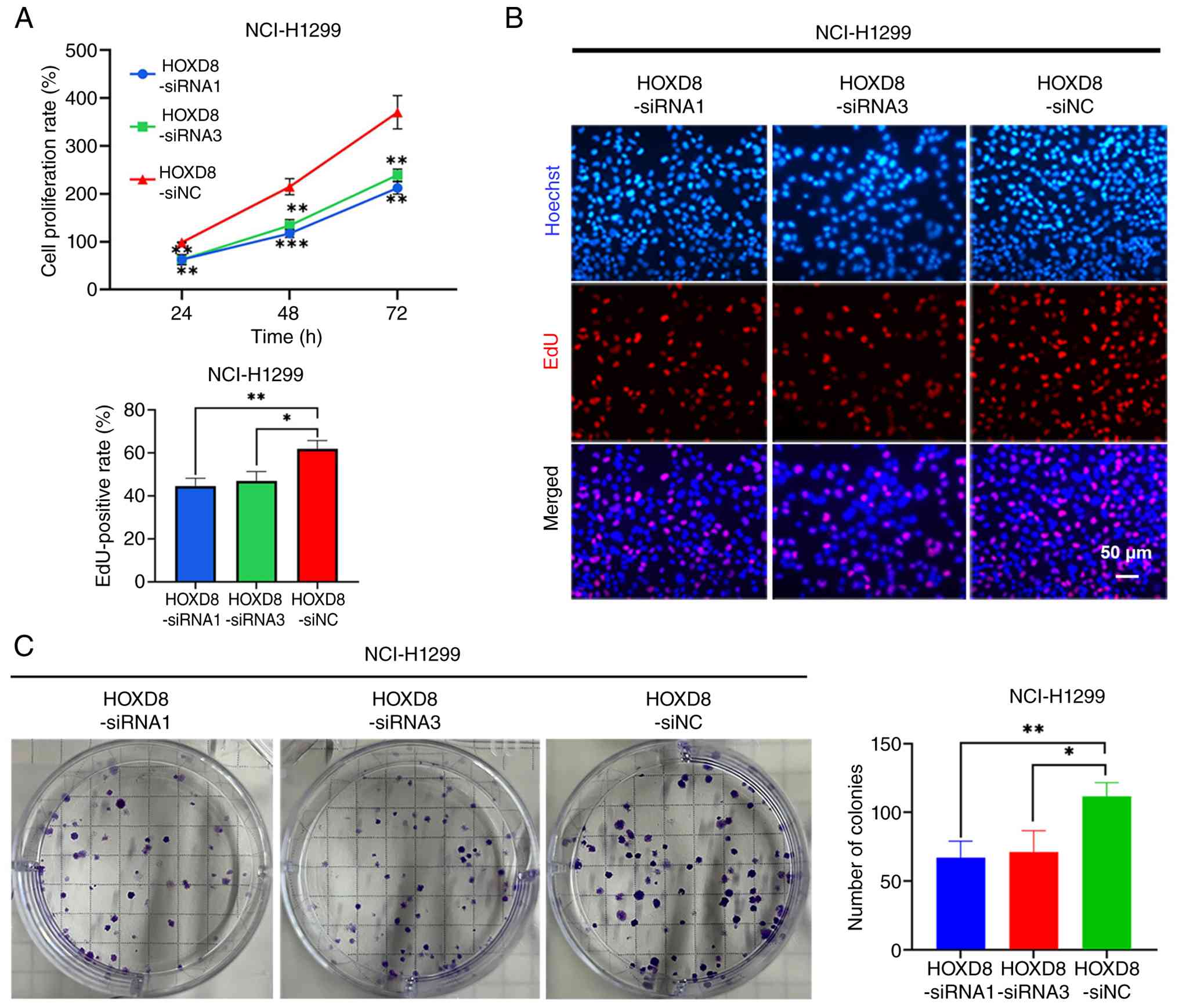
HOXD8 promoted LUAD cell proliferation.
It was thus considered that LINC01117 may promote
LUAD cell proliferation by targeting HOXD8. To test this, NCI-H1299
cells were transfected with a LINC01117 overexpression plasmid,
followed by transfection with HOXD8 siRNAs to reduce HOXD8
expression, and cell proliferation was subsequently examined. The
results of the CCK-8 assay showed that knockdown of HOXD8 in
LINC01117-overexpressing cells significantly inhibited cell
proliferation compared with the LINC01117-overexpression +
HOXD8-siNC group (Fig. 7A). In
addition, HOXD8 knockdown significantly reduced the EdU-positive
rate in LINC01117-overexpressing groups compared with in the
LINC01117-overexpression + HOXD8-siNC group (Fig. 7B). Furthermore, the colony
formation assay demonstrated that the number of colonies was
significantly reduced in HOXD8-knockdown LINC01117-overexpressing
groups compared with that in the LINC01117-overexpression +
HOXD8-siNC group (Fig. 7C). Thus,
HOXD8 knockdown reversed the pro-proliferative effect of LINC01117
overexpression. Collectively, these results indicated that
LINC01117 may promote cell proliferation through HOXD8.
LINC01117 is associated with increased
HOXD8 mRNA stability
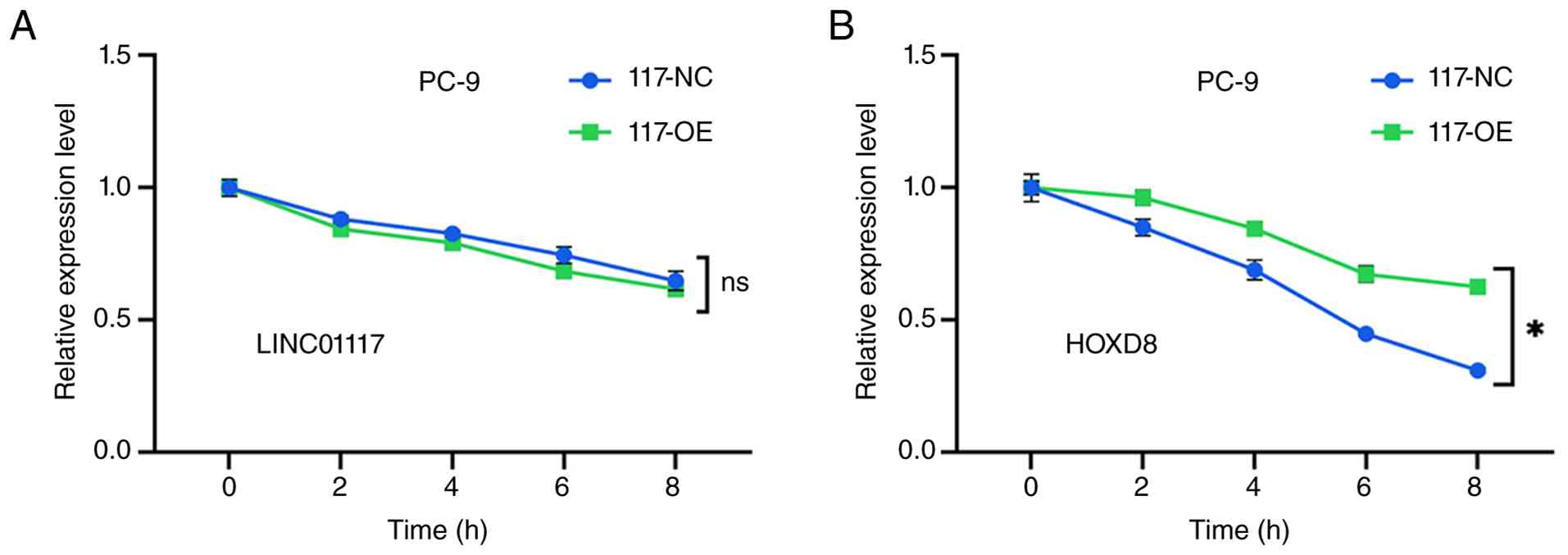
As aforementioned, changes in LINC01117 expression
lead to corresponding changes in HOXD8 expression at both the mRNA
and protein levels, suggesting that LINC01117 may stabilize HOXD8
mRNA. To assess this hypothesis, ActD was used to inhibit de
novo RNA transcription. In PC-9 cells, the degradation rate of
LINC01117 in the LINC01117 overexpression group did not differ
significantly from that in the NC group (Fig. 8A). However, the degradation rate of
HOXD8 mRNA was significantly slower in the LINC01117 overexpression
group compared with the control (Fig.
8B). These results indicated that LINC01117 was associated with
increased HOXD8 mRNA stability.
Discussion
Lung cancer accounts for the highest number of
cancer-related deaths (1);
therefore, an improved understanding of the molecular mechanisms
underlying LUAD is crucial for improving patient survival rates.
Notably, a number of studies have focused on identifying prognostic
lncRNA biomarkers, some of which have demonstrated clinical value
(37,38). LINC01117 is a relatively new
lncRNA, the functions and mechanisms of which in lung cancer remain
largely unclear. The present study demonstrated that LINC01117 was
upregulated in LUSC tumor tissues, and in LUAD tumor tissues and
cell lines. Furthermore, high LINC01117 expression was
significantly associated with a poor prognosis in patients with
LUAD, but showed no significant association with prognosis in
patients with LUSC. Thus, the present study focused on elucidating
the functions and mechanisms of LINC01117 in LUAD.
Notably, the survival curves of the high- and
low-expression groups tended to converge after ~10 years in
patients with LUAD; this suggests that the prognostic impact of
LINC01117 may be more pronounced in the early stages of disease
progression. This pattern may be influenced by several factors,
including tumor heterogeneity, subsequent treatment interventions
and dynamic changes in the tumor microenvironment, such as
cancer-associated inflammation and immune modulation. Further
studies are warranted to elucidate the underlying mechanisms.
In the present study, bioinformatics analysis
revealed a positive association between LINC01117 expression and
the T and TNM stages of LUAD, whereas no significant association
was observed with N stage. Although an increasing trend in
LINC01117 expression was observed with higher N stage, this
difference was not statistically significant. Therefore, this
finding should be interpreted with caution and requires further
validation in larger cohorts. It remains unclear whether this
result stems from limitations in TCGA sample size or indicates a
true absence of association with metastasis. To elucidate the
function and mechanism of LINC01117 in LUAD, knockdown and
overexpression experiments were performed. Collectively, these
results suggested that LINC01117 promoted LUAD cell
proliferation.
Most ncRNAs regulate biological processes by
interacting with PCGs (39).
Therefore, to identify potential targets of LINC01117, correlation
coefficients between LINC01117 and all PCGs were calculated in the
present study. Among them, HOXD8 exhibited the strongest positive
correlation coefficient with LINC01117. Notably, both LINC01117 and
HOXD8 are located at the 2q31.1 chromosomal locus, with HOXD8
positioned upstream of LINC01117. In the present study, knockdown
of LINC01117 significantly reduced HOXD8 expression, whereas its
overexpression significantly increased HOXD8 expression at both the
mRNA and protein levels. By contrast, knockdown of HOXD8 did not
significantly alter the LINC01117 expression levels. Collectively,
these findings indicated that HOXD8 may be a downstream target of
LINC01117.
A previous study reported that lncRNA LINC01116
post-transcriptionally regulates HOXD8 to promote bladder cancer
progression (40), which aligns
with the lung cancer findings in the present study. HOXD8 is a
transcription factor implicated in various cancer types (27). However, the role of HOXD8 appears
to be context-dependent. In some tumors, HOXD8 acts as an oncogene;
for example, it is upregulated in glioma, and promotes glioma cell
proliferation and migration (41).
By contrast, it may function as a tumor suppressor in certain
cancer types, such as gastric cancer, where its expression is
downregulated and negatively associated with tumor malignancy
(42). Studies have indicated that
HOXD8 contributes to chemoresistance, promotes proliferation and
metastasis, and frequently serves as a target for ncRNAs (24–26,43).
In the present study, HOXD8 knockdown reversed the
LINC01117-induced increase in cell proliferation. These results
suggested that LINC01117 may promote LUAD cell proliferation by
targeting HOXD8. Moreover, the present study demonstrated that
LINC01117 was associated with increased HOXD8 mRNA stability.
The present study has several limitations that
should be acknowledged. First, the expression of LINC01117 was only
assessed in 21 LUAD and nine LUSC tumor tissues and adjacent
non-tumor tissues. In future studies, the sample size should be
increased and additional clinical data should be collected to
further analyze the association between LINC01117 expression and
clinicopathological factors. Second, while the present study
focused on the role of LINC01117 in cell proliferation, its
potential involvement in metastasis was not investigated. Third,
the results suggested that LINC01117 may regulate HOXD8 mRNA
stability, but the precise molecular mechanism underlying this
regulation remains to be fully elucidated. Finally, the mechanism
by which LINC01117 may promote the malignant proliferation of LUAD
was not verified in vivo, and further in vivo
validation is required to confirm the biological role of LINC01117
in LUAD development. Despite these limitations, the current study
provides novel insights into the potential mechanism by which
LINC01117 contributes to LUAD progression. It offers a theoretical
basis for the development of LINC01117 as a potential therapeutic
target in LUAD.
Another limitation of the present study should be
acknowledged. Although U6 was used as a nuclear fractionation
control to verify the quality of nucleo-cytoplasmic separation
(34,35), U6 is a small nuclear RNA rather
than a lncRNA, and therefore may not represent an ideal universal
endogenous reference for lncRNA RT-qPCR normalization. Thus,
although the present data support the predominantly cytoplasmic
localization of LINC01117, this conclusion would be further
strengthened by additional approaches, such as validation with
multiple reference transcripts, absolute quantification or
RNA-fluorescence in situ hybridization analysis in future
studies.
In conclusion, the results of the present study
demonstrated that LINC01117 was upregulated in LUAD and may serve
as an independent prognostic biomarker. Mechanistically, LINC01117
promoted LUAD cell proliferation by increasing HOXD8 mRNA
stability. Overall, the present study identified LINC01117 as an
oncogene that may drive LUAD progression and predict poor patient
outcomes.
Acknowledgments
Not applicable.
Funding
The present study was supported by the Natural Science
Foundation for Young Scientists of Shaanxi Province (grant no.
2023-JC-QN-0823) and the National Natural Science Foundation of
China (grant no. 82403050).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
LiZ collected human tissue samples, validated the
expression of LINC01117 in tissues and cells, performed the
bioinformatics and subcellular localization analyses of LINC01117,
and drafted the manuscript. JS assessed the effects of LINC01117
knockdown on cell proliferation. CZ assessed the effects of
LINC01117 overexpression on cell proliferation. YD assessed HOXD8
expression following LINC01117 knockdown and overexpression, and
evaluated LINC01117 expression after HOXD8 knockdown. YC assessed
the effects of HOXD8 knockdown on cell proliferation, as well as
the effects of HOXD8 knockdown on cell proliferation in the
LINC01117-overexpression group. QZ examined the effect of LINC01117
overexpression on the mRNA stability of HOXD8. LoZ and SY
supervised the project, contributed to the study design and revised
the manuscript. LiZ, LoZ and SY confirm the authenticity of all the
raw data. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by The Ethics
Committees of The Second Affiliated Hospital of Xi'an Jiaotong
University (approval nos. 2016036 and 2022185). All subjects
provided written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Laversanne M, Sung H, Ferlay J,
Siegel RL, Soerjomataram I and Jemal A: Global cancer statistics
2022: GLOBOCAN estimates of incidence and mortality worldwide for
36 cancers in 185 countries. CA Cancer J Clin. 74:229–263.
2024.PubMed/NCBI
|
|
2
|
Bade BC and Dela Cruz CS: Lung cancer
2020: Epidemiology, etiology, and prevention. Clin Chest Med.
41:1–24. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Su PL, Furuya N, Asrar A, Rolfo C, Li Z,
Carbone DP and He K: Recent advances in therapeutic strategies for
non-small cell lung cancer. J Hematol Oncol. 18:352025. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lam S, Bai C, Baldwin DR, Chen Y, Connolly
C, de Koning H, Heuvelmans MA, Hu P, Kazerooni EA, Lancaster HL, et
al: Current and future perspectives on computed tomography
screening for lung cancer: A roadmap from 2023 to 2027 from the
international association for the study of lung cancer. J Thorac
Oncol. 19:36–51. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kolb T, Benckendorff J, Möller P, Barth
TFE and Marienfeld RB: Heterogeneous expression of predictive
biomarkers PD-L1 and TIGIT in non-mucinous lung adenocarcinoma and
corresponding lymph node metastasis: A challenge for clinical
biomarker testing. Neoplasia. 38:1008842023. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hayford CE, Tyson DR, Robbins CJ III,
Frick PL, Quaranta V and Harris LA: An in vitro model of tumor
heterogeneity resolves genetic, epigenetic, and stochastic sources
of cell state variability. PLoS Biol. 19:e30007972021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yuan Y, Tang Y, Fang Z, Wen JAO, Wicha MS
and Luo M: Long non-coding RNAs: Key regulators of tumor
Epithelial/mesenchymal plasticity and cancer stemness. Cells.
14:2272025. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li J and Wang X: Functional roles of
conserved lncRNAs and circRNAs in eukaryotes. Noncoding RNA Res.
9:1271–1279. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Song Z, Lin J, Li Z and Huang C: The
nuclear functions of long noncoding RNAs come into focus. Noncoding
RNA Res. 6:70–79. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ferrer J and Dimitrova N: Transcription
regulation by long non-coding RNAs: Mechanisms and disease
relevance. Nat Rev Mol Cell Biol. 25:396–415. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xing C, Sun SG, Yue ZQ and Bai F: Role of
lncRNA LUCAT1 in cancer. Biomed Pharmacother. 134:1111582021.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang Y, Wang Q, Xue H, Guo Y, Wei S, Li
F, Gong L, Pan W and Jiang P: Epigenetic regulation of autophagy in
bone metabolism. Function (Oxf). 5:zqae0042024. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu J, Zhang Q, Yang D, Xie F and Wang Z:
The role of long non-coding RNAs in angiogenesis and
anti-angiogenic therapy resistance in cancer. Mol Ther Nucleic
Acids. 28:397–407. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hashemi M, Moosavi MS, Abed HM, Dehghani
M, Aalipour M, Heydari EA, Behroozaghdam M, Entezari M,
Salimimoghadam S, Gunduz ES, et al: Long non-coding RNA (lncRNA)
H19 in human cancer: From proliferation and metastasis to therapy.
Pharmacol Res. 184:1064182022. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu C, Xu K, Liu J, He C, Liu P, Fu Q,
Zhang H and Qin T: LncRNA RP11-620J15.3 promotes HCC cell
proliferation and metastasis by targeting miR-326/GPI to enhance
glycolysis. Biol Direct. 18:152023. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liang Y, Chen B, Xu F, Long L, Ye F, Wang
Y, Luo D, Li Y, Zhao W, Wang L, et al: LncRNA PRBC induces
autophagy to promote breast cancer progression through modulating
PABPC1-mediated mRNA stabilization. Oncogene. 43:1019–1032. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Deng X, Xiong W, Jiang X, Zhang S, Li Z,
Zhou Y, Xiang B, Zhou M, Li X, Li G, et al: LncRNA LINC00472
regulates cell stiffness and inhibits the migration and invasion of
lung adenocarcinoma by binding to YBX1. Cell Death Dis. 11:9452020.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu T, Dong Y, Yang X, Mo L and You Y:
Crosstalk between lncRNAs and Wnt/β-catenin signaling pathways in
lung cancers: From cancer progression to therapeutic response.
Noncoding RNA Res. 9:667–677. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Leng X, Zhang M, Xu Y, Wang J, Ding N, Yu
Y, Sun S, Dai W, Xue X, Li N, et al: Non-coding RNAs as therapeutic
targets in cancer and its clinical application. J Pharm Anal.
14:1009472024. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang H, Shu L, Niu N, Zhao C, Lu S, Li Y,
Wang H, Liu Y, Zou T, Zou J, et al: Novel lncRNAs with diagnostic
or prognostic value screened out from breast cancer via
bioinformatics analyses. PeerJ. 10:e136412022. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chuang TD, Rysling S, Ton N, Baghdasarian
D and Khorram O: Comparative analysis of differentially expressed
long Non-Coding RNA in Pre- and postmenopausal fibroids. Int J Mol
Sci. 26:67982025. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu L, Ren W, Du L, Xu K and Zhou Y:
LINC01117 inhibits invasion and migration of lung adenocarcinoma
through influencing EMT process. PLoS One. 18:e02879262023.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shen C, Mao D, Tang J, Liao Z and Chen S:
Prediction of LncRNA-Protein interactions based on kernel
combinations and graph convolutional networks. IEEE J Biomed Health
Inform. 28:1937–1948. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang Y, Zhang M, Zhao Y, Deng T, Zhou X,
Qian H, Wang M, Zhang C, Huo Z, Mao Z, et al: HOXD8 suppresses
renal cell carcinoma growth by upregulating SHMT1 expression.
Cancer Sci. 114:4583–4595. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wen X, Hou Y, Zhou L and Fang X: LINC00969
inhibits proliferation with metastasis of breast cancer by
regulating phosphorylation of PI3K/AKT and ILP2 expression through
HOXD8. PeerJ. 11:e166792023. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang Y, Yu Y, Su X and Lu Y: HOXD8
inhibits the proliferation and migration of triple-negative breast
cancer cells and induces apoptosis in them through regulation of
AKT/mTOR pathway. Reprod Biol. 21:1005442021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Loi E, Zavattari C, Tommasi A, Moi L,
Canale M, Po A, Sabato C, Vega-Benedetti AF, Ziranu P, Puzzoni M,
et al: HOXD8 hypermethylation as a fully sensitive and specific
biomarker for biliary tract cancer detectable in tissue and bile
samples. Br J Cancer. 126:1783–1794. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu Y, Miao L, Ni R, Zhang H, Li L, Wang
X, Li X and Wang J: microRNA-520a-3p inhibits proliferation and
cancer stem cell phenotype by targeting HOXD8 in non-small cell
lung cancer. Oncol Rep. 36:3529–3535. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
R Core Team._R, . A Language and
Environment for Statistical Computing_. R Foundation for
Statistical Computing; Vienna, Austria: 2024, <. https://www.R-project.org/>.
|
|
30
|
Liu S, Wang Z, Zhu R, Wang F, Cheng Y and
Liu Y: Three differential expression analysis methods for RNA
sequencing: Limma, EdgeR, DESeq2. J Vis Exp. Sep 18–2021.doi:
10.3791/62528.
|
|
31
|
Therneau T: _A Package for Survival
Analysis in R_. R package version 3.7–0. 2024.<. https://CRAN.R-project.org/package=survival>.
|
|
32
|
Kassambara A, Kosinski M and Biecek P:
_survminer: Drawing Survival Curves using ‘ggplot2’_. R package
version 0.5.1. 2025.<. https://CRAN.R-project.org/package=survminer>.
|
|
33
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43:e472015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Didychuk AL, Butcher SE and Brow DA: The
life of U6 small nuclear RNA, from cradle to grave. RNA.
24:437–460. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yoon JH, Abdelmohsen K, Srikantan S, Yang
X, Martindale JL, De S, Huarte M, Zhan M, Becker KG and Gorospe M:
LincRNA-p21 suppresses target mRNA translation. Mol Cell.
47:648–655. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lin ZB, Long P, Zhao Z, Zhang YR, Chu XD,
Zhao XX, Ding H, Huan SW, Pan YL and Pan JH: Long noncoding RNA
KCNQ1OT1 is a prognostic biomarker and mediates CD8+ T cell
exhaustion by regulating CD155 expression in colorectal cancer. Int
J Biol Sci. 17:1757–1768. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen KL, Huang SW, Yao JJ, He SW, Gong S,
Tan XR, Liang YL, Li JY, Huang SY, Li YQ, et al: LncRNA DYNLRB2-AS1
promotes gemcitabine resistance of nasopharyngeal carcinoma by
inhibiting the ubiquitination degradation of DHX9 protein. Drug
Resist Updat. 76:1011112024. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yao ZT, Yang YM, Sun MM, He Y, Liao L,
Chen KS and Li B: New insights into the interplay between long
non-coding RNAs and RNA-binding proteins in cancer. Cancer Commun
(Lond). 42:117–140. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Meng L, Xing Z, Guo Z and Liu Z: LINC01106
post-transcriptionally regulates ELK3 and HOXD8 to promote bladder
cancer progression. Cell Death Dis. 11:10632020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yu K, Meng J, Chen T, Wang Y, Zhao Y,
Huang T and Gao G: HOXD8 drives Glioma progression through
epithelial-mesenchymal transition regulation: Implications for
prognosis and targeted therapy. Exp Cell Res. 446:1144762025.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yao L, Ye PC, Tan W, Luo YJ, Xiang WP, Liu
ZL, Fu ZM, Lu F, Tang LH and Xiao JW: Decreased expression of the
long non-coding RNA HOXD-AS2 promotes gastric cancer progression by
targeting HOXD8 and activating PI3K/Akt signaling pathway. World J
Gastrointest Oncol. 12:1237–1254. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ahmadi M, Bazrgar M, Akhavan S, Fathi M,
Mousavi P and Ghafouri-Fard S: HOXB and HOXD genes contribute to
the carcinogenic processes in glioblastoma: Evidence form a
bioinformatics analysis. Cancer Treat Res Commun.
43:1009232025.PubMed/NCBI
|