Introduction
Malignant tumors represent one of the leading causes
of human mortality, and the primary methods for treating tumors are
surgical resection and chemotherapy (1). Cisplatin, also known as
cis-dichlorodiamine platinum, is one of the most prevalent and
effective chemotherapy drugs at present. Cisplatin is predominantly
used to treat breast, ovarian, esophageal, bladder, head and neck
cancers, as well as non-small cell lung cancer and other solid
tumors (2–4). Mechanistically, cisplatin
predominantly induces tumor cell death by cross-linking with purine
bases in DNA, resulting in DNA damage that subsequently activates
various apoptotic or necrotic pathways (5). However, cisplatin has been shown to
not only kill tumor cells but also normal cells, resulting in toxic
side effects, such as ototoxicity, neurotoxicity, bone marrow
suppression and nephrotoxicity, which severely limit the clinical
application of cisplatin (6).
Nephrotoxicity represents the dose-limiting factor for cisplatin
use and remains the predominant factor limiting its clinical
application (7). A study has shown
that 30–40% of patients treated with cisplatin experience
nephrotoxicity, which predominantly manifests as acute kidney
injury, depletion of salt or magnesium ions, and loss of
urine-concentration function (8).
At present, drugs used clinically to alleviate or
prevent cisplatin-induced acute kidney injury (CI-AKI) remain
limited in their application due to various side effects (9). Additionally, the emergence of
chimeric antigen receptor (CAR)-T-cell therapy has demonstrated
notable potential for improving treatment of CI-AKI. A previous
clinical trial has shown that although anti-CD19 CAR-T cells
demonstrate notable clinical efficacy, their long-term safety and
sustained therapeutic benefits represent ongoing translational
challenges (10). Therefore,
exploring adjuvant drugs that can effectively alleviate kidney
injury during cisplatin chemotherapy will notably improve the
clinical application of cisplatin.
The exact mechanisms of CI-AKI remain ambiguous.
Several molecular mechanisms that contribute to CI-AKI pathogenesis
include the uptake and accumulation of cisplatin, oxidative stress,
the inflammatory response, cell apoptosis and necrosis (11,12).
In particular, oxidative stress caused by the excessive generation
of reactive oxygen species (ROS) is associated with CI-AKI
pathogenesis (13). Cisplatin
enters cells and generates ROS through normal metabolic processes,
which deplete antioxidant substances, such as superoxide dismutase
(SOD), glutathione (GSH) and malondialdehyde (MDA), and disrupt the
dynamic balance of intracellular oxidation-reduction reactions,
resulting in increased oxidative stress (14). In addition, the excessive
production of ROS can activate the NF-κB) signaling pathway,
upregulate the expression of pro-inflammatory factors, such as
tumor necrosis factor-α (TNF-α) and interleukin (IL)-1β, and alter
the activity of multiple signaling pathways that lead to cell
apoptosis (15). Pro-inflammatory
ILs have been shown to facilitate CI-AKI progression by recruiting
neutrophils and inducing apoptosis of renal cells. However,
anti-inflammatory ILs have been shown to attenuate CI-AKI by
promoting regulatory T cell activity and suppressing
pro-inflammatory cytokines (16).
Nuclear factor erythroid-2-related factor 2 (NRF2)
is a common transcription factor that can regulate oxidative
stress, drug detoxification and the inflammatory response through
various pathways (17). Primarily,
NRF2 can inhibit oxidative stress and activation of the NF-κB
signaling pathway by suppressing the accumulation of intracellular
ROS. Additionally, NRF2 has been shown to inhibit the
proteasomal-degradation of IκB-α, a negative regulator of NF-κB,
thereby suppressing the nuclear translocation of NF-κB (18). Therefore, activation of the NRF2
signaling pathway may confer protection against cisplatin-induced
oxidative stress and inflammatory responses, thus mitigating
subsequent tissue damage.
In a study on NRF2 activators, such as sulforaphane
(SFN) and dimethyl fumarate in CI-AKI, SFN was found to
significantly improve mitochondrial function in renal tubular
cells, upregulate cortical NRF2 expression and downregulate the
protein expression of its repressor Keap1, thereby alleviating
oxidative stress and tubular damage induced by cisplatin (19). Dimethyl fumarate activated the NRF2
pathway to increase the expression of downstream cytoprotective
gene NQO1, suppressed the production of inflammatory cytokines
including TNF-α and IL-6, and effectively reduced cisplatin-induced
renal tubulointerstitial fibrosis and tubular necrosis (19). SFN is the hydrolysis product of
thiogluconate, which can be extracted from a number of vegetables,
such as broccoli (20). Notably,
SFN has been proven to be non-toxic, reliable, easy to obtain and
inexpensive and exhibiting a broad range of application prospects
(21). In addition, SFN has been
shown to display a number of therapeutic effects, namely
anti-inflammatory, antioxidant, anticancer effects, in animals
(22). A previous study
demonstrated that metabolites of SFN can be detected in all organs
at up to 6 h after administration, with higher concentrations
observed in the small intestine, prostate, lungs and kidneys,
indicating that SFN is bioabsorbable (23). Furthermore, another study showed
that AKI induced by medication or ischemia can be alleviated by
SFN-activated NRF2 (24). Previous
research found that SFN treatment in mouse models of diabetic
nephropathy markedly reduced renal damage, which may be due to the
protective effect of SFN-mediated NRF2 pathway activation on renal
tissue (25).
However, it remains to be fully elucidated whether
SFN can reduce the severity of CI-AKI by upregulating NRF2
expression. Thus, the present study aimed to establish models of
CI-AKI in mice and HK-2 cells using cisplatin in order to elucidate
the protective effect of SFN on CI-AKI and investigate the
regulatory role of NRF2 expression in this condition.
Materials and methods
Establishment of a CI-AKI mouse model
and SFN treatment
A total of 32 male C57BL/6 mice (age, 6–8 weeks;
body weight, 20–25 g) was procured from the Experimental Animal
Center of Nanjing Medical University (Nanjing, China) and
maintained under specific pathogen-free conditions with ad
libitum access to food and water (22±2°C, Relative Humidity:
40–60%, Light/Dark Cycle: 12/12-h light/ dark cycle). Following a
7-day acclimation period, mice were randomly allocated into four
experimental groups (n=6 per group), including control, SFN (10
mg/kg) (26), cisplatin (20 mg/kg,
CAS No. 15663-27-1, Sigma-Aldrich) (27) and cisplatin + SFN groups. Starting
from day 7 of the experiment, SFN was administered daily to mice in
the relevant groups, whereas control and cisplatin-only groups
received equivalent volumes of saline. After 7 days of
pre-treatment with SFN or saline, mice in the cisplatin and
cisplatin + SFN groups received a single intraperitoneal injection
of cisplatin (20 mg/kg in saline). Euthanasia was performed 72 h
after cisplatin administration via intraperitoneal injection of 5%
sodium pentobarbital (150 mg/kg).
To confirm mortality following euthanasia, mice
underwent checks for: i) Cardiac and respiratory arrest; ii) pain
responses; and iii) pupillary responses to light stimuli. Blood
samples were immediately collected via terminal cardiac puncture. A
total of 0.8–1.0 ml of blood was extracted per mouse into
serum-separating tubes. Serum was obtained by centrifugation of
blood samples at 3,000 g for 15 min at 4°C and samples were stored
at −80°C for subsequent analysis. During the study, the following
humane endpoints were established and strictly observed: i) Severe
lethargy or unresponsiveness to stimuli; ii) loss of >20% body
weight; iii) inability to access food or water; and iv) signs of
severe distress, for example labored breathing or hunched posture
(28). No mice were euthanized
prematurely due to reaching predetermined humane endpoints. All
procedures and were approved by the Ethics Committee of Nanjing
Medical University (approval no. IACUC 2024-0811).
Cell culture and treatment
The human proximal tubular epithelial cell line HK-2
was obtained from the American Type Culture Collection. Cells were
maintained in DMEM/F-12 supplemented with 10% fetal bovine serum
(FBS; both Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin-streptomycin solution comprising 100 U/ml penicillin and
100 µg/ml streptomycin under standard culture conditions (37°C; 5%
CO2 humidified atmosphere). Upon achieving 70%
confluence, cells were serum-starved by switching to FBS-free
medium. Prior to experimental treatments, cells were pre-treated
with one or both of 5 µM SFN and 1.9 µM ML385, an NRF2 inhibitor,
dissolved in dimethyl sulfoxide for 4 h (37°C). Following
pre-treatment, cellular models were established by exposing HK-2
cells to 10 µg/ml cisplatin for 24 h (37°C) (27). For the control group, cells were
cultured in PBS-free medium without pre-treatment or cisplatin.
Blood biochemical analysis
Serum samples from mice in each treatment group were
analyzed to quantify specific biomarkers, namely blood urea
nitrogen (BUN; Jianglai Biotechnology Co., Ltd. (Cat. No. JL12567)
with 50 µl of serum per well, serum creatinine (Scr) from Nanjing
Jiancheng Bioengineering Institute (Cat. No. C011-2-1) with 20 µl
of serum per well, MDA from Xitang Biotechnology Co., Ltd. (Cat.
No. XT-MDA-001) with 10 µl serum per well, SOD from Abcam (Cat. No.
ab65354) using 15 µl of serum per well and GSH from Kaiji
Biotechnology Co., Ltd. (Cat. No. KGA7305) with 50 µl of serum per
well. All biochemical analyses followed established protocols
provided by the manufacturer's instructions.
ELISA
Serum and cell supernatant samples underwent
centrifugation at 3, 000 × g for 10 min at 4°C. The levels of
IL-1β, IL-6 and TNF-α were measured using ELISA kits according to
the instructions provided by the manufacturer (IL-1β, Elabscience
Biotechnology Co., Ltd., No. E-EL-M0059), (IL-6, Elabscience
Biotechnology Co., Ltd., No. E-EL-M0044) and (TNF-α, all
Elabscience Biotechnology Co., Ltd., No. E-EL-M0059,
E-EL-M0049).
Hematoxylin and eosin (H&E)
staining
Mouse renal tissues were fixed in 4% PFA at 4°C for
24 h, then embedded in paraffin. After dewaxing, tissue sections (4
µm) were rehydrated using gradient ethanol. Sections were
subsequently stained with hematoxylin for 10 min, rinsed with tap
water for 2 sec and then stained with eosin for 5 min. After
rinsing with tap water for 1 sec, the sections were dehydrated in
gradient ethanol at 75, 95 and 100%, each for 5 sec, and
subsequently air-dried. All procedures were performed at room
temperature. Changes to mouse renal structure were observed under a
light microscope in five randomly-selected visual fields.
Periodic acid-Schiff (PAS)
staining
PAS staining was performed on 4 µm-thick
paraffin-embedded mouse renal sections. The renal tissues were
fixed in 4% paraformaldehyde at 4°C overnight before embedding.
After deparaffinization in xylene and hydration in a graded ethanol
series, tissue sections were oxidized with 0.5% periodic acid for
10 min and subsequently stained with Schiff reagent for 30 min to
detect polysaccharides and glycoproteins at room temperature.
Nuclei were counterstained with Mayer's hematoxylin for 1 min and
sections were subsequently differentiated in acid alcohol and blued
in alkaline solution for 2 min at room temperature. Following
dehydration through an ethanol-xylene series, sections were mounted
with resin medium. Staining patterns in five randomly selected were
visualized using bright-field light microscopy at ×20
magnification.
TUNEL staining
Briefly, 5-µm paraffin-embedded renal tissue
sections and cultured HK-2 cells grown on coverslips were fixed in
4% paraformaldehyde (PFA) for 1 h at room temperature. Samples were
subsequently permeabilized using 0.1% Triton X-100 in 0.1% sodium
citrate for 30 min on ice. After washing with PBS, samples were
incubated with TUNEL reaction (One-Step TUNEL Apoptosis Assay kit,
Green Fluorescence, Catalog No. KTA2010, Abbkine, Wuhan, China)
mixture for 1 h at 37°C in a humidified chamber. Nuclei were
counterstained with 1 µg/ml DAPI for 5 min (22–25°C). Slides were
mounted with antifade mounting medium and images of five randomly
selected fields of view were captured using a fluorescence
microscope.
2′,7′-dichlorodihydrofluorescein
diacetate (DCFH-DA) analysis
ROS levels were quantified using the fluorescent
probe DCFH-DA. For the HK-2 cell line, cells (1–5×105)
were cultured to 70–80% confluence in 6-well plates, washed with
PBS and subsequently incubated with 10 µM DCFH-DA in serum-free
medium for 30 min at 37°C in the dark. Cells were then washed twice
with ice-cold PBS to remove extracellular dye. Finally, ROS levels
in different treatment groups were analyzed by flow cytometry.
For mouse renal tissue samples, sections were
equilibrated in PBS (pH 7.4) for 10 min at 37°C and incubated with
10 µM DCFH-DA dissolved in serum-free DMEM (Gibco, No. 12634010)
for 30 min in the dark at 37°C. Excess probe was removed by gently
washing the sections three times with PBS to minimize background
fluorescence. Data were analyzed using FACSCelesta™ and
FlowJo™ Software Version 10.8.1 from BD Biosciences.
Immunohistochemistry (IHC)
Tissue sections (−80°C) were thawed using PBS
solution and subsequently dried using absorbent paper. Sections
(5–10 µm) underwent fixation with 4% PFA for 20 min at room
temperature. Following rinsing with PBS, sections were treated with
0.1% Triton X-100 for 5 min at room temperature. Endogenous
peroxidase activity was blocked with 3% hydrogen peroxide in
methanol for 15 min at room temperature. Sections were then blocked
using a solution of 5% bovine serum albumin (BSA) (Sigma-Aldrich,
No. A7906) for 1 h at room temperature and incubated with the
primary antibodies (F4/80, Abcam, ab300421) 1:500, NRF2 (Abcam,
ab313825) 1:100) overnight at 4°C. Following three washes with PBS,
sections were incubated with the secondary antibody (Goat
Anti-Mouse IgG H&L (HRP)(Abcam, ab205719) 1:2,000) for 1 h at
room temperature. Samples underwent DAPI (Vector Laboratories, No.
SK-4100) staining for 5 min at room temperature to visualize nuclei
and were sealed beneath a coverslip. Finally, images were captured
with a light microscope.
Immunofluorescence staining
HK-2 cells (5×104 to 1×105) in
the logarithmic growth phase were lifted and centrifuged (1,000 × g
for 5 min at room temperature), and the cell density was adjusted
for inoculation in 12-well plates at 37°C with 5% CO2
overnight, with three replicate wells prepared for each group.
Cells were fixed with 4% PFA for 30 min at room temperature,
followed by permeabilization with 0.1% (v/v) Triton X-100
(Sigma-Aldrich, No. T8787) in PBS at room temperature for 10 min to
disrupt the cell membrane. Following permeabilization, samples were
blocked with 5% (w/v) BSA in PBS containing 0.1% Tween-20 at room
temperature for 60 min. Then cells were incubated with primary
antibodies (Nrf2 (Abcam, ab313825) 1:500) overnight. Samples were
washed to remove excess primary antibodies 4°C overnight and
incubated with secondary antibodies (Goat Anti-Rabbit IgG H&L
(HRP; Abcam, ab6721) 1:2,000) for 2 h at room temperature.
Following this incubation, secondary antibodies were washed off and
cells were incubated for 15 min with DAPI staining solution at room
temperature. Samples were washed again and images of cells were
captured using a fluorescence microscope.
Cell counting kit-8 (CCK-8) assay
To assess the viability of HK-2 cells, a CCK-8 assay
was performed (Beyotime Biotechnology) in accordance with the
manufacturer's instructions. HK-2 cells (5,000 cells) were seeded
into 96-well plates, which were incubated for 48 h. Subsequently,
10 µl CCK-8 reagent was introduced into each well and the plates
were incubated for a duration of 2 h. The absorbance of samples was
then quantified at a wavelength of 450 nm using a microplate reader
(SMR60047 Smart Microplate Reader, manufactured by USCNK Life
Science Co. Ltd.). All incubation steps were performed at 37°C with
5% CO2.
Cell apoptosis
HK-2 cells were harvested following centrifugation
at 300 × g for 5 min at 4°C, washed twice with cold PBS and
resuspended in 1X binding buffer at a density of 1×106
cells/ml. Subsequently, 100 µl cell suspension was incubated with 5
µl annexin V-FITC and 5 µl propidium iodide in the dark for 15 min
at room temperature. Samples were analyzed immediately using a BD
FACSCanto™ II flow cytometer (BD Biosciences) and using
FlowJo v10.8.1 (BD Biosciences, USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
RT-qPCR was utilized to analyze the mRNA expression
of NRF2. TRIzol (Thermo Fisher Scientific, Inc.) was used to
extract RNA from HK-2 cells and cDNA was synthesized utilizing the
PrimeScript™ RT Reagent Kit (Takara Bio Inc.), according
to the manufacturer's instructions. qPCR was conducted using the
SYBR-Green PCR Master Mix (Applied Biosystems; Thermo Fisher
Scientific, Inc.) on a GeneAmp® PCR System 9700 (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The primer sequences
used were as follows: NRF2, forward 5′-GCCACCGCCAGGACTACAG-3′,
reverse 5′-GCAACAAGAGCAGCCACCTC-3′; and GAPDH, forward
5′-CCCTCGTCCCGTAGACAAAATG-3′, reverse 5′-TGAGGTCAATGAAGGGGTCGT-3′.
The thermocycling conditions for the qPCR reaction included an
initial denaturation step at 95°C for 30 sec, followed by 40 cycles
of denaturation at 95°C for 10 sec and annealing and extension at
60°C for 10 sec. GAPDH served as the internal control, and the
relative expression levels of NRF2 were assessed using the
2−ΔΔCq analysis method (29).
Western blot analysis
Total protein was extracted from HK-2 cells and
mouse renal tissues using RIPA buffer (Beyotime Biotechnology) in
the presence of protease inhibitors. A BCA Protein Assay Kit
(Beyotime Biotechnology) was used to determine the total protein
content of the samples. Proteins (20–30 µg/lane) were separated
using 10% SDS-PAGE and subsequently transferred via electrophoresis
onto a polyvinylidene fluoride membrane. Membranes were blocked for
1 h with 5% skimmed milk powder in Tris-buffered saline + 20%
Tween-20 (TBST) at room temperature. Membranes were incubated with
primary antibodies (KIM-1 (Proteintech, 30948-1-AP) 1:500, NGAL
(Proteintech, 31721-1-AP) 1:500, Bax (Proteintech, 50599-2-Ig)
1:500, Bcl-2 (all Proteintech, 68103-1-Ig) 1:500, Cleaved caspase-3
(Abcam, ab214430), Caspase-3 (Abcam, ab184787) 1:500, NRF2 (Abcam,
ab313825) 1:500, GAPDH (all Abcam, ab181602; all 1:500) overnight
at 4°C. Membranes were washed with TBST solution and treated with
HRP-conjugated goat anti-rabbit secondary antibodies for 1 h at
room temperature. Protein bands were visualized utilizing an ECL
reagent (Thermo Fisher Scientific, Inc.), and analyzed by ImageJ
(Version 1.54, developed by the National Institutes of Health).
Statistical analysis
All experimental data are presented as the mean ±
standard deviation of three independent experimental repeats. Data
were analyzed using GraphPad Prism 7.0 (Dotmatics). Data were
analyzed using unpaired t-test or one-way ANOVA followed by Tukey's
post hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
SFN protects against CI-AKI in
mice
To explore the role of SFN in the development and
progression of CI-AKI, the present study established a mouse model
of CI-AKI, and mice were separated into groups for treatment with
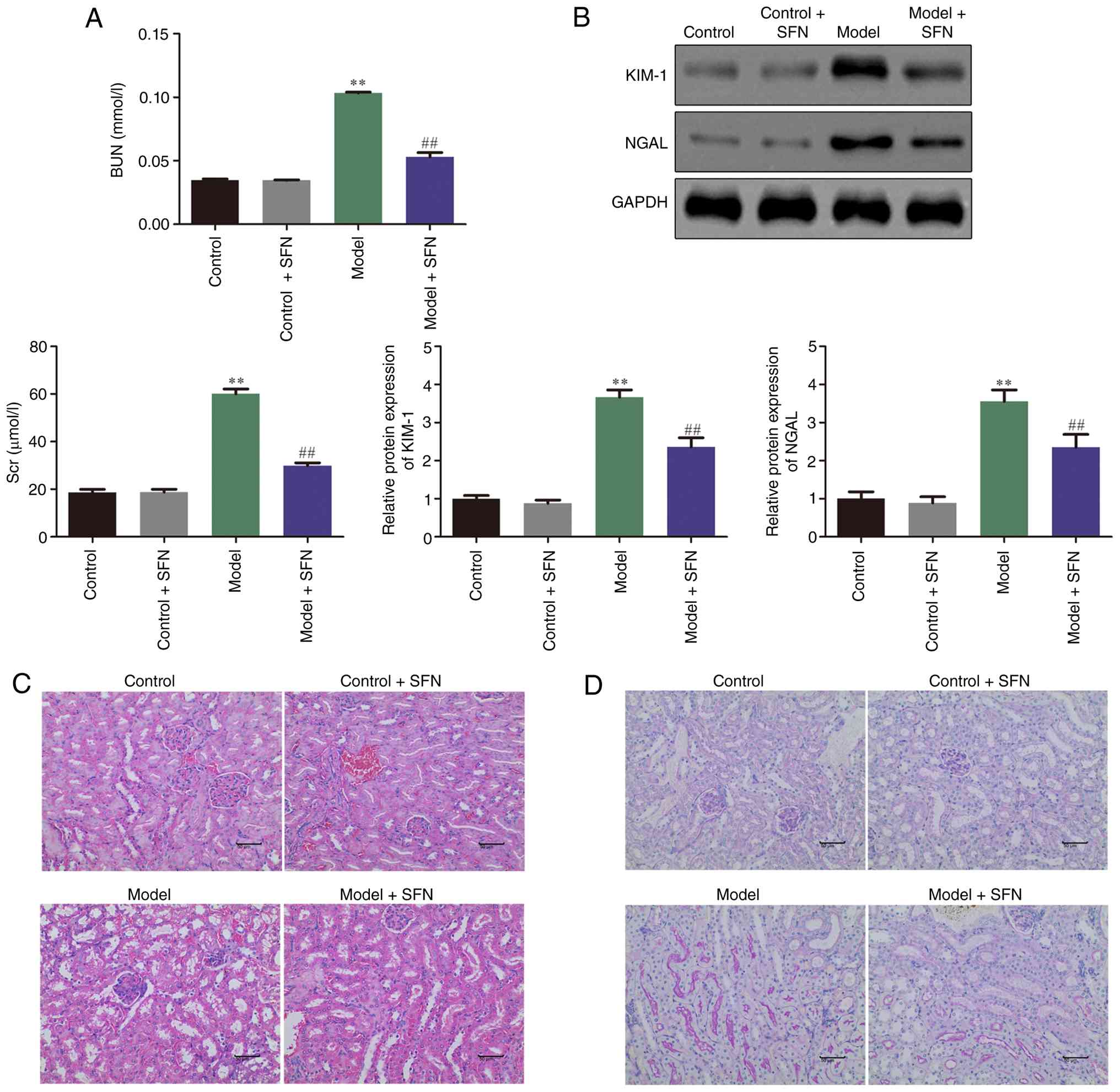
or without SFN. As shown in Fig.
1A, the levels of Scr and BUN in the CI-AKI model group were
significantly higher than the control group, whereas SFN treatment
significantly attenuated the heightened levels of Scr and BUN.
Mouse kidney markers were measured to determine the extent of
pathological damage to the kidneys in different treatment groups.
The protein expression of kidney injury molecule-1 (KIM-1) and
neutrophil gelatinase-associated lipocalin (NGAL) was significantly
increased in the model group compared with the control group,
whereas SFN significantly reduced the expression of KIM-1 and NGAL
in the model + SFN group compared with the model group (Fig. 1B). Furthermore, H&E and PAS
staining were performed to investigate whether SFN improved the
renal pathology of CI-AKI in model mice. As demonstrated in
Fig. 1C and D, the results of both
H&E and PAS staining indicated that epithelial cells in the
control group exhibited intact morphology and were arranged neatly,
whereas the renal pathological structure of cisplatin-treated model
mice demonstrated notable damage, such as irregular morphology and
detachment of renal tubular epithelial cells, marked dilation of
renal tubules and vacuolar degeneration of renal tubules. Compared
with the model group, kidney damage after prophylactic
administration of SFN was markedly reduced. Furthermore,
administration of SFN to control mice showed no effects on Scr and
BUN, KIM-1 and NGAL and renal pathology (Fig. 1A-D) compared with the control
group. These findings indicated that SFN ameliorated CI-AKI in
mice.
SFN inhibits cisplatin-induced
apoptosis, inflammation and oxidative stress
Apoptosis, inflammation and oxidative stress serve
important roles in the progression of CI-AKI (30). As shown in Fig. 2A, the number of TUNEL-positive
cells in the renal tubular lumen of cisplatin-treated model mice
was notably increased compared with the control group, and
treatment with SFN markedly reduced the apoptosis of renal tubular
cells in model mice. Furthermore, the levels of apoptosis-related
proteins, namely Bax, Bcl-2, cleaved caspase-3 and caspase-3, were
detected via western blot assay. Compared with the control group,
the expression of the pro-apoptotic protein Bax and the ratio of
cleaved caspase-3/caspase-3 in the kidneys of mice treated with
cisplatin were significantly increased, whereas the expression
level of the anti-apoptotic protein Bcl-2 significantly decreased.
SFN treatment in model mice significantly alleviated these changes
(Fig. 2B). Furthermore, the levels
of inflammatory indicators, such as F4/80, IL-1β, IL-6 and TNF-α,
were measured via IHC and ELISA. As shown in the representative
immunohistochemistry images in Fig.
2C, F4/80 levels were markedly increased in the model group
compared with the control group. However, treatment of model mice
with SFN resulted in a notable decrease in F4/80 levels compared
with untreated model mice. Similarly, IL-1β, IL-6 and TNF-α were
significantly upregulated in the model compared with the control
group; however, these levels were significantly reduced in the
model + SFN group compared with the model group (Fig. 2D). Furthermore, DCFH-DA staining
was used to visualize ROS levels in each treatment group. Staining
results revealed that ROS levels were markedly increased in the
model group compared with the control group. However, ROS levels
were notably attenuated in the model + SFN group compared with the
model group (Fig. 2E). Similarly,
biochemical analysis was utilized to evaluate the expression levels
of MDA, SOD and GSH. As demonstrated in Fig. 2F, compared with the control group,
MDA in the model group was significantly upregulated, and the
levels of SOD and GSH were significantly decreased. However,
compared with the model group, SFN treatment significantly
mitigated MDA levels and partially yet significantly restored SOD
and GSH levels. These findings indicated that SFN inhibited
cisplatin-induced apoptosis, inflammation and oxidative stress.
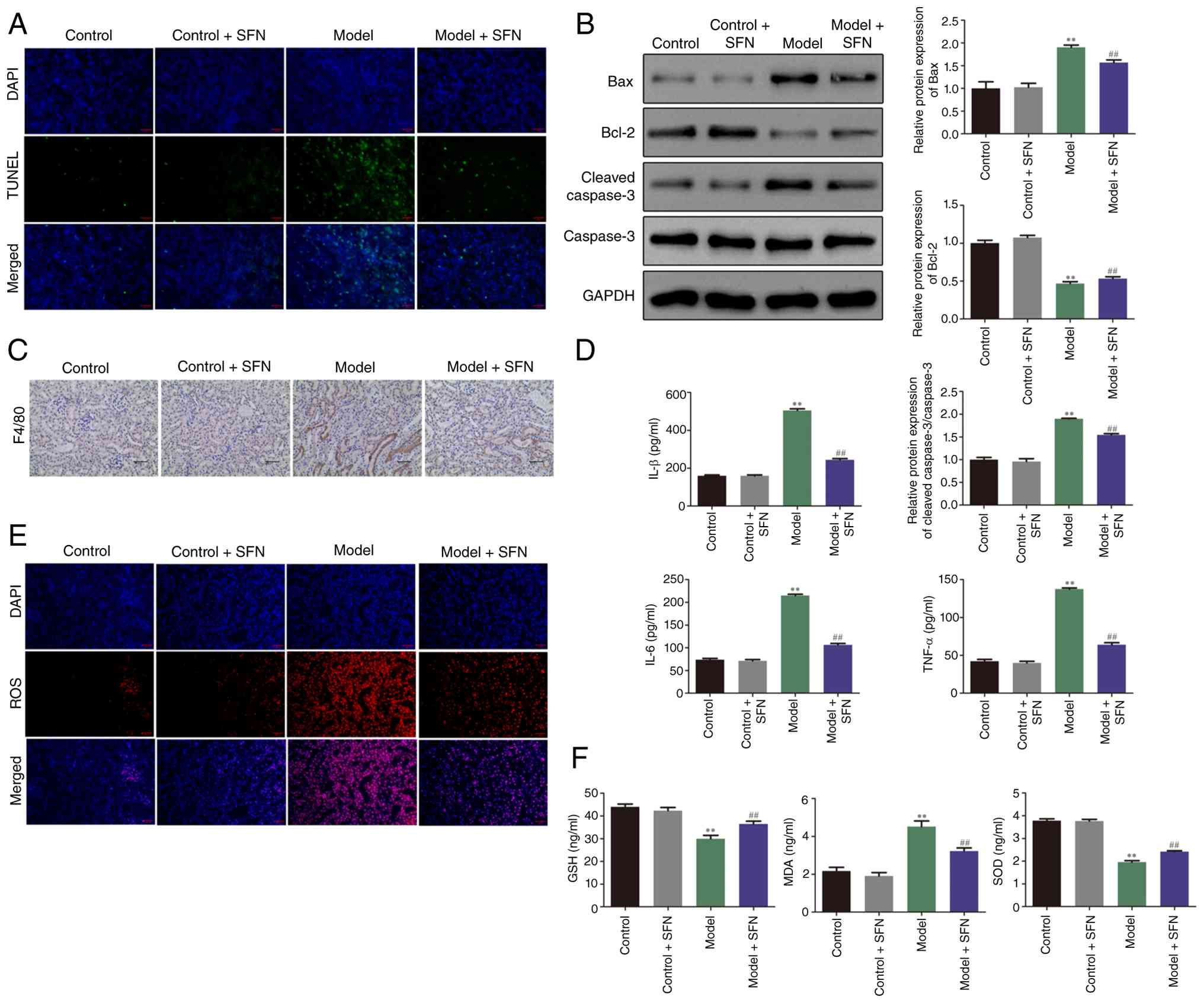 | Figure 2.SFN inhibits cisplatin-induced
apoptosis, inflammation and oxidative stress. (A) TUNEL staining
was used to determine the number of apoptotic TUNEL-positive cells
in the renal tubular lumen of mice in each treatment group.
Magnification, ×10 magnification). (B) Expression levels of the
apoptosis-related proteins Bax, Bcl-2, cleaved caspase-3 and
caspase-3 in mice were detected via western blotting. (C)
Representative images of immunohistochemical analysis showing F4/80
expression in control and model mice treated with or without SFN
(×20 magnification). (D) ELISA was performed to evaluate the
expression of IL-1β, IL-6 and TNF-α in mice across treatment
groups. (E) 2′,7′-dichlorodihydrofluorescein diacetate staining was
performed to evaluate the expression of ROS in mice in each
treatment group (×10 magnification). (F) Biochemical analysis was
used to evaluate the expression levels of MDA, SOD and GSH of mice
in each group. **P<0.01 vs. control group;
##P<0.01 vs. model group. SFN, sulforaphane; IL,
interleukin; TNF-α, tumor necrosis factor-α; MDA, malondialdehyde;
SOD, superoxide dismutase; GSH, glutathione; ROS, reactive oxygen
species. |
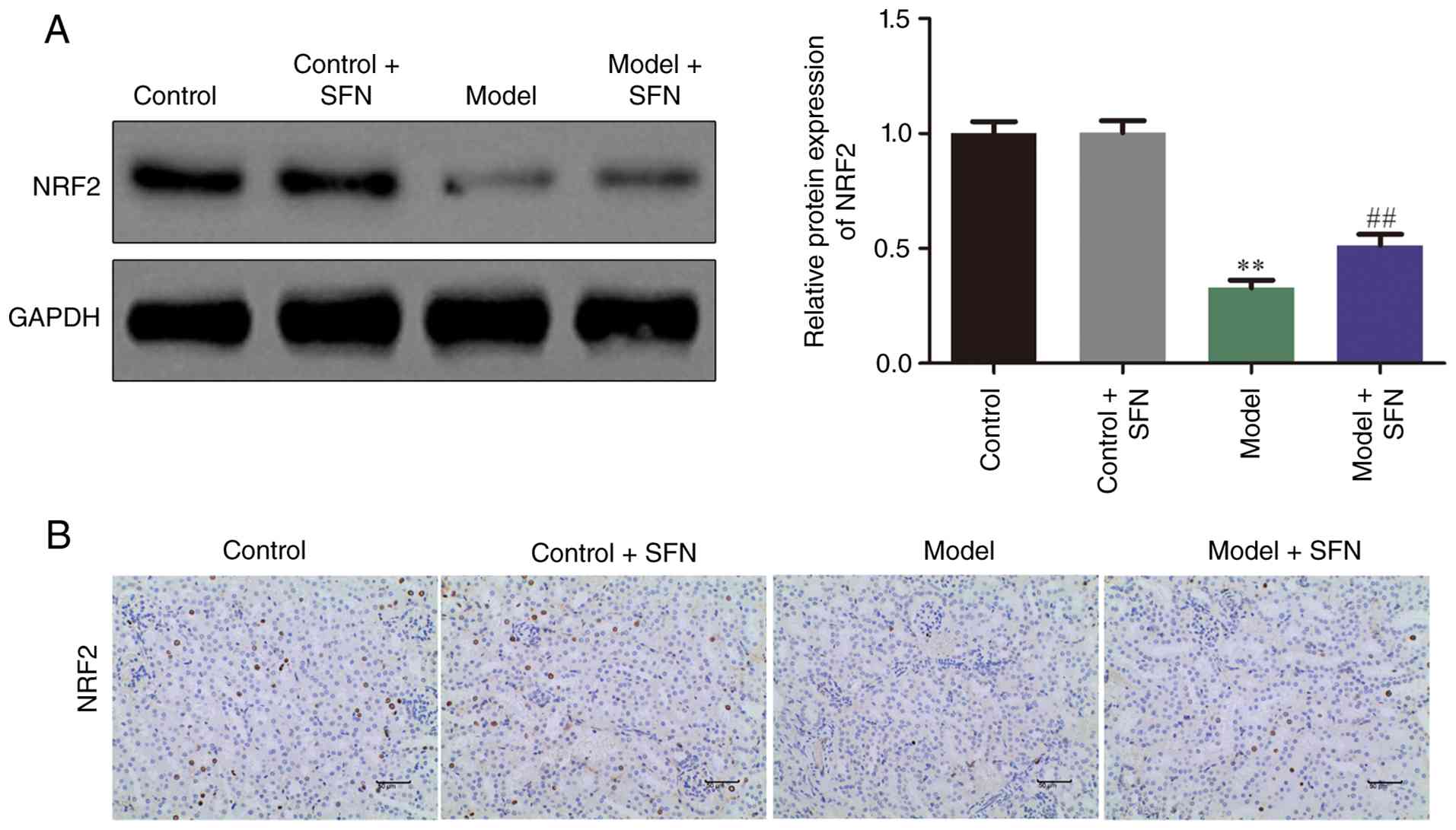
SFN activates the NRF2 signaling
pathway
The NRF2 signaling pathway, as one of the major
signal transduction pathways in CI-AKI, plays an important role in
the progression and development of CI-AKI (31). IHC and western blotting were
performed to determine the effects of SFN on the expression of
NRF2. As shown in Fig. 3, SFN
upregulated the expression of NRF2 signaling pathway in CI-AKI mice
compared with untreated model mice. These findings indicated that
SFN ameliorated CI-AKI in mice by upregulating NRF2.
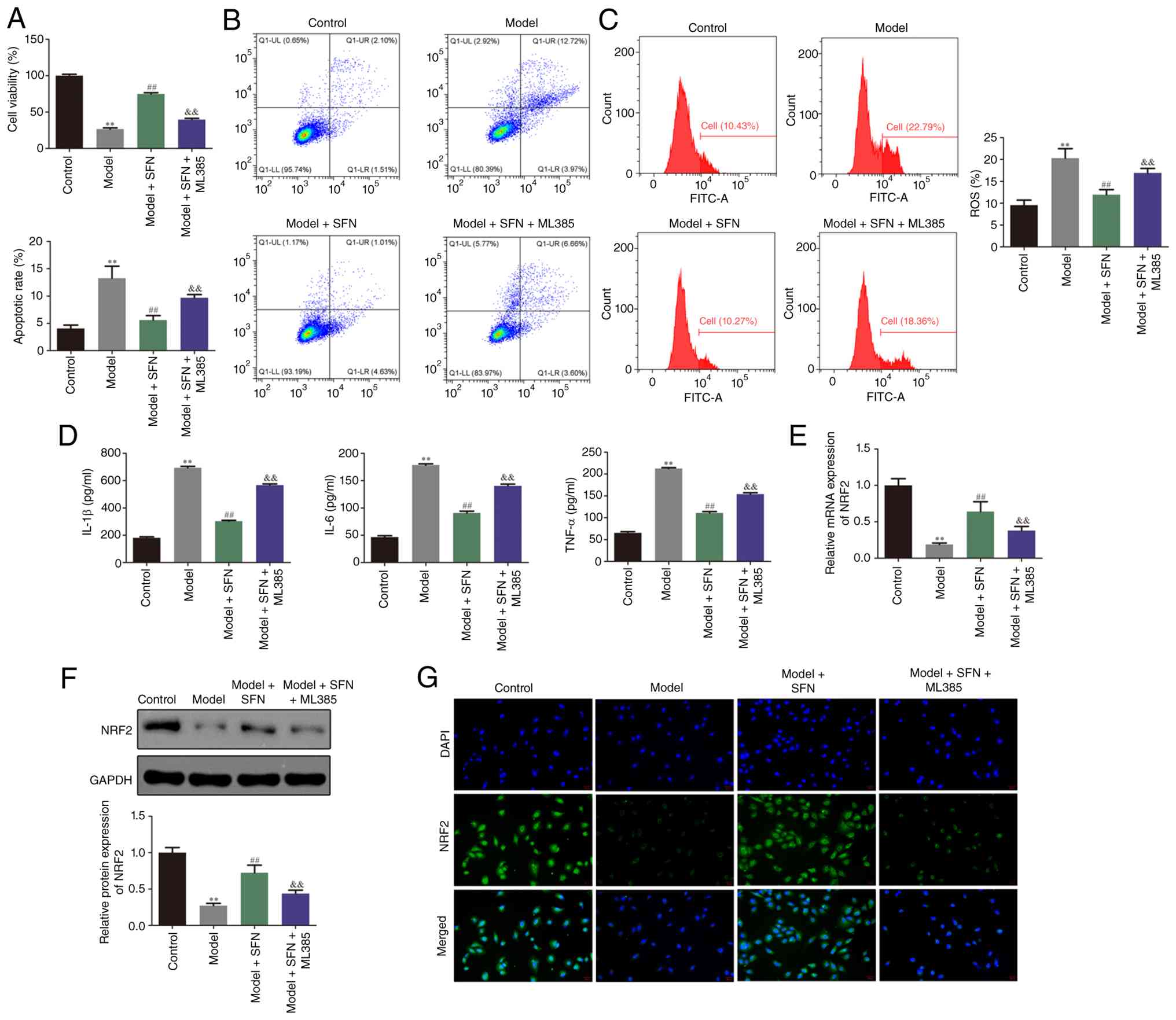
SFN protects HK-2 cells against
cisplatin-induced apoptosis, inflammation and oxidative stress in
vitro partially by regulating the NRF2 signaling pathway
To further investigate the protective effect and
mechanisms of SFN in HK-2 cells in vitro, the present study
evaluated changes in cell apoptosis, inflammation and oxidative
stress. CCK-8 assay indicated that, compared with the control
group, cisplatin significantly inhibited the viability of HK-2
cells, whereas treatment of cisplatin-induced cells with SFN
mitigated this effect (Fig. 4A).
Flow cytometry, DCFH-DA staining and ELISA demonstrated that
cisplatin significantly promoted apoptosis, inflammation and
oxidative stress in HK-2 cells relative to the control group.
Notably, treatment of model cells with SFN significantly mitigated
cisplatin-induced apoptosis, inflammation and oxidative stress
(Fig. 4B-D). Furthermore, the
present study performed RT-qPCR, western blotting and
immunofluorescence staining to assess the expression of NRF2. As
indicated in Fig. 4E-G, treatment
with SFN significantly upregulated NRF2 in cisplatin-induced HK-2
cells compared with the model group. Furthermore, co-treatment with
the NRF2 inhibitor ML385 partially yet significantly mitigated the
effects of SFN on the viability, apoptosis, inflammation and
oxidative stress of HK-2 model cells induced with cisplatin
(Fig. 4A-D). These findings
indicated that SFN protected HK-2 cells against cisplatin-induced
apoptosis, inflammation and oxidative stress in vitro via
regulation of the NRF2 signaling pathway.
Discussion
Cisplatin is one of the most widely used anticancer
drugs globally and is used in ~50% of chemotherapy regimens
(32). The kidney is the
predominant organ responsible for cisplatin metabolism, and the
concentration of cisplatin in the proximal renal tubules during
chemotherapy is ~5-fold that of the blood, therefore cisplatin is
prone to accumulate in the kidney (5). Upon accumulation, cisplatin generates
a large amount of ROS in the kidneys, which triggers oxidative
stress and inflammatory cascade reactions, ultimately leading to
kidney cell apoptosis or necrosis. This causes notable kidney
damage and limits the clinical application of cisplatin (33). Therefore, preventing or mitigating
CI-AKI is important for improving the clinical application of
cisplatin.
SFN is an isothiocyanate, which are products of the
myrosinase-mediated hydrolysis of glucosinolates found in
cruciferous vegetables, such as broccoli (34). It has been reported that SFN
exhibits anti-inflammatory, antioxidant, antitumor and
immunosuppressive activities and plays a therapeutic role in
diabetes nephropathy and psoriasis (35). A previous study demonstrated that
SFN prevents cell death and inflammation in diabetic nephropathy,
highlighting its broad renoprotective potential (36). However, to the best of our
knowledge, no in-depth research has been performed to elucidate
whether SFN exerts protective effects against CI-AKI. To the best
of our knowledge, the present study demonstrated for the first time
that SFN exhibited a protective effect in CI-AKI. The present study
explored the protective mechanisms of SFN from three aspects,
namely oxidative stress, inflammation and cell apoptosis, with a
specific focus on elucidating the role of the NRF2 signaling
pathway in mediating these effects.
Serum levels of BUN and Scr can reflect glomerular
filtration function. When renal function is impaired, the
concentrations of these biomarkers in the blood are notably
increased due to reduced excretion from the kidneys.
Histopathological examination can also reflect the survival status
of kidney cells (6). The results
of animal experiments in the present study showed that the
concentrations of BUN and Scr in the serum of mice treated with
cisplatin alone were increased compared with controls. H&E and
PAS staining of renal tissues showed that cisplatin induced
pathological changes. The levels of renal function indicators in
model mice treated with SFN were significantly reduced compared
with untreated model mice and demonstrated notable improvements to
the pathological damage of kidney tissue, indicating that SFN had a
protective effect on CI-AKI.
ROS, including peroxides, superoxide and hydroxyl
radicals, are important factors in cisplatin-induced oxidative
stress, inflammatory responses and cell apoptosis. MDA is an
important indicator for measuring lipid peroxidation and oxidative
damage, whereas SOD can inhibit lipid peroxidation by clearing
superoxide anion radicals in the body (37). GSH is one of the most important
non-enzymatic antioxidants, demonstrating multiple functions,
including the clearance of free radicals, detoxification and the
maintenance cell immunity (38).
The excessive production of ROS leads to the excessive consumption
of antioxidants, including SOD and GSH, resulting in lipid
peroxidation and the generation of excess MDA, resulting in cell or
tissue damage (39). Therefore,
the combined detection of ROS, MDA, SOD and GSH was used to
determine the degree of lipid peroxidation damage in mice.
In addition, ROS promote the production of
pro-inflammatory cytokines, for example TNF-α, IL-6 and IL-1β,
inducing a cascade of inflammatory reactions and exacerbating
kidney damage. As ROS levels increase, lipids, proteins and nucleic
acids in renal cells become damaged, leading to cell apoptosis or
necrosis (40). The in vivo
experiments of the present study demonstrated that the levels of
ROS, MDA, TNF-α, IL-6 and IL-1β were significantly increased in
cisplatin-induced mice, whereas the levels of SOD and GSH were
significantly reduced. In addition, cisplatin notably induced the
apoptosis of renal tubular cells. Intervention with SFN was shown
to significantly attenuate the aforementioned pathological changes
in model mice.
In order to further investigate the effects of SFN
on cisplatin-induced oxidative stress, inflammatory responses and
cell apoptosis, the present study constructed an in vitro
model of kidney injury using HK-2 cells. Flow cytometry was used to
detect ROS levels in HK-2 cells, and the present study found that
following cisplatin treatment, ROS levels in HK-2 cells were
significantly increased. However, following intervention with SFN,
ROS levels were significantly decreased, indicating that SFN
alleviated oxidative stress caused by cisplatin. Furthermore, ELISA
was used to detect changes in the levels of inflammatory factors,
namely TNF-α, IL-6 and IL-1β. After cisplatin treatment for model
establishment, TNF-α, IL-6 and IL-1β were significantly
upregulated, whereas concurrent SFN intervention was significantly
attenuate these heightened TNF-α, IL-6 and IL-1β levels, indicating
that SFN alleviated inflammatory responses caused by cisplatin.
Additionally, the present study evaluated the effects of SFN on the
apoptosis of HK-2 cells and found that the number of apoptotic
cells significantly increased following cisplatin treatment.
Notably, the number of apoptotic cells was significantly reduced
following SFN intervention of the model group, suggesting that SFN
inhibited cisplatin-induced cell apoptosis. The aforementioned
in vitro experimental results were consistent with the
results of in vivo experiments, supporting the implication
that the protective effects of SFN on CI-AKI were achieved by
inhibiting oxidative stress, the inflammatory response and cell
apoptosis.
NRF2 is a transcription factor that plays a
protective role in oxidative damage (41). Activation of NRF2 is closely
related to a decrease in ROS levels and an increase in antioxidant
enzyme activity, and that NRF2 plays an important regulatory role
in inhibiting oxidative stress and inflammatory responses to
protect renal function (42).
Under normal conditions, NRF2 binds to Kelch-like ECH-associated
protein 1 (Keap1) in the cytoplasm. However, under conditions of
oxidative stress, NRF2 dissociates from Keap1 and enters the
nucleus, activating downstream antioxidant proteins, such as heme
oxygenase-1 and NAD(P)H quinone dehydrogenase 1. This helps to
maintain the dynamic balance of intracellular redox reactions,
thereby protecting the body from damage caused by oxidative stress
(43). Concurrently, activation of
NRF2 can also inhibit the production of pro-inflammatory cytokines
and alleviate the inflammatory response (44). In addition, NRF2 inhibits the
activation of apoptotic pathways by suppressing oxidative stress.
Under conditions of oxidative stress, increases in ROS levels
disrupt the dynamic balance of mitochondrial integrity and enhance
the activity of the apoptotic pathway by activating various
signaling pathways, thereby inducing cell apoptosis (45).
In the present study, SFN intervention significantly
upregulated NRF2, indicating that SFN may have alleviated
cisplatin-induced oxidative stress, inflammatory responses and
apoptosis by activating the NRF2 signaling pathway. To further
support the direct regulatory role of the NRF2 signaling pathway in
alleviating CI-AKI, the present study treated HK-2 cells with the
NRF2 inhibitor ML385 to observe whether the regulatory effect of
SFN on CI-AKI pathology was reversed. The results demonstrated that
ML385 partially reversed the protective effects of SFN on oxidative
stress, inflammatory responses and apoptosis in model HK-2 cells,
suggesting that the renoprotective effect of SFN was partially
regulated by activation of the NRF2 signaling pathway.
Although the present study was, to the best of our
knowledge, the first study to demonstrate that SFN partially
alleviated CI-AKI via activation of the NRF2 signaling pathway and
provide evidence supporting its preventive administration, the
present study exhibited several key limitations. In terms of
experimental design, both animal and cell models employed a strict
pre-treatment regimen involving a single dose of cisplatin and SFN,
preventing a sufficient dose-response relationship analysis.
Furthermore, the present study primarily evaluated the preventive
potential of SFN and failed to simulate more common clinical
intervention scenarios, such as post-injury or concurrent drug
administration, which limited the elucidation of an effective and
safe therapeutic window and the evaluation of SFN intervention
efficacy in broader clinical application scenarios. In terms of
mechanism verification, although the inhibitor ML385 experiment
provided functional evidence of the involvement of the NRF2
pathway, the present study could not rule out the possibility that
SFN may have employed other protective pathways that were not
dependent on NRF2. Furthermore, the in vitro findings of the
present study were based solely on the HK-2 cell line; future
studies should include other relevant renal cell types to confirm
the generalizability of the protective mechanisms. Therefore,
future research should: i) Systematically optimize SFN dosages; ii)
evaluate the efficacy of different SFN administration timings, such
as the effect of post-injury treatment on CI-AKI; and iii) employ
more in-depth molecular tools to verify the core role of NRF2 in
the SFN-mediated alleviation of CI-AKI and explore parallel
mechanisms of SFN-mediated renoprotection in order to promote its
clinical translation.
In summary, SFN exerted a protective effect on
CI-AKI and demonstrated potential as an adjuvant drug for cisplatin
treatment. The mechanisms underlying SFN-mediated protective
effects may have been related to activation of the NRF2 signaling
pathway, thereby mediating the inhibition of oxidative stress, the
inflammatory response and cell apoptosis.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science and Technology
Development Fund of Nanjing Medical University (grant nos.
NMUB20220164 and NMUB20220057) and the Youth Fund of Nanjing
Jiangning Hospital (grant no. JNYYZXKY202210).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
ZW, MeL, HX, WL and JQ conceived and designed the
study. LX, MiL and LS performed the experiments and analyzed data.
HX and WL analyzed and interpreted data and drafted the original
manuscript. ZW, MeL and JQ critically revised the manuscript. ZW
and JQ confirmed the authenticity of the raw data generated during
the study. All authors have read and approved the final version of
the manuscript.
Ethics approval and consent to
participate
The present study was reviewed and approved by the
Ethics Committee of Nanjing Medical University (approval no. IACUC
2024-0811).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wei J and Zhu L: The role of glutathione
peroxidase 4 in the progression, drug resistance, and targeted
therapy of non-small cell lung cancer. Oncol Res. 33:863–872. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sung HY, Han J, Ju W, Kang JL, Park AK and
Ahn JH: MBNL2 enhances cisplatin resistance by regulating apoptosis
in ovarian cancer cells. BMB Rep. 58:224–231. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Teixeira AR, Mata D, Ferreira H, Paiva A,
Pelayo MJ, Rafael C, Maurício J, Calisto R and Cassiano Neves M:
Short hydration regimen in cisplatin-based chemotherapy and its
impact on nephrotoxicity: A unicentric prospective study. Cureus.
17:e797742025.PubMed/NCBI
|
|
4
|
Sugino Y, Nishikawa T, Inaba S, Owa S,
Kato M, Higashi S, Sasaki T, Masui S, Nishikawa K, Nakamura A, et
al: Cisplatin-induced therapy-related myelodysplastic syndrome
during avelumab maintenance therapy for metastatic urothelial
carcinoma. Int Cancer Conf J. 14:73–78. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tao S, Qi Y, Gao J, Yuan H, Wang R, Shen
X, Wei G and Peng Z: Ameliorative effect of sipunculus nudus
hydrolysate on cisplatin-induced nephrotoxicity by mitigating
oxidative stress, inflammation and apoptosis. Mar Drugs.
23:1002025. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Park I, Kim S, Um YW, Kim HE, Lee JH, Kim
S, Kim P and Jo YH: Intravital imaging of peritubular
microcirculation impairment in cisplatin-induced acute kidney
injury. JCI Insight. 10:e1786892025. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen C, Wang W, Poklis JL, Li PL, Lichtman
AH, Gewirtz DA and Li N: Mitigation of cisplatin-induced acute
kidney injury through oral administration of fatty acid amide
hydrolase inhibitor PF-04457845. J Pharmacol Exp Ther.
392:1000322025. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bui AP, Pham TTM, Kim M, Park JH, Kim JI,
Seo JH, Jung J, Kim JY and Ha E: GLDC alleviates cisplatin-induced
apoptosis, cellular senescence, and production of reactive oxygen
species via regulating UCP1 in the kidney. Life Sci.
368:1235022025. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tian Z, Wu Y, Yi B, Li L, Liu Y, Zhang H
and Li A: ESCRT III-mediated lysosomal repair improve renal tubular
cell injury in cisplatin-induced AKI. Autophagy. 21:1927–1944.
2025. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gu R, Shen J, Zhang J, Mao J and Ye Q:
Revolutionizing autoimmune kidney disease treatment with chimeric
antigen receptor-T cell therapy. Research (Wash D C).
8:07122025.PubMed/NCBI
|
|
11
|
Oh CJ, Choi W, Lee HY, Lee IK, Kim MJ and
Jeon JH: Sodium phenylbutyrate attenuates cisplatin-induced acute
kidney injury through inhibition of pyruvate dehydrogenase kinase
4. Biomedicines. 12:28152024. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yang XP, Wang YJ, Xu YS, Peng S and Yuan
HF: Bergamottin pretreatment attenuates cisplatin-induced acute
kidney injury in mice by inhibiting BACE-1-mediated ferroptosis.
Ren Fail. 48:26418472026. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Airik M, Clayton K, Wipf P and Airik R:
JP4-039 mitigates cisplatin-induced acute kidney injury by
inhibiting oxidative stress and blocking apoptosis and ferroptosis
in mice. Antioxidants (Basel). 13:15342024. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang H, Deng Z, Wang Y, Zheng X, Zhou L,
Yan S, Wang Y, Dai Y, Kanwar YS, Chen F and Deng F: CHIP drives
proteasomal degradation of NUR77 to alleviate oxidative stress and
intrinsic apoptosis in cisplatin-induced nephropathy. Commun Biol.
7:14032024. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yin H, Yan Q, Li Y and Tang H:
Dihydromyricetin nanoparticles alleviate lipopolysaccharide-induced
acute kidney injury by decreasing inflammation and cell apoptosis
via the TLR4/NF-κB pathway. J Funct Biomater. 15:2492024.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ji Y, Zhao Z, Yang Y, Wang X, Qiao R, Yu
X, Gong X, Feng Z and Hong Q: Mechanisms underlying the impact of
interleukin family on acute kidney injury: Pathogenesis,
progression, and therapy. Research (Wash D C).
8:07382025.PubMed/NCBI
|
|
17
|
Zhang Y, Hu J, Zhang Y and Ci X:
Amentoflavone protects against cisplatin-induced acute kidney
injury by modulating Nrf2-mediated oxidative stress and ferroptosis
and partially by activating Nrf2-dependent PANoptosis. Front
pharmacol. 16:15080472025. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ramadan SA, Kamel EM, Ewais MA, Khowailed
AA, Hassanein EHM and Mahmoud AM: Flavonoids of Haloxylon
salicornicum (Rimth) prevent cisplatin-induced acute kidney injury
by modulating oxidative stress, inflammation, Nrf2, and SIRT1.
Environ Sci Pollut Res Int. 30:49197–49214. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu J, Wang Y, Qiao P, Ying Y, Lin S, Lu
F, Gao C, Li M, Yang B and Zhou H: Mechanisms of cisplatin-induced
acute kidney injury: The role of NRF2 in mitochondrial dysfunction
and metabolic reprogramming. Antioxidants (Basel). 14:7752025.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Adtani PN, Al-Bayati SAAF and Elsayed WS:
Sulforaphane from brassica oleracea induces apoptosis in oral
squamous carcinoma cells via p53 activation and mitochondrial
membrane potential dysfunction. Pharmaceuticals (Basel).
18:3932025. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Song F, Verheust Y, Sampers I and Raes K:
The stability of isothiocyanates in broccoli extract: Oxidation
from erucin to sulforaphane was discovered. Food Chem.
480:1438722025. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lv D, Chu L, Du Y, Li C, Bao N, Su Y, Wang
G, Zheng Y and Yu Y: Sulforaphane alleviates membranous nephropathy
by inhibiting oxidative stress-associated podocyte pyroptosis. Iran
J Basic Med Sci. 28:237–244. 2025.PubMed/NCBI
|
|
23
|
Josa E, Barril G and Ruperto M: Potential
effects of bioactive compounds of plant-based foods and medicinal
plants in chronic kidney disease and dialysis: A systematic review.
Nutrients. 16:43212024. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zaghlool SS, Abdelaal N, El-Shoura EAM,
Mahmoud NI and Ahmed YM: Restoring glomerular filtration rate by
sulforaphane modulates ERK1/2/JNK/p38MAPK, IRF3/iNOS, Nrf2/HO-1
signaling pathways against folic acid-induced acute renal injury in
rats. Int Immunopharmacol. 123:1107772023. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li Z, Guo H, Li J, Ma T, Zhou S, Zhang Z,
Miao L and Cai L: Sulforaphane prevents type 2 diabetes-induced
nephropathy via AMPK-mediated activation of lipid metabolic
pathways and Nrf2 antioxidative function. Clin Sci (Lond).
134:2469–2487. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pańczyszyn-Trzewik P, Stachowicz K,
Misztak P, Nowak G and Sowa-Kućma M: Repeated sulforaphane
treatment reverses depressive-like behavior and exerts antioxidant
effects in the olfactory bulbectomy model in mice. Pharmaceuticals
(Basel). 17:7622024. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu N, Mu R, Deng S, Han Y, Shi Y, Fu X, Li
H and Yao Q: Reserpine alleviates cisplatin-induced acute kidney
injury via anti-ferroptosis and cGAS/STING pathway. Ren Fai.
46:24063952024. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cai F, Li D, Zhou K, Zhang W and Yang Y:
Tiliroside attenuates acute kidney injury by inhibiting ferroptosis
through the disruption of NRF2-KEAP1 interaction. Phytomedicine.
126:1554072024. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zheng D, Ruan X, Wu Q, Qiu Y and Ruan S:
Yishen Jiangzhuo decoction attenuates cisplatin-induced acute
kidney injury by inhibiting inflammation, oxidative stress and
apoptosis through the TNF signal pathway. Exp Ther Med. 28:3312024.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhao L, Yue Z, Wang G, Qin J, Ma H, Tang D
and Yin G: Smilax glabra roxb. alleviates cisplatin-induced acute
kidney injury in mice by activating the Nrf2/HO-1 signalling
pathway. Phytomedicine. 139:1565502025. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pan L, Zhang D, Shao Q, Cheng M, Liao Z,
Yu L, Wang Y, Jia P and Zhang J: Panax notoginseng improves the
sensitivity of non-small cell lung cancer to cisplatin by
inhibiting Akt signaling. Cancer Biomark. 42:187585922413033772025.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Abd-Eldayem AM, Ali MF and Ahmed EA:
Nebivolol rescued the liver and kidney from the coadministration of
rivaroxaban and cisplatin by targeting inflammation, oxidative
stress, and apoptosis in rats. Int Immunopharmacol. 153:1144862025.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu Y and Zhang M: Synergistic anticancer
effects of silibinin and sulforaphane: Targeting gastric cancer via
PI3K/AKT and ERK1/2 MAPK pathway inhibition and molecular docking
insights. J Biochem Mol Toxicol. 39:e702372025. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhao L, Li J, Dang Y, Fisher D, Hien NTT,
Musabaev E, Pronyuk K and Zhao L: Protective role of sulforaphane
in lipid metabolism-related diseases. Mol Biol Rep. 52:2412025.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Guerrero-Beltrán CE, Mukhopadhyay P,
Horváth B, Rajesh M, Tapia E, García-Torres I, Pedraza-Chaverri J
and Pacher P: Sulforaphane, a natural constituent of broccoli,
prevents cell death and inflammation in nephropathy. J Nutr
Biochem. 23:494–500. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wu T, Ma W, Lu W, Huangshen Z, Chen S,
Yang Q, Li C, Li Z, Li N, Feng X, et al: Vaccarin alleviates
cisplatin-induced acute kidney injury via decreasing NOX4-derived
ROS. Heliyon. 9:e212312023. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ren Y, Wu F, Huo L, Wang X, Zhang Y, Fan
M, Tan M, Zhao J, Cheng J, Zhao Z and Bao J: Switchable ROS
generator and scavenger to prevent the cisplatin induced acute
kidney injury and improve efficacy via synergistic
chemodynamic/immune therapy. Mater Today Bio. 29:1013282024.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li P, Li D, Lu Y, Pan S, Cheng F, Li S,
Zhang X, Huo J, Liu D and Liu Z: GSTT1/GSTM1 deficiency aggravated
cisplatin-induced acute kidney injury via ROS-triggered
ferroptosis. Front Immunol. 15:14572302024. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Qi J, Luo Q, Zhang Q, Wu M, Zhang L, Qin
L, Xue Q and Nie X: Yi-Shen-Xie-Zhuo formula alleviates
cisplatin-induced AKI by regulating inflammation and apoptosis via
the cGAS/STING pathway. J Ethnopharmacol. 309:1163272023.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hu J, Zhang Y, Zhang Y, Shi N, Miu Y,
Huang J, Miao M and Ci X: Bergenin inhibits ferritinophagy and
ferroptosis in cisplatin-induced acute kidney injury by activating
the p-GSK3β/Nrf2/PPARγ pathway. Int Immunopharmacol.
147:1140042025. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Qi H, Shi H, Yan M, Zhao L, Yin Y, Tan X,
Qi H, Li H, Weng K, Tang Y and Dai Y: Ammonium tetrathiomolybdate
relieves oxidative stress in cisplatin-induced acute kidney injury
via NRF2 signaling pathway. Cell Death Discov. 9:2592023.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang S, Zheng Y, Jin S, Fu Y and Liu Y:
Dioscin protects against cisplatin-induced acute kidney injury by
reducing ferroptosis and apoptosis through activating Nrf2/HO-1
signaling. Antioxidants (Basel). 11:24432022. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sami DH, Soliman AS, Khowailed AA,
Hassanein EHM, Kamel EM and Mahmoud AM: 7-Hydroxycoumarin modulates
Nrf2/HO-1 and microRNA-34a/SIRT1 signaling and prevents
cisplatin-induced oxidative stress, inflammation, and kidney injury
in rats. Life Sci. 310:1211042022. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hu J, Gu W, Ma N, Fan X and Ci X:
Leonurine alleviates ferroptosis in cisplatin-induced acute kidney
injury by activating the Nrf2 signalling pathway. Br J Pharmacol.
179:3991–4009. 2022. View Article : Google Scholar : PubMed/NCBI
|