Introduction
Cardiovascular diseases (CVDs) are a leading cause
of morbidity and mortality worldwide, accounting for an estimated
19.7 million deaths annually, representing ~32% of all global
deaths (1). Among the risk factors
for CVDs, oxidized low-density lipoprotein (ox-LDL) is a key driver
of atherosclerosis, a disease characterized by the accumulation of
fatty deposits in arterial walls (2,3).
Previous studies have highlighted the detrimental effects of ox-LDL
on macrophage function, especially its ability to promote
inflammation and cell senescence (4,5).
Moreover, the fact that ox-LDL contributes to atherosclerosis by
inducing oxidative stress and inflammatory responses is
well-established (6,7).
Autophagy is a key cellular process that degrades
and recycles damaged organelles and proteins, thereby maintaining
cell homeostasis (8). In
macrophages, autophagy is particularly important because it
regulates inflammation and lipid metabolism (9,10).
Impaired autophagic activity is associated with pathophysiological
conditions, including CVD and neurodegeneration (11). Macrophage autophagy promotes the
catabolism of cytoplasmic lipid droplets, maintains cell lipid
homeostasis and plays an important role in anti-atherosclerotic
processes (12). Moreover,
mitochondrial dysfunction, typically a consequence of oxidative
stress, exacerbates these conditions (13). The interaction between autophagy
and mitochondrial function is complex and understanding this
interplay is essential for developing effective therapeutic
interventions.
β-hydroxybutyrate (BHB), a ketone body generated
during fatty acid metabolism, possesses anti-inflammatory
properties and may protect cells from oxidative stress-induced
damage (14). BHB also promotes
autophagosome formation, suggesting that it may be used
therapeutically in metabolic disorders (15). The STAT signaling pathway is a
critical component of cellular communication, primarily involved in
regulating the transcription of target genes that are essential for
cellular function (16). The
clinical significance of the relationship between STAT signaling
and autophagy is becoming increasingly evident (17,18).
In cancer, for example, the activation of the JAK/STAT pathway
leads to enhanced autophagic activity, which may contribute to
tumor cell survival and resistance to therapy (19). Conversely, inhibiting the JAK/STAT
pathway is associated with the promotion of autophagy and tumor
cell death, indicating that targeting this signaling pathway could
be a viable strategy for cancer treatment (20). The activation of the IL-12/STAT4
signaling pathway enhances autophagic activity, thereby
contributing to the clearance of Mycobacterium tuberculosis
in infected macrophages (21).
Autophagy-related protein 5 can modulate immune responses by
enhancing the production of CCL2, which influences STAT4 activation
in CD4+ T helper 1 cells during malaria infections
(22). When macrophages are
exposed to ox-LDL, STAT4 is phosphorylated and translocates to the
nucleus, where it regulates the transcription of genes involved in
inflammation (23). This leads to
the production of pro-inflammatory cytokines such as IFN-γ and
TNF-α, which are essential for the activation and recruitment of
immune cells to the site of inflammation (24).
The molecular mechanisms by which BHB regulates
autophagy and the role of STAT4 in this process remain incompletely
understood. Further investigations are needed to delineate these
pathways and assess their therapeutic relevance for
autophagy-targeted interventions in lipid-related diseases. The
present study aimed to examine the effects of BHB on autophagy and
mitochondrial function in THP-1 macrophage exposed to ox-LDL and
determine how BHB may restore cellular functions that are impaired
by ox-LDL, with particular emphasis on the signaling molecule
STAT4. The present findings could inform the development of novel
therapeutic strategies for metabolic disorders and CVD.
Materials and methods
Reagents
(R)-(−)-β-Hydroxybutyric acid sodium salt (cat. no.
298360) was purchased from Sigma-Aldrich (Merck KGaA). ox-LDL (cat.
no. YB-002) and
1,1′-dioctadecyl--3,3,3′,3′-tetramethylindocarbocyanine perchlorate
(Dil)-ox-LDL (cat. no. YB-0010) were obtained from Guangzhou Yiyuan
Biotech Co., Ltd. Phorbol 12-myristate 13-acetate (PMA; cat. no.
S1819) and the JC-1 Mitochondrial Membrane Potential (ΔΨm) Assay
kit (cat. no. C2003S) were obtained from Beyotime Biotechnology.
FBS (cat. no. 11011-8611) was purchased from Beijing Solarbio
Science & Technology Co., Ltd. The Senescence-Associated
β-Galactosidase (SA-β-Gal) Stain Kit (cat. no. G1580), BSA (cat.
no. IA0910), Autophagy Staining Detection kit (cat. no. G0170),
DAPI Staining Solution (cat. no. C0065), High-Efficiency RIPA
Cell/Tissue Lysis Buffer (cat. no. R0010) and trypsin (cat. no.
T1300) were obtained from Beijing Solarbio Science & Technology
Co., Ltd. Rabbit anti-rat p62 (cat. no. ab109012), anti-STAT4 (cat.
no. ab28815) and anti-β-actin antibody (cat. no. ab8226) were
obtained from Abcam. ECL Plus Ultra-Sensitive Detection Reagent
(cat. no. P0018AM), PVDF membrane (cat. no. FFP28), HRP-labeled
goat anti-mouse IgG (cat. no. A0216) and HRP-labeled goat
anti-rabbit IgG (cat. no. A0208) were obtained from Beyotime
Biotechnology. The PAGE Gel Rapid Preparation kit (cat. no. PG112)
was obtained from Epizyme Biotech. High-glucose DMEM (cat. no.
SH30022.01) was purchased from Cytiva. Lipofectamine 2000 (cat. no.
11668030) and Opti-MEM reduced-serum medium (cat. no. 31985070)
were purchased from Thermo Fisher Scientific, Inc.
Cell culture and treatment
THP-1 cells were purchased from GenChem Inc. and
maintained in high-glucose DMEM supplemented with 10% FBS and 1%
penicillin/streptomycin at 37°C in a humidified atmosphere
containing 5% CO2. All experiments were performed using
cells between passages 3 and 10 at ~80% confluency.
For macrophage differentiation, 2×105
THP-1 cells per well were seeded in 6-well plates and stimulated
with 100 ng/ml PMA at 37°C for 24 h. Following PMA-induced
monocyte-to-macrophage differentiation (confirmed by the transition
to an adherent morphology), cells were washed twice with PBS. In
the experiments involving both ox-LDL and BHB, the differentiated
THP-1 macrophages were first incubated with 50 mg/l ox-LDL for 24 h
at 37°C, and then incubated with BHB (1, 2 or 3 mM) for 24 h at
37°C.
Western blot analysis
A total of 2×105 THP-1 cells per well
seeded in 6-well plates were stimulated with 100 ng/ml PMA for 24 h
at 37°C. The differentiated THP-1 macrophages were treated with
either ox-LDL or BHB for an additional 24 h at 37°C. Cells were
harvested and washed twice with ice-cold PBS. Protein extraction
was performed using 100 µl high-efficiency RIPA lysis buffer
supplemented with 1 µl PMSF by incubating on ice for 30 min.
Lysates were centrifuged at 12,000 × g for 20 min at 4°C, mixed
with 25 µl 5X loading buffer, denatured at 100°C for 10 min and
immediately chilled on ice. Samples were stored at −20°C until
analysis.
Protein concentration was determined using the BCA
assay. Equal amounts of protein (20 µg/lane) were separated on 10%
SDS-polyacrylamide gels and transferred onto PVDF membranes at 100
V. Membranes were blocked with 5% BSA for 2 h at room temperature,
followed by incubation with primary antibodies (1:1,000) overnight
at 4°C. HRP-conjugated goat anti-rabbit secondary antibody
(1:5,000) or HRP-conjugated goat anti-mouse secondary antibody
(1:5,000) was applied for 2 h at room temperature. Protein signals
were detected using ECL reagents. Band intensities were quantified
using ImageJ software (ImageJ 1.53t; National Institutes of Health)
with β-actin as the loading control.
Autophagy detection
A total of 4×104 THP-1 cells per well
were seeded onto sterile glass coverslips placed in 24-well plates.
Following stimulation with 100 ng/ml PMA at 37°C for 24 h, cells
were washed twice with PBS. THP-1 cells in different experimental
groups were treated as follows: In the experiment to determine the
optimal concentration of ox-LDL, differentiated THP-1 macrophages
were incubated with ox-LDL (0, 50 and 100 mg/l) at 37°C for 24 h;
in experiments involving ox-LDL and BHB, differentiated THP-1
macrophages were first incubated with 50 mg/l ox-LDL at 37°C for 24
h, and then incubated with BHB (1, 2 or 3 mM) at 37°C for 24 h; and
in experiments involving overexpression and inhibition of STAT4,
after the overexpression and inhibition of STAT4, 50 mg/l ox-LDL
was added (or not added) and cells were incubated at 37°C for 24 h,
followed by the addition (or non-addition) of 3 mM BHB and
incubation at 37°C for 24 h.
After the aforementioned treatment, the cells were
processed with an Autophagy Staining Detection kit, according to
the manufacturer's protocol. Briefly, cells were gently rinsed
twice with wash buffer, then incubated with dansylcadaverine for
15–30 min at room temperature under light-protected conditions.
Following two additional washes, nuclei were counterstained with
DAPI (1 µg/ml) for 5–10 min at 25°C. Following final rinsing with
wash buffer, the coverslips were mounted onto adhesive microscope
slides using an anti-fade mounting medium (cat. no. S2100; Beijing
Solarbio Science & Technology Co., Ltd.).
Autophagic vesicles were visualized with a
laser-scanning confocal microscope (FV3000; Olympus Corporation).
Excitation/emission settings were set to optimal dye parameters
(488 nm excitation/530 nm emission) and for DAPI (405 nm
excitation/461 nm emission). Images were acquired using sequential
channel scanning to avoid fluorescence bleed-through.
ΔΨm assessment
A total of 4×104 THP-1 cells per well
grown on glass coverslips in 24-well plates. Following stimulation
with 100 ng/ml PMA at 37°C for 24 h, cells were washed twice with
PBS. Cell treatments and grouping were the same as described in the
autophagy detection section.
After the aforementioned treatment, the cells were
processed with the JC-1 Mitochondrial Membrane Potential (ΔΨm)
Assay kit (cat. no. C2003S; Beyotime Biotechnology) according to
the manufacturer's protocol. Briefly, a JC-1 dye working solution
was prepared by diluting the stock solution 1:1 in complete culture
medium (high-glucose DMEM supplemented with 10% FBS and 1%
penicillin/streptomycin). The cells were incubated with this
mixture for 20 min at 37°C under light-protected conditions. After
incubation, the coverslips were washed twice with JC-1 assay
buffer, transferred to fresh culture medium and mounted face-down
onto adhesive microscope slides using an anti-fade mounting medium.
The ΔΨm assessment was performed by observing the aggregated (red)
or monomeric (green) state of JC-1 in the cells with a
laser-scanning confocal microscope (FV3000; Olympus Corporation).
Fluorescence intensities were quantified using ImageJ software
(ImageJ 1.53t; National Institutes of Health).
Senescence-associated β-Gal (SA-β-Gal)
staining
A total of 4×104 THP-1 cells per well
were seeded in 24-well plates. Following stimulation with 100 ng/ml
PMA at 37°C for 24 h, cells were washed twice with PBS.
Differentiated THP-1 macrophages were first incubated with 50 mg/l
ox-LDL at 37°C for 24 h, and then incubated with BHB (0,1, 2 or 3
mM) at 37°C for 24 h. After the aforementioned treatment, the cells
were washed twice with PBS and fixed with 4% formaldehyde for 15
min at room temperature. Following three PBS washes, the cells were
incubated with 1 ml β-Gal staining solution [1 mg/ml X-Gal, 5 mM
potassium ferrocyanide, 5 mM potassium ferricyanide, 150 mM NaCl, 2
mM MgCl2 in 40 mM citric-acid/sodium-phosphate buffer
(pH 6.0)] under an airtight seal with paraffin film at 37°C in a
non-CO2 atmosphere for 24 h. After two additional PBS
washes, SA-β-Gal-positive cells (blue staining) were examined under
a fluorescence microscope. Semi-quantitative analysis of
fluorescence intensity was performed using ImageJ software (version
1.54f; National Institutes of Health). In total, five random fields
of view/well were imaged for statistical analysis.
Fluorescent ox-LDL uptake assay
A total of 4×104 THP-1 cells per well
were seeded in 24-well plates and differentiated with 100 ng/ml PMA
for 24 h at 37°C. The resulting macrophages were incubated with 100
µg/ml DiI-ox-LDL together with BHB (0, 1, 2, 3 mM) for 24 h at
37°C, 5 % CO2. After incubation, the cells were washed
three times with PBS. Fluorescent micrographs were captured with an
inverted fluorescence microscope. Intracellular DiI-ox-LDL
accumulation was quantified with ImageJ software (version no.
1.53t, National Institutes of Health), by analyzing fluorescence
intensity in three independent biological replicates.
Total RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was isolated from THP-1 cells using an RNA
isolation kit (cat. no. R0026; Beyotime Biotechnology) according to
the manufacturer's instructions. Briefly, after adding the lysis
buffer to the THP-1 cell culture plate, the cells were lysed and
incubated at room temperature for 5 min. Subsequently, 200 µl
chloroform was added, and the mixture was vigorously shaken for 15
sec, followed by incubation at room temperature for 3 min. The
sample was then centrifuged at 13,000 × g for 10 min at 4°C. The
supernatant was transferred to a new tube. An equal volume of
ethanol was slowly added, followed by thorough mixing. The solution
along with any precipitate was transferred into an adsorption
column (CR3) and centrifuged at 13,000 × g for 30 sec at 4°C.
Subsequently, 500 µl protein removal solution was added, followed
by centrifugation at 13,000 × g for 30 sec at 4°C, and the
flow-through was discarded. The CR3 column was placed into a
collection tube, and 500 µl wash buffer was added. After incubation
at room temperature for 2 min, the column was centrifuged at 13,000
× g for 30 sec at 4°C, and the flow-through was discarded. The
column was transferred to a 2 ml collection tube and centrifuged at
13,000 × g for 2 min at 4°C to remove residual liquid. Finally, the
CR3 column was placed into a new 1.5-ml microcentrifuge tube, and
40 µl RNase-free ddH2O was added. After incubation at
room temperature for 2 min, the tube was centrifuged at 13,000 × g
for 2 min at 4°C. The extracted RNA was used for subsequent
experiments.
Reverse transcription was performed using an RT-PCR
kit (cat. no. RP1100; Beijing Solarbio Science & Technology
Co., Ltd.). Briefly, in a total reaction volume of 14.5 µl, 2 µg of
RNA and 2 µl of the specific primer Oligo(dT)16 were
added, and the volume was adjusted with RNase-free
ddH2O. The mixture was incubated at 70°C for 5 min and
then rapidly cooled on ice for 2 min. After a brief centrifugation
at 500 × g for 30 sec at 4°C to collect the reaction mixture, the
following components were added: 4 µl 5X M-MLV Buffer, 1 µl dNTPs,
0.5 µl RNasin and 1 µl M-MLV reverse transcriptase. The reaction
was incubated at 42°C for 60 min, followed by heating at 95°C for 5
min to terminate the reaction. The product was placed on ice for
subsequent experiments.
qPCR was performed using 2X SYBRGreen PCR Mastermix
(cat. no. SR1110; Beijing Solarbio Science & Technology Co.,
Ltd.). Briefly, the following components were added to a reaction
tube: 12.5 µl of 2X SYBR Green PCR mix, 1 µl of primers (10 µM), 5
µl of template DNA and 5.5 µl of ddH2O, to a final
volume of 25 µl. The reaction was performed using an Applied
Biosystems StepOnePlus real-time PCR system (Thermo Fisher
Scientific, Inc.) under the following conditions: Initial
denaturation at 95°C for 3 min; 40 cycles of denaturation at 95°C
for 20 sec, annealing at 60°C for 25 sec and extension at 72°C for
30 sec (with fluorescence acquisition during the extension step).
The cycle threshold (Ct) value was analyzed using StepOne software
v2.3 (Thermo Fisher Scientific, Inc.), semilog amplification curves
were evaluated using the comparative quantification method
(2−ΔΔCq) (25) and the
gene expression levels were normalized to those of human β-actin.
The primer pairs for qPCR were: STAT4 forward,
5′-CCTGACATTCCCAAAGACAAAGC-3′ and reverse,
5′-TCTCTCAACACCGCATACACAC-3′; and β-actin forward,
5′-GTGGACATCCGCAAAGAC-3′ and reverse,
5′-AAAGGGTGTAACGCAACTA-3′.
STAT4 overexpression via
Lipofectamine-mediated transfection
The PCDNA3.1-STAT4 (Homo sapiens,
XM_054343566.1) expression plasmid was purchased from BGI Genomics
and the empty PCDNA3.1 vector (BGI Genomics) was also transfected
as a negative control. A total of 2×105 THP-1 cells per
well were seeded in 6-well plates at ~80 % confluency and
differentiated with 100 ng/ml PMA for 24 h at 37°C. The medium was
replaced with antibiotic-free complete DMEM (Cytiva). Lipofectamine
2000 (10 µl) was diluted in 250 µl Opti-MEM® I
Reduced-Serum Medium. Plasmid DNA (5 µg) was diluted in 250 µl
Opti-MEM. Following a 5 min incubation at room temperature, the
diluted DNA and Lipofectamine solutions were combined, vortexed
gently and incubated for 15 min at room temperature. The resulting
DNA-lipid complexes were added dropwise to the cells. Following 6 h
of incubation at 37°C with 5 % CO2, the transfection
medium was replaced with fresh complete DMEM (Cytiva). The
expression of STAT4 was validated by RT-qPCR. RT-qPCR was used to
verify the overexpression efficiency of STAT4 before treating cells
with ox-LDL or BHB. THP-1 cells were seeded into 6-well plates at a
density of 2×105 cells per well and stimulated with 100
ng/ml PMA at 37°C for 24 h. Following transfection according to the
aforementioned procedure, the cells were collected for total RNA
extraction. Subsequently, RT-qPCR was conducted to detect STAT4
mRNA levels in order to confirm the transfection efficiency.
STAT4 silencing
The STAT4 gene-silencing kit (genOFF h-STAT4_2500A)
was used according to the manufacturer's protocol (cat. no.
SIGS0000957-4; Guangzhou RiboBio Co., Ltd.). The STAT4 siRNA target
sequence was 5′-GCCTGACCATAGATTTGGA-3′ (the corresponding siRNA was
provided by Guangzhou RiboBio Co., Ltd.), and the negative control
siRNA (cat. no. SIGS0000957-4; Guangzhou RiboBio Co., Ltd.) was
provided by Guangzhou RiboBio Co., Ltd. A total of 2×105
THP-1 cells per well were seeded in 6-well plates. Following 24 h
of stimulation with 100 ng/ml PMA at 37°C, the medium was replaced
with antibiotic-free complete DMEM. The Ribo FECT™ CP buffer was
diluted with PBS and the STAT4 siRNA and the negative control siRNA
included in the STAT4 gene-silencing kit were briefly centrifuged
(500 × g; 1 min) at 25°C, resuspended in 250 µl sterile de-ionized
water, aliquoted and stored at −20°C. For the transfection mixture,
120 µl diluted Ribo FECT CP buffer, 10 µl 50 nM STAT4 siRNA and 12
µl Ribo FECT CP reagent were incubated at room temperature for 15
min and then added dropwise to the 6-well plates. Following 48 h of
incubation at 37°C with 5% CO2, the expression of STAT4
was validated by RT-qPCR. RT-qPCR was used to verify the silencing
efficiency of STAT4 before treating cells with ox-LDL or BHB. THP-1
cells were seeded into 6-well plates at a density of
2×105 cells per well and stimulated with 100 ng/ml PMA
at 37°C for 24 h. Following transfection according to the
aforementioned procedure, the cells were collected for total RNA
extraction. Subsequently, RT-qPCR was conducted as aforementioned
to detect STAT4 mRNA levels in order to confirm the transfection
efficiency.
Statistical analysis
Data were analyzed using GraphPad Prism®
software (version 10.1.2; GraphPad Software, Inc.; Dotmatics). All
experiments were independently repeated at least three times. All
data are presented as the mean ± SEM. Statistical analysis was
performed using one-way ANOVA followed by Bonferroni's post hoc
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
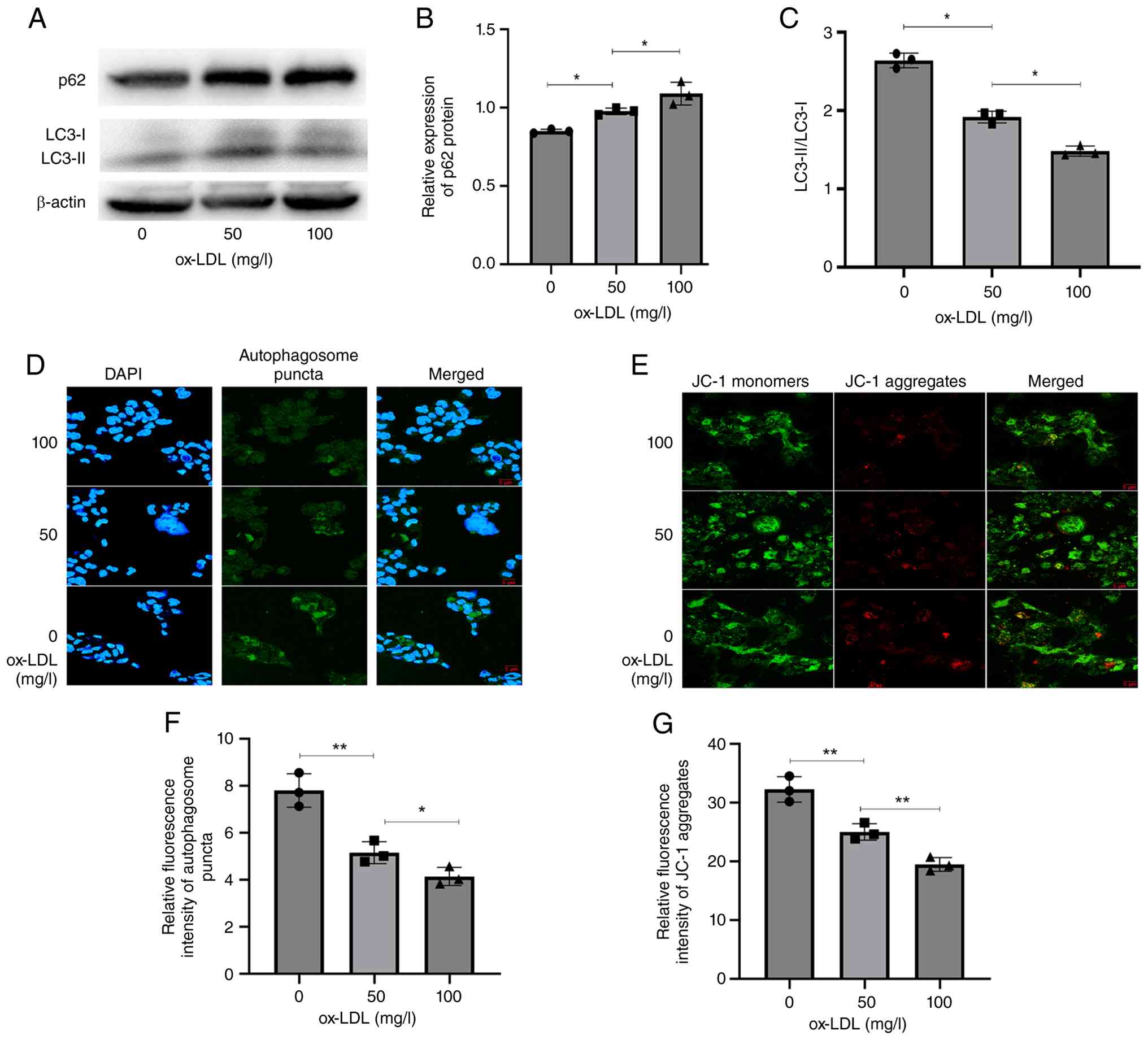
ox-LDL induces impaired autophagic
flux and decreased ΔΨm in THP-1 cells
The present study assessed the expression of p62, a
selective substrate for autophagic degradation, in THP-1 cells
following ox-LDL stimulation. Immunoblot analysis revealed that
ox-LDL (50 and 100 mg/l) significantly increased p62 expression and
decreased the ratio of LC3-II/LC3-I compared with untreated
controls (Fig. 1A-C). To
corroborate these findings, autophagosome formation was evaluated
using confocal laser scanning microscopy. Cells treated with ox-LDL
(100 mg/l) exhibited a marked reduction in autophagic puncta,
consistent with the immunoblot data, indicating impaired autophagy
(Fig. 1D and F). ΔΨm was measured
using the JC-1 fluorescent probe. Red fluorescence, indicating high
ΔΨm, was observed in control cells. By contrast, ox-LDL treatment
(100 mg/l) led to significantly attenuated red fluorescence
intensity and a concomitant increase in green fluorescence
(indicating a reduction in ΔΨm; Fig.
1E and G). These results demonstrate that ox-LDL exposure
induces a substantial decrease in ΔΨm in THP-1 cells. Collectively,
these findings indicated mitochondrial dysfunction, reflecting a
diminished capacity to maintain cell energy metabolism and
homeostasis.
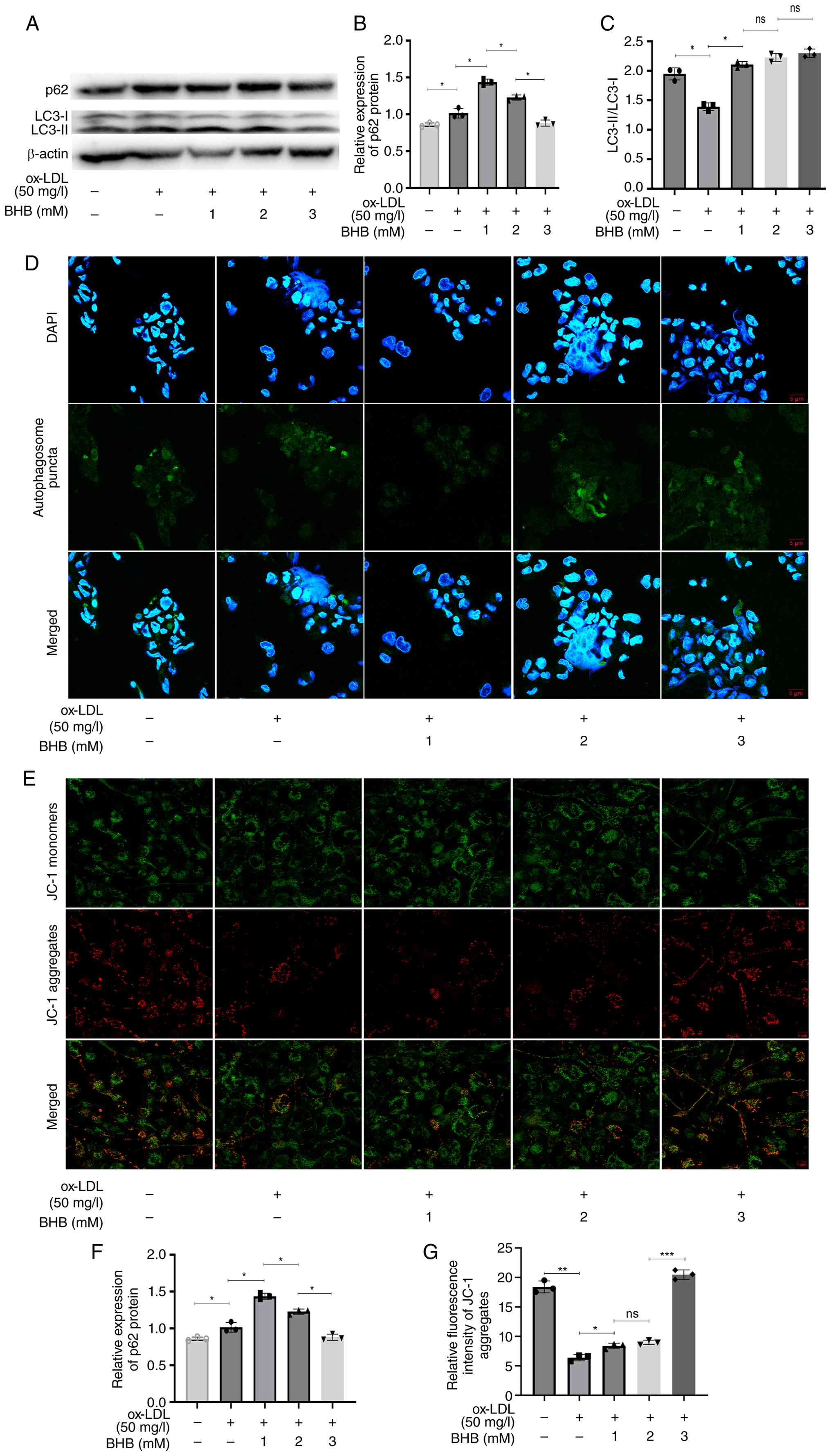
BHB restores impaired autophagic flux
and ΔΨm in ox-LDL-treated THP-1 cells
The present study assessed the effect of BHB on
autophagic flux in THP-1 cells. p62 expression levels decreased
significantly, while ratio of LC3-II/LC3-I increased markedly
following BHB treatment, indicating that BHB effectively restored
the impaired autophagic flux in THP-1 cells (Fig. 2A-C). Cells treated with both ox-LDL
and BHB at 1 and 2 mM exhibited a higher expression of p62 compared
with those exposed solely to ox-LDL, while a notable decrease in
expression was observed at 3 mM BHB. Fluorescent staining confirmed
this restoration: While 1 mM and 2 mM BHB reduced autophagic puncta
formation and ΔΨm, 3 mM BHB markedly increased both autophagic
puncta (Fig. 2D and F) and ΔΨm
(Fig. 2E and G). Collectively,
these results demonstrate that BHB enhances autophagic capacity and
mitochondrial function in THP-1 macrophages by ameliorating
ox-LDL-induced impairment of autophagic flux.
BHB decrease ox-LDL accumulation and
β-Gal activity in THP-1 cells
BHB co-treatment significantly decreased Dil-ox-LDL
levels in THP-1 cells, indicating potent inhibition of ox-LDL
accumulation in macrophages (Fig. 3A
and B). As elevated β-Gal activity serves as a key marker of
cell senescence (26), this was
measured following 24 h of co-treatment with ox-LDL and BHB. BHB
suppressed intracellular β-Gal activity in a dose-dependent manner
(Fig. 3C and D), suggesting
attenuation of cell aging processes.
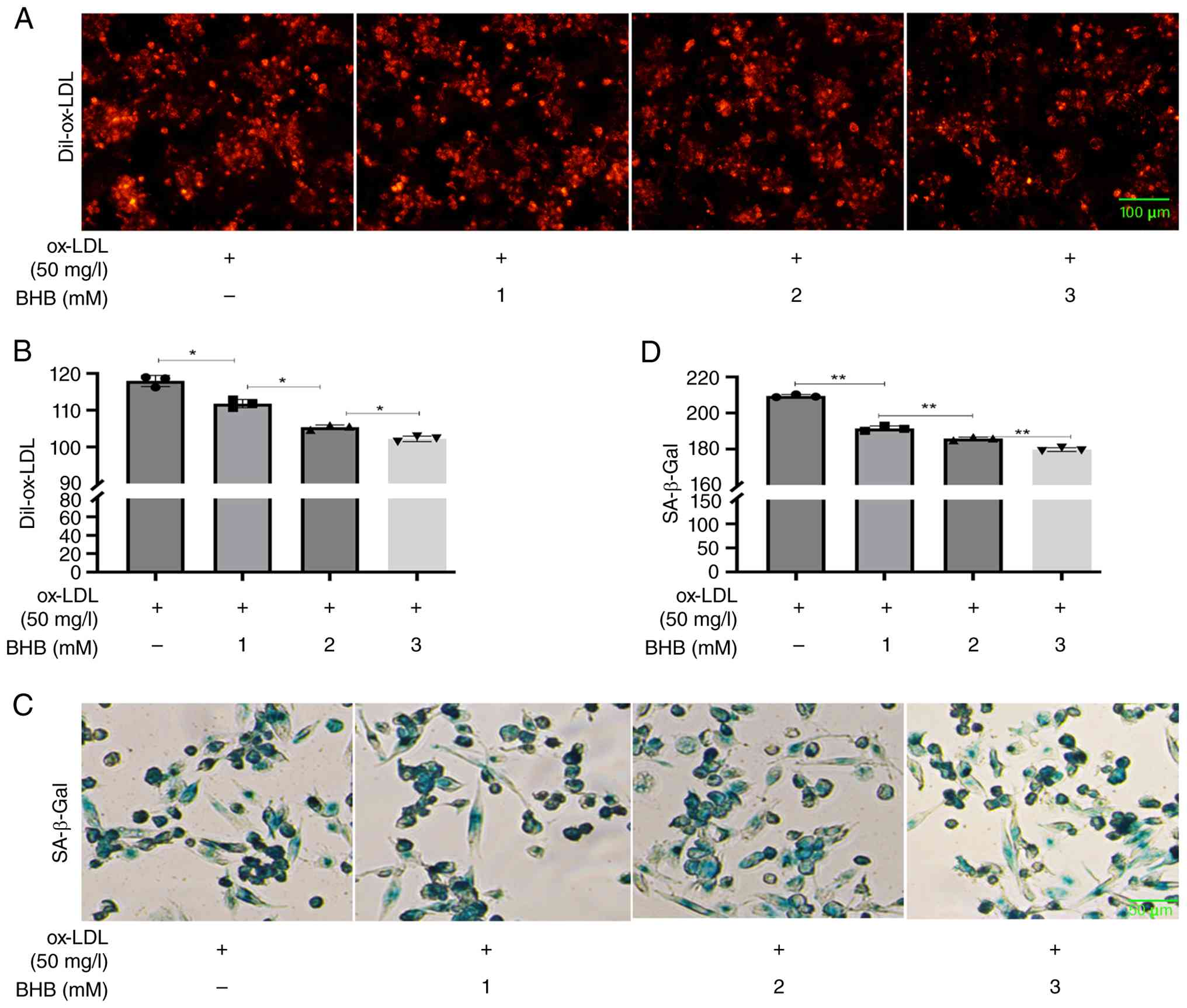 | Figure 3.BHB decreases ox-LDL accumulation and
β-Gal activity. (A) Representative fluorescence microscopy of (B)
Dil-ox-LDL uptake in THP-1 cells. Scale bar, 100 µm. (C)
Representative (D) SA-β-Gal staining. Scale bar, 50 µm. *P<0.05,
**P<0.01. BHB, β-hydroxybutyrate; Dil-ox-LDL,
1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine
perchlorate-oxidized low-density lipoprotein; SA-β-Gal,
senescence-associated β-galactosidase. |
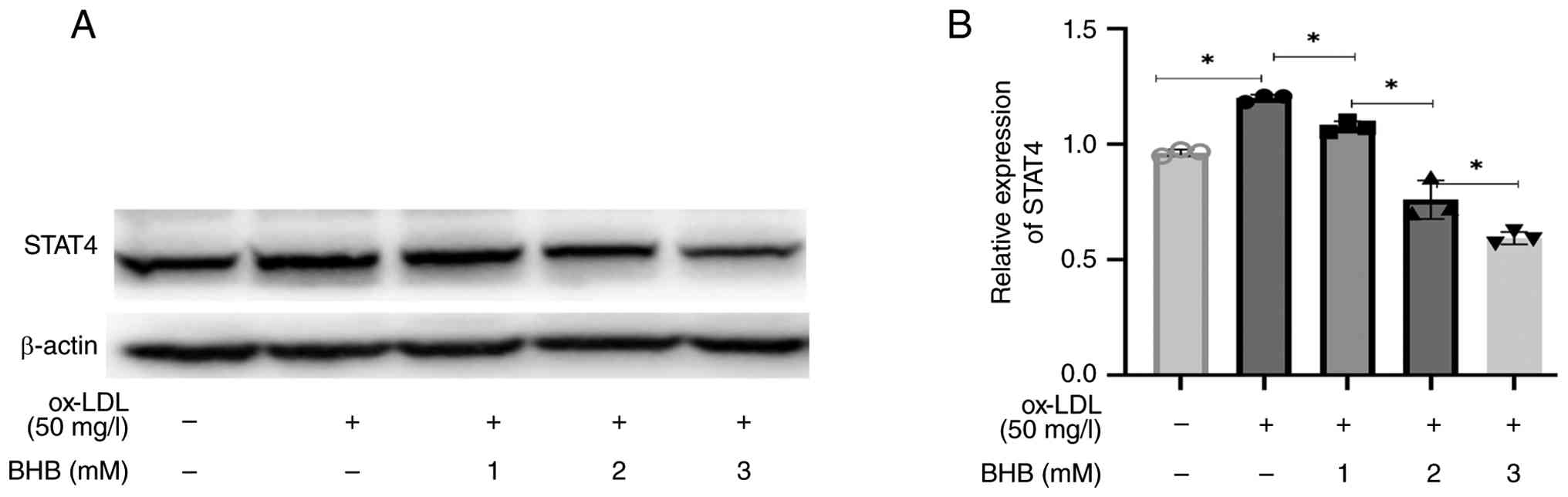
BHB downregulates STAT4 expression in
ox-LDL-treated THP-1 cells
ox-LDL significantly upregulated STAT4 protein
expression in THP-1 cells (Fig. 4A and
B). Conversely, BHB substantially decreased STAT4 levels in a
dose-dependent manner.
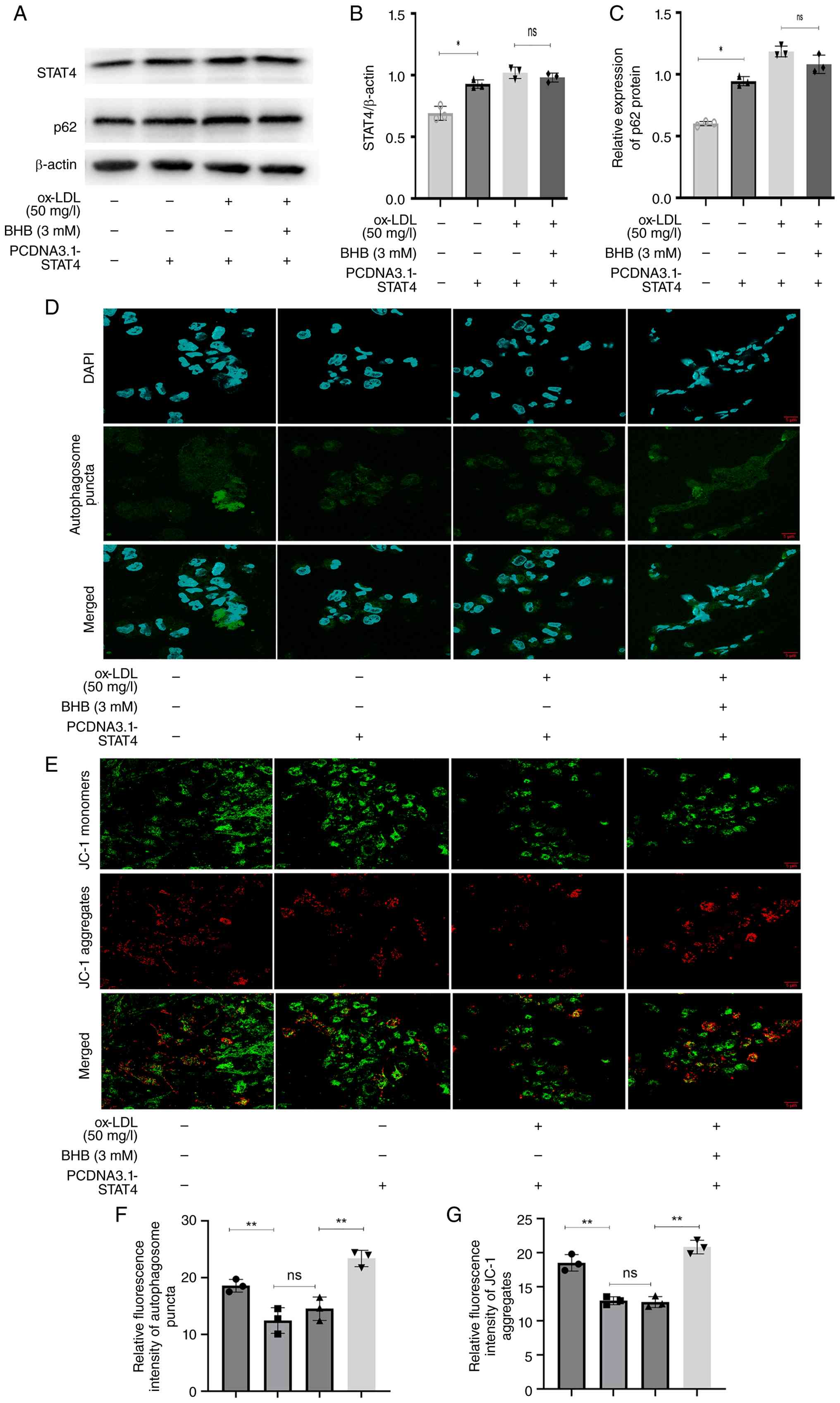
STAT4 overexpression impairs
BHB-mediated restoration of autophagic flux
The present study examined the role of STAT4 in
mediating the rescue effect of BHB on ox-LDL-induced impairment of
autophagic flux. Following overexpression of STAT4 (Fig. S1A), p62 levels increased
significantly (Fig. 5A-C), whereas
autophagic puncta decreased (Fig. 5D
and F) and the ΔΨm declined (Fig.
5E and G). Following co-treatment with ox-LDL and BHB,
STAT4-overexpressing cells exhibited no significant alterations in
p62 expression, autophagosome count or ΔΨm. Collectively, these
findings demonstrate that STAT4 overexpression abrogated
BHB-mediated restoration of impaired autophagic flux and
mitochondrial function.
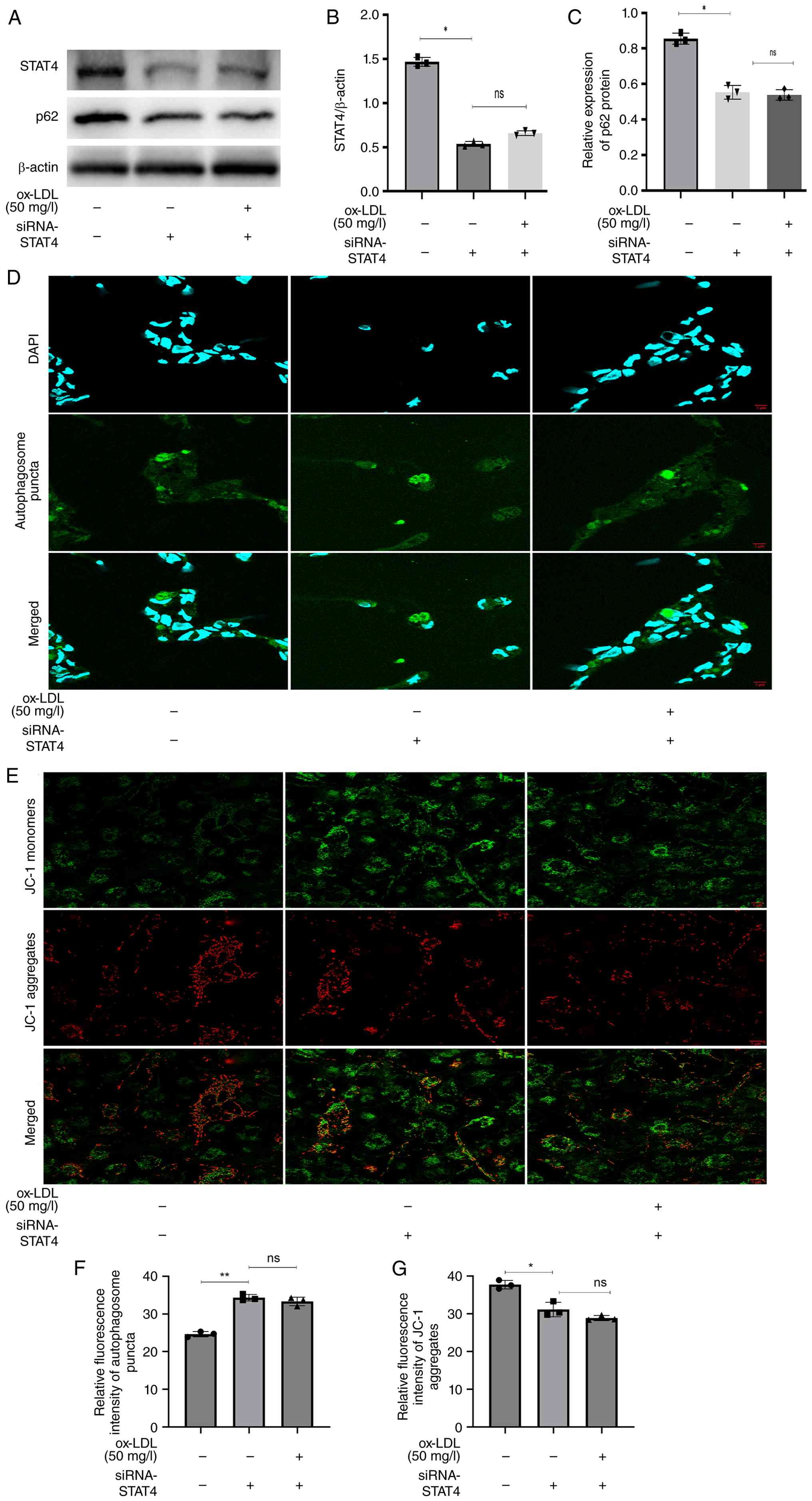
STAT4 silencing prevents
Ox-LDL-induced impairment of autophagic flux and Ox-LDL-induced
reduction of the ΔΨm in THP-1 cells
siRNA transfection was performed to explore the role
of STAT4 in autophagic recovery and ox-LDL uptake. Compared with
cells transfected with scrambled RNA (Fig. S1B), STAT4 silencing significantly
decreased p62 expression (Fig.
6A-C), increased autophagosome formation (Fig. 6D and F) and elevated ΔΨm (Fig. 6E and G). However, when ox-LDL was
added following siRNA-STAT4 transfection in THP-1 cells, the
results did not show the same pattern as before in
non-siRNA-STAT4-transfected THP-1 cells [increased p62 expression
levels, impaired autophagic flux (Fig.
1A-F) and decreased ΔΨm (Fig. 1E
and G)]. As aforementioned, silencing of STAT4 markedly
attenuated ox-LDL-induced impairment of autophagic flux and the
accompanying decline in ΔΨm in THP-1 cells.
Discussion
The pathophysiology of CVD, particularly
atherosclerosis, is influenced by the accumulation of ox-LDL, which
serves a pivotal role in macrophage dysfunction and foam cell
formation (27). This accumulation
initiates a cascade of inflammatory responses and promotes cell
senescence, contributing to disease progression and enhancing
susceptibility to other types of metabolic disorder, such as
glucose metabolism disorders (28,29).
The present study examined the impact of BHB on
autophagy and mitochondrial function in THP-1 macrophages exposed
to ox-LDL. ox-LDL impairs autophagic flux and compromises
mitochondrial integrity, resulting in elevated cell stress and
inflammation (30). Here, BHB
significantly restored autophagic function and enhanced ΔΨm in
these cells, implying a potential therapeutic benefit of BHB in
mitigating the deleterious effects of ox-LDL. Moreover, modulation
of the STAT4 signaling pathway is a key mechanism by which BHB
exerts its restorative effects, underscoring the importance of
targeting autophagy to improve macrophage function in
atherosclerosis (31).
Recent studies have demonstrated that ox-LDL-induced
impairment of autophagy flux and mitochondrial dysfunction in THP-1
macrophages serve critical roles in the progression of
atherosclerosis (32,33). Gu et al (34) showed that ox-LDL impairs autophagy
flux in THP-1 cells, whereas treatment with nicotinate-curcumin
rescues the impaired autophagic flux by significantly increasing
LC3-II levels, autolysosome numbers and p62 degradation in
ox-LDL-treated THP-1 cells, similar to the present study. The
present study examined the expression of the key selective
autophagy protein p62. Accumulation of p62 protein typically
suggests impairment or inhibition of autophagic flux (35). Changes in p62 alongside LC3-II
levels reflect autophagic activity (36). The present study demonstrated an
increase in LC3-II levels along with a decrease in p62, indicating
overall activation of autophagic flux (37,38).
In human umbilical vein endothelial cells (HUVECs)
treated with ox-LDL, silencing of syntaxin 17, a key regulator of
autophagosome-lysosome fusion, exacerbates autophagic flux
impairment and inflammatory responses (39). Here, ox-LDL induced decreased ΔΨm
in THP-1 macrophages, a phenomenon increasingly associated with
oxidative stress and inflammatory signaling (40,41).
Erianin reverses ox-LDL-induced decreased ΔΨm in HUVECs by
activating the Nrf2 pathway, which enhances antioxidant defenses
(42). These findings align with
the present observations, suggesting that decreased ΔΨm may stem
from impaired electron transport chain activity and ROS
accumulation.
BHB, a primary ketone body, serves a key role in
regulating macrophage function and foam cell formation. A recent
study has demonstrated that BHB suppresses pro-inflammatory M1
macrophage polarization by inducing histone β-hydroxybutyrylation
(Kbhb) of STAT1 at lysine 679, thereby inhibiting its
phosphorylation and inflammatory cytokine production (such as IL-1β
and TNF-α) (43). Conversely, BHB
promotes anti-inflammatory M2 polarization via STAT6-dependent
pathways, enhancing the secretion of IL-10 and arginase-1 (Arg-1),
which facilitates tissue repair and decreases lipid accumulation in
macrophages (44). In
atherosclerosis models, BHB decreases ox-LDL uptake by
downregulating scavenger receptors (such as CD36) and upregulating
cholesterol efflux transporters (ATP-binding cassette protein A1
and ATP-binding cassette transporter G1), thereby attenuating foam
cell formation (45,46). These immunometabolic shifts
underscore the dual role of BGB in mitigating inflammation and
lipid overload, positioning it as a promising therapeutic candidate
for vascular pathology.
BHB directly modulates STAT transcription factors
through epigenetic and post-translational mechanisms (47). Bai et al (43) identified that BHB induces Kbhb
modification of STAT1 at lysine 679, which impairs its
phosphorylation and nuclear translocation, thereby inhibiting
lipopolysaccharide-driven M1 polarization. Conversely, BHB
activates STAT6 phosphorylation via IL-4 receptor α signaling,
driving M2-associated gene expression. In endothelial cells, BHB
enhances STAT6-mediated Nrf2 activation, thereby boosting
antioxidant defenses and decreasing oxidative stress (48). This STAT-centric regulation
underpins the ability of BHB to rebalance macrophage phenotypes by
suppressing STAT1/NF-κB-driven inflammation and amplifying
STAT6-mediated resolution pathways. These mechanisms may contribute
to the decreased plaque instability observed in atherosclerotic
models treated with BHB or ketogenic diets (49,50).
STAT signaling intersects with mitochondrial
homeostasis and autophagy, which are key for macrophage lipid
metabolism (51). STAT1
hyperactivation promotes mitochondrial ROS overproduction and
membrane depolarization, thereby exacerbating oxidative damage and
impairing mitophagy (52). By
contrast, STAT6 activation by BHB upregulates Nrf2, which enhances
mitochondrial biogenesis and stabilizes membrane potential via
peroxisome proliferator-activated receptor γ coactivator 1-α
(48,53). Furthermore, STAT6-dependent
induction of Arg-1 supports autophagy flux through mTOR inhibition,
facilitating the clearance of ox-lipids (44). In BHB-treated macrophages, the
suppression of STAT1 coupled with the activation of STAT6 preserves
mitochondrial integrity and promotes autophagy-mediated cholesterol
efflux, thereby decreasing foam cell transformation (54). This crosstalk highlights STATs as a
link between metabolic reprogramming and cell quality control.
Mitochondrial dysfunction is increasingly recognized
as a central driver of vascular injury (55,56).
The NADH dehydrogenase (ubiquinone) Fe-S protein 4 (NDUFS4)-sirtuin
5 (SIRT5)-dual specificity phosphatase 1 (DUSP1) axis regulates
mitochondrial unfolded protein response (mtUPR) and energy
metabolism, thereby protecting against coronary microvascular
injury during ischemia-reperfusion (57). Similarly, targeting the SIRT5/DUSP1
pathway is a promising strategy for mitochondrial repair in
doxorubicin-induced cardiotoxicity (58). In the present study, BHB restored
autophagic activity and ΔΨm, which may involve analogous pathways,
as STAT4 modulation may intersect with these mitochondrial
regulatory networks.
The interplay between mitochondria and the
endoplasmic reticulum (ER) has emerged as a key determinant of cell
stress responses (59).
Astragaloside IV attenuates mechanical stress-induced cardiac
remodeling through the Piezo1-voltage-dependent anion channel 1
axis, which regulates ER UPR (60). Tanshinone IIA modulates METTL3- and
SIRT5-mediated UPR pathways to govern mitochondria-ER crosstalk in
coronary microvascular injury (61). The potential involvement of
ER-mitochondria coupling in ox-LDL-challenged macrophages warrants
further investigation.
Mitophagy and mitochondrial dynamics are key nodes
in mitochondrial quality control networks. The nuclear receptor
subfamily 4 group A member 1-mitochondrial fission factor-FUN14
domain containing 1 axis regulates mitochondrial fragmentation and
mitophagy in myocardial ischemia-reperfusion injury (62). In atherosclerosis, targeting of the
transmembrane BAX inhibitor motif containing 6/NDUFS4 pathway
restores mitophagy and suppresses necroptosis (63). The present study demonstrated that
BHB enhancement autophagic function may extend to mitophagy
regulation, potentially via STAT4-dependent mechanisms. In
addition, the DNA-dependent protein kinase catalytic
subunit-yes-associated protein 1-ferroptosis axis is implicated in
diabetic cardiomyopathy (64),
suggesting broader crosstalk between DNA damage responses and
organelle homeostasis.
The present study had certain limitations that
warrant consideration. A key limitation of this study is the
assessment of autophagy primarily through steady-state levels of
LC3-II and p62 without the use of lysosomal inhibitors to directly
measure autophagic flux. Future investigations should establish the
dynamics of the autophagic process and include standard
pharmacological approaches using bafilomycin A1 or chloroquine to
trace LC3-II and p62 turnover. The present study used
monodansylcadaverine-based staining, which does not distinguish
between autophagosomes and autolysosomes and does not give a
definitive assessment of autophagic flux. Furthermore, employing
more advanced techniques such as live-cell imaging with tandem
fluorescent-tagged LC3 [red fluorescent protein (RFP)-green
fluorescent protein (GFP)-LC3] is required to confirm whether the
present changes represent an upregulation of functional autophagy
or a potential impairment at the fusion/degradation stage. This
method allows precise discrimination between autophagosomes
(GFP+/RFP+) and autolysosomes (GFP-/RFP+), providing a
direct and quantitative measure of autophagic flux and maturation
independently of lysosomal inhibitors (65). The present findings were derived
from in vitro experiments using THP-1 macrophage cell lines,
which may not fully replicate the complex physiological and
pathological contexts of human atherosclerosis. The modulation of
STAT4 underscores the complexity of the signaling pathways
involved. Specifically, the present data do not demonstrate whether
STAT4 acts upstream or downstream of mitochondrial functional
changes. To identify the mechanistic interplay between STAT4
signaling and autophagy, subsequent experiments should assess
levels of phosphorylated STAT4 and STAT4 nuclear localization to
validate the activation status and subcellular distribution of
STAT4 in response to BHB.
In conclusion, the present study demonstrated the
detrimental effects of ox-LDL on autophagic flux and mitochondrial
function in THP-1 macrophages, while highlighting the potential of
BHB to restore these processes. The present findings contribute to
understanding of the cell mechanisms underlying lipid metabolism
and inflammation and suggest BHB may be a promising therapeutic
candidate for the treatment of atherosclerosis. Future studies
should investigate the translational potential of these findings
in vivo to validate the clinical applicability of BHB in
cardiomyocyte energy metabolism and CVD.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Joint Construction
Project of the Henan Provincial Medical Science and Technology
Research Program (grant no. LHGJ20230035), Henan Natural Science
Foundation (grant no. 222300420355) from the Henan Provincial
Department of Education and the Henan Province Science and
Technology Research Program Project (grant no. SBGJ202103016) from
the Henan Provincial Health Commission.
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
WL designed the study and wrote the manuscript. MH
performed the experiments. WL and HW analyzed the data. WL and HW
confirm the authenticity of all the raw data. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
CVD
|
cardiovascular disease
|
|
ox-LDL
|
oxidized low-density lipoprotein
|
|
BHB:
|
β-hydroxybutyrate
|
|
PMA
|
phorbol 12-myristate 13-acetate
|
|
SA-β-Gal
|
senescence-associated
β-galactosidase
|
|
ROS
|
reactive oxygen species
|
References
|
1
|
Roth GA, Mensah GA, Johnson CO, Addolorato
G, Ammirati E, Baddour LM, Barengo NC, Beaton AZ, Benjamin EJ,
Benziger CP, et al: Global burden of cardiovascular diseases and
risk factors, 1990–2019: Update from the GBD 2019 study. J Am Coll
Cardiol. 76:2982–3021. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhi X, Sun Y, Cai F, Wang S, Gao H, Wu F,
Zhang L and Shen Z: Oxidized Low-density lipoprotein
(Ox-LDL)-triggered double-lock probe for spatiotemporal lipoprotein
oxidation and atherosclerotic plaque imaging. Adv Healthc Mater.
12:e23015952023. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Qiao YN, Zou YL and Guo SD: Low-density
lipoprotein particles in atherosclerosis. Front Physiol.
13:9319312022. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Obermayer G, Afonyushkin T and Binder CJ:
Oxidized low-density lipoprotein in inflammation-driven thrombosis.
J Thromb Haemost. 16:418–428. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Thangasparan S, Kamisah Y, Ugusman A,
Mohamad Anuar NN and Ibrahim N: Unravelling the Mechanisms of
oxidised Low-density lipoprotein in cardiovascular health: Current
evidence from in vitro and in vivo studies. Int J Mol Sci.
25:132922024. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Munno M, Mallia A, Greco A, Modafferi G,
Banfi C and Eligini S: Radical oxygen species, oxidized low-density
lipoproteins, and lectin-like oxidized low-density lipoprotein
receptor 1: A vicious circle in atherosclerotic process.
Antioxidants (Basel). 13:5832024. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
He Y and Liu T: Oxidized low-density
lipoprotein regulates macrophage polarization in atherosclerosis.
Int Immunopharmacol. 120:1103382023. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang L and Guo H: Acetylation
modification in the regulation of macroautophagy. Adv Biotechnol
(Singap). 2:192024. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang X, Misra SK, Moitra P, Zhang X,
Jeong SJ, Stitham J, Rodriguez-Velez A, Park A, Yeh YS, Gillanders
WE, et al: Use of acidic nanoparticles to rescue macrophage
lysosomal dysfunction in atherosclerosis. Autophagy. 19:886–903.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li P, Li H, Li X, Li S, Xu H, Cui J, Cheng
G, Liu Y, Xu X, Xin Y and Liu A: San Jie tong Mai fang protects
against atherosclerosis progression by regulating macroautophagy
through the PI3K/AKT/mTOR signaling pathway. J Cardiovasc
Pharmacol. 82:333–343. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Salminen A and Kaarniranta K: Regulation
of the aging process by autophagy. Trends Mol Med. 15:217–224.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Robichaud S, Fairman G, Vijithakumar V,
Mak E, Cook DP, Pelletier AR, Huard S, Vanderhyden BC, Figeys D,
Lavallée-Adam M, et al: Identification of novel lipid droplet
factors that regulate lipophagy and cholesterol efflux in
macrophage foam cells. Autophagy. 17:3671–389. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Seidenberg J, Stellato M, Hukara A,
Ludewig B, Klingel K, Distler O, Błyszczuk P and Kania G: The AP-1
transcription factor Fosl-2 regulates autophagy in cardiac
fibroblasts during myocardial fibrogenesis. Int J Mol Sci.
22:18612021. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu Y, Teng Y, Zhang C, Pan Y, Zhang Q, Zhu
X, Liu N, Su X and Lin J: The ketone body β-hydroxybutyrate
alleviates CoCrMo alloy particles induced osteolysis by regulating
NLRP3 inflammasome and osteoclast differentiation. J
Nanobiotechnology. 20:1202022. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chu Y, Hua Y, He L, He J, Chen Y, Yang J,
Mahmoud I, Zeng F, Zeng X, Benavides GA, et al:
beta-hydroxybutyrate administered at reperfusion reduces infarct
size and preserves cardiac function by improving mitochondrial
function through autophagy in male mice. J Mol Cell Cardiol.
186:31–44. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu Y, Wang W, Zhang J, Gao S, Xu T and
Yin Y: JAK/STAT signaling in diabetic kidney disease. Front Cell
Dev Biol. 11:12332592023. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Billah M, Ridiandries A, Allahwala UK,
Mudaliar H, Dona A, Hunyor S, Khachigian LM and Bhindi R: Remote
ischemic preconditioning induces cardioprotective autophagy and
signals through the IL-6-Dependent JAK-STAT pathway. Int J Mol Sci.
21:16922020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen D, Liu Y, Chen J, Lin H, Guo H, Wu Y,
Xu Y, Zhou Y, Zhou W, Lu R, et al: JAK/STAT pathway promotes the
progression of diabetic kidney disease via autophagy in podocytes.
Eur J Pharmacol. 902:1741212021. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu Z, Hu K, Chen YS, Huang YJ, Hu Q, Zeng
W, Cao Y, Xiao Q and Zhang XK: JAK2/STAT3 inhibition attenuates
intestinal ischemia-reperfusion injury via promoting autophagy: In
vitro and in vivo study. Mol Biol Rep. 49:2857–2867. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang Y, Liu M, Wu Y, Xu Y, Hong Y and
Xiang H: Insulin-like growth factor 1 knockdown attenuates high
glucose-induced podocyte injury by promoting the JAK2/STAT
signalling-mediated autophagy. Nephrology (Carlton). 29:394–404.
2024. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Madhavan A, Arun KB, Pushparajan AR,
Balaji M and Kumar RA: Transcription repressor protein ZBTB25
associates with HDAC1-Sin3a complex in Mycobacterium
tuberculosis-infected macrophages, and its inhibition clears
pathogen by autophagy. mSphere. 6:e00036–21. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gao Y, Chen S, Jiao S, Fan Y, Li X, Tan N,
Fang J, Xu L, Huang Y, Zhao J, et al: ATG5-regulated CCL2/MCP-1
production in myeloid cells selectively modulates anti-malarial
CD4+ Th1 responses. Autophagy. 20:1398–1417. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Scarno G, Mazej J, Laffranchi M, Di Censo
C, Mattiola I, Candelotti AM, Pietropaolo G, Stabile H, Fionda C,
Peruzzi G, et al: Divergent roles for STAT4 in shaping
differentiation of cytotoxic ILC1 and NK cells during gut
inflammation. Proc Natl Acad Sci USA. 120:e23067611202023.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang X, Wang Y and Lv J: STAT4 targets
KISS1 to inhibit the oxidative damage, inflammation and neuronal
apoptosis in experimental PD models by inactivating the MAPK
pathway. Neurochem Int. 175:1056832024. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Semenova N, Yakisich JS, Ascue R, Iyer AKV
and Azad N: Reversible upregulation of the Senescence-Associated
Beta-Galactosidase marker induced by cell detachment in cancer
cells. Cells. 14:16672025. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sanchez-Leon ME, Loaeza-Reyes KJ,
Matias-Cervantes CA, Mayoral-Andrade G, Pérez-Campos EL,
Pérez-Campos-Mayoral L, Hernández-Huerta MT, Zenteno E,
Pérez-Cervera Y and Pina-Canseco S: LOX-1 in cardiovascular
disease: A comprehensive molecular and clinical review. Int J Mol
Sci. 25:52762024. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kishimoto H, Akagi M, Zushi S, Teramura T,
Onodera Y, Sawamura T and Hamanishi C: Induction of hypertrophic
chondrocyte-like phenotypes by oxidized LDL in cultured bovine
articular chondrocytes through increase in oxidative stress.
Osteoarthritis Cartilage. 18:1284–1290. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Palmer AK, Spinelli R, Prata LGL, Chaib S,
Suda M, Tchkonia T, Smith U and Kirkland JL: Senotherapeutics for
metabolic disease and diabetic complications. J Intern Med.
299:2–19. 2026. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hao T, Fang W, Xu D, Chen Q, Liu Q, Cui K,
Cao X, Li Y, Mai K and Ai Q: Phosphatidylethanolamine alleviates
OX-LDL-induced macrophage inflammation by upregulating autophagy
and inhibiting NLRP1 inflammasome activation. Free Radic Biol Med.
208:402–417. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang G, Chen JH, Qiang Y, Wang DZ and Chen
Z: Decreased STAT4 indicates poor prognosis and enhanced cell
proliferation in hepatocellular carcinoma. World J Gastroenterol.
21:3983–3993. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Miao X, Pan R and Chang F: Down-regulation
of ATP8B2 in foam cells inhibits autophagic flux and ox-LDL
degradation in atherosclerosis. Cell Biochem Biophys. 83:3451–3463.
2025. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang J, Ma X, Niu D, Sun Y, Chai X, Deng
Y, Wang J and Dong J: PCSK9 inhibitors suppress oxidative stress
and inflammation in atherosclerotic development by promoting
macrophage autophagy. Am J Transl Res. 15:5129–5144.
2023.PubMed/NCBI
|
|
34
|
Gu HF, Li HZ, Tang YL, Tang XQ, Zheng XL
and Liao DF: Nicotinate-Curcumin impedes foam cell formation from
THP-1 cells through restoring autophagy flux. PLoS One.
11:e01548202016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhou P, Zhang Q, Yang Y, Wu W, Chen D,
Zheng Z, Jongkaewwattana A, Jin H, Zhou H and Luo R: Cleavage of
SQSTM1/p62 by the Zika virus protease NS2B3 prevents autophagic
degradation of viral NS3 and NS5 proteins. Autophagy. 20:2769–2784.
2024. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wu K, Seylani A, Wu J, Wu X, Bleck CKE and
Sack MN: BLOC1S1/GCN5L1/BORCS1 is a critical mediator for the
initiation of autolysosomal tubulation. Autophagy. 17:3707–3724.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhan Q, Jeon J, Li Y, Huang Y, Xiong J,
Wang Q, Xu TL, Li Y, Ji FH, Du G and Zhu MX: CAMK2/CaMKII activates
MLKL in short-term starvation to facilitate autophagic flux.
Autophagy. 18:726–744. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yuan X, Tian GG, Pei X, Hu X and Wu J:
Spermidine induces cytoprotective autophagy of female germline stem
cells in vitro and ameliorates aging caused by oxidative stress
through upregulated sequestosome-1/p62 expression. Cell Biosci.
11:1072021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cui X, Wang B, Han D, Cheng M, Yuan P, Du
P, Hou Y, Su C, Tang J and Zhang J: Exacerbation of atherosclerosis
by STX17 knockdown: Unravelling the role of autophagy and
inflammation. J Cell Mol Med. 28:e184022024. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yuan L, Fan L and Zhang Z, Huang X, Liu Q
and Zhang Z: Procyanidin B2 alleviates oxidized low-density
lipoprotein-induced cell injury, inflammation, monocyte chemotaxis,
and oxidative stress by inhibiting the nuclear factor kappa-B
pathway in human umbilical vein endothelial cells. BMC Cardiovasc
Disord. 24:2312024. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hua J, Gao Z, Zhong S, Wei B, Zhu J and
Ying R: CISD1 protects against atherosclerosis by suppressing lipid
accumulation and inflammation via mediating Drp1. Biochem Biophys
Res Commun. 577:80–88. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang Z, Wang L, Wang Y and Zhang J:
Erianin protects human umbilical vein endothelial cells from
oxidized Low-Density Lipoprotein-Induced apoptosis and oxidative
stress through activation of nuclear factor E2-Related factor 2
signaling. Chem Biol Drug Des. 105:e701042025. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Bai YP, Xing YJ, Ma T, Li K, Zhang T, Wang
DG, Wan SJ, Zhang CW, Sun Y and Wang MY: beta-Hydroxybutyrate
suppresses M1 macrophage polarization through
beta-hydroxybutyrylation of the STAT1 protein. Cell Death Dis.
15:8742024. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Huang C, Wang J, Liu H, Huang R, Yan X,
Song M, Tan G and Zhi F: Ketone body β-hydroxybutyrate ameliorates
colitis by promoting M2 macrophage polarization through the
STAT6-dependent signaling pathway. BMC Med. 20:1482022. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang N, Liu C, Jin L, Zhang R, Wang T,
Wang Q, Chen J, Yang F, Siebert HC and Zheng X: Ketogenic diet
elicits antitumor properties through inducing oxidative stress,
inhibiting MMP-9 expression, and rebalancing M1/M2 Tumor-associated
macrophage phenotype in a mouse model of colon cancer. J Agric Food
Chem. 68:11182–11196. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Xu Z, Zhang M, Li X, Wang Y and Du R:
Exercise ameliorates atherosclerosis via Up-regulating serum
β-Hydroxybutyrate levels. Int J Mol Sci. 23:37882022. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ehtiati S, Hatami B, Khatami SH,
Tajernarenj K, Abdi S, Sirati-Sabet M, Ghazizadeh Hashemi SAH,
Ahmadzade R, Hamed N, Goudarzi M, et al: The multifaceted influence
of Beta-Hydroxybutyrate on autophagy, mitochondrial metabolism, and
epigenetic regulation. J Cell Biochem. 126:e700502025. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ji LW, Deng Y and Li T: Effect of Ketone
Body β-Hydroxybutyrate to attenuate inflammation-induced
mitochondrial oxidative stress in vascular endothelial cells.
Sichuan Da Xue Xue Bao Yi Xue Ban. 52:954–959. 2021.(In Chinese).
PubMed/NCBI
|
|
49
|
Huang X, Xie M, Wang Y, Lu X, Mei F, Zhang
K, Yang X, Chen G, Yin Y, Feng G, et al: Porphyromonas gingivalis
aggravates atherosclerotic plaque instability by promoting
lipid-laden macrophage necroptosis. Signal Transduct Target Ther.
10:1712025. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Cui Y, Chen Y, Li H, Zhang W, Wang X, Xia
M, Gan N, Zhou Y, Li M, Zhang H, et al: PCSK9 Promotes
Atherosclerotic Plaque Instability by Inducing VSMC Ferroptosis
through the YAP1-NUPR1 Axis. Research (Wash D C).
8:09222025.PubMed/NCBI
|
|
51
|
Hu W, Liu H, Lin L, Li M, Fan Z, Zhang Y,
Lin X and Qi Y: Feedback inhibition of the Janus kinase/signal
transducer and activator of transcription signaling pathway by
CG5953 through Ptp61F. Int J Biol Macromol. 330:1478752025.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ji L, He Q, Liu Y, Deng Y, Xie M, Luo K,
Cai X, Zuo Y, Wu W, Li Q, et al: Ketone Body β-hydroxybutyrate
prevents myocardial oxidative stress in septic cardiomyopathy. Oxid
Med Cell Longev. 2022:25138372022. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Lu F, Wang R, Cheng Y and Li X:
Preconditioning with β-hydroxybutyrate attenuates lung
ischemia-reperfusion injury by suppressing alveolar macrophage
pyroptosis through the SIRT1-FOXO3 signaling pathway. FASEB J.
38:e700272024. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wang C, Xu W, Jiang S, Wu Y, Shu J, Gao X
and Huang K: β-Hydroxybutyrate facilitates postinfarction cardiac
repair via targeting PHD2. Circ Res. 136:704–718. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Marcheggiani F, Nunzi I, Rao L, Dhaouadi
N, Nesci S, Pinton P and Marchi S: Mitochondrial dysfunction in
cerebrovascular diseases. Trends Mol Med. Apr 30–2026.doi:
10.1016/j.molmed.2026.04.002 (Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yang W, He J, Yu H, Wu Y and Shi S:
Mitochondrial dysfunction: Potential therapy for abdominal aortic
aneurysms. Curr Vasc Pharmacol. 23:255–271. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Pu X, Liu J, Wang Y, Guan X, Wu Q, Zhang
Q, Liu R and Chang X: Ginsenoside Rb1 attenuates coronary
microvascular inflammatory injury via NDUFS4-SIRT5-DUSP1-mediated
mitochondrial quality control in a murine ischemia-reperfusion
model. J Ginseng Res. 49:509–522. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Shi H, Pang B, Zhang F, Guo Z, Xu W, Zheng
M, You Y, Liu G, Nie Y, Liang J and Chang X: A novel
ligustrazine-based nanodelivery system protects against
doxorubicin-induced cardiotoxicity by targeting the SIRT5-DUSP1
axis for mitochondrial repair. J Nanobiotechnology. 23:6812025.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Padhy B, Xie J, Idrees D, Cheng CJ and
Huang CL: Endoplasmic reticulum-mitochondrion disconnection
promotes metabolic reprogramming and cystogenesis in polycystic
kidney disease. bioRxiv. Nov 12–2025.(Epub ahead of print). doi:
10.1101/2025.10.31.685870.
|
|
60
|
Zhang S, Gao W, Gao X, Xu W, Liu Y, Guo Z,
Liu G, Zhang P, Shi H and Chang X: Astragaloside VI attenuates
mechanical stress-induced cardiac remodeling through piezo1-VDAC1
dependent endoplasmic reticulum unfolded protein response.
Phytomedicine. 148:1572882025. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Pu X, Wu Q, Yan Z, Zhou S, Zhang Q, Zhang
X, Cai Y, Liu Z, Liu R and Chang X: Tanshinone IIA modulates Sirt5
and Metll3 interaction to govern mitochondria-endoplasmic reticulum
unfolded protein response in coronary microvascular injury.
Phytomedicine. 145:1569822025. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wang J, Zhuang H, Jia L, He X, Zheng S, Ji
K, Xie K, Ying T, Zhang Y, Li C and Chang X: Nuclear receptor
subfamily 4 group A member 1 promotes myocardial
ischemia/reperfusion injury through inducing mitochondrial fission
factor-mediated mitochondrial fragmentation and inhibiting FUN14
domain containing 1-depedent mitophagy. Int J Biol Sci.
20:4458–4475. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Chang X, Zhou H, Hu J, Ge T, He K, Chen Y,
Zou R and Fan X: Targeting mitochondria by lipid-selenium conjugate
drug results in malate/fumarate exhaustion and induces
mitophagy-mediated necroptosis suppression. Int J Biol Sci.
20:5793–5811. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Wang J, Chang X, Li C, Gao J, Guo Z,
Zhuang H, Wang L, Huang Y, Wang W, Li C and He Q: DNA-PKcs-Driven
YAP1 phosphorylation and nuclear translocation: A key regulator of
ferroptosis in hyperglycemia-induced cardiac dysfunction in type 1
diabetes. Adv Sci (Weinh). 12:e24126982025. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Jiang XL, Liu B, Li JK, Lin YF, Zhu PL,
Zhang Z, Wang Y, Deng B, Zhang JZ and Yung KK: Przewaquinone A, as
a natural STAT3 inhibitor, suppresses the growth of melanoma cells
and induces autophagy. Phytomedicine. 142:1568102025. View Article : Google Scholar : PubMed/NCBI
|