Introduction
Breast cancer is the most commonly diagnosed
malignancy worldwide, with ~2.3 million novel cases and 685,000
mortalities reported in 2020 (1).
Among its subtypes, triple-negative breast cancer, defined by the
absence of estrogen receptor, progesterone receptor and HER2
expression, is the most aggressive and treatment resistant, largely
due to its high metastatic potential (2). Although surgery, radiotherapy,
chemotherapy, targeted therapy and immunotherapy are extensively
used in breast cancer management, their effectiveness is often
limited by adverse effects and suboptimal outcomes, highlighting
the need for safer and more effective therapeutic strategies in the
future (3).
Natural bioactive compounds have gained attention as
anticancer agents due to their pleiotropic effects, ability to
synergize with conventional chemotherapeutics and capacity to
target multiple cancer-related pathways (4,5). For
instance, medicinal plant bioactive compounds exhibit anticancer
potential by suppressing the G2/M and G0/G1 phases,
inducing cell cycle arrest, and promoting apoptosis (5). Among these compounds, polysaccharides
are biologically key macromolecules with well-documented
antioxidant, immunomodulatory, anticancer and anti-inflammatory
properties (6–8). Since their anticancer potential was
first recognized, numerous polysaccharides have been developed as
adjuvant cancer therapies, including those derived from Trametes
versicolor (synonym, Coriolus versicolor), fucoidan and
sepia ink, which are notable for their low toxicity and favorable
safety profiles (7). Sulfated
galactan (SG), a polysaccharide isolated from Gracilaria
fisheri, has been reported to inhibit the proliferation and
migration of human cholangiocarcinoma HuCCA-1 cells through
interaction with and inactivation of EGFR signaling (8). Furthermore, SG suppresses
proliferation of human breast cancer MCF-7 cells by inducing cell
cycle arrest, without cytotoxic effects on normal fibroblast L292
cells (9).
Immunotherapy has transformed cancer treatment and
improved outcomes across multiple malignancies such as melanoma and
breast cancer (10,11); however, majority of patients fail
to respond because several tumors exhibit low intrinsic
immunogenicity, enabling immune evasion (12). Immunogenic cell death (ICD) is a
regulated form of cell death that increases tumor immunogenicity by
promoting the release of damage-associated molecular patterns
(DAMPs) and activating tumor-specific immune responses within the
tumor microenvironment. Beyond direct tumor cell killing, ICD
inducers enhance tumor antigenicity and adjuvanticity, thereby
stimulating both innate and adaptive antitumor immunity (13,14).
ICD is primarily driven by endoplasmic reticulum (ER) stress and/or
reactive oxygen species (ROS), leading to the exposure or release
of key DAMPs, such as calreticulin (CRT) (15). Furthermore, Fas receptor (Fas-R)
and major histocompatibility complex class I (MHC class I) are key
mediators of immune-dependent tumor cell killing and immune
surveillance, with their expression levels influencing tumor
sensitivity to immunotherapy (16,17).
Sulfated polysaccharides exhibit notable
anti-inflammatory and immunostimulatory properties by modulating
immune responses, regulating oxidative stress and inhibiting
pro-inflammatory signaling pathways (18,19).
Notably, SG from Gracilaria fisheri has been reported to
enhance immunostimulatory activity in murine J774A1 macrophages by
increasing the secretion of pro-inflammatory cytokines, including
TNF-α, IL-1β and IL-6 (20),
suggesting its potential to induce ICD. However, despite increasing
evidence that supports the antiproliferative, anticancer and
immunomodulatory effects of SG (8,9,20),
its ability to induce ICD has not been elucidated. Furthermore, the
biological activity of SG is notably dependent on its structural
characteristics, particularly molecular weight (21). To date, no studies have
investigated whether SG or its degraded derivative (DSG) induce ICD
in triple-negative breast cancer cells, to the best of our
knowledge. Therefore, the present study aimed to investigate the
microstructural features and ICD-inducing effects of SG and DSG
from Gracilaria fisheri in triple-negative breast cancer
cells (MDA-MB-231).
Materials and methods
Gracilaria fisheri SG and DSG
SG and DSG were derived from Gracilaria
fisheri, which was collected from the Shrimp Genetic
Improvement Center (Surat Thani, Thailand) between October and
December 2024. The polysaccharides were prepared according to
previously established methods (21,22).
SG had a molecular weight of 217 kDa, whereas DSG had a molecular
weight of 8 kDa, as determined by gel permeation chromatography
(Fig. S1). The structures of SG
and DSG were confirmed by 1H- and 13C-nuclear
magnetic resonance (Bruker Corporation) analyses. Both compounds
consist of alternating 3-linked β-D-galactopyranose and 4-linked
3,6-anhydro-α-L-galactopyranose or α-L-galactopyranose-6-sulfate
residues, as presented in Fig.
S2, consistent with previous studies (21,22).
Analysis of surface morphology and
elemental composition of SG and DSG
The surface morphology and elemental composition of
SG and DSG were examined using scanning electron microscopy coupled
with energy-dispersive X-ray spectroscopy (SEM-EDX). Samples (5–10
µg) were deposited onto SEM stubs and gold-coated for 2 min at a
coating current of 20 mA. Surface morphology was visualized using
an SEM (JSM-IT200™; JEOL, Ltd.) and elemental composition was
analyzed using an EDX detector (Ultim® Max 40; Oxford
Instruments plc). For EDX measurements, the accelerating voltage
was set to 10 kV, with a working distance of ~8.8 mm. Spectra were
acquired from randomly selected regions of each sample to ensure
representative elemental characterization. At least three
independent areas were analyzed and the acquisition time for each
spectrum was ~30 sec. Elemental mapping and point analyses were
performed using the instrument software (AZtec® version
6.1; Oxford Instruments) to identify and quantify the elemental
distribution on the sample surface. The elemental composition was
reported as weight (%).
Sulfate content and Fourier-transform
infrared (FTIR) spectroscopy analysis
Sulfate groups in SG and DSG were quantified using
the barium chloride (BaCl2)-gelatin method and confirmed
by FTIR spectroscopy. Samples (20 mg) were hydrolyzed with 2 N HCl
at 100°C for 2 h, centrifuged (3,000 × g, for 10 min at room
temperature), and the supernatants were diluted with Milli-Q water
and 0.5 N HCl. BaCl2-gelatin reagent (1 ml) was added to
the diluted supernatants, and the mixtures were incubated at room
temperature for 30 min. Absorbance was measured at 550 nm to
determine sulfate content as described previously (22). For FTIR analysis, SG and DSG (2 mg)
were mixed with dried potassium bromide to prepare pellets and
spectra were recorded using a Bruker TENSOR 27 spectrometer
(attenuated total reflectance mode) over the range of 400–4,000
cm-1 at a resolution of 1 cm-1 with 16
scans.
Cell culture and cytotoxicity
evaluation
Mammary epithelial cells (MCF-10A; cat. no.
CRL-10317™) and breast cancer cells (MDA-MB-231; cat. no. HTB-26™)
were obtained from the American Type Culture Collection. Cells were
cultured in Dulbecco's Modified Eagle Medium (Gibco™; Thermo Fisher
Scientific, Inc.) supplemented with 10% fetal bovine serum
(Invitrogen; Thermo Fisher Scientific, Inc.) and 0.5 M sodium
bicarbonate in a humidified incubator at 37°C with 5%
CO2. The cytotoxicity of SG, DSG and doxorubicin [DOXO;
Fresenius Kabi (Thailand) Ltd.], used as a positive control, was
evaluated using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl
tetrazolium bromide (MTT) assay. Cells were seeded in 96-well
plates (3×104 cells/well; 200 µl per well) and cultured
overnight at 37°C. Cells were then treated with various
concentrations of SG or DSG (100–1,000 µg/ml) or DOXO (0.02–1.50
µg/ml) for 24 h at 37°C. Cell viability was assessed using the MTT
assay, in which dimethyl sulfoxide (DMSO) was used to solubilize
formazan crystals, and absorbance was measured at OD 595 nm.
Cytotoxicity was expressed as the 50% cytotoxic concentration.
Cell morphology and membrane
permeability evaluation
Morphological changes in MCF-10A and MDA-MB-231
cells following treatment with DOXO, SG or DSG were evaluated.
Cells (3×106 cells/well) were seeded in 6-well plates
and cultured for 24 h at 37°C, then assigned to four groups: i)
Normal control (NC); ii) DOXO (0.5 µg/ml); iii) SG (1,000 µg/ml);
and iv) DSG (1,000 µg/ml). After 24 h of treatment, cell morphology
was examined using phase-contrast microscopy (Leica DFC 7000 T;
Leica Microsystems GmbH). Morphological changes were quantified in
three randomly selected fields at ×200 magnification using ImageJ
software (version 1.54g; National Institutes of Health). To further
assess membrane permeability in MDA-MB-231 cells, Hoechst/propidium
iodide (PI) dual staining was performed. Cells (6×104
cells/well) were seeded onto round coverslips in 24-well plates and
cultured for 24 h at 37°C, followed by an additional 24 h of
treatment at 37°C with DOXO, SG or DSG. Cells were then washed with
PBS and stained with Hoechst (Merck KGaA; MilliporeSigma) and PI
(BioChemica; PanReac AppliChem) at a 1:1 µg/ml ratio for 30 min at
room temperature in the dark. After washing, coverslips were
mounted with antifade medium (Invitrogen; Thermo Fisher Scientific,
Inc.) and stained cells were visualized using a confocal microscope
(ZEISS LSM 800; Carl Zeiss AG), as described previously (21). PI fluorescence intensity was
quantified in three randomly selected fields at ×200 magnification
using ImageJ software (version 1.54g; National Institutes of
Health).
Intracellular ROS generation
evaluation
Based on cytotoxicity, morphological and membrane
permeability results, intracellular ROS generation was further
assessed in MDA-MB-231 cells using an ROS detection assay kit
(BioVision, Inc.; Abcam). Following treatment with DOXO, SG or DSG,
cells were washed with 100 µl of ROS assay buffer and incubated
with 100 µl of 1X ROS labeling solution containing the DCFH-DA
fluorophore (BioVision, Inc.; Abcam) diluted in assay buffer at
37°C for 45 min in the dark. Cells were then washed again with ROS
assay buffer and fluorescence was measured immediately at
excitation/emission wavelengths of 495/529 nm. Intracellular ROS
levels were expressed as fold changes compared with the
control.
Cell ultrastructure evaluation by
transmission electron microscopy (TEM)
MDA-MB-231 cells were further examined for
ultrastructural changes following compound treatment using TEM
analysis. After treatment with DOXO, SG or DSG, cells were fixed in
Karnovsky's fixative (2.5% glutaraldehyde+2.0% paraformaldehyde, in
0.1 M sodium cacodylate buffer, pH 7.4) for 30 min at room
temperature, washed with PBS and centrifuged at 1,500 × g for 1 min
at 25°C. Cells were post-fixed with 1% OSO4
in 0.1 M phosphate buffer for 1 h at room temperature, washed with
PBS and centrifuged at 1,500 × g, for 1 min at room temperature.
Cell pellets were embedded in 2% agarose for 20 min at room
temperature, cut into small pieces and dehydrated using a graded
ethanol series. Samples were infiltrated with propylene oxide and
propylene oxide/EPON 812 mixtures, followed by pure EPON 812
overnight, then embedded and polymerized at 60°C for 48 h.
Ultrathin sections were prepared, stained with uranyl acetate and
lead citrate, and examined using JEM-1010-TEM (JEOL Ltd.).
Western blotting analysis of ICD
markers
After 6, 12 and 24 h of treatment, MDA-MB-231 cells
were harvested and proteins were extracted using a protein lysis
buffer (20 mM Tris-HCl, 100 mM NaCl, 5 mM phenylmethylsulfonyl
fluoride; PMSF) supplemented with a 100X protease inhibitor
solution. Protein concentration was determined using a NanoDrop™
2000 spectrophotometer (Thermo Fisher Scientific, Inc.) by
measuring absorbance at 280 nm. Equal amounts of protein (50 µg per
sample) were separated by 12.5% SDS-PAGE and transferred onto
nitrocellulose membranes (MilliporeSigma; Merck KGaA). Membranes
were blocked with 2% BSA (cat. no. BSA-1S; Capricorn Scientific) in
PBS for 2 h at room temperature and then incubated overnight at 4°C
with primary antibodies (1:1,000 dilution) against CRT (cat. no.
MA5-15382), Fas-R (cat. no. MA5-32489) and MHC class I (cat. no.
MA5-35712; Invitrogen; Thermo Fisher Scientific, Inc.), followed by
HRP-conjugated goat anti-rabbit secondary antibody (cat. no. 31460;
1:2,000 dilution; Invitrogen; Thermo Fisher Scientific, Inc.) for 1
h at room temperature. Immunoreactive bands were visualized using
Clarity™ Western ECL substrate (Bio-Rad Laboratories, Inc.).
Protein expression levels were normalized to β-actin (cat. no.
AF7018; Affinity Biosciences, Inc.) and quantified by densitometric
analysis using Scion Image version 4.0.2 software (Scion
Corporation, Frederick, MD, USA). All experiments were performed in
triplicate.
Immunofluorescent staining of ICD
Immunofluorescence staining was performed to assess
the expression levels and localization of CRT, Fas-R and MHC class
I in MDA-MB-231 cells. Cells grown on coated round glass coverslips
were treated with DOXO (0.5 µg/ml) or DSG (1,000 µg/ml). After 12 h
of treatment, cells were washed with PBS and fixed with 4%
paraformaldehyde for 20 min at room temperature. After washing,
cells were incubated overnight at 4°C with primary antibodies
against CRT, Fas-R and MHC class I. Cells were then incubated with
a FITC-conjugated goat anti-rabbit IgG secondary antibody (1:500;
cat. no. 701078; Invitrogen; Thermo Fisher Scientific, Inc.) for 1
h at room temperature. Negative controls were prepared by omitting
the primary antibody. Cell membranes were counterstained with
CellMask™ Deep Red Plasma Membrane (cat. no. C10046; Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions and nuclei were stained using ProLong™ Diamond
Antifade Mountant with DAPI (Invitrogen; Thermo Fisher Scientific,
Inc.) for 5 min at room temperature. Immunofluorescence images were
acquired using a confocal microscope (ZEISS LSM 800; Carl Zeiss
AG). FITC fluorescence intensity for CRT, Fas-R and MHC class I was
quantified in three randomly selected fields at ×200 magnification
using ImageJ software (version 1.54g; National Institutes of
Health).
Determination of ICD-related genes by
reverse transcription-quantitative PCR (RT-qPCR) analysis
Total RNA was extracted using TRIzol®
reagent (200 µl; MilliporeSigma) and RNA purity and concentration
were assessed by measuring the A260/280 ratio using a NanoDrop™
2000 spectrophotometer (Invitrogen; Thermo Fisher Scientific,
Inc.). Complementary DNA (cDNA) was synthesized from 1 µg of RNA
using the RevertAid™ First Strand cDNA Synthesis Kit (Invitrogen;
Thermo Fisher Scientific, Inc.) by incubation at 42°C for 60 min,
followed by 70°C for 5 min. Gene expression was analyzed by RT-qPCR
using synthesized cDNA, SYBR Green PCR Master Mix (Invitrogen;
Thermo Fisher Scientific, Inc.) and gene-specific forward and
reverse primers. The RT-qPCR thermocycling conditions were as
follows: 50°C for 2 min, 95°C for 10 min and 40 cycles of 95°C for
15 sec, 60°C for 30 sec and 72°C for 30 sec. Relative mRNA
expression levels were calculated using the 2-ΔΔCq
method (23) and analyzed using
Bio-Rad CFX Maestro software version 2.3 (Bio-Rad Laboratories,
Inc.). Primers targeting protein kinase RNA-like ER kinase (PERK),
inositol-requiring enzyme 1 (IRE1), activating transcription factor
(ATF)6, ATF4, eukaryotic initiation factor 2 α subunit (eIF2α),
CRT, Fas-R, MHC class I and GAPDH were obtained from BIONICS Co.,
Ltd. Primer sequences were designed using National Center for
Biotechnology Information Primer-Basic Local Alignment Search Tool
and are listed in Table I.
 | Table I.Primer sequences used in the present
study. |
Table I.
Primer sequences used in the present
study.
| Gene | Primer sequences
(5′-3′) |
|---|
| PERK | F:
ATTGCATCTGCCTGGTTAC |
|
| R:
GACTCCTTCCTTTGCCTGT |
| IRE1 | F:
GACAGGCTCAATCAAATGG |
|
| R:
CGGTCAGGAGGTCAATAACA |
| ATF4 | F:
CCAGCAAAGCACCGCAACA |
|
| R:
CCATCCACAGCCAGCCATT |
| ATF6 | F:
CAGGGAGAAGGAACTTGTGA |
|
| R:
ACTGACCGAGGAGACGAGA |
| elF2α | F:
AAGGCGTATCCGTTCTATCA |
|
| R:
CTTCCCGTTCATCTTCATTC |
| CRT | F:
TCAAGGAGCAGTTTCTGGACGG |
|
| R:
GCATCCTGGCTTGTCTGCAAAC |
| Fas-R | F:
TGAAGGACATGGCTTAGAAGTG |
|
| R:
GGTGCAAGGGTCACAGTGTT |
| MHC class I | F:
CAGTTCGTGAGGTTCGACAG |
|
| R:
CAGCCGTACATGCTCTGGA |
| GAPDH | F:
GGTGAAGGTCGGTGTGAACG |
|
| R:
CTCGCTCCTGGAAGATGGTG |
Statistical analysis
Data are presented as the mean±standard error of the
mean from three or more independent experiments. Statistical
significance was assessed using one-way analysis of variance
followed by Tukey's multiple comparison test in GraphPad Prism
software (version 10.3.1.509; Dotmatics). P<0.05 was considered
to indicate a statistically significant difference.
Results
Surface morphology and elemental
composition of Gracilaria fisheri SG and DSG
SEM-EDX was used to examine the microstructure and
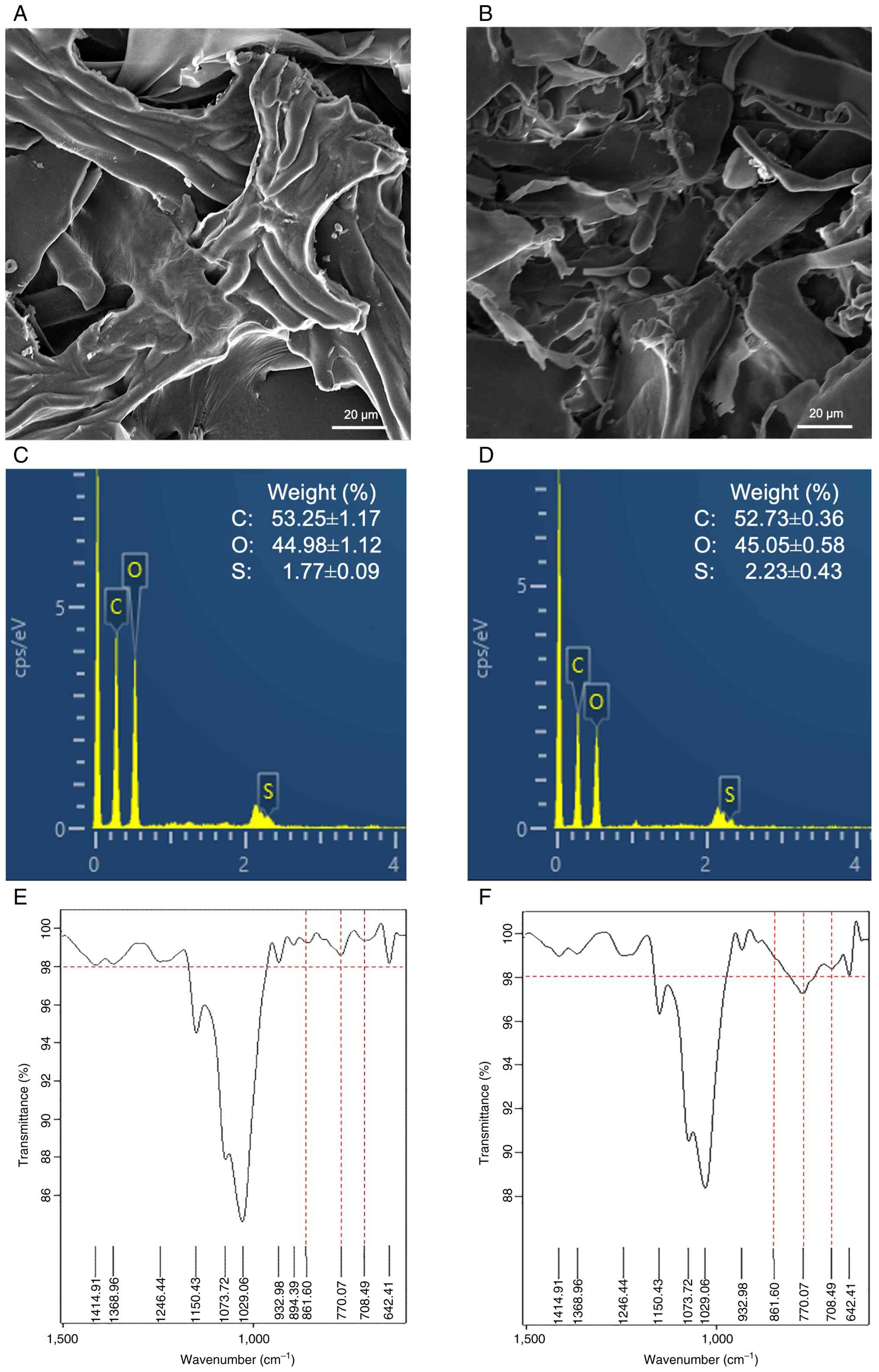
elemental composition of SG and DSG (24). As presented in Fig. 1A and B, SG and DSG exhibited
distinct surface morphologies. SG appeared as large lamellar,
bundle-like fiber structures, whereas DSG consisted of shorter,
fragmented sheet-like structures with irregular shapes. EDX
analysis confirmed the presence of carbon, oxygen and sulfur in
both samples (Fig. 1C and D). In
SG, carbon, oxygen and sulfur contents were 53.25±1.17%,
44.98±1.12% and 1.77±0.09%, respectively, whereas in DSG these
values were 52.73±0.36%, 45.05±0.58% and 2.23±0.43%. The sulfate
contents of SG and DSG were further quantified as 11.41±0.14% and
12.92±0.07%, respectively (Fig.
S1). FTIR analysis revealed characteristic absorption bands
between 400 and 4,000 cm-1, consistent with
polysaccharide functional groups (Fig. S2). Notably, absorbance bands at
861.60, 770.07 and 708.49 cm-1, corresponding to sulfate
group stretching vibrations (21),
were more notable in DSG (Fig. 1E and
F). These results confirmed the sulfated nature of both
polysaccharides and suggested that degradation to a lower molecular
weight is associated with increased sulfate content within the
structure.
Effects of SG and DSG on cell
viability in normal breast epithelial and human breast cancer
cells
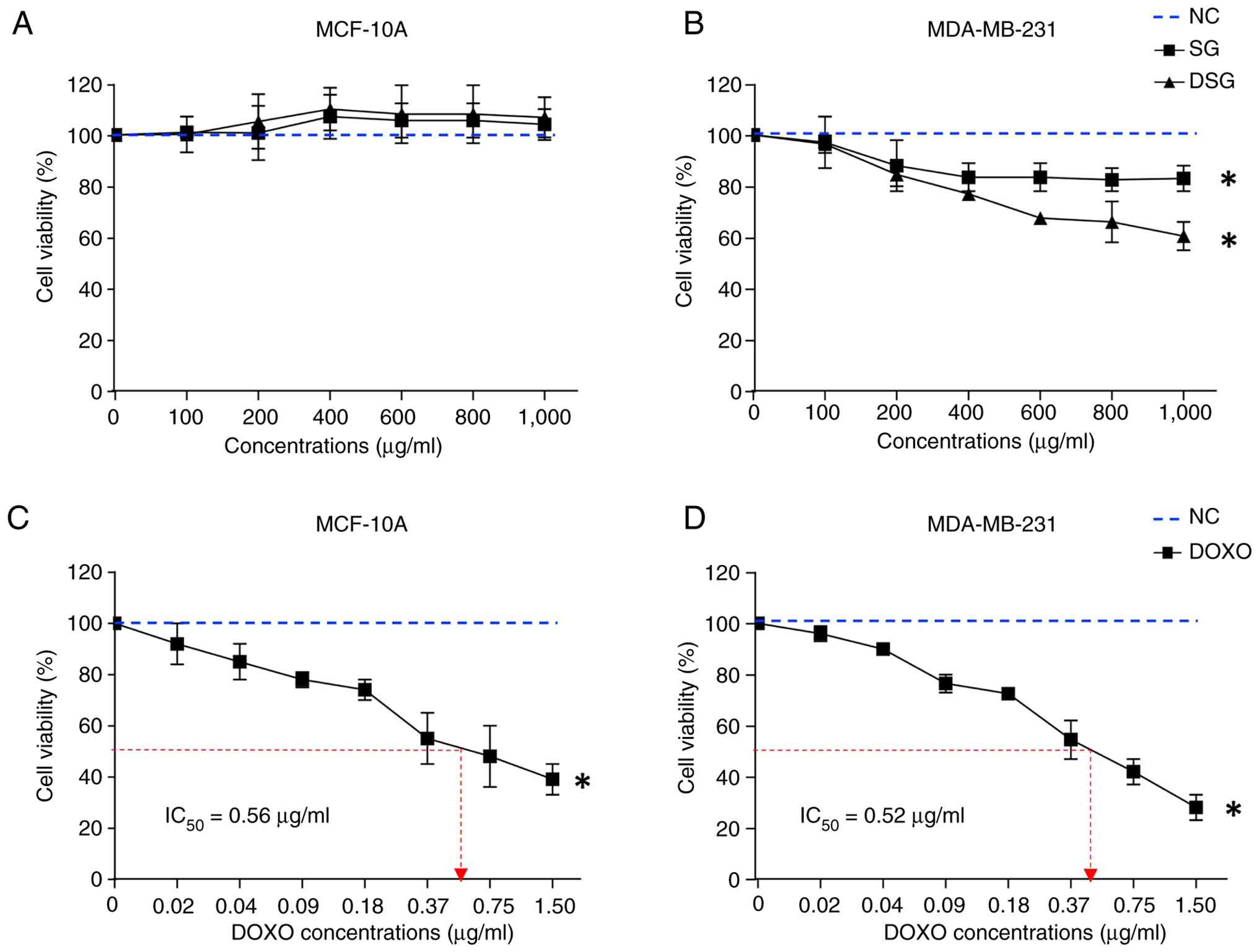
The cytotoxic effects of SG and DSG on normal breast
epithelial MCF-10A cells and MDA-MB-231 breast cancer cells were
evaluated after 24 h of exposure and expressed as a percentage of
control at concentrations ranging from 100 to 1,000 µg/ml. SG and
DSG exhibited no significant cytotoxicity toward MCF-10A cells at
concentrations up to 1,000 µg/ml (Fig.
2A). By contrast, both compounds significantly reduced
viability in MDA-MB-231 cells at concentrations of 400–1,000 µg/ml,
with DSG exhibiting greater cytotoxic potency compared with SG
(Fig. 2B). The IC50
values for SG and DSG in MDA-MB-231 cells were 2,736.17 and
1,927.11 µg/ml, respectively, calculated from the slope and
equation obtained from Fig. 2B.
DOXO, used as a positive control, significantly reduced the
viability of both MCF-10A and MDA-MB-231 cells, with
IC50 values of 0.56 and 0.52 µg/ml, respectively
(Fig. 2C and D). Since SG and DSG
did not achieve 50% cytotoxicity at the highest concentration
tested, IC50 values could not be determined within the
experimental range. Therefore, a concentration of 1,000 µg/ml was
selected for subsequent experiments.
Effects of SG and DSG on morphology
and intracellular ROS generation in MDA-MB-231 cells
To assess the effects of SG and DSG on cellular
morphology, morphological features and membrane permeability in
MDA-MB-231 cells were examined. Phase-contrast microscopy revealed
that MDA-MB-231 cells treated with DOXO, SG and DSG (Fig. 3A) exhibited clear morphological
changes compared with untreated controls, including cell shrinkage,
rounding and detachment. By contrast, MCF-10A cells treated with SG
or DSG indicated no noticeable morphological alterations compared
with controls (Fig. S3). As
presented in Fig. 3B, MDA-MB-231
cells treated with SG and DSG displayed prominent cytoplasmic PI
fluorescence, similar to that observed in DOXO-treated cells,
indicating increased membrane permeability consistent with cell
death. Quantitative analysis of morphological changes and PI
fluorescence intensity in MDA-MB-231 cells demonstrated a
significantly higher incidence in the order of DOXO>DSG>SG
(Fig. 3C and D).
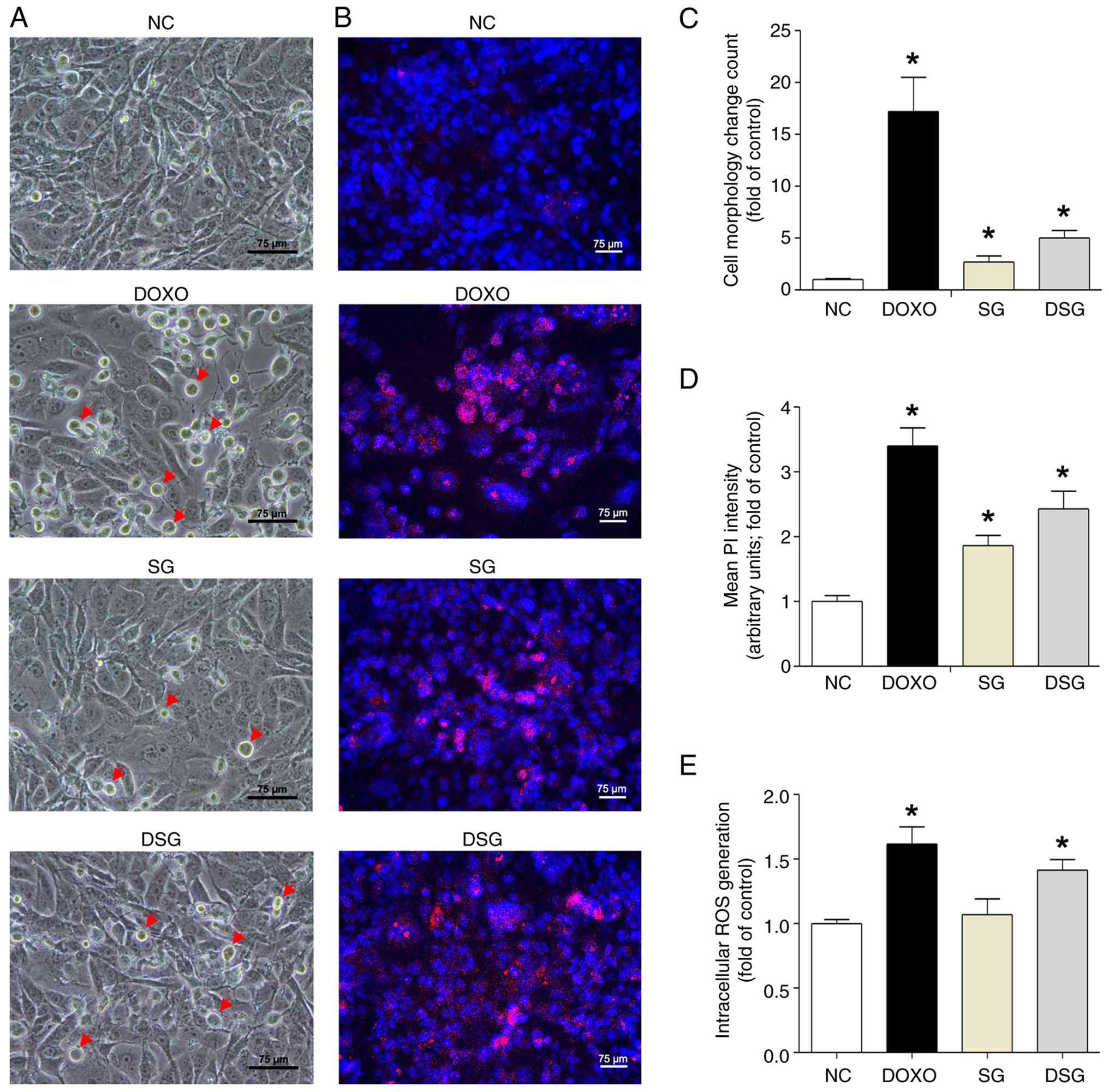 | Figure 3.Cell morphology, membrane
permeability alterations and intracellular ROS generation in
MDA-MB-231 cells following treatment with DOXO (0.5 µg/ml), SG and
DSG (both 1,000 µg/ml) for 24 h. (A) Phase-contrast micrographs
exhibiting morphological features of MDA-MB-231 cells after
treatment; red arrows indicate cells exhibiting morphological
changes (scale bar, 75 µm). (B) Immunofluorescence micrographs
indicating Hoechst and PI dual staining in MDA-MB-231 cells after
treatment (scale bar, 75 µm). (C) Quantitative analysis of
morphological changes in MDA-MB-231 cells treated with DOXO, SG and
DSG. (D) Quantitative analysis of PI penetration intensity in
MDA-MB-231 cells treated with DOXO, SG and DSG. (E) Quantitative
analysis of intracellular ROS generation in MDA-MB-231 cells after
treatment with DOXO, SG and DSG. Results are presented as the
mean±standard error of the mean (n=3) from three independent
experiments. *P<0.05 indicates a statistically significant
difference compared with the NC group. SG, sulfated galactan; DSG,
degraded sulfated galactan; NC, normal control; ROS, reactive
oxygen species; DOXO, doxorubicin; PI, propidium iodide. |
Furthermore, ROS levels exceeding the antioxidant
capacity of cancer cells can cause biomolecular damage and trigger
cell death (25). The present
study evaluated whether SG and DSG increase intracellular ROS
levels. The effects of DOXO, SG and DSG on intracellular ROS
generation in MDA-MB-231 cells are presented in Fig. 3E. Cells treated with DOXO and DSG
exhibited significantly higher ROS levels (1.62-fold for DOXO and
1.41-fold for DSG compared with the control), whereas SG-treated
cells exhibited ROS levels similar to those of the control. These
findings suggested that DSG induces changes in cell morphology and
membrane permeability in MDA-MB-231 cells through increased
intracellular ROS generation.
Ultrastructural alterations in
MDA-MB-231 cells induced by SG and DSG
To further characterize the ultrastructural effects
of SG and DSG on MDA-MB-231 cells, TEM analysis was performed. As
presented in Fig. 4, distinct
ultrastructural differences were observed among the NC, DOXO-, SG-
and DSG-treated groups. Cells in the NC group exhibited
well-preserved cellular architecture, including intact nuclear
membranes, prominent nucleoli, evenly distributed chromatin and a
uniform, electron-lucent cytoplasm (Fig. 4A). By contrast, DOXO-treated cells
revealed marked ultrastructural damage, characterized by
cytoplasmic condensation, extensive vacuolization and accumulation
of electron-dense bodies, consistent with severe cellular stress
and cytotoxicity (Fig. 4B).
SG-treated cells largely maintained ultrastructural features
similar to those of the NC group, indicating minimal cytotoxic
effects (Fig. 4C). By contrast,
DSG-treated cells retained nuclear integrity but exhibited notable
cytoplasmic alterations, including extensive vacuolization,
electron-dense inclusions and organelle disorganization (Fig. 4D). These findings indicated that
DSG induces notable ultrastructural damage in MDA-MB-231 cells,
with cytotoxic effects similar to those observed with DOXO.
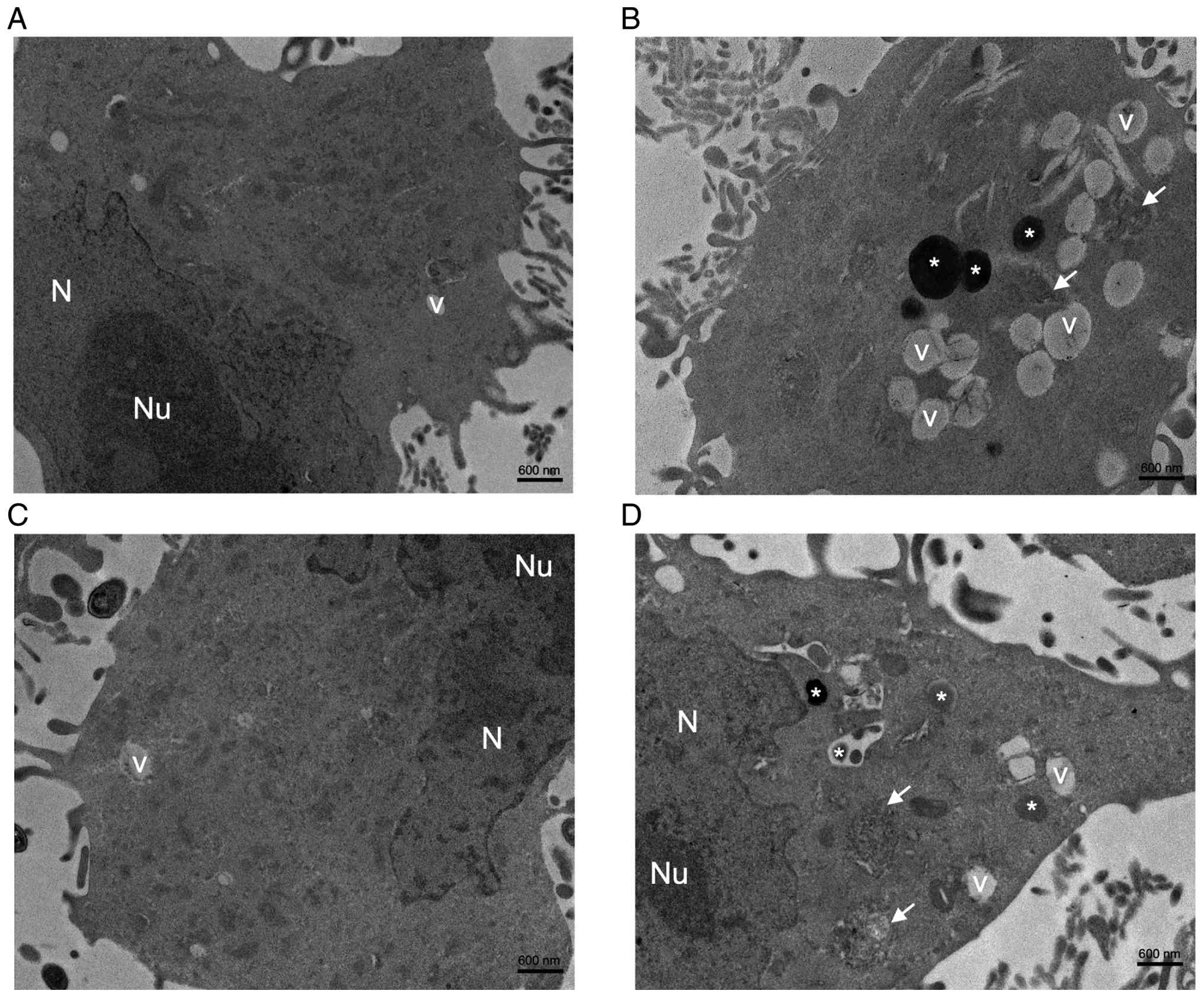 | Figure 4.TEM micrographs of cultured
MDA-MB-231 cells after treatment with DOXO (0.5 µg/ml), SG and DSG
(both 1,000 µg/ml) for 24 h. TEM micrographs indicate (A) NC (scale
bar, 600 nm), (B) DOXO-treated MDA-MB-231 cells (scale bar, 600
nm), (C) SG-treated MDA-MB-231 cells (scale bar, 600 nm) and (D)
DSG-treated MDA-MB-231 cells (scale bar, 600 nm). SG, sulfated
galactan; DSG, degraded sulfated galactan; N, nucleus; Nu,
nucleolus; V, vacuole; *, electron-dense bodies; white arrow,
organelle disorganization; TEM, transmission electron
microscopy. |
Effects of DSG on ICD and its
underlying mechanisms in MDA-MB-231 cells
ICD is a regulated form of cell death that
stimulates tumor-specific immunity through the release of DAMPs
from dying cancer cells, thereby activating immune responses within
the tumor microenvironment. ICD can be induced by several
conventional therapies, including chemotherapy, targeted therapy,
radiotherapy and photodynamic therapy (15). Therefore, the present study
examined the effects of DSG on ICD-related marker expression and
the underlying mechanisms in MDA-MB-231 cells. DSG treatment
increased the expression levels of CRT and Fas-R, as presented in
Figs. 5 and 6. Western blotting analysis demonstrated
that DSG significantly upregulated CRT and Fas-R expression levels
at 12 and 24 h, whereas expression level of MHC class I did not
differ compared with that in control cells (Fig. 5). The DSG-induced increases in CRT
and Fas-R expression levels were similar to those observed in
DOXO-treated cells, a known ICD inducer (26). Specifically, CRT expression
increased to 1.46±0.41-fold at 6 h, 1.39±0.24-fold at 12 h and
2.09±0.26-fold at 24 h following DSG treatment compared with the
control. Fas-R expression increased to 1.38±0.09-fold at 12 h and
2.14±0.74-fold at 24 h following DSG treatment.
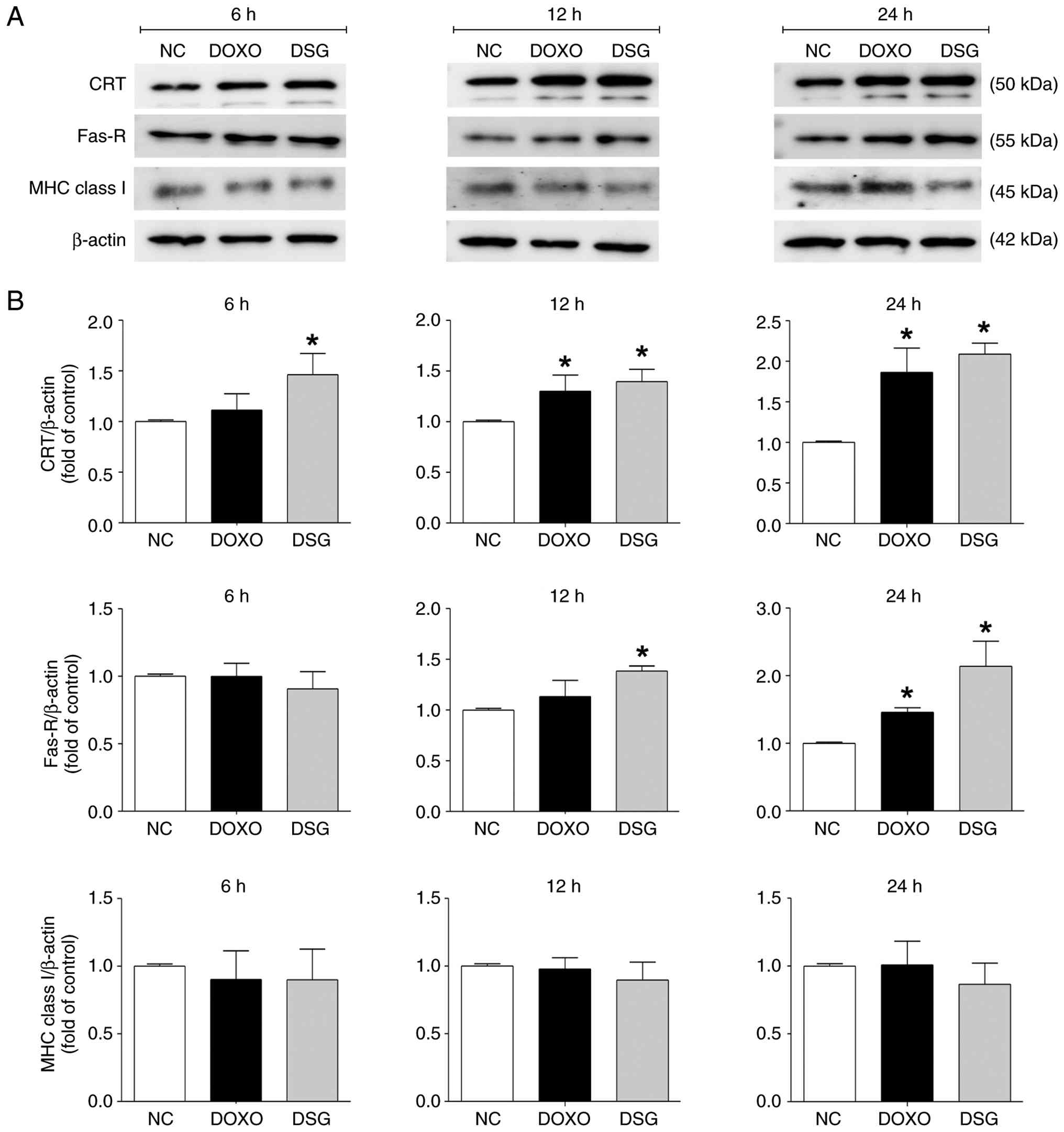 | Figure 5.Expression levels of immunogenic cell
death-related proteins in MDA-MB-231 cells treated with DOXO and
DSG for 6, 12 and 24 h, as assessed by western blotting analysis.
(A) Representative immunoblot bands of CRT, Fas-R and MHC class I
in MDA-MB-231 cells from the experimental groups (NC, DOXO and
DSG). (B) Quantitative analysis of CRT, Fas-R and MHC class I
expression levels normalized to β-actin, with fold changes
calculated relative to the NC. Results are presented as the
mean±standard error of the mean (n=3) from three independent
experiments. *P<0.05 indicates a statistically significant
difference compared with the NC group. DSG, degraded sulfated
galactan; NC, normal control; DOXO, doxorubicin; CRT, calreticulin;
Fas-R, Fas receptor; MHC, major histocompatibility complex. |
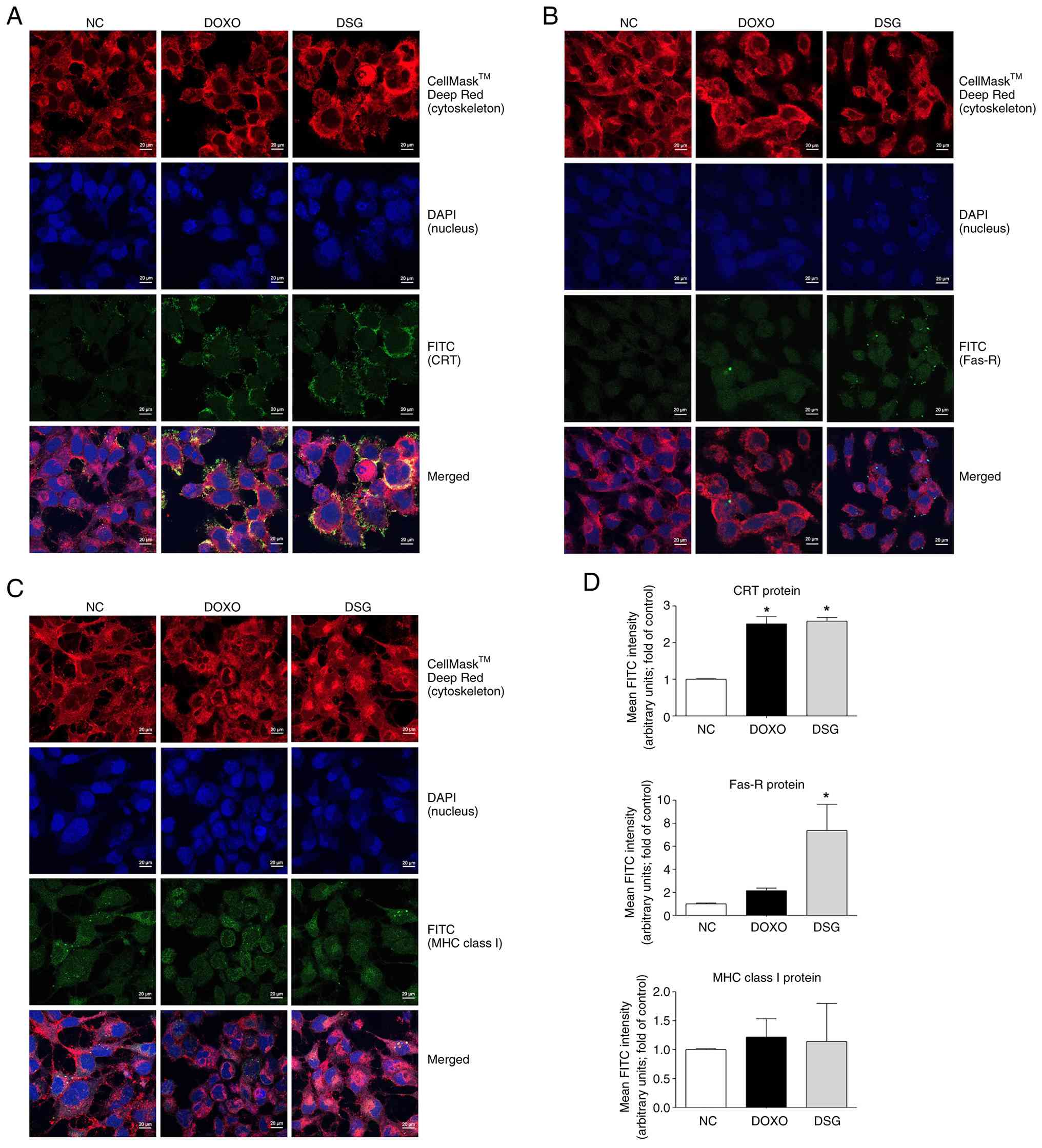 | Figure 6.Expression and localization of
immunogenic cell death-related proteins in MDA-MB-231 cells treated
with DOXO and DSG for 12 h, as assessed by immunofluorescence
staining. Confocal immunofluorescence micrographs indicate the
expression and subcellular localization of CRT (A), Fas-R (B) and
MHC class I (C) in MDA-MB-231 cells following treatment. Target
proteins (CRT, Fas-R and MHC class I) are shown in green, nuclei
are stained with DAPI (blue) and the plasma membrane is stained
with CellMask™ Deep Red (red) (Scale bar, 20 µm). (D) Quantitative
analysis of FITC fluorescence intensity for CRT, Fas-R and MHC
class I in MDA-MB-231 cells following treatment. Data are presented
as the mean±standard error of the mean (n=3) from three independent
experiments. *P<0.05 indicates a statistically significant
difference compared with the NC group. DSG, degraded sulfated
galactan; NC, normal control; DOXO, doxorubicin; CRT, calreticulin;
Fas-R, Fas receptor; MHC, major histocompatibility complex. |
Furthermore, confocal immunofluorescence staining
was performed to confirm the expression levels and subcellular
localization of CRT, Fas-R and MHC class I in MDA-MB-231 cells.
Immunoreactivity of CRT and Fas-R was markedly increased in cells
treated with DOXO and DSG compared with control cells, with both
proteins predominantly localized at the cell surface membrane. By
contrast, no significant differences in MHC class I
immunoreactivity were observed among the experimental groups
(Fig. 6). Quantitative analysis of
FITC fluorescence intensity further revealed that DSG significantly
increased CRT and Fas-R expression compared with the control
(Fig. 6D), supporting the
ICD-inducing activity of DSG in MDA-MB-231 cells.
Furthermore, the expression levels of key mRNAs
involved in ICD-inducing mechanisms in MDA-MB-231 cells, including
those associated with ER stress (15), was evaluated after DSG treatment
using RT-qPCR. Compared with the control group, DSG-treated cells
exhibited significant upregulation of ER stress- and ICD-related
genes, including PERK, IRE1, ATF6, ATF4, eIF2α, CRT and Fas-R,
consistent with the effects observed in DOXO-treated cells. By
contrast, MHC class I mRNA expression remained unchanged following
DSG treatment (Fig. 7).
Collectively, these results indicated that DSG is associated with
activation ER stress-related ICD signaling pathways at the
transcriptional level, supporting its ICD-inducing potential in
MDA-MB-231 breast cancer cells.
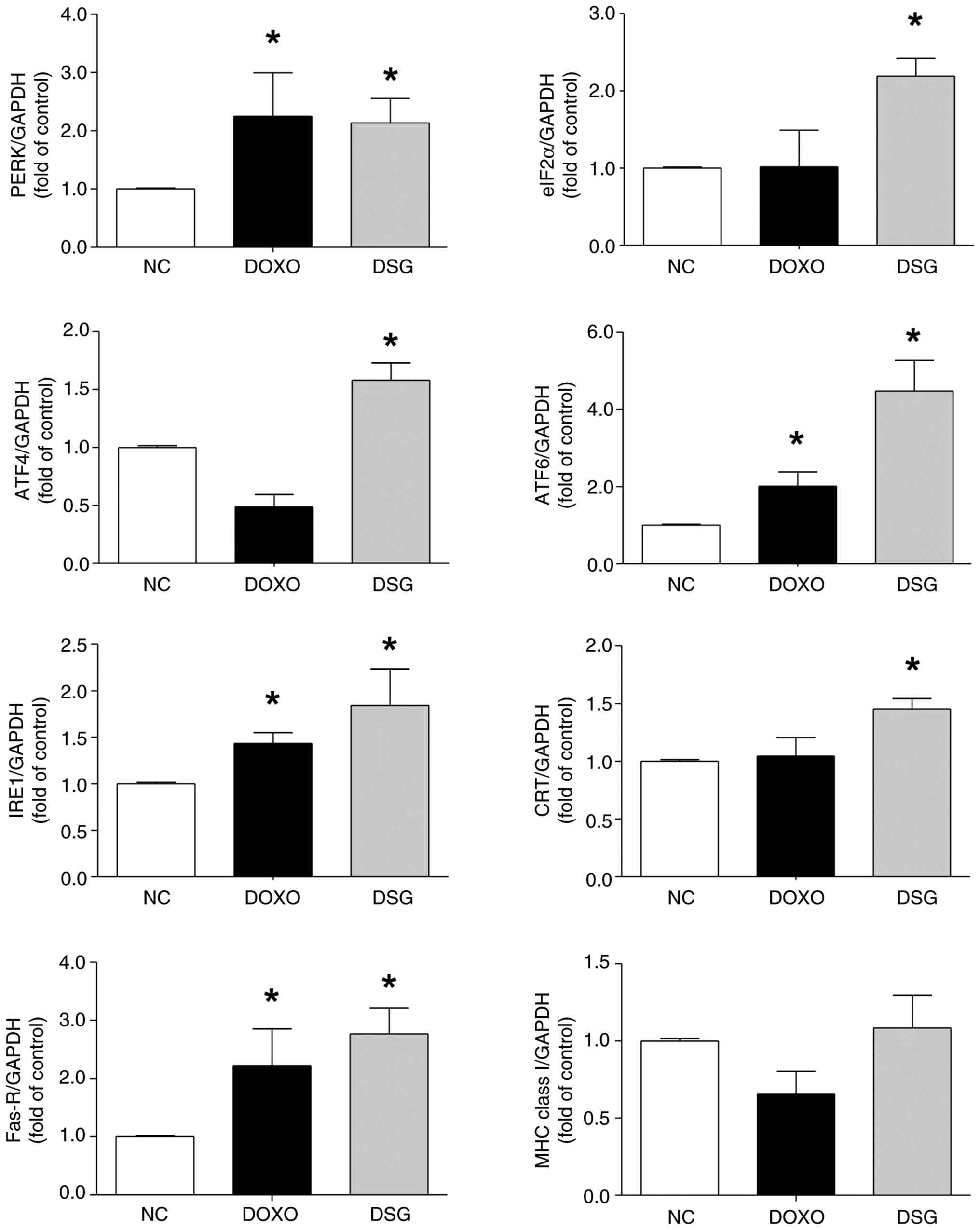 | Figure 7.mRNA expression levels of PERK,
eIF2α, ATF4, ATF6, IRE1, CRT, Fas-R and MHC class I in MDA-MB-231
cells treated with DOXO and DSG, as assessed by reverse
transcription quantitative PCR and normalized to GAPDH. Results are
presented as the mean±standard error of the mean (n=3) from three
independent experiments. *P<0.05 indicates a statistically
significant difference compared with the NC group. PERK, protein
kinase RNA-like endoplasmic reticulum kinase; ATF, activating
transcription factor; IRE1, inositol-requiring enzyme 1; eIF2α,
eukaryotic initiation factor 2 α subunit; DSG, degraded sulfated
galactan; NC, normal control; DOXO, doxorubicin; CRT, calreticulin;
Fas-R, Fas receptor; MHC, major histocompatibility complex. |
Discussion
There is growing interest in the anticancer
potential of sulfated polysaccharides, including their application
in cancer immunotherapy (18,27).
Their structural diversity, extensive biological activity and
notably low toxicity make sulfated polysaccharides key candidates
for anticancer development. Key structural features, such as
molecular weight, polysaccharide type and degree of substitution,
strongly influence their biological effects (28). In particular, reduced molecular
weight combined with increased sulfation has been reported to
enhance polysaccharide bioactivity (29). To generate low-molecular weight SG
from Gracilaria fisheri, acid hydrolysis has been used to
produce a homogeneous DSG (21).
In the present study, the surface morphology and elemental
composition of SG and DSG were characterized using SEM-EDX
analysis. To the best of our knowledge, the present study also
provides the first evidence that DSG induces ICD in triple-negative
breast cancer MDA-MB-231 cells. SEM is extensively used to examine
the morphology and microstructural features of polysaccharides
(30,31), while SEM-EDX allows simultaneous
assessment of microstructure and elemental composition (24,32).
The SEM-EDX analysis in the present study demonstrated clear
differences between SG and DSG in surface morphology, including
particle size and shape, as well as in elemental composition,
particularly sulfur content. Sulfate quantification and FTIR
spectroscopy further confirmed the presence of sulfur in both SG
and DSG, verifying their sulfated nature. Notably, degradation to a
lower molecular weight was associated with increased sulfate
content within the polysaccharide structure (33). These findings were consistent with
previous studies on seaweed-derived sulfated polysaccharides, which
reported that molecular weight and chemical composition strongly
influence surface morphology and physicochemical properties
(34,35).
The cytotoxic effects of SG and DSG on normal breast
epithelial MCF-10A cells and triple-negative breast cancer
MDA-MB-231 cells were evaluated and compared with those of DOXO, an
extensively used anticancer drug and a well-established inducer of
ICD (36,37). SG and DSG significantly reduced the
viability of MDA-MB-231 cells while having no detectable effect on
MCF-10A cell viability, whereas DOXO reduced viability in both cell
lines. These findings were consistent with the notable
morphological changes and increased membrane permeability observed
in MDA-MB-231 cells after treatment. Together, these results
indicated that SG and DSG exert selective cytotoxicity toward
cancer cells while sparing normal cells (38). This pattern aligns with previous
studies that demonstrated sulfated polysaccharides from Padina
tetrastromatica exhibit no cytotoxic effects on normal cells at
concentrations up to 2,000 µg/ml (39). Similarly, SG derivatives at
concentrations of 500 and 1,000 µg/ml were cytotoxic to MCF-7
breast cancer cells but not to L929 normal fibroblasts (9). The selective anticancer activity of
SG and DSG may reflect differences in cellular metabolism, surface
receptor expression and intracellular signaling pathways between
normal and cancer cells (40).
Cell morphological alterations and increased
membrane permeability are often associated with elevated ROS
generation induced by natural products used in cancer therapy
(41). For example, Artemisia
monosperma extracts exert cytotoxic effects on human colorectal
carcinoma HCT-116 cells by inducing ROS overproduction, leading to
notable morphological changes and membrane blebbing (42). Similarly, sulfated polysaccharides
derived from Padina tetrastromatica induce morphological
alterations in HeLa cells through ROS-mediated mitochondrial
membrane depolarization and dysfunction (43), consistent with the present study
observation that DSG increased ROS generation in MDA-MB-231 cells.
By contrast, SG did not elevate ROS levels in MDA-MB-231 cells,
suggesting that the observed membrane damage was not primarily
ROS-mediated. This effect may instead be associated with increased
membrane permeability resulting, at least in part, from disruption
of cytoskeletal proteins such as actin. Proteolytic degradation of
cytoskeletal components can compromise plasma membrane integrity
and lead to alterations in cell morphology (44). Furthermore, treatment with DSG,
characterized by lower-molecular weight and higher sulfate content,
exhibits enhanced biological activity compared with SG, indicating
a positive association between its structural features and
bioactivity (45). Molecular
weight and sulfate content are key determinants of the biological
activity of sulfated polysaccharides (29). Low-molecular weight sulfated
polysaccharides generally exhibit improved anticancer effects due
to enhanced solubility, tissue penetration and cellular uptake,
leading to greater inhibition of cancer cell proliferation,
induction of apoptosis and suppression of tumor progression
compared with the native form of sulfated polysaccharides (46). Additionally, higher sulfate content
has been associated with increased bioactivity, as evidenced by
greater inhibition of colony formation in moderately sulfated
fucoidans compared with low-sulfated forms (47). Consistent with these findings, DSG
likely exerts stronger anticancer activity by more effectively
interacting with cell surface molecules and/or enhancing its
penetration and uptake in MDA-MB-231 cells.
The impact of DSG treatment on the ultrastructure of
MDA-MB-231 cells was further examined using TEM analysis.
DSG-treated cells exhibited notable cytoplasmic alterations,
including prominent vacuolization, electron-dense inclusions and
marked organelle disorganization, closely resembling the
ultrastructural features observed in DOXO-treated cells. These
changes are indicative of cellular stress and cytotoxicity
(48). The present study findings
were consistent with previous studies that demonstrated
ultrastructural analysis of MDA-MB-231 cells exposed to
nanoencapsulated tarin revealed accumulation of autophagosomes and
damaged organelles (49).
Similarly, DOXO-treated human breast adenocarcinoma cells have been
reported to contain abundant abnormal vacuoles containing fibrous,
heterogeneous and flocculent material (50).
Notably, the presence of prominent cytoplasmic
vacuoles and electron-dense structures suggests autophagosome
formation and lysosomal involvement, features commonly associated
with ER stress-mediated ICD (15).
ER stress can trigger autophagic flux while promoting the exposure
and release of DAMPs, thereby enhancing tumor immunogenicity
(14,15). In the present study, DSG
upregulated CRT and Fas-R expression but did not alter MHC class I
levels compared with control cells. CRT exposure is a canonical
hallmark of ICD that facilitates phagocytic uptake of dying tumor
cells (14), whereas increased
Fas-R expression enhances tumor cell susceptibility to
immune-mediated apoptosis (16,51).
By contrast, MHC class I primarily governs antigen presentation to
CD8+ T cells and reflects adaptive immune recognition rather than
serving as a direct marker of ICD (17). The absence of changes in MHC class
I expression suggested that DSG preferentially activates
ICD-related danger signaling and immune susceptibility pathways
without extensively enhancing antigen presentation machinery.
Furthermore, low-molecular weight sulfated
polysaccharides exhibit improved cellular accessibility and
bioavailability, which facilitates robust activation of
ICD-associated ER stress signaling pathways, including
PERK-eIF2α-ATF4, IRE1 and ATF6 (15). Consistent with this mechanism, DSG
significantly upregulated the transcription of ER stress- and
ICD-related genes, including PERK, eIF2α, ATF4, ATF6, IRE1, CRT and
Fas-R, supporting its role as an ICD inducer. In line with these
findings, lentinan derived from shiitake mushrooms suppresses tumor
growth by inducing autophagy and apoptosis in HT-29 tumor models,
accompanied by marked ER stress activation (52). Sustained ER stress can ultimately
drive cell death through cytotoxic autophagy and disruption of Ca2+
homeostasis via pathways such as PERK/ATF4/CHOP and IRE1α (53). Beyond polysaccharides, the flavonol
glycoside afzelin induces ICD in lung cancer cells by activating ER
stress and increasing ATP, high mobility group box 1 and CRT
release (54). Similarly, extracts
of Marsdenia tenacissima inhibit non-small cell lung cancer
cell viability by concurrently activating ER stress (ATF6,
glucose-regulated protein 78 kDa, ATF4, X-box binding protein 1s
and CHOP) and ICD pathways (55).
Trametes robiniophila Murr. (Huaier) has also been reported
to promote ICD in triple-negative breast cancer cells by enhancing
CRT exposure, potentially through the circular RNA cytoplasmic
linker associated protein 1/double-stranded RNA-dependent protein
kinase/eIF2α signaling axis (56).
The present study demonstrated that DSG derived from
Gracilaria fisheri selectively exerts mild cytotoxic effects
on triple-negative breast cancer MDA-MB-231 cells while sparing
normal MCF-10A cells. Of note, DSG effectively triggers ICD, likely
through activation of ER stress-mediated pathways, highlighting a
mechanistic association between its structural features and
immunomodulatory antitumor activity. Mild to moderate cytotoxicity
induced by DSG may still serve a key role in promoting
ICD-associated signaling and immunomodulatory activity, rather than
diminishing its therapeutic relevance. ICD is not solely dependent
on extensive tumor cell killing but is primarily driven by the
induction of cellular stress responses that lead to the emission of
DAMPs (14,15). In this context, mild cytotoxic
stress caused by DSG may be sufficient to activate ER stress
pathways, particularly the PERK-eIF2α axis, which is closely
associated with CRT exposure on the cell surface, a key hallmark of
ICD (15). These signals
collectively enhance dendritic cell recruitment, antigen uptake and
subsequent T-cell-mediated antitumor immunity. Notably, sulfated
polysaccharides have also been reported to directly modulate immune
cell functions, including macrophage activation and cytokine
production, suggesting that their antitumor effects may arise from
a combined action of tumor cell stress induction and immune system
stimulation (57,58). Therefore, even in the presence of
only mild cytotoxicity, DSG may still effectively contribute to
anticancer immunity by functioning as an ICD-inducing
immunomodulatory agent that amplifies both tumor-derived danger
signals and host immune responses.
However, several limitations in the present study
should be noted. Definitive confirmation of DSG-induced ICD will
require additional functional assays, including co-culture of
DSG-treated cancer cells with immune cells, phagocytosis assays and
immune cell-mediated cytotoxicity assays. Furthermore, confirmatory
experiments, such as the use of ROS scavengers (for example,
N-acetylcysteine), PERK inhibition or gene knockdown, are warranted
to verify that DSG triggers ICD through activation of ER
stress-mediated pathways. Further studies are also warranted to
determine whether DSG induces ICD in other breast cancer subtypes
or additional cancer types and whether these effects can be
reproduced in vivo. Addressing these points will be key to
advancing DSG toward translational and clinical application in the
future.
Supplementary Material
Supporting Data
Acknowledgements
We would like to thank Dr Dylan Southard,
International Affairs, Khon Kaen University International College,
Thailand for language editing of the manuscript.
Funding
The present study received grants from the Khon Kaen University
Faculty of Medicine, Thailand (grant no. IN69033) and Mahasarakham
University Faculty of Medicine, Thailand (grant no. Med.Msu
01/09/2566).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
TR, YP, KW and CP conceived and designed the present
study. TR, YP, WS, JP and CP conducted the experiments. TR, WS, SS,
JEA and CP performed the data analysis. TR, WS, SS and CP drafted
the manuscript. TR, KW and CP revised and finalized the manuscript.
TR and CP obtained funding, managed the project and confirm the
authenticity of all the raw data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249.
2021.PubMed/NCBI
|
|
2
|
Łukasiewicz S, Czeczelewski M, Forma A,
Baj J, Sitarz R and Stanisławek A: Breast cancer-epidemiology, risk
factors, classification, prognostic markers, and current treatment
strategies-an updated review. Cancers (Basel). 13:42872021.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mun EJ, Babiker HM, Weinberg U, Kirson ED
and Von Hoff DD: Tumor treating fields: A fourth modality in cancer
treatment. Clin Cancer Res. 24:266–275. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vijayalakshmi M, Meganathan S, Surendhar
SK, Umamaheswari A and Prabu SL: Exploring the systematic
anticancer mechanism in selected medicinal plants: A review. Oncol
Adv. 2:141–147. 2024. View Article : Google Scholar
|
|
5
|
Armonavičius D, Maruška A, Jakštys B,
Stankevičius M, Drevinskas T, Bimbiraitė-Survilienė K, Čaplikaitė
M, Ihara H, Takafuji M, Skrzydlewska E, et al: Evaluation of the
anticancer activity of medicinal plants predominantly accumulating
ellagic acid compounds. Antioxidants (Basel). 14:13392025.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Guo R, Chen M, Ding Y, Yang P, Wang M,
Zhang H, He Y and Ma H: Polysaccharides as potential anti-tumor
biomacromolecules-A review. Front Nutr. 9:8381792022. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Habtemariam S: Trametes versicolor
(Synn. Coriolus versicolor) polysaccharides in cancer
therapy: Targets and efficacy. Biomedicines. 8:1352020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sae-Lao T, Tohtong R, Bates DO and
Wongprasert K: Sulfated galactans from red seaweed gracilaria
fisheri target EGFR and inhibit cholangiocarcinoma cell
proliferation. Am J Chin Med. 45:615–633. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Phanphak J, Somintara S, Sakaew W, Senarai
T, Kovensky J, Wongprasert K and Rudtanatip T: Sulfated galactan
derivatives from Gracilaria fisheri suppress the
proliferation of MCF7 breast cancer cells by inducing cell cycle
arrest. World Acad Sci J. 7:772025. View Article : Google Scholar
|
|
10
|
Van Allen EM, Miao D, Schilling B, Shukla
SA, Blank C, Zimmer L, Sucker A, Hillen U, Foppen MHG, Goldinger
SM, et al: Genomic correlates of response to CTLA-4 blockade in
metastatic melanoma. Science. 350:207–211. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schmid P, Cortes J, Pusztai L, McArthur H,
Kümmel S, Bergh J, Denkert C, Park YH, Hui R, Harbeck N, et al:
Pembrolizumab for early triple-negative breast Cancer. N Engl J
Med. 382:810–821. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Peng K, Zhao X, Fu YX and Liang Y:
Eliciting antitumor immunity via therapeutic cancer vaccines. Cell
Mol Immunol. 22:840–868. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fabian KP, Kowalczyk JT, Reynolds ST and
Hodge JW: Dying of stress: Chemotherapy, radiotherapy, and
small-molecule inhibitors in immunogenic cell death and immunogenic
modulation. Cells. 11:38262022. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Arimoto KI, Miyauchi S, Liu M and Zhang
DE: Emerging role of immunogenic cell death in cancer
immunotherapy. Front Immunol. 15:13902632024. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Han Y, Tian X, Zhai J and Zhang Z:
Clinical application of immunogenic cell death inducers in cancer
immunotherapy: Turning cold tumors hot. Front Cell Dev Biol.
12:13631212024. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Szarynska M, Olejniczak A, Wierzbicki P,
Kobiela J, Laski D, Sledzinski Z, Adrych K, Guzek M and Kmiec Z:
FasR and FasL in colorectal cancer. Int J Oncol. 51:975–986. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu K and Fong L: CD4+ T cells help
myeloid-mediated killing of immune-evasive tumors. Trends Cancer.
9:777–779. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pradhan B, Rout L and Ki JS:
Immunomodulatory and anti-inflammatory and anticancer activities of
porphyran, a sulfated galactan. Carbohydr Polym. 301:1203262023.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu W, Shen M, Li J and Xie J:
Structure-activity relationships of sulfated polysaccharides in
inflammatory bowel disease: Sources, mechanisms, and therapeutic
potential. Carbohydr Polym. 372:1245742026. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Khongthong S, Theapparat Y, Roekngam N,
Tantisuwanno C, Otto M and Piewngam P: Characterization and
immunomodulatory activity of sulfated galactan from the red seaweed
gracilaria fisheri. Int J Biol Macromol. 189:705–714. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rudtanatip T, Somintara S, Sakaew W,
ElAbid J, Cano ME, Jongsomchai K, Wongprasert K and Kovensky J:
Sulfated galactans from gracilaria fisheri with
supplementation of octanoyl promote wound healing activity in vitro
and in vivo. Macromol Biosci. 22:e22001722022. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wongprasert K, Rudtanatip T and Praiboon
J: Immunostimulatory activity of sulfated galactans isolated from
the red seaweed gracilaria fisheri and development of resistance
against white spot syndrome virus (WSSV) in shrimp. Fish Shellfish
Immunol. 36:52–60. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−delta delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Patel MA, Manna S, Vo A, Xu X, Conti DS,
Choi S, Kozak D and Zheng J: Scanning electron microscope (SEM)
coupled with energy dispersive X-ray spectroscopy (EDS)-a potential
analytical tool for physicochemical characterization of API in
complex drug formulations. Microsc Microanal. 26:2254–2255. 2020.
View Article : Google Scholar
|
|
25
|
Wang J, Sun D, Huang L, Wang S and Jin Y:
Targeting reactive oxygen species capacity of tumor cells with
repurposed drug as an anticancer therapy. Oxid Med Cell Longev.
2021:85329402021. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhao H, Zhao Y, Zhang S, Wang Z, Yu W,
Dong N, Yang X, Zhang X, Sun Q, Hao X and Ren X: Effects of
immunogenic cell death-inducing chemotherapeutics on the immune
cell activation and tertiary lymphoid structure formation in
melanoma. Front Immunol. 15:13027512024. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
B J and R, . R: A critical review on
pharmacological properties of sulfated polysaccharides from marine
macroalgae. Carbohydr Polym. 344:1224882024. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nagahawatta DP, Liyanage NM, Jayawardena
TU, Yang F, Jayawardena HHACK, Kurera MJMS, Wang F, Fu X and Jeon
YJ: Functions and values of sulfated polysaccharides from seaweed.
Algae. 38:217–240. 2023. View Article : Google Scholar
|
|
29
|
Kang J, Jia X, Wang N, Xiao M, Song S, Wu
S, Li Z, Wang S, Cui SW and Guo Q: Insights into the
structure-bioactivity relationships of marine sulfated
polysaccharides: A review. Food Hydrocoll. 123:1070492022.
View Article : Google Scholar
|
|
30
|
Yue JR, Lu JM, Fan QF, Sun P, Li YJ, Zhou
SL, Wang XY, Niu JM, Xu YK and Zhou J: Comparative study of the
structural characteristics and bioactivity of polysaccharides
extracted from aspidopterys obcordate hemsl. using different
solvents. Molecules. 28:79772023. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang M, Zhao P, Ochir S, Bai W, Lu J, Dan
M, Bo S and Chen C: Sulfated modification, characterizations,
anti-oxidant and immunostimulatory activities of a polysaccharide
from arnebia euchroma (royle) johnst. Roots. Results Chem.
17:1026242025. View Article : Google Scholar
|
|
32
|
Olasehinde TA, Mabinya LV, Olaniran AO and
Okoh AI: Chemical characterization of sulfated polysaccharides from
gracilaria gracilis and ulva lactuca and their radical scavenging,
metal chelating, and cholinesterase inhibitory activities. Int J
Food Prop. 22:100–110. 2019. View Article : Google Scholar
|
|
33
|
Gong P, Wang M, Guo Y, Long H, Wang Z, Cui
D, Yao W, Yang W, Chen F and Xie J: Structure characterization, in
vitro antioxidant and anti-tumor activity of sulfated
polysaccharide from siraitia grosvenorii. Foods.
12:21332023. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tang L, Chen Y, Jiang Z, Zhong S, Chen W,
Zheng F and Shi G: Purification, partial characterization and
bioactivity of sulfated polysaccharides from grateloupia lívida.
Int J Biol Macromol. 94:642–652. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Netanel Liberman G, Ochbaum G,
Mejubovsky-Mikhelis M, Bitton R and Malis Arad S: Physicochemical
characteristics of the sulfated polysaccharides of the red
microalgae dixoniella grisea and porphyridium aerugineum. Int J
Biol Macromol. 145:1171–1179. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kciuk M, Gielecińska A, Mujwar S, Kołat D,
Kałuzińska-Kołat Ż, Celik I and Kontek R: Doxorubicin-An agent with
multiple mechanisms of anticancer activity. Cells. 12:6592023.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cardador CM, de Castro TB, de Castro RJA,
Bocca AL, Camargo LC, Pacheco TA, Muehlmann LA and Longo JPF:
Doxorubicin-induced immunogenic cell death impairs tumor
progression and distant metastasis in a 4T1 breast cancer tumor
model. Curr Pharm Des. 30:2493–2504. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Blagosklonny MV: Selective protection of
normal cells from chemotherapy, while killing drug-resistant cancer
cells. Oncotarget. 14:193–206. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jose GM, Raghavankutty M and Kurup GM:
Attenuation of hydrogenperoxide-induced oxidative damages in L929
fibroblast cells by sulfated polysaccharides isolated from the
edible marine algae padina tetrastromatica. J Bioact Compat
Polym. 34:150–162. 2019. View Article : Google Scholar
|
|
40
|
Akhtar MJ, Ahamed M, Alhadlaq HA,
Alrokayan SA and Kumar S: Targeted anticancer therapy:
Overexpressed receptors and nanotechnology. Clin Chim Acta.
436:78–92. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Muchtaridi M, Az-Zahra F, Wongso H,
Setyawati LU, Novitasari D and Ikram EHK: Molecular mechanism of
natural food antioxidants to regulate ROS in treating cancer: A
review. Antioxidants (Basel). 13:2072024. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Farshori NN, Al-Sheddi ES, Al-Oqail MM,
Al-Massarani SM, Al-Jassas EA, Ahmad J, Saquib Q, Wahab R,
Al-Khedhairy AA and Siddiqui MA: Artemisia monosperma
induces ROS-mediated cell death in human colorectal carcinoma cells
via modulating apoptotic genes. J King Saud Univ Sci.
35:1027632023. View Article : Google Scholar
|
|
43
|
Jose GM, Raghavankutty M and Kurup GM:
Sulfated polysaccharides from Padina tetrastromatica induce
apoptosis in HeLa cells through ROS triggered mitochondrial
pathway. Process Biochem. 68:197–204. 2018. View Article : Google Scholar
|
|
44
|
Nakajima K, Takakura H, Shimizu Y and
Ogawa M: Changes in plasma membrane damage inducing cell death
after treatment with near-infrared photoimmunotherapy. Cancer Sci.
109:2889–2896. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wang Y, Xing M, Cao Q, Ji A, Liang H and
Song S: Biological activities of fucoidan and the factors mediating
its therapeutic effects: A review of recent studies. Mar Drugs.
17:1832019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liyanage NM, Dissanayake DS, Li Y, Ko KY,
Nagahawatta DP and Jeon YJ: Sulfated polysaccharides in cancer
therapy: A focus on algal-derived bioactive. Mar Drugs. 24:1312026.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Trung DT, Surits VV, Zueva AO, Cao HTT,
Shevchenko NM, Ermakova SP and Thinh PD: Anticancer activity in
vitro of sulfated polysaccharides from the brown alga
spatoglossum vietnamense. Molecules. 29:49822024. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Perrotta I: Live and let die: Analyzing
ultrastructural features in cell death. Ultrastruct Pathol.
49:1–19. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cardoso RV, Pereira PR, Freitas CS, Mattos
ÉBA, Silva AVF, Midlej VDV, Vericimo MA, Conte-Júnior CA and
Paschoalin VMF: Tarin-loaded nanoliposomes activate apoptosis and
autophagy and inhibit the migration of human mammary adenocarcinoma
cells. Int J Nanomed. 18:6393–6408. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Rembiałkowska N, Dubińska-Magiera M,
Sikora A, Szlasa W, Szewczyk A, Czapor-Irzabek H, Daczewska M,
Saczko J and Kulbacka J: Doxorubicin assisted by microsecond
electroporation promotes irreparable morphological alternations in
sensitive and resistant human breast adenocarcinoma cells. Appl
Sci. 10:27652020. View Article : Google Scholar
|
|
51
|
Sica M, Roussel M and Legembre P: CD95/Fas
stoichiometry in future precision medicine. Cell Death Differ.
32:1570–1577. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhang Y, Liu Y, Zhou Y, Zheng Z, Tang W,
Song M, Wang J and Wang K: Lentinan inhibited colon cancer growth
by inducing endoplasmic reticulum stress-mediated autophagic cell
death and apoptosis. Carbohydr Polym. 267:1181542021. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Fan J, Zhu J, Zhu H, Zhang Y and Xu H:
Potential therapeutic target for polysaccharide inhibition of colon
cancer progression. Front Med (Lausanne). 10:13254912024.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Xia L, Xu X, Li M, Zhang X and Cao F:
Afzelin induces immunogenic cell death against lung cancer by
targeting NQO2. BMC Complement Med Ther. 23:3812023. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Yuan Y, Guo Y, Guo ZW, Hao HF, Jiao YN,
Deng XX and Han SY: Marsdenia tenacissima extract induces
endoplasmic reticulum stress-associated immunogenic cell death in
non-small cell lung cancer cells through targeting AXL. J
Ethnopharmacol. 314:1166202023. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Li C, Wang X, Chen T, Li W, Zhou X, Wang L
and Yang Q: Huaier induces immunogenic cell death via
circCLASP1/PKR/eIF2α signaling pathway in triple negative breast
cancer. Front Cell Dev Biol. 10:9138242022. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Bhuyan PP, Nayak R, Patra S, Abdulabbas
HS, Jena M and Pradhan B: Seaweed-derived sulfated polysaccharides;
The new age chemopreventives: A comprehensive review. Cancers
(Basel). 15:7152023. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Wang Y, Luo J, Xu C, Hu D, Li Y, Ye Y,
Yang J, Chen X, Li C and Zhu K: Recent advance in marine
polysaccharides: Structure, anti-inflammatory mechanisms, and
functional applications. Mar Drugs. 24:1292026. View Article : Google Scholar : PubMed/NCBI
|