Introduction
Colorectal cancer (CRC) remains one of the most
prevalent malignancies worldwide and continues to represent a major
contributor to cancer-related mortality (1). Recent data indicate that, as of 2022,
CRC ranks as the third most common cancer globally and is the
second leading cause of cancer mortality following lung cancer
(2). In Korea, statistics show
that as of 2022, CRC is the second most frequently diagnosed cancer
after thyroid cancer, and by 2023, it has become the third-leading
cause of cancer-related death following lung and liver cancer
(3). A particular concern in Korea
is the notably high incidence of CRC among younger individuals, the
highest rate reported globally (4). Although advances have been achieved
in surgical procedures, chemotherapy protocols, and targeted
therapies, the prognosis for patients with advanced CRC remains
poor, primarily due to therapeutic resistance, the development of
metastases, and adverse treatment-related effects (5,6).
These ongoing obstacles underscore the urgent need for innovative
therapeutic approaches capable of effectively inhibiting tumor
progression with reduced toxicity. Within this framework, natural
compounds, including phytochemicals and dietary constituents, have
emerged as promising candidates for the development of new
therapeutic modalities for CRC.
β-glucans are naturally occurring polysaccharides
that serve as key structural components within the cell walls of
yeast, fungi, and cereal grains. In recent years, extensive
research has demonstrated their wide-ranging physiological
activities, such as modulation of immune responses (7,8),
regulation of metabolic processes (9–11),
and potential anticancer properties (12–14).
These advances support a growing interest in β-glucans as bioactive
agents suitable for use both as functional food ingredients and as
adjunctive therapies. Notably, β-(1→3),(1→6)-D-glucan, derived from
an edible cultured fungus through fermentation with the strain
Aureobasidium pullulans (AP-FBG), has gained attention for
its increased solubility and enhanced bioavailability, attributes
that may improve its therapeutic efficacy (15–19).
Furthermore, emerging data suggest that derivatives of AP-FBG with
low molecular weight (LMW-AP-FBG) exhibit considerable promise for
therapeutic development (17,20–22).
In parallel, low-molar-mass oat β-glucan has been reported to
influence autophagy- and apoptosis-associated processes during the
early stages of chemically induced colorectal carcinogenesis in
animal models (23). Nevertheless,
such in vivo observations provide limited insight into the
direct cellular responses and molecular signaling events elicited
by β-glucans in human colorectal cancer cells.
Programmed cell death pathways, including apoptosis
and autophagy, are essential for maintaining cellular homeostasis
and have a significant impact on cancer development (24,25).
Apoptosis (type I cell death) is characterized by caspase
activation, DNA fragmentation, and notable morphological changes,
while autophagy (type II cell death) involves the lysosomal
degradation of intracellular components (24–26).
Dysregulation of these mechanisms is strongly associated with
tumorigenesis and the development of resistance to therapy, which
underscores their relevance as potential targets for cancer
treatment (24–26). Recent studies suggest that
β-glucans, in addition to their recognized immunomodulatory
effects, might influence programmed cell death pathways,
particularly apoptosis and autophagy, across diverse cancer types
(20,23,27,28).
Nevertheless, the detailed molecular mechanisms underlying these
effects in CRC remain largely unknown. Considering the pivotal
function of apoptosis-autophagy crosstalk in the regulation of
tumor progression and therapeutic response, investigating the
modulatory effects of β-glucans on these pathways in CRC is
critical for advancing their potential therapeutic applications and
designing new intervention strategies. In this study, we sought to
investigate the effects of LMW-AP-FBG on apoptosis and autophagy in
colorectal cancer cell lines. Understanding the influence of this
agent on cell death pathways could support its further development
as a promising therapeutic option for CRC.
Materials and methods
Materials
RPMI-1640 medium and Dulbecco's modified Eagle's
medium (DMEM) were purchased from WELGENE (Gyeongsan, Korea).
Penicillin/streptomycin was obtained from Cytiva (Marlborough, MA,
USA). Fetal bovine serum (FBS) was purchased from Atlas (Fort
Collins, USA). Z-VAD-FMK, a pan-caspase inhibitor, was procured
from Enzo Life Sciences (New York, USA). WestGlow™ PICO PLUS ECL
Chemiluminescent substrate was acquired from BioMax (Guri, Korea).
Pierce™ BCA protein assay Kit was sourced from Pierce (Rockford,
IL, USA). PRO-PREP™ solution was provided by iNtRON Biotechnology
(Seongnam, Korea). Primary antibodies targeting caspase-3 (#9662),
cleaved caspase-3 (#9664), caspase-7 (#9492), cleaved caspase-7
(#9491), caspase-8 (#4790), cleaved caspase-8 (#9496), caspase-9
(#9502), cleaved caspase-9 (#9501), Poly (ADP-ribose) polymerase
(PARP, #9542), Bcl-2 (#3498), Bax (#2772), autophagy-related 3
(Atg3, #3415), Atg5 (#12994), Atg7 (#8558), microtubule-associated
proteins 1A/1B light chain 3 (LC3, #12741), p62 (#5114),
phosphatidylinositol 3-kinase (PI3K, #4249), phospho-PI3K (p-PI3K,
#4228), protein kinase B (AKT, #9272), phospho-AKT (p-AKT, #9271),
mammalian target of rapamycin (mTOR, #2972), and phospho-mTOR
(p-mTOR, #2971) were obtained from Cell Signaling Technology
(Danvers, MA, USA). Antibodies for apoptosis-inducing factor (AIF,
sc-13116), cytochrome C (sc-7159) and beclin-1 (BCEN-1, sc-12427)
were obtained from Santa Cruz Biotechnology (Santa Cruz, CA).
Horseradish peroxidase-conjugated goat anti-rabbit (GTX213110) and
goat anti-mouse IgG (GTX213111) were supplied by Genetex (Irvine,
CA, USA). GAPDH loading control monoclonal antibody as well as
other unspecified reagents were purchased from Sigma (St. Louis,
MO, USA).
Preparation of LMW-AP-FBG
LMW-AP-FBG, kindly provided by Professor Hisashi
Kimoto (Fukui Prefectural University, Fukui, Japan), was dissolved
in RPMI-1640 medium at a concentration of 10 mg/ml and stored at
4°C until use. To briefly outline the manufacturing process of
LMW-AP-FBG, AP-FBG was suspended in distilled water to reach a
final concentration of 1% (w/v), and sodium hydroxide was then
added to attain an ultimate alkali concentration of 2% (w/v). The
resulting mixture underwent heat treatment in a boiling water bath
for 1 h to achieve alkaline hydrolysis. The alkaline hydrolysate
was neutralized with hydrochloric acid and desalted by dialysis
against ultrapure water using a dialysis membrane. Size-exclusion
high-performance liquid chromatography (SE-HPLC, Tosoh Corporation,
Tokyo, Japan) with a KW 802.5 Shodex column (30 cm × 8 mm i.d.,
Resonac Corporation, Tokyo, Japan) using a phosphate buffer (0.05
M) containing NaCl (0.3 M, pH 7.0) as the mobile phase was employed
to evaluate the degradation of the polymer. Chromatographic
separation was conducted at 30°C with a flow rate of 1.0 ml/min,
and detection was accomplished by a refractive index detector.
According to SE-HPLC analysis, the average molecular weight of
native AP-FBG was 183,683 Da, which decreased to 2,675 Da after
alkaline hydrolysis.
Cell Culture
Human colorectal cancer cell lines (SW480, HCT116,
and Caco-2), human pancreatic cancer cell lines (AsPC-1, PANC-1,
and MIA-PaCa-2), a human lung cancer cell line (A549), an ovarian
cancer cell line (SKOV3), a mouse colon cancer cell line (CT26),
and a human normal colon fibroblast cell line (CCD-18Co) were
obtained from the Korea Cell Bank (Seoul, Korea). All experiments
were performed using low-passage cells (fewer than 10 passages) in
accordance with the supplier's guidelines. These cell lines were
maintained in either RPMI-1640 or DMEM, each supplemented with 10%
heat-inactivated FBS and 1% penicillin-streptomycin solution at
37°C in a 5% CO2 atmosphere.
Cell viability assay
Cell viability was evaluated using the
CellTiter-Glo® 2.0 Cell Viability Assay (Promega,
Wisconsin, USA). In brief, SW480 cells were plated
(5×103 cells) in 96-well plates. Following exposure to
different concentrations of LMW-AP-FBG (prepared in complete
medium) for 24, 48, or 72 h, the assay reagents were added and the
plates were placed on an orbital shaker for 2 min. Luminescence was
measured using the GloMax 96 Microplate Luminometer (Promega,
Wisconsin, USA). The IC50 values were determined using
GraphPad PRISM software 4.0 (GraphPad Software Inc., San Diego, CA,
USA).
In vitro wound healing assay
SW480 cells were seeded (2×104 cells) in
24-well plates and cultured for 24 h to reach confluence. A linear
scratch was created using a sterile 200-µl pipette tip, followed by
gentle washing to remove detached cells. The cells were then
treated with the indicated concentrations of LMW-AP-FBG and
incubated for an additional 24 h. Cells that migrated into the
wound region were visualized using a DM-IL-LED inverted microscope
equipped with an EC3 camera (Leica Microsystems, Heerbrugg,
Switzerland). Wound closure was quantified using ImageJ software
(National Institutes of Health, Bethesda, MD, USA) by calculating
the percentage of wound closure as
[(A0-A24)/A0] ×100, where A0 and
A24 represent the wound area at 0 and 24 h,
respectively. Three random fields per well were analyzed, and
experiments were performed in triplicate.
Measurement of apoptotic cells
The proportion of apoptotic cells was determined
with the Muse™ Annexin V & Dead Cell Kit (Luminex, Austin, TX,
USA), following the manufacturer's protocols. SW480 cells were
cultured in 24-well plates at a density of 5×104
cells/well and treated with increasing doses (0.25, 0.5, and 1
mg/ml) of LMW-AP-FBG for 48 h. Cells were washed with PBS,
collected, and incubated with Annexin V and 7-Aminoactinomycin
D-dead cell marker (7-AAD) for 20 min at room temperature in the
dark. The percentages of cells undergoing early and late apoptosis
were analyzed using the Muse™ Cell Analyzer (Luminex, Austin, TX,
USA).
Western Blot analysis
For whole cell lysate preparation, SW480 cells were
treated with various concentrations of LMW-AP-FBG for 48 h, washed
with ice-cold PBS, and lysed with 50 µl of PRO-PREP™ protein
extraction solution containing a freshly prepared mixture of
complete protease inhibitors (Roche, Switzerland). A total of 20 µg
of protein was separated by 12% or 15% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis, and transferred onto a
PVDF membrane (Millipore, Massachusetts, USA). After blocking the
membranes with 5% non-fat milk in 0.1% Tween 20 Tris Buffered
Saline (TBS-T) for 2 h, membranes were incubated overnight at 4°C
in the presence of primary antibodies. The membranes were
subsequently treated for 3 h at 4°C with secondary antibodies.
Chemiluminescent substrate was then applied, and signals were
visualized using the DAVINCH-Chemi CAS-400 SM (Davinch-k, Seoul,
Korea) and quantified by Total Lab analysis software (Nonlinear
Dynamics, Dunham, NC, USA).
Measurement of the Mitochondrial
Membrane Potential
Mitochondrial membrane potential (MMP) levels were
assessed using the JC-1 Mitochondrial Membrane Potential Detection
Kit (Biotium, Fremont, CA, USA) following the manufacturer's
protocol. In brief, SW480 cells cultured in 24-well plates
(0.5×105 cells/well) were exposed to the indicated
concentrations of LMW-AP-FBG for 48 h. Following treatment, the
cells were washed, collected using trypsin-EDTA, and incubated with
JC-1 Dye (Biotium) for 15 min in a 37°C in a CO2
incubator. The proportion of depolarized cells was determined by
flow cytometry (BD Bioscience, Becton Dickinson, USA) and results
were analyzed with FlowJo Software (Tree Star, Inc, Ashland, OR,
USA).
Statistical analysis
All data are presented as mean ± SEM. Statistical
significance was determined using a two-tailed Student's t test or
a one-way ANOVA followed by Bonferroni's post-hoc test in GraphPad
PRISM software 4.0. A value of P<0.05 was considered
statistically significant.
Results
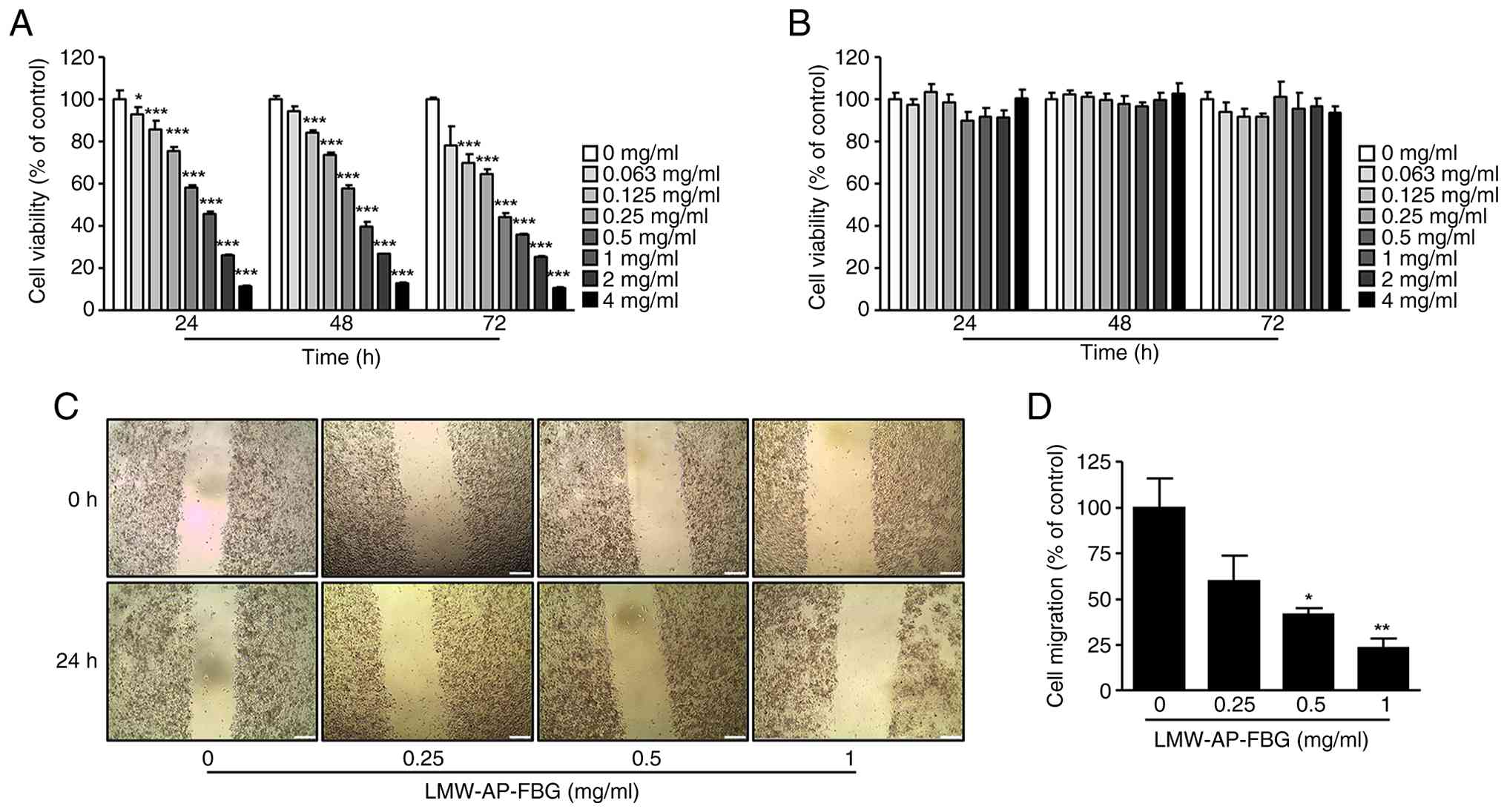
LMW-AP-FBG treatment reduced the cell
viability of SW480 cells
To assess the inhibitory effects of LMW-AP-FBG on
the viability of various cancer cell lines, an initial screening
was performed using the CellTiter-Glo 2.0 Cell Viability Assay.
LMW-AP-FBG treatment at 0.5 mg/ml markedly reduced cell viability
across multiple cancer cell types, including colon, pancreatic,
ovarian, and lung cancer cells, with PANC-1 cells showing the most
pronounced sensitivity (Fig. S1).
These results indicate that LMW-AP-FBG exerts broad
antiproliferative effects against diverse cancer cell lines.
Based on this preliminary screening, SW480 cells
were selected for subsequent mechanistic studies owing to their
well-characterized genetic alterations relevant to colorectal
cancer and their suitability for in vitro anticancer
evaluations. SW480 cells were then treated with increasing
concentrations of LMW-AP-FBG for 24, 48, and 72 h, resulting in a
clear concentration- and time-dependent decrease in cell viability
(Fig. 1A). The calculated
half-maximal inhibitory concentrations (IC50) were 557.2
µg/ml at 24 h, 497.4 µg/ml at 48 h, and 300.6 µg/ml at 72 h.
To evaluate the selectivity of LMW-AP-FBG toward
cancer cells, its cytotoxic effects were examined in normal human
colorectal CCD-18Co cells under identical experimental conditions.
No significant reduction in cell viability was observed at any
tested concentration over 24, 48, and 72 h (Fig. 1B), indicating minimal cytotoxicity
in normal cells.
Given that enhanced cell migration is a key
characteristic of malignant progression, we next assessed the
effect of LMW-AP-FBG on the migratory behavior of SW480 cells using
a wound healing assay. Notably, LMW-AP-FBG significantly suppressed
migratory activity at concentrations of 0.5 and 1 mg/ml (Fig. 1C and D), suggesting a potential
role in suppressing cancer cell motility.
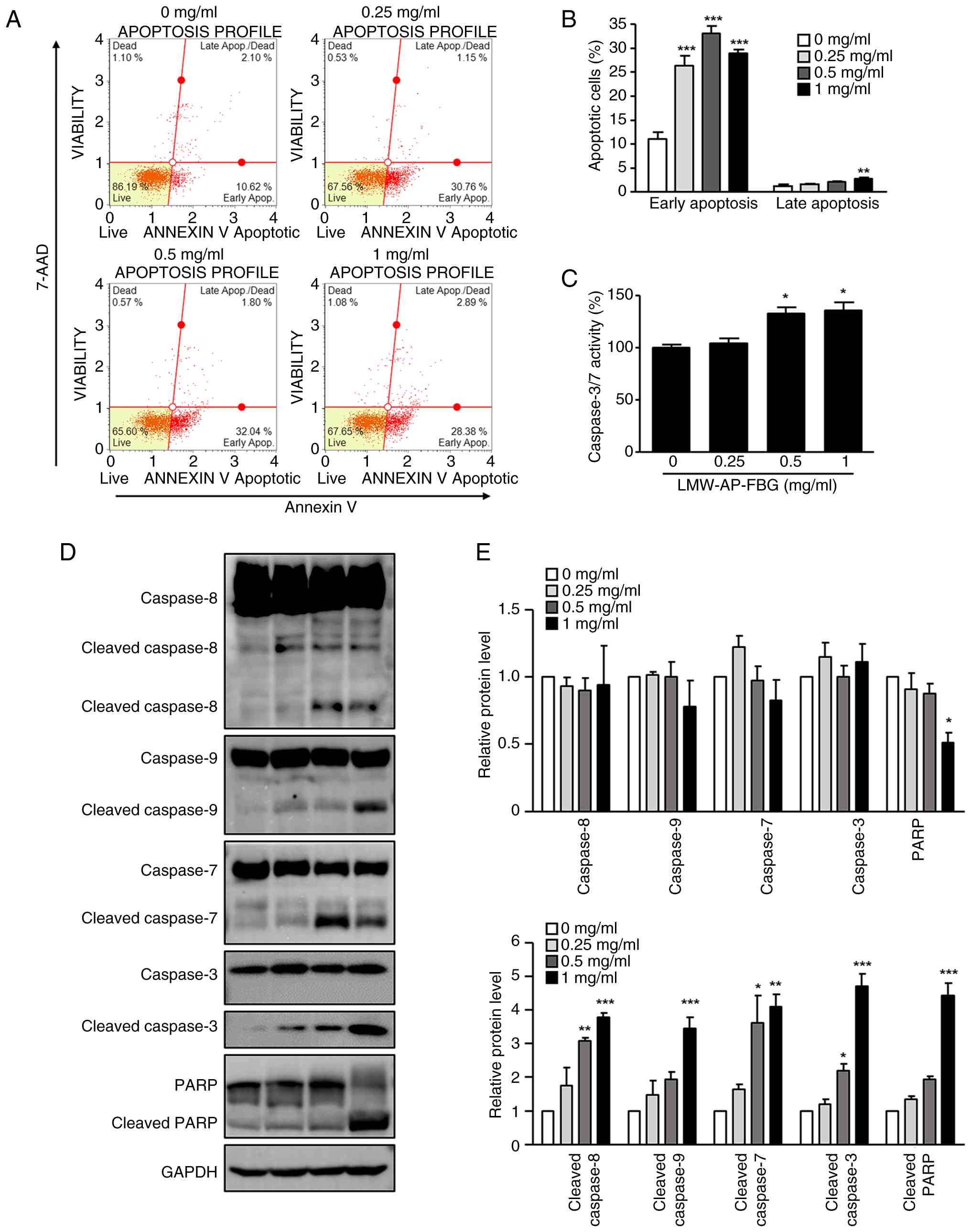
LMW-AP-FBG enhances apoptosis in SW480
cells
Given that LMW-AP-FBG reduced SW480 cell viability,
we subsequently assessed whether LMW-AP-FBG triggers apoptosis
using the Muse Annexin V assay kit. Early and late apoptotic
populations were quantified following 48 h of LMW-AP-FBG exposure.
LMW-AP-FBG led to an approximate 2- to 3-fold increase in early
apoptotic cells compared to controls. Specifically, the proportions
of early apoptotic cells at concentrations of 0.25, 0.5, and 1
mg/ml were 28.4±4.2%, 33.0±3.1%, and 29.0±1.6%, respectively, while
the control was 11.0±2.9% (Fig. 2A and
B). These data suggest that LMW-AP-FBG promotes apoptosis in
SW480 cells.
Furthermore, to investigate the involvement of
caspases in LMW-AP-FBG-induced apoptosis in SW480 cells, caspase-3
and −7 activities were quantitatively assessed using the Caspase
Glo 3/7 luminescence assay. The activity levels of caspase-3 and −7
were markedly increased at both 0.5 and 1 mg/ml concentrations of
LMW-AP-FBG, as demonstrated in Fig.
2C. Given that caspases function as crucial proteases in
apoptosis by cleaving numerous substrate proteins (23,29),
we next examined caspase activation in SW480 cells following
LMW-AP-FBG treatment. Western blot analysis showed that the total
protein levels of caspase-3, −7, −8, and −9 remained largely
unchanged across the tested concentrations (Fig. 2D and E). In contrast, their cleaved
forms were markedly elevated in a concentration-dependent manner,
as confirmed by densitometric analysis (Fig. 2D and E). In parallel, PARP cleavage
was prominently induced, as reflected by the accumulation of
cleaved PARP alongside a concomitant decrease in full-length PARP.
Quantitative analysis further substantiated these findings,
demonstrating significant upregulation of cleaved caspases and PARP
relative to the control group. Collectively, these findings
indicate that LMW-AP-FBG triggers apoptosis by activating caspases
involved in both extrinsic and intrinsic apoptotic processes.
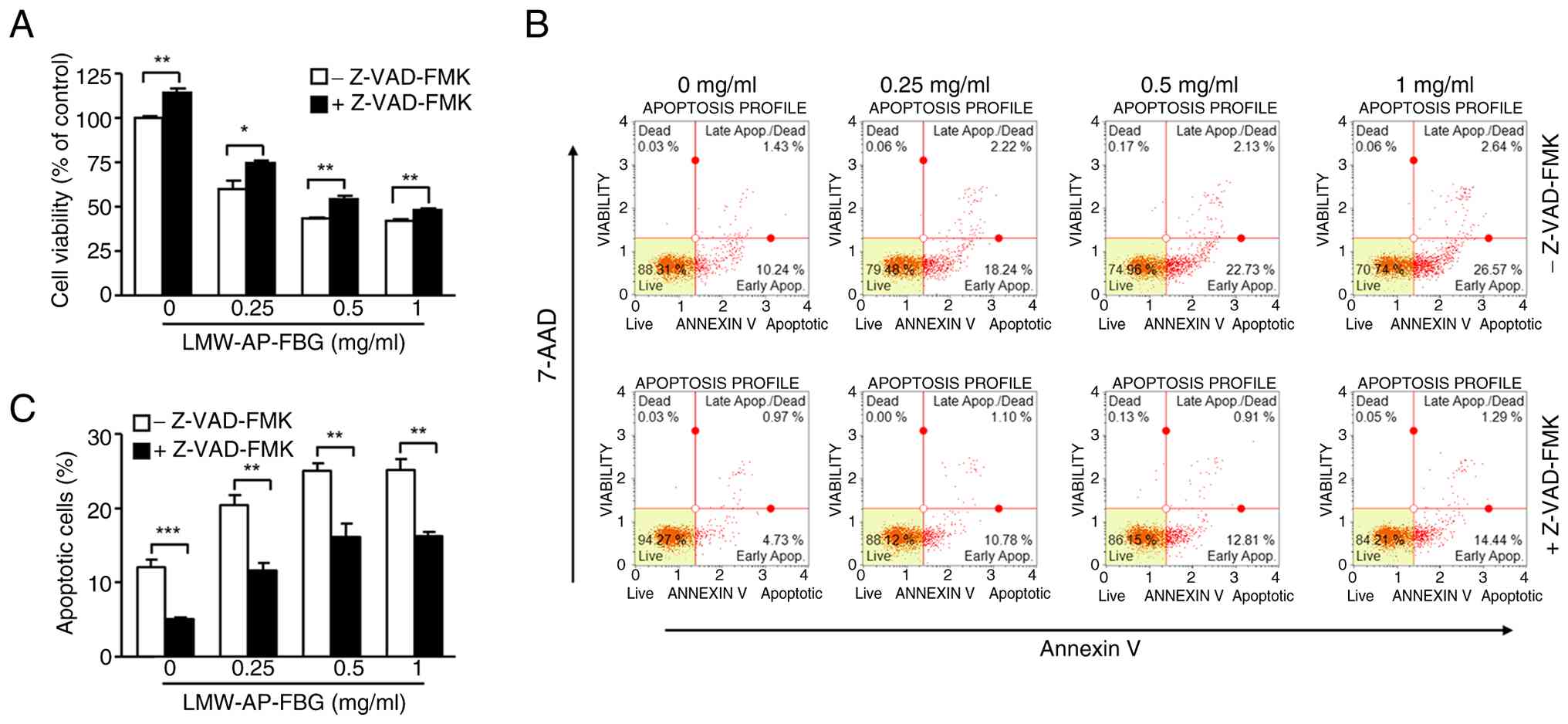
Apoptosis induced by LMW-AP-FBG in
SW480 cells is partially caspase-dependent
To further clarify whether LMW-AP-FBG-induced cell
death in SW480 cells is caspase-dependent, experiments employing
the pan-caspase inhibitor Z-VAD-FMK were conducted. Specifically,
SW480 cells were co-incubated with Z-VAD-FMK (20 µM) and various
concentrations of LMW-AP-FBG for 48 h, followed by assessment of
cell viability using the CellTiter-Glo 2.0 Cell Viability Assay. As
illustrated in Fig. 3A, treatment
with 0.25 mg/ml LMW-AP-FBG alone resulted in 59.8% cell viability,
while co-treatment with Z-VAD-FMK restored viability to 74.3%.
Furthermore, at 1 mg/ml LMW-AP-FBG, the presence of Z-VAD-FMK led
to an approximately 13% increase in viability compared to
LMW-AP-FBG alone (Fig. 3A).
Under equivalent conditions, apoptosis measurements
revealed that the proportion of total apoptotic cells was reduced
in all Z-VAD-FMK co-treated groups in comparison to those receiving
only LMW-AP-FBG (Fig. 3B and C).
More precisely, Z-VAD-FMK co-treatment reduced LMW-AP-FBG-induced
apoptosis from 20.5±2.6% to 11.6±1.97% at 0.25 mg/ml, and from
25.1±2.1% to 16.2±3.1% at 0.5 mg/ml (Fig. 3C). Collectively, these findings
support the notion that the reduction in SW480 cell viability
induced by LMW-AP-FBG is at least partially mediated through
caspase-dependent apoptotic pathways.
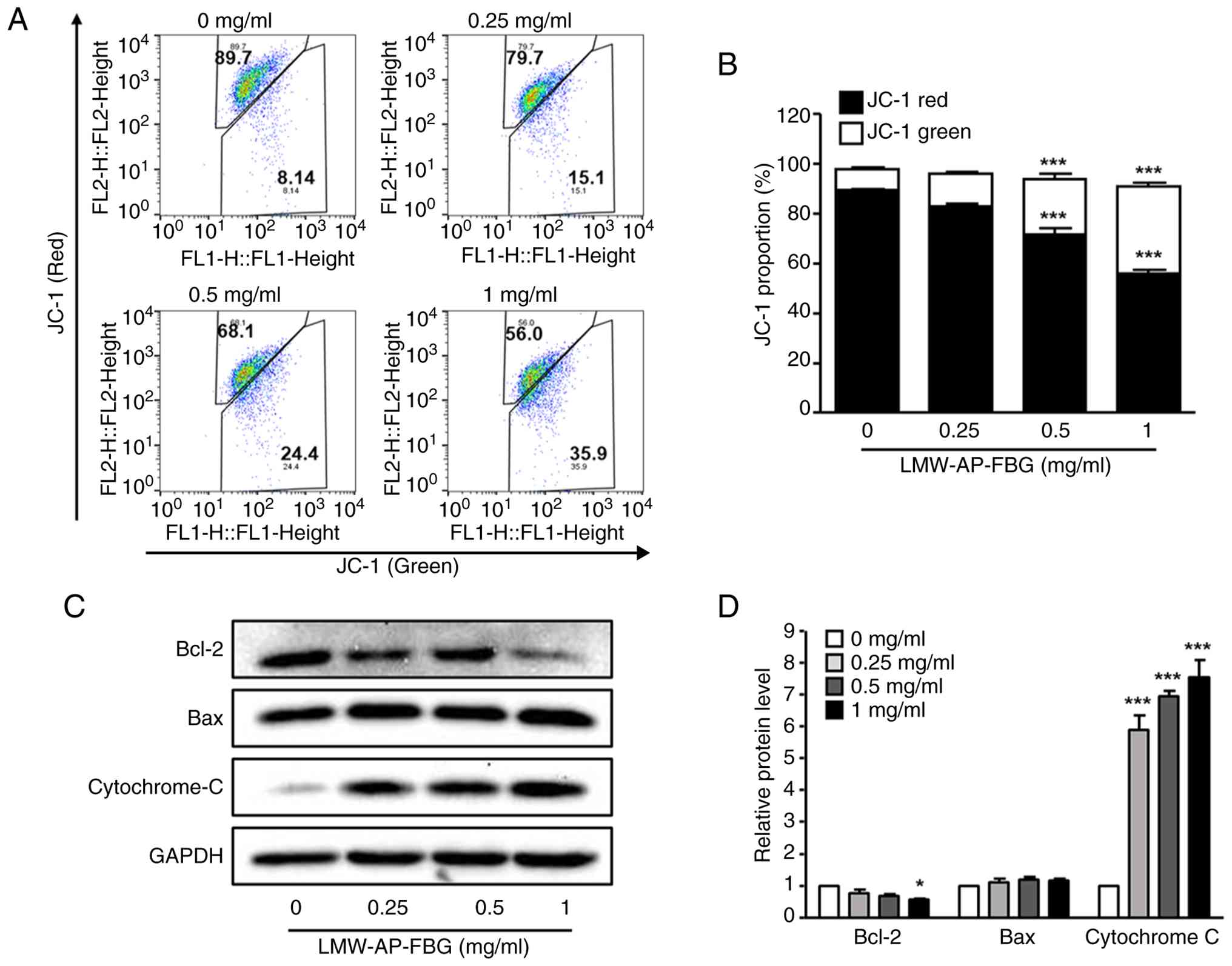
LMW-AP-FBG induces
mitochondria-mediated apoptosis in SW480 cells
Mitochondria play a pivotal role in apoptosis.
During this process, mitochondrial outer membrane permeabilization
(MOMP) markedly increases, facilitating cytochrome c release via
mitochondrial pores, which subsequently activates caspases and
triggers cell death. To evaluate whether LMW-AP-FBG promotes an
increase in MOMP in SW480 cells, we assessed MMP using JC-1
staining. When MMP decreases, JC-1 aggregates (red fluorescence)
dissociate into monomers, resulting in enhanced green fluorescence.
Upon treatment with LMW-AP-FBG concentrations of 0.5 and 1 mg/ml,
the proportion of JC-1 aggregates decreased from 89.58±0.58% of the
control to 71.58±5.64% and 56.05±2.84%, respectively; conversely,
JC-1 monomers increased from 8.49±0.87% of the control to
22.28±4.60% and 34.95±2.77%, respectively (Fig. 4A and B).
In line with the observed decrease in MMP, treatment
of SW480 cells with LMW-AP-FBG resulted in reduced Bcl-2 protein
levels and elevated cytosolic cytochrome c levels. Notably,
cytochrome c increased approximately 6–8-fold compared to the
control group after 48 h of LMW-AP-FBG exposure, while Bcl-2
expression was significantly diminished at 1 mg/ml of LMW-AP-FBG
(Fig. 4C and D). Collectively,
these findings indicate that LMW-AP-FBG induces apoptosis via
mitochondrial pathways regulated by Bcl-2 family proteins.
To further determine whether LMW-AP-FBG induces
AIF-mediated caspase-independent apoptosis, AIF localization was
examined. However, LMW-AP-FBG treatment did not promote AIF
translocation compared with the control group (Fig. S2) and there was no evidence to
suggest that LMW-AP-FBG promoted AIF translocation.
LMW-AP-FBG induces
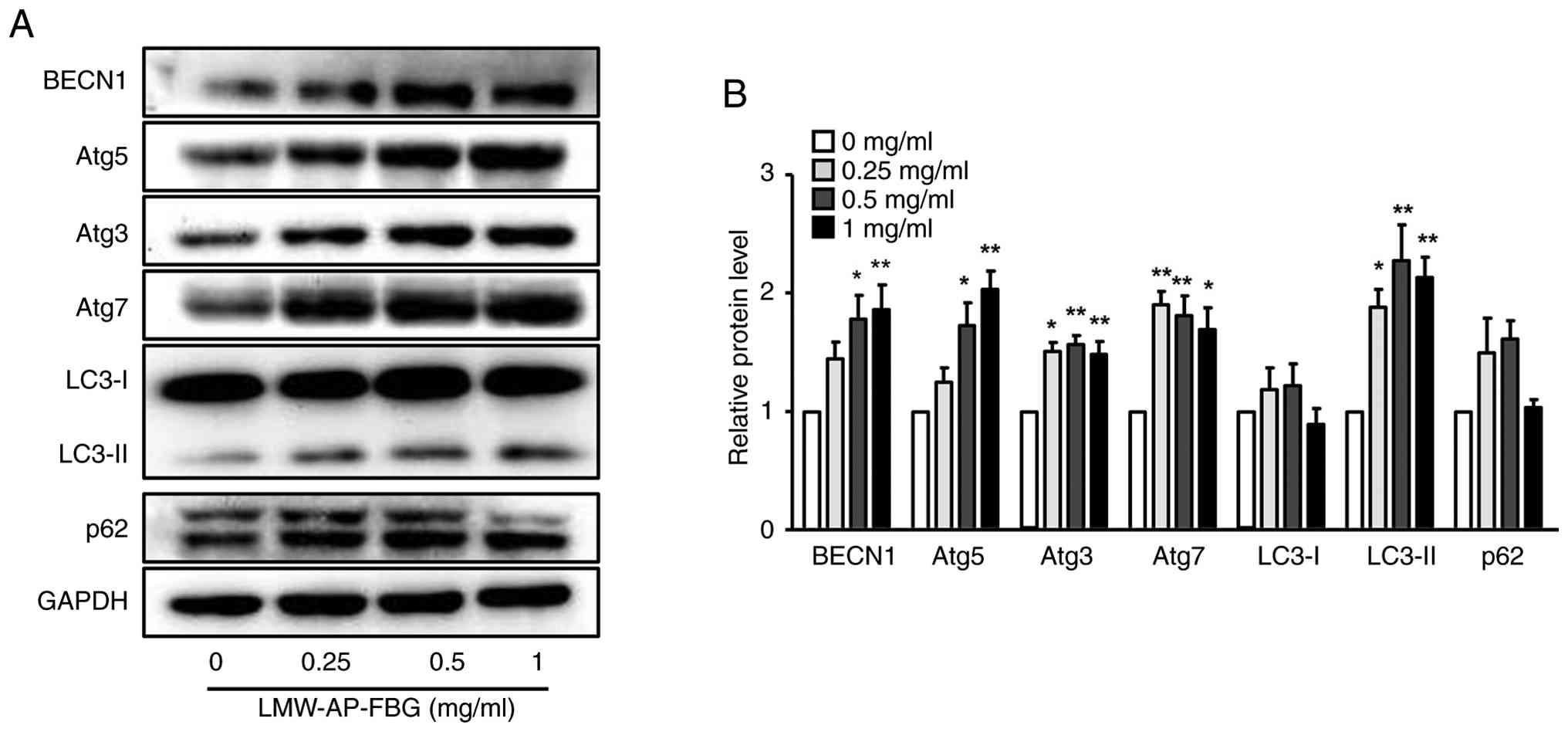
autophagy-associated molecular changes in SW480 cells
Since the extent of apoptosis induced by LMW-AP-FBG
in SW480 cells was less than the observed inhibition of cell
viability, we postulated that LMW-AP-FBG may trigger other forms of
cell death in addition to apoptosis. To investigate this
possibility, we quantified levels of autophagy-associated proteins
using Western blot analysis. At concentrations of 0.5 and 1 mg/ml,
expression levels of BECN1, Atg5, Atg3, and Atg7 were 1.5-fold
greater compared with the control group (Fig. 5A and B). Furthermore, while LC3-I
levels remained unchanged, LC3-II expression was significantly
upregulated in a dose-dependent manner with LMW-AP-FBG treatment
(Fig. 5A and B). For p62, no
significant alteration was observed upon LMW-AP-FBG administration,
suggesting that although autophagy-related machinery was activated,
enhanced autophagic flux was not clearly evident under the
experimental conditions.
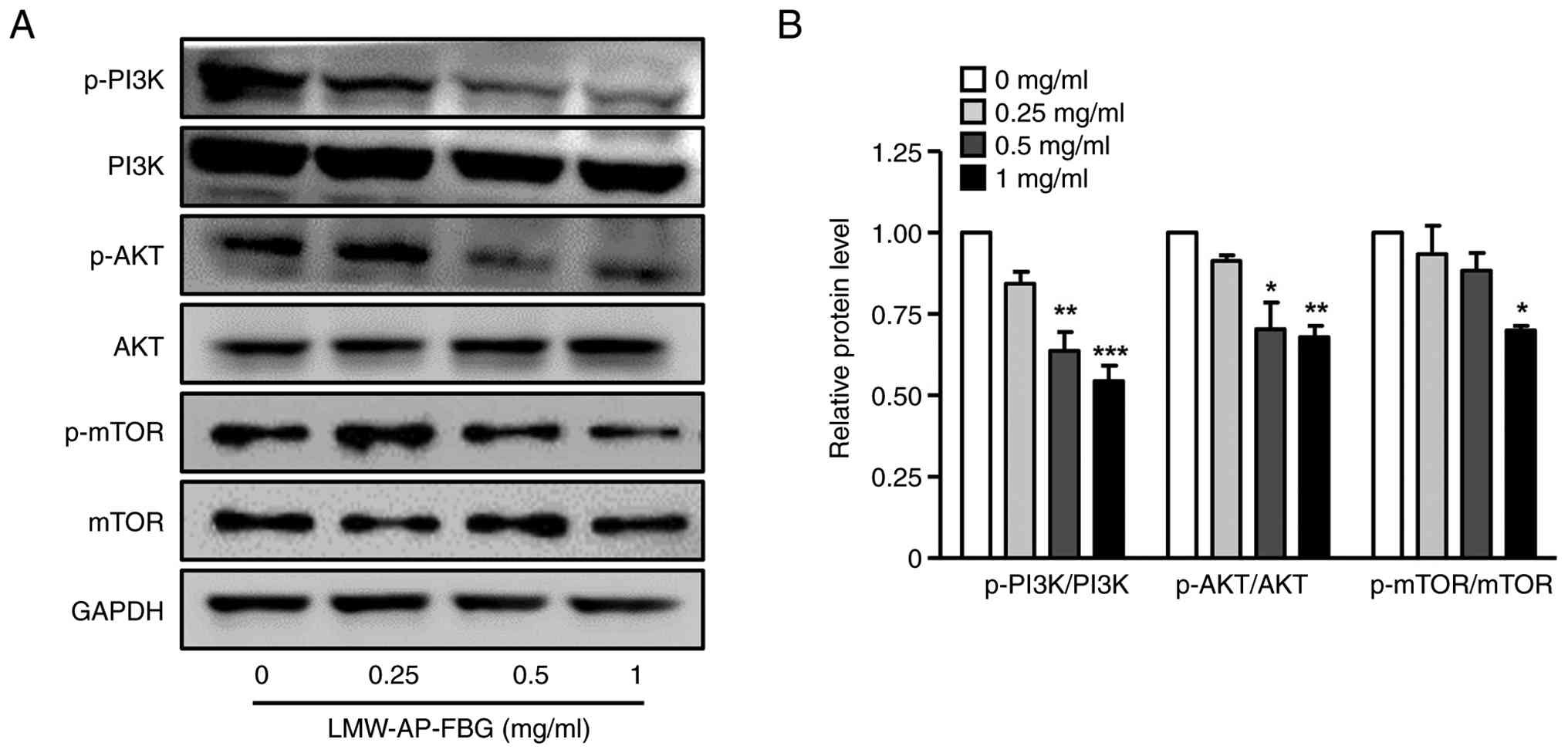
LMW-AP-FBG reduces PI3K/AKT/mTOR
signal transduction pathway
Given that our results demonstrated that LMW-AP-FBG
could induce apoptosis and activate autophagy-related pathways, we
subsequently investigated the PI3K/AKT/mTOR signaling pathway,
which is known to play a central role in modulating the balance
between these two processes (30,31).
As depicted in Fig. 6, treatment
with LMW-AP-FBG resulted in a marked reduction in the
phosphorylation of PI3K, AKT, and mTOR compared to their respective
total protein levels. These outcomes imply that LMW-AP-FBG
substantially suppresses the activation of the PI3K/AKT/mTOR
pathway, thereby inhibiting downstream signaling events that govern
cellular proliferation and survival. Consequently, the inhibition
of this pathway is likely to contribute to the observed induction
of apoptotic and autophagy-related responses in SW480 cells.
Discussion
β-glucans are well recognized for a wide array of
physicochemical and biological activities; however, certain
intrinsic limitations-including challenges in purification,
variability in structural conformations, and inadequate aqueous
solubility-have consistently impeded detailed clarification of
their structure-activity relationships (32,33).
To address these issues, we adopted a sodium hydroxide-mediated
hydrolysis approach aimed at reducing both molecular weight and
viscosity, thereby enhancing both the consistency and
bioavailability of β-glucan samples. In particular, we subjected
crude AP-FBG to hydrolysis and then filtration, obtaining
LMW-AP-FBG with a mean molecular weight below 3,000 Da. In
subsequent functional assays, this LMW-AP-FBG demonstrated
significant growth-inhibitory effects on SW480 CRC cells at
concentrations as low as 0.25 mg/ml. Notably, this active
concentration is approximately one-tenth of concentrations
typically reported in the literature (20,21).
For example, earlier studies in which LMW-AP-FBG was generated
through mechanochemical ball milling instead of chemical hydrolysis
using sodium hydroxide, showed that anticancer effects were seen in
CT-26 cells only at concentrations of 2.5 mg/ml (20). These data underscore the
methodological strengths of the sodium hydroxide-mediated
hydrolysis approach, which enables efficient production of
LMW-AP-FBG with greater bioactivity. This further supports the
importance of processing methods in determining both the structural
attributes and therapeutic efficacy of β-glucans, offering critical
guidance for the development of β-glucan-based therapeutics.
Our results demonstrate that LMW-AP-FBG suppresses
the proliferation of SW480 cells primarily by inducing apoptotic
cell death. This conclusion is substantiated by cell viability
assays and Annexin V/7-AAD staining, which revealed a significant
increase in the proportion of cells undergoing early apoptosis,
while progression to late apoptosis was comparatively limited.
Importantly, administration of the pan-caspase inhibitor Z-VAD-FMK
inhibited apoptosis induction but resulted in only partial
restoration of cell viability. Collectively, these observations
indicate that, although apoptosis is a key mediator of the
antiproliferative effect, other pathways likely contribute. The
partial recovery of proliferation upon caspase inhibition suggests
involvement of additional caspase-independent cell death pathways,
such as AIF-mediated apoptosis or autophagy. Under physiological
conditions, the anti-apoptotic protein Bcl-2 inhibits mitochondrial
apoptosis by binding and sequestering the pro-apoptotic protein
Bax, thereby preventing Bax oligomerization and pore formation in
the mitochondrial outer membrane (34). In this study, exposure to
LMW-AP-FBG resulted in a pronounced decrease in Bcl-2 expression in
SW480 cells and was accompanied by a simultaneous increase in
cytosolic cytochrome c. This observation indicates that LMW-AP-FBG
compromises mitochondrial membrane integrity, promoting cytochrome
c release and subsequent activation of the intrinsic apoptotic
pathway. In addition to caspase-dependent signaling, the intrinsic
pathway includes caspase-independent mechanisms involving
mitochondrial proteins such as AIF and Endo G, which may
translocate to the nucleus (35,36).
However, our findings suggest that AIF-dependent signaling is not a
major contributor to LMW-AP-FBG-induced apoptosis. -Taken together,
these findings indicate that LMW-AP-FBG induces apoptosis in SW480
cells primarily through mechanisms associated with Bcl-2
downregulation and cytochrome c release rather than through
AIF-mediated caspase-independent pathways.
Of note, the extent of apoptosis observed did not
entirely explain the decrease in cell viability, implying that
other cell death modalities may also be operative. Supporting this
notion, Western blot analyses revealed elevated levels of
autophagy-related initiation and elongation proteins, including
BECN1, Atg5, Atg3, and Atg7, together with a dose-dependent
increase in LC3-II accumulation. The absence of significant
alteration in p62 expression indicates that, although LMW-AP-FBG
triggers autophagy, it may not fully promote autophagic flux.
Collectively, these findings indicate that autophagy, in parallel
with apoptosis, may be involved in the cellular response to
LMW-AP-FBG treatment and potentially contribute to its cytotoxic
effects in SW480 cells. Further studies employing lysosomal
inhibitors or LC3 turnover assays will be required to determine
whether this autophagic response primarily serves cytoprotective or
cytotoxic functions.
Our findings are consistent with a growing body of
evidence indicating that natural polysaccharides, such as β-glucans
and fucoidans, exert antitumor effects through modulation of
mitochondrial function, Bcl-2 family proteins, and
autophagy-related signaling pathways (37–39).
Significantly, concurrent engagement of apoptotic and
autophagy-related pathways by LMW-AP-FBG highlights its potential
as a multi-targeted anticancer candidate. The participation of
multiple cell death pathways in cancer cells is now widely
acknowledged as a key determinant of anticancer treatment efficacy
(40,41). Our results support the hypothesis
that LMW-AP-FBG may leverage these complementary mechanisms to
augment its anticancer properties.
The PI3K/AKT/mTOR pathway orchestrates essential
cellular functions, such as proliferation, energy metabolism, and
survival (30,31). Persistent activation of this
signaling axis has been associated with colorectal cancer
development and resistance to therapies (31,42).
Notably, inhibition of this pathway enhances apoptosis mediated by
mitochondria and increases autophagic processes across a range of
cancer cell systems (43). As
such, investigating this network in our study was crucial to
elucidate the molecular basis of the anticancer actions of
LMW-AP-FBG in SW480 colon cancer cells. We found that LMW-AP-FBG
treatment caused significant suppression of PI3K/AKT/mTOR
signaling, accompanied by enhanced apoptotic and autophagy-related
responses. These observations indicate that LMW-AP-FBG may modulate
two major cell death pathways, at least in part, through disruption
of a central signaling cascade. Collectively, these data broaden
the perspective on its molecular mechanisms and emphasize its
potential as a candidate for therapeutic intervention in CRC.
The findings of this study suggest that LMW-AP-FBG
may have potential clinical applications as an adjunct therapeutic
agent in colorectal cancer treatment. In particular, β-glucans are
known to modulate immune responses and enhance the efficacy of
conventional anticancer therapies. Therefore, LMW-AP-FBG may be
considered as a promising candidate for combination therapy with
existing chemotherapeutic agents or targeted therapies. However,
further in vivo and clinical studies are required to
validate its therapeutic efficacy and safety.
There are several limitations to this study. It
should be noted that the present findings were obtained using an
in vitro cell culture system. Although these results provide
important mechanistic insights into the anticancer effects of
LMW-AP-FBG, further studies using animal models and clinical
samples are required to confirm its therapeutic potential in
vivo. First, as our data are derived from in vitro
assays using a single colon cancer cell line, the results may not
accurately represent in vivo biology. Second, even though
both apoptosis and autophagy were observed, the distinct roles each
process plays in LMW-AP-FBG-induced cell death remain to be
clarified. Thus, subsequent investigations should focus on defining
the contribution of autophagy by employing pharmacological
inhibitors or genetic silencing approaches targeting
autophagy-related genes. Evaluation using in vivo xenograft
models will be required to further assess both the therapeutic
efficacy and safety profile of LMW-AP-FBG. In addition, a detailed
analysis of the crosstalk between Bcl-2 family members and
autophagy regulators could yield a deeper understanding of the
complex anticancer mechanisms underlying LMW-AP-FBG action.
Collectively, our results indicate that LMW-AP-FBG
exerts anticancer effects against SW480 cells through induction of
mitochondria-mediated apoptosis accompanied by engagement of
autophagy-related processes, a mechanism that is associated, at
least in part, with inhibition of the PI3K/AKT/mTOR pathway. These
findings underscore the potential of LMW-AP-FBG as a promising
natural candidate for anticancer applications and justify further
mechanistic and translational research.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was supported by a National Research Foundation of
Korea grant funded by the government of Korea (grant no.
RS-2023-NR076775) and by sabbatical research funding from Daegu
Catholic University in 2025.
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
JWP performed the experiments and prepared the
original draft of the manuscript. GMM, TMJ, JAS and TK performed
experiments and contributed to the methodology. HK contributed to
the conceptualization and validation of the study and critically
reviewed and edited the manuscript. JKK conceived and designed the
study, acquired funding, administered the project, and supervised
the review and editing of the manuscript. JWP, TMJ and JKK confirm
the authenticity of all the raw data. All authors have read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Olfatifar M, Rafiei F, Sadeghi A, Ataei E,
Habibi MA, Pezeshgi Modarres M, Ghalavand Z and Houri H: Assessing
the colorectal cancer landscape: A comprehensive exploration of
future trends in 216 countries and territories from 2021 to 2040. J
Epidemiol Glob Health. 15:52025. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bray F, Laversanne M, Sung H, Ferlay J,
Siegel RL, Soerjomataram I and Jemal A: Global cancer statistics
2022: GLOBOCAN estimates of incidence and mortality worldwide for
36 cancers in 185 countries. CA Cancer J Clin. 74:229–263.
2024.PubMed/NCBI
|
|
3
|
Park EH, Jung KW, Park NJ, Kang MJ, Yun
EH, Kim HJ, Kim JE, Kong HJ, Choi KS, Yang HK, et al: Cancer
statistics in Korea: Incidence, mortality, survival, and prevalence
in 2022. Cancer Res Treat. 57:312–330. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shin A, Jung KW and Jeong SY: Right then,
wrong now: Early-onset colorectal cancer in Korea. Cancer Res
Treat. 55:1058–1060. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Alese OB, Wu C, Chapin WJ, Ulanja MB,
Zheng-Lin B, Amankwah M and Eads J: Update on emerging therapies
for advanced colorectal cancer. Am Soc Clin Oncol Educ Book.
43:e3895742023. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shinji S, Yamada T, Matsuda A, Sonoda H,
Ohta R, Iwai T, Takeda K, Yonaga K, Masuda Y and Yoshida H: Recent
advances in the treatment of colorectal cancer: A review. J Nippon
Med Sch. 89:246–254. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
De Marco Castro E, Calder PC and Roche HM:
β-1,3/1,6-glucans and immunity: State of the art and future
directions. Mol Nutr Food Res. 65:e1901072021. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhong X, Wang G, Li F, Fang S, Zhou S,
Ishiwata A, Tonevitsky AG, Shkurnikov M, Cai H and Ding F:
Immunomodulatory effect and biological significance of β-glucans.
Pharmaceutic. 15:16152023. View Article : Google Scholar
|
|
9
|
Murphy EJ, Rezoagli E, Major I, Rowan NJ
and Laffey JG: β-Glucan metabolic and immunomodulatory properties
and potential for clinical application. J Fungi (Basel). 6:3562020.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rahmani J, Miri A, Černevičiūtė R,
Thompson J, de Souza NN, Sultana R, Kord Varkaneh H, Mousavi SM and
Hekmatdoost A: Effects of cereal beta-glucan consumption on body
weight, body mass index, waist circumference and total energy
intake: A meta-analysis of randomized controlled trials. Complement
Ther Med. 43:131–139. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Aoe S, Ichinose Y, Kohyama N, Komae K,
Takahashi A, Abe D, Yoshioka T and Yanagisawa T: Effects of high
β-glucan barley on visceral fat obesity in Japanese individuals: A
randomized, double-blind study. Nutrition. 42:1–6. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mishra AP, Salehi B, Sharifi-Rad M,
Pezzani R, Kobarfard F, Sharifi-Rad J and Nigam M: Programmed cell
death, from a cancer perspective: An overview. Mol Diagn Ther.
22:281–295. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vetvicka V, Teplyakova TV, Shintyapina AB
and Korolenko TA: Effects of medicinal fungi-derived β-glucan on
tumor progression. J Fungi (Basel). 7:2502021. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu Z, Wu XM, Luo YB, Li H, Zhou YQ, Liu ZQ
and Li ZY: Exploring the therapeutic potential of yeast β-glucan:
Prebiotic, anti-infective, and anticancer properties-A review. Int
J Biol Macromol. 283:1374362024. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ikewaki N, Sonoda T, Kurosawa G, Iwasaki
M, Devaprasad Dedeepiya V, Senthilkumar R, Preethy S and Abraham
SJK: Beta 1,3-1,6 glucans produced by two novel strains of
Aureobasidium pullulans exert immune and metabolic
beneficial effects in healthy middle-aged Japanese men: Results of
an exploratory randomized control study. JAR Life. 12:61–71.
2023.PubMed/NCBI
|
|
16
|
Jeong JH, Kim DJ, Hong SJ, Ahn JH, Lee DJ,
Jang AR, Kim S, Cho HJ, Lee JY, Park JH, et al: Investigating the
immune-stimulating potential of β-glucan from Aureobasidium
pullulans in cancer immunotherapy. Biomol Ther (Seoul).
32:556–567. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Suzuki T, Kusano K, Kondo N, Nishikawa K,
Kuge T and Ohno N: Biological activity of high-purity
β-1,3-1,6-glucan derived from the black yeast Aureobasidium
pullulans: A literature review. Nutrients. 13:2422021.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tada R, Yoshikawa M, Ikeda F, Adachi Y,
Kato Y, Kuge T, Tanioka A, Ishibashi KI, Tsubaki K and Ohno N:
Induction of IFN-γ by a highly branched 1,3-β-d-glucan from
Aureobasidium pullulans in mouse-derived splenocytes via
dectin-1-independent pathways. Biochem Biophys Res Commun.
404:1105–1110. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tanioka A, Tanabe K, Hosono A, Kawakami H,
Kaminogawa S, Tsubaki K and Hachimura S: Enhancement of intestinal
immune function in mice by β-D-glucan from Aureobasidium
pullulans ADK-34. Scand J Immunol. 78:61–68. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kim JH, Seo J, No H, Kuge T, Mori T,
Kimoto H and Kim JK: Low-molecular-weight β-1,3-1,6-glucan derived
from Aureobasidium pullulans exhibits anticancer activity by
inducing apoptosis in colorectal cancer cells. Biomedicines.
11:5292023. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
No H, Kim J, Seo CR, Lee DE, Kim JH, Kuge
T, Mori T, Kimoto H and Kim JK: Anti-inflammatory effects of
β-1,3-1,6-glucan derived from black yeast Aureobasidium
pullulans in RAW264.7 cells. Int J Biol Macromol. 193:592–600.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hayashi N, Shoubayashi Y, Kondo N and
Fukudome K: Hydrothermal processing of β-glucan from
Aureobasidium pullulans produces a low molecular weight
reagent that regulates inflammatory responses induced by TLR
ligands. Biochem Biophys Res Commun. 511:318–322. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kopiasz Ł, Dziendzikowska K, Oczkowski M,
Harasym J and Gromadzka-Ostrowska J: Low-molar-mass oat beta-glucan
impacts autophagy and apoptosis in early stages of induced
colorectal carcinogenesis in rats. Int J Biol Macromol.
254:1278322024. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Qian S, Long Y, Tan G, Li X, Xiang B, Tao
Y, Xie Z and Zhang X: Programmed cell death: Molecular mechanisms,
biological functions, diseases, and therapeutic targets. MedComm
(2020). 5:e700242024. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Saxena R, Welsh CM and He YW: Targeting
regulated cell death pathways in cancers for effective treatment: A
comprehensive review. Front Cell Dev Biol. 12:14623392024.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Park W, Wei S, Kim BS, Kim B, Bae SJ, Chae
YC, Ryu D and Ha KT: Diversity and complexity of cell death: A
historical review. Exp Mol Med. 55:1573–1594. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hu S, Meng Y, Guo L and Xu X: A novel
strategy to enhance inhibition of Hela cervical cancer by combining
Lentinus β-glucan and autophagic flux blockage. Int J Biol
Macromol. 281:1363092024. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang Y, Liu Y, Zhou Y, Zheng Z, Tang W,
Song M, Wang J and Wang K: Lentinan inhibited colon cancer growth
by inducing endoplasmic reticulum stress-mediated autophagic cell
death and apoptosis. Carbohydr Polym. 267:1181542021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pfeffer CM and Singh ATK: Apoptosis: A
target for anticancer therapy. Int J Mol Sci. 19:4482018.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fruman DA, Chiu H, Hopkins BD, Bagrodia S,
Cantley LC and Abraham RT: The PI3K pathway in human disease. Cell.
170:605–635. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Leiphrakpam PD and Are C: PI3K/Akt/mTOR
signaling pathway as a target for colorectal cancer treatment. Int
J Mol Sci. 25:31782024. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Elder MJ, Webster SJ, Chee R, Williams DL,
Hill Gaston JS and Goodall JC: β-Glucan size controls
dectin-1-mediated immune responses in human dendritic cells by
regulating IL-1β production. Front Immunol. 8:7912017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Luo Y, Geng J, Feng J, Liu L, Zhang J, Liu
Y and Guo Q: A comprehensive review on the structure-activity
relationships and applications of β-D-glucans. Bioact Carbohydr
Diet Fibre. 32:1004602024. View Article : Google Scholar
|
|
34
|
Shamas-Din A, Kale J, Leber B and Andrews
DW: Mechanisms of action of Bcl-2 family proteins. Cold Spring Harb
Perspect Biol. 5:a0087142013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Candé C, Vahsen N, Garrido C and Kroemer
G: Apoptosis-inducing factor (AIF): Caspase-independent after all.
Cell Death Differ. 11:591–595. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bano D and Prehn JHM: Apoptosis-Inducing
Factor (AIF) in physiology and disease: The tale of a repented
natural born killer. EBioMedicine. 30:29–37. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang N, Liu H, Liu G, Li M, He X, Yin C,
Tu Q, Shen X, Bai W, Wang Q, et al: Yeast β-D-glucan exerts
antitumour activity in liver cancer through impairing autophagy and
lysosomal function, promoting reactive oxygen species production
and apoptosis. Redox Biol. 32:1014952020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Park M, Bang C, Yun WS and Jeong YM:
Low-molecular-weight fucoidan inhibits the proliferation of
melanoma via Bcl-2 phosphorylation and PTEN/AKT pathway. Oncol Res.
32:273–282. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chantree P, Surarak T, Sangpairoj K,
Aguilar P and Hitakomate E: Antitumor effects of fucoidan via
apoptotic and autophagic induction on HSC-3 oral squamous cell
carcinoma. Asian Pac J Cancer Prev. 21:2469–2477. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pentimalli F, Grelli S, Di Daniele N,
Melino G and Amelio I: Cell death pathologies: Targeting death
pathways and the immune system for cancer therapy. Genes Immun.
20:539–554. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Santofimia-Castaño P and Iovanna J:
Combating pancreatic cancer chemoresistance by triggering multiple
cell death pathways. Pancreatology. 21:522–529. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Narayanankutty A: PI3K/Akt/mTOR pathway as
a therapeutic target for colorectal cancer: A review of preclinical
and clinical evidence. Curr Drug Targets. 20:1217–1226. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Meng Y, Si Y, Guo T, Zhao W, Zhang L, Wang
Y, Wang L, Sun K and Feng S: Ethoxychelerythrine as a potential
therapeutic strategy targets PI3K/AKT/mTOR induced mitochondrial
apoptosis in the treatment of colorectal cancer. Sci Rep.
15:66422025. View Article : Google Scholar : PubMed/NCBI
|