Introduction
As a key component in maintaining gut homeostasis,
the intestinal epithelial barrier permits selective nutrient uptake
while preventing the infiltration of luminal pathogens, toxins and
antigens. The functional integrity of this barrier relies on
epithelial cell viability, the proper organization of tight
junction proteins and the maintenance of intracellular redox
balance. Epidemiological evidence indicates that inflammatory bowel
disease affects ~4.9 million individuals globally, with a steadily
increasing incidence and prevalence over recent decades across both
high-income and newly industrialized regions (1,2).
Disruption of epithelial barrier integrity is widely recognized as
a key pathological feature in numerous gastrointestinal disorders,
including inflammatory bowel disease and oxidative
stress-associated mucosal injury (3,4).
Among the pathogenic mechanisms involved, oxidative stress serves
as a key contributor to epithelial dysfunction. Excessive
accumulation of reactive oxygen species (ROS) can directly damage
cellular macromolecules, compromise tight junction architecture and
amplify inflammatory responses, ultimately leading to barrier
breakdown (5,6). Although oxidative stress has been
increasingly acknowledged as an important therapeutic target
(3,7), effective strategies that directly
protect intestinal epithelial cells against oxidative
stress-mediated barrier impairment remain limited. Several
antioxidants, phytochemicals and microbiota-targeted interventions
have demonstrated protective effects in experimental models
(8,9); however, their clinical translation
remains constrained by poor bioavailability, insufficient target
specificity and limited robust evidence from human studies
(10,11).
Nuclear factor erythroid 2-related factor 2 (Nrf2)
functions as a central regulator of cellular antioxidant defense
and redox homeostasis. Upon oxidative challenge, Nrf2 undergoes
nuclear translocation, where it activates the transcription of
numerous cytoprotective genes, including heme oxygenase-1 (HO-1)
and NADPH quinone dehydrogenase 1 (NQO1). Increasing evidence over
the past decade has indicated that Nrf2 pathway activation not only
alleviates oxidative damage, but also contributes to the
preservation of epithelial integrity, modulation of inflammatory
responses and maintenance of gastrointestinal barrier function
(12,13). Concurrently, bioactive compounds
derived from natural sources have garnered marked attention as
modulators of oxidative stress due to their favorable safety
profiles and multitarget biological activities. Numerous studies
have demonstrated that plant-derived polyphenols and glycosides can
attenuate intestinal oxidative stress and improve epithelial
barrier function through regulation of Nrf2-dependent antioxidant
pathways (14,15).
Mulberroside A is a naturally occurring stilbene
glycoside isolated from Morus species and has been reported
to exhibit a broad spectrum of pharmacological properties,
including antioxidant and anti-inflammatory effects (16). Previous studies have shown that
mulberroside A may effectively reduce oxidative stress and confer
cytoprotection in cardiovascular, hepatic and neural cell models
(17–19). Emerging evidence has further
suggested that this compound may influence redox-sensitive
signaling pathways and antioxidant enzyme expression, implying its
potential regulatory effect upon Nrf2-associated mechanisms.
However, to the best of our knowledge, despite these findings, the
role of mulberroside A in intestinal epithelial oxidative stress
has not been systematically explored (12). In particular, its effects on
oxidative stress-induced intestinal barrier impairment and tight
junction integrity, as well as the associated molecular mechanisms,
remain largely undefined.
Therefore, the present study evaluated the
protective actions of mulberroside A on hydrogen peroxide
(H2O2)-evoked oxidative stress and barrier
dysfunction using Caco-2 intestinal epithelial cells. Its impact on
oxidative stress parameters, epithelial barrier integrity and
activation of the Nrf2-mediated antioxidant response were examined.
In addition, Nrf2 gene silencing was employed to clarify the
mechanistic contribution of Nrf2 signaling to the protective
actions of mulberroside A.
Materials and methods
Cell culture and reagents
Caco-2, the human colorectal adenocarcinoma cell
line, was obtained from the American Type Culture Collection (cat.
no. ATCC-HTB-37). Cell line authentication was provided by the
supplier and routine testing determined that the cells were
Mycoplasma-free. Mulberroside A (purity ≥98%) was purchased
from MedChemExpress (cat. no. HY-N0619). Mulberroside A was
dissolved in DMSO to prepare a stock solution and subsequently
diluted with culture medium to the indicated working
concentrations. The final DMSO concentration in all treatment
groups was maintained <0.1% (v/v). Caco-2 cells were cultured in
DMEM containing 10% FBS and 1% penicillin-streptomycin. Cells were
maintained at 37°C in 5% CO2. The medium was changed
every 2–3 days and cells at the logarithmic growth stage were
selected for subsequent experiments. All experiments were performed
using Caco-2 cells between passages 20–35 to ensure the formation
of a physiological intestinal epithelial barrier with
transepithelial electrical resistance values >50
Ωcm2, consistent with previously established criteria
for barrier-competent Caco-2 monolayers (20).
Establishment of oxidative stress
model and experimental grouping
To establish an in vitro model of oxidative
stress-mediated intestinal epithelial damage, Caco-2 cells were
treated with H2O2, which is commonly used to
induce acute oxidative damage in intestinal epithelial cells
(21). H2O2
was administered at 500 µM (final concentration), a condition
selected based on previous studies and preliminary experiments
(22).
To assess the effects of mulberroside A, cells were
initially treated with increasing concentrations of mulberroside A
(0, 5, 10, 20 and 40 µM) at 37°C for 24 h to evaluate cellular
responses. Based on the dose-screening results, three
concentrations, 5 µM (low), 10 µM (medium) and 20 µM (high), were
selected for subsequent experiments.
For functional and mechanistic analyses, cells were
assigned to the following groups: i) Control; ii)
H2O2; and iii) H2O2
combined with mulberroside A (5, 10 or 20 µM). In the mulberroside
A treatment groups, cells were pretreated with mulberroside A at
37°C for 30 min, followed by exposure to 500 µM
H2O2 for 6 h at 37°C in the continued
presence of mulberroside A. To further determine the involvement of
Nrf2 signaling, additional groups were established in which cells
were transfected with Nrf2-specific small interfering RNA (siNrf2)
or negative control siRNA (siNC) prior to mulberroside A
pretreatment and H2O2 exposure. Subsequently,
two independent siRNA sequences targeting Nrf2 were initially
evaluated for knockdown efficiency and the sequence exhibiting
higher efficiency was selected for further functional
experiments.
siRNA transfection
siRNAs targeting human Nrf2 and a corresponding
negative control siRNA were commercially synthesized by GenePharma
Co., Ltd. Transient siRNA transfection was carried out in Caco-2
cells using Lipofectamine™ RNAiMAX Transfection Reagent
(cat. no. 13778075; Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions. Briefly, cells were transfected
with siRNAs at a final concentration of 50 nM and incubated at 37°C
in a humidified atmosphere containing 5% CO2 for 48 h to
achieve effective knockdown of Nrf2 expression before subsequent
treatments. The efficiency of Nrf2 silencing was verified at the
mRNA level by reverse transcription-quantitative PCR (RT-qPCR)
using GAPDH as the internal control prior to functional and
molecular analyses. The detailed sequences of the siRNAs used are
provided in Table SI.
Cell viability assay
Caco-2 cells were plated in 96-well plates and
exposed to the designated treatments. After treatment, CCK-8
reagent (cat. no. CK04; Dojindo Molecular Technologies, Inc.) was
added to each well at a final concentration of 10% (v/v) and cells
were incubated under standard culture conditions for 2 h.
Absorbance at 450 nm was recorded with a microplate
spectrophotometer (SpectraMax® iD3; Molecular Devices,
LLC.). Cell viability was calculated as a percentage of the control
group.
Determination of intracellular
ROS
ROS levels were assessed with a fluorescent ROS
detection kit based on 2′,7′-dichlorofluorescein diacetate (cat.
no. S0033S; Beyotime Biotechnology). Following the indicated
treatments, Caco-2 cells were incubated with the fluorescent probe
at 37°C for 30 min under light-protected conditions, followed by
fluorescence microscopy. The fluorescence signal was analyzed as
integrated density using ImageJ software (version 1.53c; National
Institutes of Health). ROS levels were quantified as relative
integrated fluorescence values, and the results were normalized to
the control group.
Transepithelial electrical resistance
(TEER) measurement
Intestinal epithelial barrier integrity was assessed
by TEER. Caco-2 cells were seeded onto Corning®
Transwell® inserts (cat. no. 3413; Corning, Inc.) to
form confluent monolayers. TEER values were measured using an EVOM2
epithelial volt/ohm meter with STX2 electrodes (World Precision
Instruments), corrected by subtracting blank insert resistance and
normalized to the membrane surface area.
Western blotting
Using a nuclear/cytoplasmic protein extraction kit
(cat. no. P0027; Beyotime Biotechnology), nuclear and cytoplasmic
fractions were prepared and protein concentrations were quantified
using a Pierce™ BCA assay kit (cat. no. 23227; Thermo Fisher
Scientific, Inc.). After 30 µg of protein per sample was separated
by SDS-PAGE and transferred to PVDF membranes (cat. no. IPVH00010;
MilliporeSigma; Merck KGaA). Membranes were blocked with 5% non-fat
milk (Beyotime Biotechnology) at room temperature for 60 min.
Subsequently, membranes were incubated overnight at 4°C with the
following primary antibodies: Nrf2 (cat. no. ab62352; 1:1,000;
Abcam), HO-1 (cat. no. ab189491; 1:2,000; Abcam), NQO1 (cat. no.
ab80588; 1:10,000; Abcam), lamin B1 (cat. no. 12987-1-AP; 1:5,000;
Proteintech Group, Inc.) and GAPDH (cat. no. 81640-5-RR; 1:10,000;
Proteintech Group, Inc.). After washing three times with TBST (10
min each), membranes were incubated with HRP-conjugated secondary
antibodies at room temperature for 60 min: Goat anti-rabbit IgG
H&L/HRP (bs-0295G-HRP; 1:3,000; BIOSS) or goat anti-mouse IgG
H&L/HRP (bs-0296G-HRP; 1:3,000; BIOSS), depending on the
primary antibody host species. Signals were developed using
Pierce™ ECL reagent (cat. no. 32106; Thermo Fisher
Scientific, Inc.) and densitometry analysis was performed using
ImageJ software (version 1.53c; National Institutes of Health).
RT-qPCR
Total RNA was extracted from Caco-2 cells using
TRIzol™ reagent (cat. no. 15596026; Thermo Fisher Scientific,
Inc.). Complementary DNA was synthesized using the PrimeScript™
reverse transcription kit (cat. no. RR037A; Takara Bio, Inc.)
according to the manufacturer's instructions, with a reverse
transcription reaction at 37°C for 15 min followed by enzyme
inactivation at 85°C for 5 sec. Using SYBR Green Master Mix (cat.
no. RR420A; Takara Bio, Inc.), quantitative real-time PCR was
conducted on a QuantStudio 5 real-time PCR system (Applied
Biosystems). The thermocycling conditions were as follows: Initial
denaturation at 95°C for 30 sec, followed by 40 cycles of
denaturation at 95°C for 5 sec and annealing/extension at 60°C for
30 sec. A melt curve analysis was subsequently performed to verify
amplification specificity. Relative mRNA expression was determined
using the 2−ΔΔCq method (23) with GAPDH as the internal control.
The primer used sequences were as follows: Nrf2 forward,
5′-ACAAACATTCAAGCCGCTTGG-3′ and reverse,
5′-CGTAGCATGCTGAAAACTTCG-3′; GAPDH forward,
5′-AAGATCATCAGCAATGCCTCC-3′ and reverse,
5′-AGGTTTTTCTAGACGGCAGG-3′.
Measurement of malondialdehyde (MDA)
and superoxide dismutase (SOD) levels
Following the indicated treatments, Caco-2 cells
were lysed using Cell Lysis Buffer (cat. no. P0013; Beyotime
Biotechnology) according to the manufacturer's instructions. Cell
lysates were collected and used for the determination of lipid
peroxidation and antioxidant capacity by measuring MDA content and
total SOD activity using a Lipid Peroxidation (MDA) Assay Kit (cat.
no. S0131S; Beyotime Biotechnology) and a Total SOD Activity Assay
Kit (cat. no. S0101S; Beyotime Biotechnology), respectively,
according to the manufacturers' protocols. Absorbance was recorded
spectrophotometrically using a SpectraMax® iD3
microplate reader (Molecular Devices, LLC.) and MDA and SOD values
were normalized to total protein concentration.
Immunofluorescence staining
After treatment, Caco-2 cells cultured on glass
coverslips were fixed with 4% paraformaldehyde at room temperature
for 20 min, permeabilized and blocked with 5% bovine serum albumin
(cat. no. ST025; Beyotime Biotechnology) at room temperature for 30
min. Immunofluorescence staining was subsequently performed to
assess the expression and localization of tight junction proteins.
Cells were then incubated with primary antibodies against zonula
occludens 1 (ZO-1; cat. no. ab96587; 1:200; Abcam) and occludin
(cat. no. ab216327; 1:200; Abcam) at 4°C overnight. After washing
with PBS, cells were incubated with Alexa Fluor™
488-conjugated goat anti-rabbit IgG (H+L) secondary antibody (cat.
no. A-11008; 1:500; Thermo Fisher Scientific, Inc.) at room
temperature for 1 h in the dark. Nuclei were counterstained with
DAPI (cat. no. D9542; Sigma-Aldrich; Merck KGaA) at room
temperature for 10 min. Fluorescence images were acquired using the
IX73 fluorescence microscope (Olympus Corporation).
Statistical analysis
Each experiment was independently repeated at least
three times. Data are reported as the mean ± SD. Statistical
analyses were conducted using SPSS software (version 26.0; IBM
Corp.). Differences among multiple groups were analyzed using
one-way ANOVA tests followed by Tukey's post hoc test. P<0.05
was considered to indicate a statistically significant
difference.
Results
Impact of mulberroside A on Caco-2
cell viability
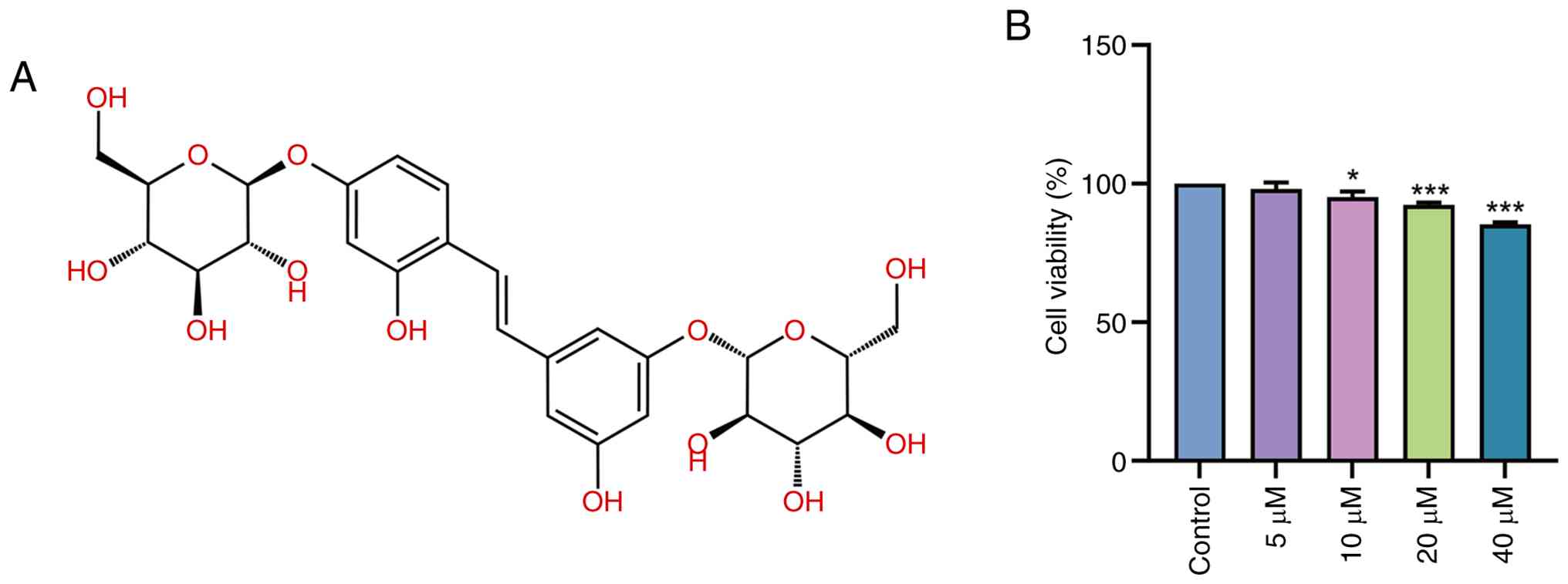
Fig. 1A shows the
chemical structure of mulberroside A, which was obtained from the
ChEMBL database (https://www.ebi.ac.uk/chembl/). Mulberroside A is a
stilbene glycoside composed of a central stilbene backbone bearing
numerous phenolic hydroxyl groups and glycosidic moieties. Based on
the CCK-8 assay, treatment of Caco-2 cells with 5 µM mulberroside A
for 24 h did not significantly affect cell viability compared with
the 0 µM (control) group (P>0.05; Fig. 1B). By contrast, treatment with 10
µM mulberroside A significantly decreased cell viability compared
with the control group (P<0.05). A further decline in viability
was observed in the 20 µM mulberroside A-treated group (P<0.001
vs. 0 µM), whereas treatment with 40 µM mulberroside A resulted in
a more pronounced reduction (P<0.001 vs. 0 µM). These findings
suggested that mulberroside A exerted limited effects on basal
Caco-2 cell viability at 5 µM, whereas higher concentrations tended
to reduce cell viability.
Effect of mulberroside A on
H2O2-induced oxidative stress and barrier
dysfunction in Caco-2 cells
As shown in Fig.
2A, H2O2 exposure significantly decreased
Caco-2 cell viability compared with the control group (P<0.001).
Pretreatment with mulberroside A at low, medium and high
concentration significantly restored cell viability compared with
the H2O2 group (all, P<0.001). Significant
differences in cell viability were observed among the mulberroside
A-treated groups (P<0.001), with the medium-dose group
exhibiting the greatest increase in viability.
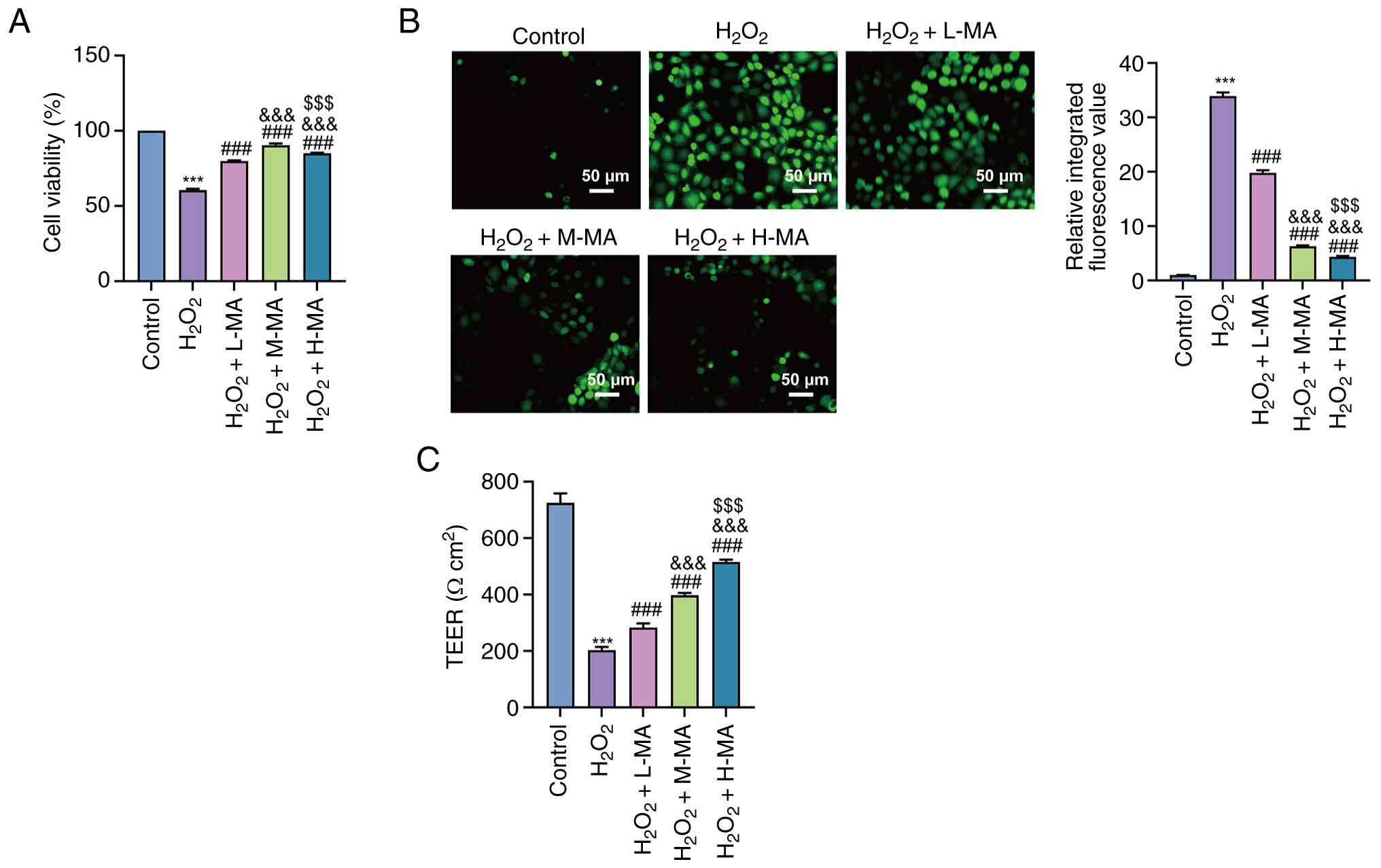 | Figure 2.Effect of MA on
H2O2-induced oxidative stress and barrier
dysfunction in Caco-2 cells. (A) Cell viability of Caco-2 cells in
the control, H2O2, H2O2
+ L-MA (5 µM), H2O2 + M-MA (10 µM) and
H2O2 + H-MA (20 µM) groups, as assessed using
a Cell Counting Kit-8 assay. (B) Intracellular reactive oxygen
species levels (representative fluorescence images shown; scale
bar, 50 µm). (C) TEER of Caco-2 cell monolayers following the
indicated treatments. Data are presented as the mean ± SD from
three independent experiments. ***P<0.001 vs. control,
###P<0.001 vs. H2O2,
&&&P<0.001 vs. H2O2
+ L-MA and $$$P<0.001 vs. H2O2
+ M-MA. H2O2, hydrogen peroxide; MA,
mulberroside A; L-MA, low-dose MA; M-MA, medium-dose MA; H-MA,
high-dose MA; TEER, transepithelial electrical resistance. |
Intracellular ROS levels were significantly
increased following H2O2 treatment compared
with the control group (P<0.001; Fig. 2B), whereas mulberroside A
significantly suppressed ROS accumulation at all tested
concentrations compared with the H2O2 group
(all, P<0.001). Significant differences were also observed among
the low-, medium- and high-dose mulberroside A groups (all,
P<0.001), demonstrating a concentration-dependent reduction in
ROS levels.
H2O2 exposure also caused a
significant reduction in TEER compared with the control group
(P<0.001; Fig. 2C).
Mulberroside A significantly elevated TEER values compared with the
H2O2 group at all tested concentrations (all,
P<0.001). TEER values progressively increased with increasing
mulberroside A concentrations, supporting a concentration-dependent
restoration of epithelial barrier integrity (P<0.001).
Effect of mulberroside A on
Nrf2/HO-1/NQO1 pathway protein levels in Caco-2 cells
As shown in Fig. 3A and
B, H2O2 treatment significantly decreased
nuclear Nrf2 protein levels compared with the control group
(P<0.001). Mulberroside A treatment at low, medium and high
concentrations significantly increased nuclear Nrf2 levels compared
with the H2O2 group (all, P<0.001). In
addition, significant inter-dose differences were observed among
the mulberroside A-treated groups (P<0.001), indicating a
concentration-dependent increase in nuclear Nrf2 accumulation.
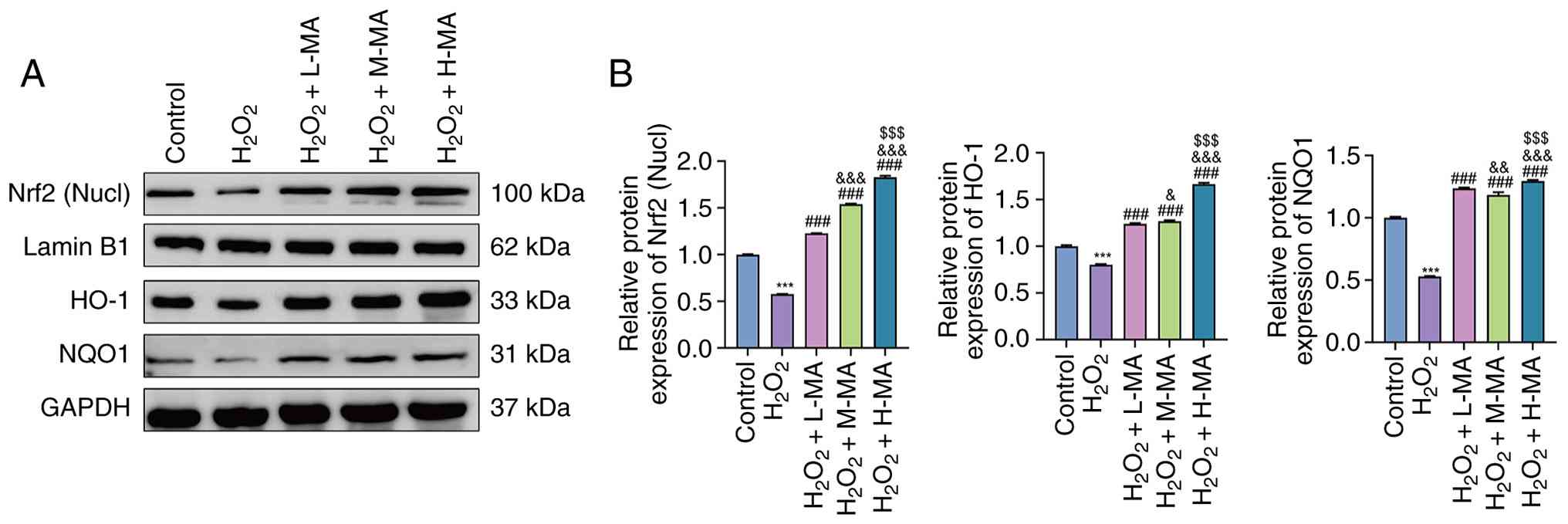 | Figure 3.Effect of MA on Nrf2/HO-1/NQO1
protein expression in Caco-2 cells. (A) Representative western
blotting of nucl Nrf2 and total HO-1 and NQO1 in control,
H2O2, H2O2 + L-MA,
H2O2 + M-MA and H2O2 +
H-MA groups. (B) Densitometric quantification normalized to the
corresponding internal controls. Data are presented as the mean ±
SD from three independent experiments. ***P<0.001 vs. control,
###P<0.001 vs. H2O2,
&P<0.05 vs. H2O2 + L-MA,
&&P<0.01 vs. H2O2 +
L-MA, &&&P<0.001 vs.
H2O2 + L-MA; and $$$P<0.001 vs.
H2O2 + M-MA. H2O2,
hydrogen peroxide; MA, mulberroside A; L-MA, low-dose MA; M-MA,
medium-dose MA; H-MA, high-dose MA; Nucl, nuclear; Nrf2, nuclear
factor erythroid 2-related factor 2; HO-1, heme oxygenase 1; NQO1,
NADPH quinone dehydrogenase 1. |
Similarly, total HO-1 protein levels were
significantly reduced following H2O2
treatment compared with the control group (P<0.001).
Mulberroside A treatment then significantly increased HO-1 protein
levels compared with the H2O2 group
(P<0.001) and HO-1 expression progressively increased with
increasing mulberroside A concentrations (P<0.05).
Consistent changes were observed for NQO1 protein
expression. H2O2 exposure significantly
decreased NQO1 levels compared with the control group (P<0.001),
whereas mulberroside A significantly restored NQO1 expression at
all tested concentrations compared with the
H2O2 group (all, P<0.001). Significant
differences among all mulberroside A dose groups were also detected
(P<0.01), further supporting a concentration-associated
activation of the Nrf2/HO-1/NQO1 antioxidant pathway.
Effect of Nrf2 knockdown on
mulberroside A-associated alterations in oxidative
stress-associated parameters in Caco-2 cells
As depicted in Fig.
4A, reverse transcription-quantitative PCR analysis
demonstrated that transfection with siNrf2#1 and siNrf2#2
significantly reduced Nrf2 mRNA levels in Caco-2 cells compared
with the siNC group (P<0.001). Based on knockdown efficiency,
siNrf2#2 was selected for subsequent experiments.
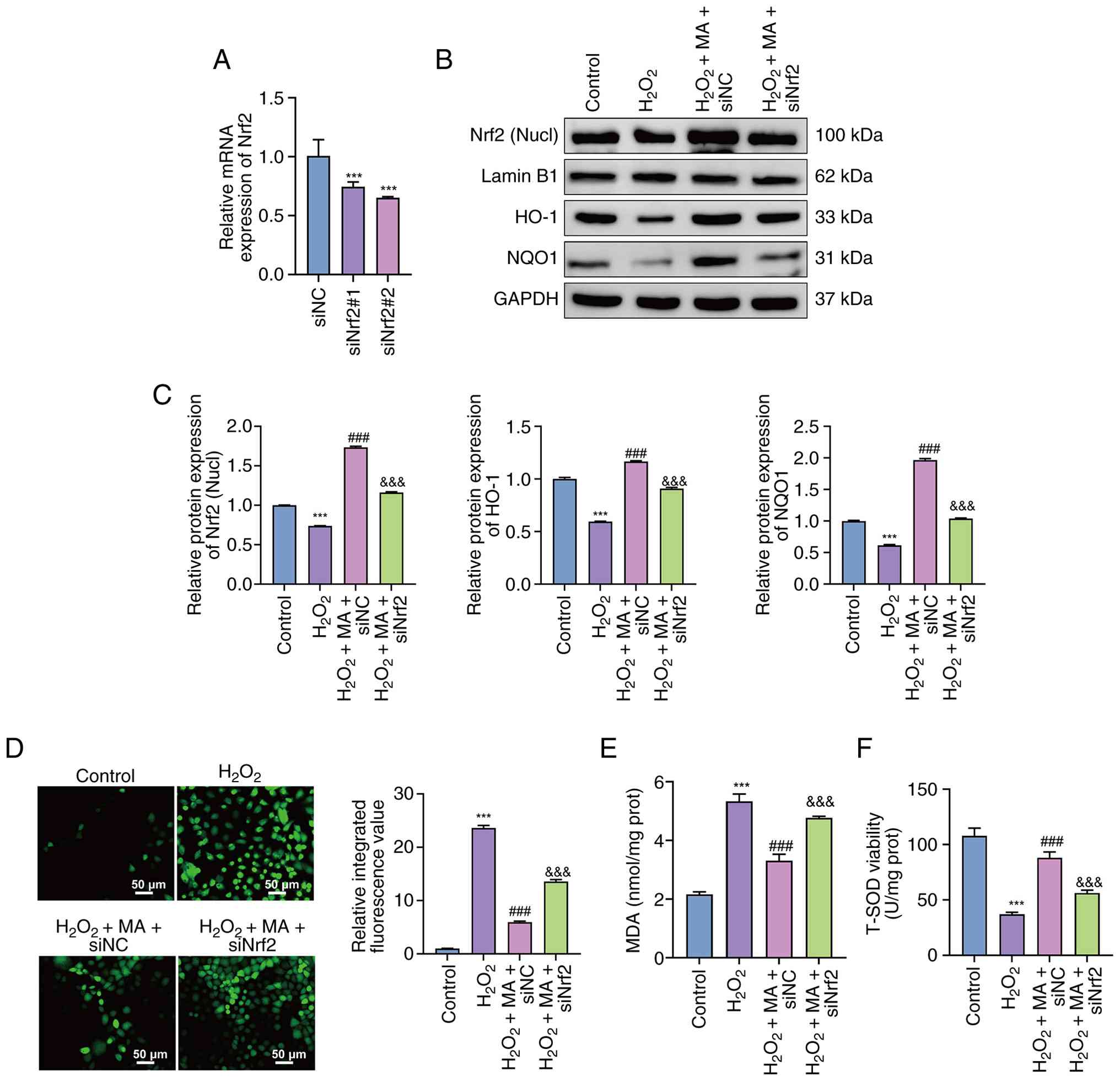 | Figure 4.Impact of Nrf2 knockdown on
MA-associated changes in oxidative stress-associated parameters in
Caco-2 cells. (A) Nrf2 mRNA levels after transfection with siNC,
siNrf2#1 or siNrf2#2, as determined by reverse
transcription-quantitative PCR. ***P<0.001 vs. siNC. (B)
Representative western blotting of nucl Nrf2 and total HO-1 and
NQO1 in control, H2O2,
H2O2 + MA + siNC and
H2O2 + MA + siNrf2 groups. (C) Densitometric
quantification normalized to internal controls. (D) Intracellular
reactive oxygen species levels, quantified as relative integrated
fluorescence intensity of DCF fluorescence images (representative
fluorescence images; scale bar, 50 µm). (E) MDA levels. (F) SOD
activity. Data are presented as the mean ± SD from three
independent experiments. ***P<0.001 vs. control,
###P<0.001 vs. H2O2 and
&&&P<0.001 vs. H2O2
+ MA+ siNC. H2O2, hydrogen peroxide; MA,
mulberroside A; L-MA, low-dose MA; M-MA, medium-dose MA; H-MA,
high-dose MA; Nucl, nuclear; Nrf2, nuclear factor erythroid
2-related factor 2; si, small interfering RNA; NC, negative
control; MDA, malondialdehyde; SOD, superoxide dismutase. |
Western blotting further demonstrated that
H2O2 treatment significantly reduced nuclear
Nrf2 and total HO-1 and NQO1 protein levels compared with the
control group (P<0.001; Fig. 4B and
C). In the H2O2 + mulberroside A + siNC
group, these proteins were significantly increased compared with
the H2O2 group (P<0.001). By contrast,
Nrf2 silencing significantly reduced nuclear Nrf2, HO-1 and NQO1
protein levels compared with the H2O2 +
mulberroside A + siNC group (all P<0.001).
Quantification of DCF fluorescence revealed that the
relative integrated fluorescence value was significantly increased
following H2O2 exposure compared with the
control group (P<0.001; Fig.
4D), whereas mulberroside A significantly reduced ROS
accumulation in siNC-transfected cells compared with the
H2O2 group (P<0.001). This inhibitory
effect was significantly attenuated by Nrf2 knockdown, as indicated
by higher ROS levels in the H2O2 +
mulberroside A + siNrf2 group compared with the
H2O2 + mulberroside A + siNC group
(P<0.001).
Consistently, H2O2
significantly increased MDA levels and decreased SOD activity
compared with the control group (both, P<0.001; Fig. 4E and F). Mulberroside A
significantly reduced MDA levels, but restored SOD activity in
siNC-transfected cells compared with the H2O2
group (both, P<0.001). However, Nrf2 knockdown significantly
weakened these effects, resulting in higher MDA levels and lower
SOD activity compared with the H2O2 +
mulberroside A + siNC group (both, P<0.001).
Effect of Nrf2 knockdown on epithelial
barrier integrity and tight junction protein expression in Caco-2
cells
As shown in Fig.
5A, H2O2 exposure significantly decreased
TEER compared with the control group (P<0.001). Mulberroside A
significantly restored TEER in siNC-transfected cells compared with
the H2O2 group (P<0.001). By contrast,
Nrf2 knockdown significantly reduced TEER compared with the
H2O2 + mulberroside A + siNC group
(P<0.001), indicating that Nrf2 silencing attenuated the
barrier-protective effect of mulberroside A.
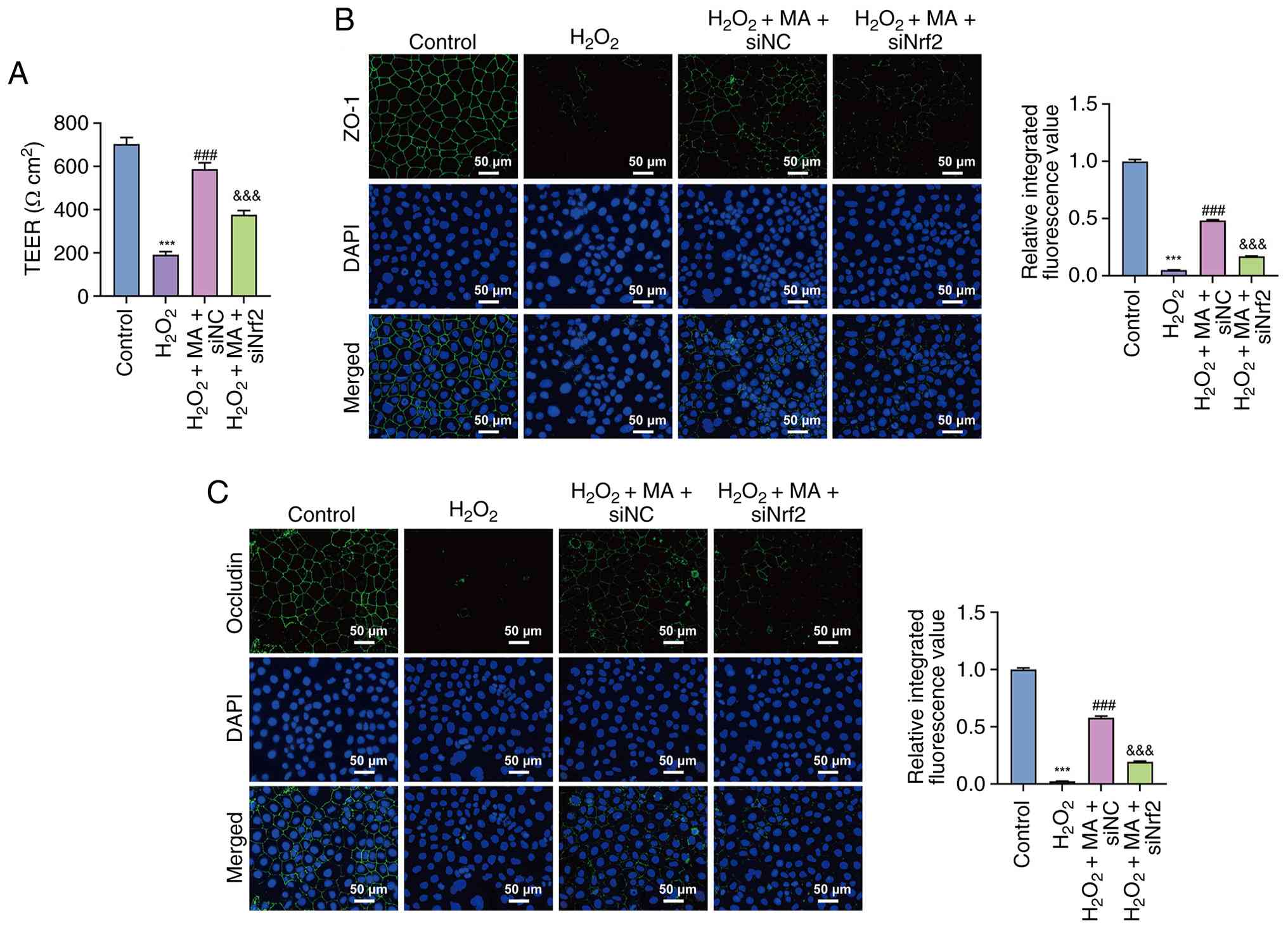 | Figure 5.Effects of Nrf2 knockdown on
epithelial barrier function and tight junction protein distribution
in Caco-2 cells. (A) TEER in control, H2O2,
H2O2 + MA + siNC and
H2O2 + MA + siNrf2 groups. Immunofluorescence
staining of (B) ZO-1 and (C) occludin. Data are presented as the
mean ± SD from three independent experiments. ***P<0.001 vs.
control, ###P<0.001 vs. H2O2
and &&&P<0.001 vs.
H2O2 + MA + siNC. H2O2,
hydrogen peroxide; MA, mulberroside A; L-MA, low-dose MA; M-MA,
medium-dose MA; H-MA, high-dose MA; Nucl, nuclear; Nrf2, nuclear
factor erythroid 2-related factor 2; si, small interfering RNA; NC,
negative control; TEER, transepithelial electrical resistance;
ZO-1, zonula occludens 1. |
Consistently, immunofluorescence analysis showed
that H2O2 markedly decreased ZO-1 and
occludin fluorescence intensities compared with the control group
(both, P<0.001; Fig. 5B and C).
Mulberroside A significantly increased both ZO-1 and occludin
signals in siNC-transfected cells compared with the
H2O2 group (both, P<0.001). However, Nrf2
knockdown significantly attenuated these increases, as reflected by
lower ZO-1 and occludin fluorescence intensities in the
H2O2 + mulberroside A + siNrf2 group compared
with the H2O2 + mulberroside A + siNC group
(both, P<0.001).
Discussion
In the present study, the impact of mulberroside A
on oxidative stress-induced intestinal epithelial injury was
systematically characterized using an
H2O2-challenged Caco-2 cell model. Exposure
to H2O2 resulted in notable oxidative injury
and epithelial barrier impairment, as evidenced by increased
intracellular ROS levels, reduced transepithelial electrical
resistance and disrupted expression of the tight junction proteins
ZO-1 and occludin. Within the concentration range examined,
mulberroside A markedly attenuated these alterations, accompanied
by enhanced nuclear accumulation of Nrf2 and upregulated expression
of the downstream antioxidant proteins HO-1 and NQO1. Notably,
genetic silencing of Nrf2 markedly weakened the mulberroside
A-associated changes in oxidative stress markers, barrier integrity
and tight junction protein expression. Collectively, these findings
revealed a coherent pattern showing an association between
oxidative stress modulation, antioxidant signaling activation and
epithelial barrier preservation in the context of mulberroside A
treatment.
Oxidative stress is widely recognized as a central
pathological driver of intestinal epithelial barrier disruption and
excessive ROS production has been shown to compromise epithelial
integrity by directly damaging cellular components and
destabilizing tight junction architecture (22,24,25).
Previous studies have demonstrated that elevated ROS levels are
associated with reductions in transepithelial electrical resistance
and disorganization of key tight junction proteins, including ZO-1
and occludin, ultimately leading to increased epithelial
permeability (24,26). These observations are consistent
with the present findings, in which H2O2
exposure triggered marked oxidative stress in Caco-2 cells,
accompanied by notable decreases in TEER and disrupted expression
of tight junction proteins. Furthermore, the coordinated recovery
of ROS levels, TEER and tight junction integrity observed following
mulberroside A treatment aligns with the established concept that
restoration of redox homeostasis is associated with the
preservation of epithelial barrier integrity (26–28).
From a molecular perspective, the Nrf2 pathway has
been extensively characterized as a key regulator of cellular
antioxidant defense and epithelial protection (29,30).
Activation of Nrf2 promotes its nuclear translocation and
subsequent transcriptional upregulation of cytoprotective enzymes,
including HO-1 and NQO1, which serve key roles in mitigating
oxidative injury and maintaining epithelial homeostasis.
Accumulating evidence has indicated that impairment of Nrf2
signaling exacerbates oxidative stress-induced epithelial damage,
whereas enhancement of Nrf2 activity confers resistance to
oxidative insults in the gastrointestinal tract (31,32).
In line with these reports, the present data demonstrated that
H2O2 exposure suppressed nuclear Nrf2
accumulation and reduced HO-1 and NQO1 expression, whereas
mulberroside A treatment was associated with concurrent increases
in nuclear Nrf2 and downstream antioxidant proteins, aligning with
improvements in oxidative stress markers and barrier-associated
parameters. The attenuation of these changes following Nrf2
knockdown further supports the association between Nrf2 signaling
status and epithelial redox balance observed in previous studies
(33,34).
Notably, the cell viability response to mulberroside
A did not exhibit a strictly linear dose-response pattern.
Mulberroside A alone caused only limited changes in basal Caco-2
cell viability across the tested concentrations, although higher
concentrations tended to reduce viability. Similarly, in the
H2O2-challenged model, the medium dose showed
the greatest improvement in cell viability, whereas the high dose
did not further enhance this effect. This may indicate that
CCK-8-based viability, which mainly reflects cellular metabolic
activity, does not necessarily parallel antioxidant and
barrier-protective responses. By contrast, the effects of
mulberroside A on ROS suppression and TEER restoration exhibited a
clearer concentration-associated pattern.
The compound
2,3,5,4′-tetrahydroxystilbene-2-O-β-D-glycoside, a stilbene
glycoside analogous to mulberroside A, has been shown to attenuate
H2O2-induced oxidative damage in MC3T3-E1
osteoblasts in a dose-dependent manner through Nrf2/HO-1/NQO1
activation, with maximal effects at the highest dose tested
(35). Mulberroside A itself has
also been shown to inhibit oxidative stress in
H2O2-stimulated Caco-2 cells, with higher
doses conferring greater intestinal barrier protection (36). Thus, the enhanced ROS suppression
and TEER restoration observed at 20 µM may reflect stronger
activation of antioxidant responses involving the Nrf2/HO-1/NQO1
axis, strengthening antioxidant defense and tight junction
preservation at higher concentrations.
The present study identified mulberroside A as a
previously underexplored modulator of oxidative stress-associated
intestinal epithelial barrier impairment. Using an established
Caco-2 cell-based model, the present study showed that mulberroside
A treatment under oxidative stress conditions was associated with
concurrent alterations in Nrf2 nuclear accumulation, downstream
antioxidant gene expression (HO-1 and NQO1), intracellular redox
status and epithelial barrier integrity. Notably, genetic silencing
of Nrf2 partially attenuated the protective effects of mulberroside
A, supporting the involvement of Nrf2 signaling in mediating these
cellular responses rather than solely representing a parallel
event. By integrating molecular indicators of antioxidant signaling
with biochemical measurements of oxidative stress, functional
assessment of transepithelial resistance and structural evaluation
of tight junction proteins, the present findings provide a
multilevel view associating intracellular redox regulation with
epithelial barrier preservation. Collectively, these findings
extend current knowledge of mulberroside A beyond its reported
antioxidant properties in non-intestinal tissues and offer
mechanistic insights into its potential relevance in oxidative
stress-associated intestinal epithelial injury, with possible
implications for conditions such as inflammatory bowel disease
(36,37).
The present study was performed in a Caco-2
cell-based in vitro model and further validation in animal
models and in vivo systems is required to establish the
physiological importance of the findings. In addition, epithelial
barrier integrity was assessed mainly by TEER measurement and
immunofluorescence staining of ZO-1 and occludin; therefore,
additional western blotting validation of these tight junction
proteins would further strengthen the protein-level evidence.
Furthermore, as a glycoside, mulberroside A may undergo hydrolysis
by intestinal microflora to its aglycone oxyresveratrol,
potentially affecting its oral bioavailability; future
pharmacokinetic studies are needed to clarify its in vivo
absorption characteristics. Despite activation of Nrf2 signaling
being closely associated with the observed effects of mulberroside
A, the upstream regulatory mechanisms responsible for Nrf2
activation were not examined and require further investigation,
particularly with regard to kelch-like ECH-associated protein
1-related regulation. In addition, the H2O2
model primarily represents acute oxidative stress and the impact of
mulberroside A under chronic inflammatory or disease-relevant
conditions remains to be determined. In addition, the present study
focused primarily on nuclear Nrf2 levels; cytoplasmic and total
Nrf2 protein data would provide a more complete view of Nrf2
nuclear translocation dynamics and warrants examination in future
investigations. Collectively, the present findings provide a
mechanistic basis for subsequent studies aimed at elucidating the
function of mulberroside A in oxidative stress-associated
intestinal disorders.
In conclusion, the present results indicated that
mulberroside A attenuated oxidative stress-induced intestinal
epithelial barrier dysfunction in Caco-2 cells, accompanied by
reduced oxidative stress and preservation of tight junction
integrity. These effects were closely associated with activation of
the Nrf2/HO-1 antioxidant pathway, as supported by increased Nrf2
nuclear accumulation and downstream antioxidant responses.
Collectively, these findings provided mechanistic insight into the
intestinal protective potential of mulberroside A and thus support
further investigation of mulberroside A in oxidative
stress-associated intestinal epithelial injury.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
JC, XH and XC conceived and designed the study. JC
and WJ performed the experiments and collected the data. DL
conducted the statistical analyses and interpreted the results. XH
and XC supervised the project and revised it critically for
important intellectual content. JC drafted the manuscript. All
authors read and approved the final version of the manuscript. JC
and XH confirm the authenticity of all the raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang R, Li Z, Liu S and Zhang D: Global,
regional and national burden of inflammatory bowel disease in 204
countries and territories from 1990 to 2019: A systematic analysis
based on the Global Burden of disease study 2019. BMJ Open.
13:e0651862023. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kaplan GG: The global burden of
inflammatory bowel disease: From 2025 to 2045. Nat Rev
Gastroenterol Hepatol. 22:708–720. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Arumugam P, Saha K and Nighot P:
Intestinal epithelial tight junction barrier regulation by novel
pathways. Inflamm Bowel Dis. 31:259–271. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Muro P, Zhang L, Li S, Zhao Z, Jin T, Mao
F and Mao Z: The emerging role of oxidative stress in inflammatory
bowel disease. Front Endocrinol (Lausanne). 15:13903512024.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jin J, Cen T, Huang M, Xu F, Ding Q, Lv D,
Wang S, Fei L, Ma H and Fu P: Involvement of mitochondrial unfolded
protein response and activating transcription factor 4 in the
mitochondrial damage pathway of BEAS-2B cells induced by cigarette
smoke extracts. J Thorac Dis. 17:9586–9597. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kustra A, Maliszewska-Olejniczak K,
Sekrecka-Belniak A, Kulawiak B and Bednarczyk P: Polystyrene
nanoplastics in human gastrointestinal Models-cellular and
molecular mechanisms of toxicity. Int J Mol Sci. 26:117382025.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tratenšek A, Locatelli I, Grabnar I,
Drobne D and Vovk T: Oxidative stress-related biomarkers as
promising indicators of inflammatory bowel disease activity: A
systematic review and meta-analysis. Redox Biol. 77:1033802024.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rodrigues Junior JI, Vasconcelos JKG,
Xavier LEMDS, Gomes ADS, Santos JCF, Campos SBG, Martins ASDP,
Goulart MOF and Moura FA: Antioxidant therapy in inflammatory bowel
disease: A systematic review and a Meta-analysis of randomized
clinical trials. Pharmaceuticals (Basel). 16:13742023. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bora SJ, Bhattacharyya S, Deb S and Sarkar
D: Phytochemicals as potential therapeutic agents for inflammatory
bowel disease: A comprehensive review. Phytochem Rev. 25:1985–2026.
2026. View Article : Google Scholar
|
|
10
|
Paudel D, Nair DVT, Joseph G, Castro R,
Tiwari AK and Singh V: Gastrointestinal microbiota-directed
nutritional and therapeutic interventions for inflammatory bowel
disease: Opportunities and challenges. Gastroenterol Rep (Oxf).
12:goae0332024. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang X, Cheng Y, Huang J, Xu F, Jiang J,
Nalinratana N, Jin L and Xue Y: Engineered probiotics for
inflammatory bowel disease therapy: Mechanisms, delivery
strategies, and precision medicine. Front Microbiol.
16:16965242025. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yuan L, Wang Y, Li N, Yang X, Sun X, Tian
H and Zhang Y: Mechanism of action and therapeutic implications of
Nrf2/HO-1 in inflammatory bowel disease. Antioxidants (Basel).
13:10122024. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Khan MZ, Li S, Ullah A, Li Y, Abohashrh M,
Alzahrani FM, Alzahrani KJ, Alsharif KF, Wang C and Ma Q:
Therapeutic agents targeting the Nrf2 signaling pathway to combat
oxidative stress and intestinal inflammation in veterinary and
translational medicine. Vet Sci. 13:252025.PubMed/NCBI
|
|
14
|
Chen H, Li Y, Wang J, Zheng T, Wu C, Cui
M, Feng Y, Ye H, Dong Z and Dang Y: Plant polyphenols attenuate
DSS-induced ulcerative colitis in mice via antioxidation,
anti-inflammation and microbiota regulation. Int J Mol Sci.
24:108282023. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wallner M, Stadlbauer V,
Blank-Landeshammer B, Heckmann M, Sadova N, Iken M, Pitari GM and
Weghuber J: Plant extracts identified by in vitro high-content
screening improve epithelial barrier function and attenuate
oxidative and inflammatory stress. Pharmacol Res Nat Prod.
7:1002262025.
|
|
16
|
Li J, Wang J, Li Y, Guo J, Jin Z, Qiao S,
Zhang Y, Li G, Liu H and Wu C: Mulberroside A: A Multi-target
neuroprotective agent in Alzheimer's disease via cholinergic
restoration and PI3K/AKT pathway activation. Biology (Basel).
14:11142025.PubMed/NCBI
|
|
17
|
Liu T, Zhang Q, Zhao F, Yin J, Liu H, Wang
L and Liu B: Mulberroside A alleviates myocardial infarction by
inhibiting oxidative stress and apoptosis via the PI3K/Akt
signaling pathway. Bratislava Med J. 126:2148–2156. 2025.
View Article : Google Scholar
|
|
18
|
Shi B, Qian J, Miao H, Zhang S, Hu Y, Liu
P and Xu L: Mulberroside A ameliorates CCl4-induced liver fibrosis
in mice via inhibiting pro-inflammatory response. Food Sci Nutr.
11:3433–3441. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu Y, Chen M, Chen C and Huang G: In
vitro and in vivo investigation of the capacity of mulberroside A
to inhibit senescence. NPJ Sci Food. 9:1982025. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Donetti E, Bendinelli P, Correnti M,
Gammella E, Recalcati S and Ferraretto A: Caco2/HT-29 in vitro cell
Co-Culture: Barrier integrity, permeability, and tight Junctions'
composition during progressive passages of parental cells. Biology
(Basel). 14:2672025.PubMed/NCBI
|
|
21
|
Dong Y, Hou Q, Lei J, Wolf PG, Ayansola H
and Zhang B: Quercetin alleviates intestinal oxidative damage
induced by H2O2 via modulation of GSH: In vitro screening and in
vivo evaluation in a colitis model of mice. ACS Omega. 5:8334–8346.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hasegawa T, Mizugaki A, Inoue Y, Kato H
and Murakami H: Cystine reduces tight junction permeability and
intestinal inflammation induced by oxidative stress in Caco-2
cells. Amino Acids. 53:1021–1032. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shen C, Luo Z, Ma S, Yu C, Lai T, Tang S,
Zhang H, Zhang J, Xu W and Xu J: Microbe-derived antioxidants
protect IPEC-1 cells from H2O2-induced oxidative stress,
inflammation and tight junction protein disruption via activating
the Nrf2 pathway to inhibit the ROS/NLRP3/IL-1β signaling pathway.
Antioxidants (Basel). 13:5332024. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen L, Chu H, Hu L, Li Z, Yang L and Hou
X: The role of NADPH oxidase 1 in alcohol-induced oxidative stress
injury of intestinal epithelial cells. Cell Biol Toxicol.
39:2345–2364. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lin PY, Stern A, Peng HH, Chen JH and Yang
HC: Redox and metabolic regulation of intestinal barrier function
and associated disorders. Int J Mol Sci. 23:144632022. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li G, Wang S and Fan Z: Oxidative stress
in intestinal Ischemia-reperfusion. Front Med (Lausanne).
8:7507312021. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Stojanovic B, Milivojcevic Bevc I,
Dimitrijevic Stojanovic M, Stojanovic BS, Jovanovic M, Lazarevic S,
Milosevic B, Radosavljevic I, Tasic-Uros D, Markovic N, et al: Nrf2
as a molecular guardian of redox balance and barrier integrity in
IBD. Antioxidants (Basel). 14:14072025. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Peng S, Shen L, Yu X, Zhang L, Xu K, Xia
Y, Zha L, Wu J and Luo H: The role of Nrf2 in the pathogenesis and
treatment of ulcerative colitis. Front Immunol. 14:12001112023.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shuhua Y, Lingqi M, Yunlong D, He T, Yao S
and Peng L: Proanthocyanidins activate Nrf2/ARE signaling pathway
in intestinal epithelial cells by inhibiting the ubiquitinated
degradation of Nrf2. Biomed Res Int. 2022:85627952022. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang Y, Chai H, Lan T, Gan Q, Liang Y,
Tang C, Zeng Y and Liang H: SMP30 May protect human lens epithelial
cells against high glucose-induced oxidative damage by regulating
the Keap1/Nrf2/NQO1 pathway. Sci Rep. 15:432332025. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shi M, Luan Y, Zhang Z, Xi X and Li W:
FSTL1 silencing protects against lipopolysaccharide-induced
ferroptosis in renal tubular cells by regulating the PI3K/Akt
pathway. Peptides. 194:1714542025. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen C, Tao M, Wang L, Yan W, Lin R and Fu
Y: Nicotinamide N-methyltransferase mediates redox regulation and
colonic epithelial barrier impairment by SIRT1/PPAR-γ/NLRP6/NRF2
axis in inflammatory bowel disease. Int Immunopharmacol.
168:1158822026. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lim HS, Park J, Whang WJ, Kang WS, Kim S,
Yoo YS and Park G: Melatonin ameliorates desiccation Stress-induced
ocular inflammation in an in vitro model by activating the Nrf2
pathway. J Cell Mol Med. 29:e708792025. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cheng J, Wang H, Zhang Z and Liang K:
Stilbene glycoside protects osteoblasts against oxidative damage
via Nrf2/HO-1 and NF-κB signaling pathways. Arch Med Sci.
15:196–203. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yu R, Wen S, Wang Q, Wang C, Zhang L, Wu
X, Li J and Kong L: Mulberroside A repairs high fructose
diet-induced damage of intestinal epithelial and blood-brain
barriers in mice: A potential for preventing hippocampal
neuroinflammatory injury. J Neurochem. 157:1979–1991. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang M, Hu X and Xiong S: Pharmacological
activities and pharmacokinetic properties of mulberroside: A
systematical review. Fitoterapia. 186:1067992025. View Article : Google Scholar : PubMed/NCBI
|