Introduction
Sepsis is a life-threatening syndrome defined by
organ dysfunction arising from a dysregulated host response to
infection, with up to 60% of patients progressing to acute kidney
injury (AKI) (1). Its pathogenesis
is multifactorial, involving excessive immune activation, cytokine
storms and microvascular dysfunction (2). Despite progress in supportive care,
sepsis remains a major clinical challenge, contributing to ~11
million deaths worldwide annually, and often results in poor
outcomes for survivors (3).
Current treatments primarily rely on early recognition and
non-specific supportive interventions, such as broad-spectrum
antibiotics, fluid resuscitation and hemodynamic stabilization
(4). However, no effective
targeted pharmacotherapies exist, and present strategies remain
largely symptomatic and insufficient (3).
Traditional Chinese medicine (TCM) offers
therapeutic benefits through multi-component, multi-pathway and
multi-target mechanisms and is considered a potential complementary
strategy for the management of sepsis-related inflammatory
disorders (5). Laggera alata
(D. Don) Sch. Bip. ex Oliv, a member of the Asteraceae family,
is native to eastern, southeastern and southwestern China and has
been traditionally used in TCM for effects traditionally described
as dispelling wind, eliminating dampness and detoxification
(6). As a traditional Chinese
medicinal herb, Laggera alata has long been used in Chinese
folk medicine to treat various disorders and has been reported to
exhibit antibacterial, anti-inflammatory and antipyretic activities
(7). Traditionally, Laggera
alata has been commonly used for the treatment of upper
respiratory tract infections, influenza, parotitis and recurrent
herpes viral infections, with its extracts showing notable
anti-inflammatory and antiviral activities (8). Additionally, this plant is frequently
employed to eliminate phlegm and treat bronchitis and jaundice,
which is closely related to its rich essential oil content. These
essential oils are mainly composed of oxygenated monoterpenes and
sesquiterpenes, among which the aromatic ether
2,5-dimethoxy-p-cymene is one of the major active constituents
(9). Modern pharmacological
studies have shown that extracts of Laggera alata possess
anti-inflammatory properties and show protective effects in various
disease models, including carbon tetrachloride-induced hepatic
fibrosis (10), collagen-induced
rheumatoid arthritis (11) and
sepsis-associated AKI (12).
However, the molecular mechanisms underlying its effects in
endotoxin-induced systemic inflammation remain poorly
understood.
The authors' previous research demonstrated that
total phenolics extracted from Laggera alata (TPLA)
attenuated inflammation in a rheumatoid arthritis model by
suppressing M1 macrophage polarization (11). Among the key drivers of multi-organ
dysfunction in sepsis is mitochondrial impairment, especially in
the kidneys (13). Mitophagy, an
essential mitochondrial quality-control process that selectively
eliminates damaged or superfluous mitochondria, has recently been
recognized as a critical regulator of mitochondrial homeostasis,
immune regulation and disease progression (14–16).
Among the currently identified mitophagy pathways, the PTEN-induced
putative kinase 1 (PINK1)/Parkin-dependent pathway remains one of
the best-characterized mechanisms, in which PINK1 accumulates on
damaged mitochondria and recruits Parkin to initiate mitochondrial
ubiquitination and autophagic clearance (16). Recent evidence further indicates
that mitochondrial dysfunction, impaired mitophagy, excessive
mitochondrial reactive oxygen species (ROS) production and altered
mitochondrial metabolism are closely associated with macrophage
polarization imbalance and immune dysregulation during sepsis
(17,18). Moreover, activation of the
PINK1/Parkin pathway has been shown to mitigate renal injury and
improve outcomes in AKI models (19). In particular, recent studies have
linked PINK1/Parkin-mediated mitophagy to macrophage phenotype
remodeling, suggesting that mitophagy may serve as a mechanistic
bridge between mitochondrial quality control and inflammatory
resolution (20,21). While mitophagy has emerged as a key
mechanism in sepsis pathophysiology, the potential role of TPLA in
modulating PINK1/Parkin-associated mitophagy and immune regulation
has not yet been investigated. The present study aimed to elucidate
the mechanisms by which TPLA alleviates LPS-induced systemic
inflammation and organ injury, with a focus on
PINK1/Parkin-associated mitophagy and macrophage polarization.
Materials and methods
Preparation of TPLA
Plant material was collected on November 29, 2019,
from Sanfang Reservoir (Longxu, Cangwu, Guangxi, China). The
voucher specimen (collection no. 450421191129020LY) was deposited
in the Herbarium of Guangxi Institute of Botany (IBK) under voucher
number IBK00430105. The specimen was identified by Yusong Huang on
October 24, 2020, as Laggera alata (D. Don) Sch. Bip. ex
Oliv, a species belonging to the genus Laggera of the
family Asteraceae. The whole plant was used for extraction. The
plant name was verified against The Plant List (http://www.theplantlist.org). TPLA was extracted
according to previously published protocols (11). Dried Laggera alata herb (10
kg) was ground into coarse powder and extracted twice by reflux
with 95% ethanol at ~84°C, using solvent-to-material ratios of 12:1
and 8:1 (L/kg) for 1.5 and 1 h, respectively. The combined extracts
were filtered, concentrated under reduced pressure using a rotary
evaporator, and subsequently dried in a 65°C water bath. The yield
of TPLA (total phenolics: 13.52%) was 2.156 kg, with an extraction
efficiency of 21.56%. The extract was sealed and stored in a dry
environment. For experimental use, an accurately weighed amount of
TPLA powder was dissolved in distilled water and diluted to the
required concentration immediately before use.
Cell culture and treatment
The mouse macrophage cell line RAW264.7 (cat. no.
CL-0190; Wuhan Pricella Biotechnology Co., Ltd.) was cultured in
high-glucose Dulbecco's Modified Eagle Medium (DMEM; cat. no.
E600003; Sangon Biotech Co., Ltd.) supplemented with 10% fetal
bovine serum (FBS; Guangzhou Yunoer Biotechnology Co., Ltd.) and 1%
penicillin-streptomycin (cat. no. P1400; Beijing Solarbio Science
& Technology Co., Ltd.) at 37°C in a 5% CO2
incubator. Cells were tested and confirmed to be mycoplasma-free
and authenticated by short tandem repeat (STR) profiling (Wuhan
Pricella Biotechnology Co., Ltd.). Cells at 80–90% confluence were
used for experiments. To establish an in vitro LPS model
group, cells were treated with 1 µg/ml lipopolysaccharide (LPS;
cat. no. L2630; MilliporeSigma) for 24 h (22), followed by treatment with TPLA at
low (10 µg/ml), medium (50 µg/ml) and high (100 µg/ml)
concentrations or dexamethasone (DEX; 10 µM; cat. no. D4902;
MilliporeSigma). The control group received no treatment. For
mitophagy inhibition experiments, cells were additionally assigned
to the LPS, LPS + TPLA-high, and LPS + TPLA-high + Mdivi-1 groups.
Mdivi-1 (10 µM; cat. no. M0199; Sigma-Aldrich; Merck KGaA) was
added 1 h before TPLA-high treatment. Mitophagy-related protein
expression was subsequently evaluated by western blotting. In the
Mdivi-1 inhibition experiment, macrophage polarization was further
assessed by flow cytometry using CD80/CD86 as M1-associated markers
and CD206/CD163 as M2-associated markers.
Enzyme-linked immunosorbent assay
(ELISA)
Levels of IL-1β (cat. no. MLB00C-1; R&D Systems,
Inc.), IL-6 (cat. no. BMS603-2), TNF-α (cat. no. BMS607-3TEN),
IL-1α (cat. no. 900-K82K), IL-4 (cat. no. 88-7044-88) and IL-10
(cat. no. 88-7105-88; all from Thermo Fisher Scientific, Inc.) were
quantified using ELISA kits. Samples were added to 96-well plates
pre-coated with capture antibodies and incubated for 2 h at 37°C.
After washing, biotin-conjugated detection antibodies were added
and incubated for 1 h at 37°C, after which the substrate solution
(cat. no. A8043; Thermo Fisher Scientific, Inc.) was added for
color development. Absorbance was measured at 450 nm using a
microplate reader (Varioskan™ LUX; Thermo Fisher
Scientific, Inc.), and concentrations were calculated from standard
curves.
JC-1 staining for mitochondrial
membrane potential (MMP)
MMP was assessed using JC-1 dye (cat. no. C2006;
Beyotime Institute of Biotechnology). After 24 h of treatment,
cells were washed with PBS and incubated with 5 µM JC-1 at 37°C for
30 min in the dark. After staining, cells were washed with JC-1
buffer and immediately analyzed by flow cytometry (Accuri C6; BD
Biosciences). Fluorescence signals were collected in the FL1-H
(green, JC-1 monomers, 530±15 nm) and FL2-H (red, JC-1 aggregates,
585±20 nm) channels under logarithmic scaling. At least 10,000
events were recorded per sample. The red/green fluorescence ratio
was calculated to represent MMP, where red fluorescence indicates
polarized mitochondria and green fluorescence indicates
depolarization. Data were analyzed using FlowJo software (v10.8; BD
Biosciences).
Cell Counting Kit-8 (CCK-8) assay
RAW264.7 cells were seeded at 6×104
cell/ml (200 µl per well) in 96-well plates and treated according
to group allocation. After 24 h of treatment, 20 µl of CCK-8
reagent (cat. no. CK04; Dojindo Laboratories, Inc.) was added to
each well and incubated for 2–4 h at 37°C. Absorbance was then
measured at 450 nm using a microplate reader (Synergy H1; BioTek;
Agilent Technologies, Inc.).
Flow cytometric analysis of
apoptosis
Apoptosis was evaluated using the Annexin V-FITC/PI
Apoptosis Detection Kit (cat. no. A211-01; Vazyme Biotech Co.,
Ltd.) following the manufacturer's instructions. After washing with
PBS (cat. no. E600003; Sangon Biotech Co., Ltd.), cells were
digested with 0.25% trypsin (cat. no. C0201; Beyotime Institute of
Biotechnology) without EDTA, resuspended in 1X binding buffer, and
incubated with 5 µl Annexin V-FITC and 5 µl propidium iodide (PI)
for 15 min at room temperature in the dark. Samples were
immediately analyzed by flow cytometry (Accuri C6; BD Biosciences).
Fluorescence was detected using the FL1-H (FITC, 530±15 nm) and
FL3-H (PI, >670 nm) channels under logarithmic scaling.
Compensation was applied using single-stained controls. At least
10,000 events were acquired per sample, and debris and doublets
were excluded by forward- and side-scatter gating. Early and late
apoptotic cell percentages were calculated using FlowJo software
(v10.8; BD Biosciences).
Western blotting
RAW264.7 cells were lysed in RIPA buffer (cat. no.
P0013B; Beyotime Institute of Biotechnology) after washing with
PBS, and lysates were centrifuged at 12,000 × g for 15 min at 4°C
to collect supernatants. Protein concentration was determined using
a BCA protein assay kit (cat. no. P0012; Beyotime Biotechnology)
according to the manufacturer's instructions. Equal protein amounts
(30 µg) were resolved on 10% gels using SDS-PAGE together with a
pre-stained protein marker (cat. no. G2086; Wuhan Servicebio
Technology Co., Ltd.), followed by transfer onto PVDF membranes
(MilliporeSigma). Membranes were blocked with 5% non-fat milk in
TBST containing 0.1% Tween-20 for 1 h at room temperature and then
incubated with primary antibodies overnight at 4°C: LC3B rabbit
monoclonal antibody (1:2,000; cat. no. ab192890; Abcam), p62 rabbit
polyclonal antibody (1:10,000; cat. no. ab109012; Abcam), Parkin
rabbit polyclonal antibody (1:1,000; cat. no. GB11596; Wuhan
Servicebio Technology Co., Ltd.), PINK1 rabbit polyclonal antibody
(1:500; cat. no. GB114934; Wuhan Servicebio Technology Co., Ltd.)
and GAPDH rabbit polyclonal antibody (1:2,500; cat. no. ab9485;
Abcam). After incubation with horseradish peroxidase
(HRP)-conjugated goat anti-rabbit IgG secondary antibody (1:10,000;
cat. no. ab6721; Abcam), chemiluminescent signals were detected on
X-ray films using ECL substrate (Thermo Fisher Scientific, Inc.).
Band intensities were analyzed with ImageJ software (v1.80;
National Institutes of Health).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted using RNAiso Plus reagent
(cat. no. 9109; Takara Bio, Inc.), and RNA purity and concentration
were determined using a NanoDrop spectrophotometer (Micro Drop;
Bio-DL). cDNA synthesis was performed using PrimeScript RT Reagent
Kit (cat. no. RR037A; Takara Bio, Inc.) under the following
conditions: 37°C for 15 min, 85°C for 5 sec. qPCR was performed
using PowerTrack™ SYBR Green Master Mix (Thermo Fisher
Scientific, Inc.) and specific primers as follows: inducible nitric
oxide synthase (iNOS) forward, 5′-TCCTGGACATTACGACCCCT-3′ and
reverse, 5′-AGGCCTCCAATCTCTGCCTA-3′; IL-12 forward,
5′-CTCCTGTGGGAGAAGCAGAC-3′ and reverse, 5′-CAGATAGCCCATCACCCTGT-3′;
arginase 1 (Arg1) forward, 5′-AGATTATCGGAGCGCCTTTCT-3′ and reverse,
5′-CGTGGTCTCTCACGTCATACT-3′; IL-10 forward,
5′-CCAAGCCTTATCGGAAATGA-3′ and reverse, 5′-TTTTCACAGGGGAGAAATCG-3′;
and GAPDH forward, 5′-GTGTTCCTACCCCCAATGTGT-3′ and reverse,
5′-ATTGTCATACCAGGAAATGAGCTT-3′. The PCR cycling conditions were
95°C for 5 min, followed by 40 cycles of 95°C for 10 sec and 60°C
for 30 sec. A melting curve analysis was performed at the end.
Relative gene expression was calculated using the 2−ΔΔCq
method (23). GAPDH was used as
the internal reference gene.
Immunofluorescence (IF) staining
IF staining of CD80, CD86, CD206 and CD163 was
performed to assess changes in macrophage polarization. After 24 h
of cell culture, cells were washed twice with phosphate-buffered
saline and fixed with 4% paraformaldehyde (cat. no. A0001; Sangon
Biotech Co., Ltd.) for 20 min at room temperature. Cells were then
permeabilized with 0.1% Triton X-100 (cat. no. A600413; Sangon
Biotech Co., Ltd.) for 10 min, followed by blocking with 5% normal
goat serum (cat. no. C0265; Beyotime Institute of Biotechnology)
for 1 h at room temperature. Subsequently, cells were incubated
overnight at 4°C with the following primary antibodies: CD80
(1:200; cat. no. 66406-1-Ig), CD86 (1:200; cat. no. 13395-1-AP),
CD206 (1:200; cat. no. 83485-1-RR) and CD163 (1:200; cat. no.
16646-1-AP; all from Proteintech Group, Inc.). The next day, cells
were washed three times with PBS and incubated for 1 h at room
temperature with Alexa Fluor 488-conjugated secondary antibody
(1:1,000; cat. no. A11034) and Alexa Fluor 594-conjugated secondary
antibody (1:1,000; cat. no. A11032; both from Thermo Fisher
Scientific, Inc.). Nuclei were stained with 1 µg/ml DAPI (cat. no.
C1002; Beyotime Institute of Biotechnology) for 10 min, and images
were captured using a fluorescence microscope (IX71; Olympus
Corporation).
Flow cytometric analysis of macrophage
polarization
Macrophage polarization was further quantified by
flow cytometry. After the indicated treatments, RAW264.7 cells were
collected, washed with PBS, and incubated with
fluorochrome-conjugated anti-mouse antibodies against CD80-FITC
(cat. no. 104705), CD86-PE (cat. no. 105007),
CD206-PerCP/Cyanine5.5 (cat. no. 141715) and CD163-APC (cat. no.
155305; all from BioLegend, Inc.) according to the manufacturers'
instructions. After incubation in the dark at 4°C for 30 min, cells
were washed twice with PBS and analyzed using a BD Accuri C6 flow
cytometer (BD Biosciences). Debris and doublets were excluded by
forward- and side-scatter gating, and at least 10,000 events were
acquired for each sample. The percentages of CD80/CD86-positive and
CD206/CD163-positive cells were analyzed using FlowJo software
(v10.8; BD Biosciences).
Experimental animals
Male C57BL/6 mice (7–8 weeks old, 20–25 g) were
obtained from Hunan Silaike Jingda Laboratory Animal Co., Ltd.
[production license No. SCXK (Xiang) 2019-0004]. Only male mice
were used in this initial in vivo experiment to reduce
variability associated with sex-dependent immune and hormonal
differences and because previous LPS/endotoxemia studies have
reported sex-dependent inflammatory and organ-injury responses,
with male mice often showing greater susceptibility or more
pronounced injury than female mice (24). All animal experiments were
conducted at the Guangxi University of Chinese Medicine, which
holds the laboratory animal use license No. SYXK [Gui] 2019-0001.
All procedures were approved by the Animal Ethics Committee of
Guangxi University of Chinese Medicine (approval no. DW20240923-01;
Nanning, China). Animals were housed in a controlled environment
(25±2°C, 65% humidity, 12/12-h light/dark cycle) with free access
to standard chow and purified water. All animal procedures complied
with the ARRIVE guidelines (25)
and the National Institutes of Health Guide for the Care and Use of
Laboratory Animals.
Establishment of the LPS-induced
systemic inflammation model and pretreatment regimen
Mice were randomly assigned to the following groups:
control, LPS model, TPLA (low, medium and high dose) and DEX (n=6
per group). A total of 36 mice were used in the in vivo
experiment, with six mice allocated to each group. All 36 animals
completed the experimental protocol and were included in the final
analysis. No animals were excluded, no animals were found dead, and
no premature euthanasia was required during the study. At the
scheduled endpoint, all 36 mice were euthanized for sample
collection. TPLA was administered orally at 50, 100, or 200 mg/kg
once daily for 27 consecutive days as a pretreatment regimen
(11). The DEX group received DEX
at 1 mg/kg (26). On the last two
days of the experiment, mice were challenged with intraperitoneal
LPS (20 mg/kg), twice daily for two consecutive days, to induce
acute systemic inflammatory injury (27). Mice exhibiting lethargy,
piloerection and labored breathing within 24 h were considered
successfully modeled (28). Core
body temperature was measured 6 h after the final LPS injection.
The total duration of the in vivo experiment was 27 days,
and mice were euthanized 6 h after the final LPS injection on day
27. At the scheduled endpoint, mice were deeply anesthetized and
euthanized by intraperitoneal injection of 1% sodium pentobarbital
solution (10 mg/ml; Sigma-Aldrich, P3761) at 150 mg/kg body weight
(29,30), followed by cervical dislocation as
a secondary physical method. Death was verified by the absence of
spontaneous respiration and heartbeat, loss of corneal and pedal
withdrawal reflexes, and absence of response to toe pinch before
tissue collection. Humane endpoints were defined before the
experiment to minimize animal suffering. Animals were monitored at
least twice daily for signs of distress, including persistent
lethargy, severe weight loss (>20% of body weight), hypothermia,
hunched posture, tremors, unresponsiveness, or labored breathing.
Mice showing any of these signs would have been immediately
euthanized using the same sodium pentobarbital protocol.
Hematoxylin and eosin (H&E)
staining
Lung and liver tissues were collected and fixed in
10% neutral-buffered formalin (cat. no. HT501128; MilliporeSigma)
for 24 h at room temperature. After dehydration, clearing and
paraffin embedding, 4 µm-thick sections were cut using a microtome
(cat. no. RM2255; Leica Biosystems). Sections were deparaffinized
using xylene (X5; MilliporeSigma) and rehydrated through graded
ethanol. Hematoxylin (cat. no. GHS132) and eosin (cat. no. E4003;
both from MilliporeSigma) were used for nuclear and cytoplasmic
staining, respectively. After dehydration and clearing, slides were
mounted with neutral resin (cat. no. G8110; MilliporeSigma) and
examined under a light microscope (BX53; Olympus Corporation).
Semi-quantitative histopathological scoring was performed to
evaluate lung and liver injury. For each animal, representative
non-overlapping fields were observed, and the overall pathological
injury was scored by two independent investigators blinded to the
experimental groups. Lung injury was assessed according to alveolar
structural destruction, alveolar septal thickening, inflammatory
cell infiltration, congestion and alveolar collapse. Liver injury
was assessed according to hepatocellular swelling, cytoplasmic
rarefaction, inflammatory cell infiltration, nuclear pyknosis and
disruption of hepatic cord architecture. The severity of
pathological injury was graded using a 0–4 scale as follows: 0, no
obvious injury; 1, mild injury; 2, moderate injury; 3, severe
injury; and 4, very severe injury.
Statistical analysis
Statistical analyses and graph generation were
performed using GraphPad Prism 10.0 (GraphPad Software; Dotmatics).
Data are presented as the mean ± standard deviation.
Semi-quantitative histopathological injury scores are presented as
median with interquartile range. Data normality was assessed using
the Shapiro-Wilk test. For normally distributed data,
independent-samples t-tests were used for two-group comparisons,
and one-way ANOVA with Tukey's post hoc test was used for multiple
group comparisons. For non-normally distributed data, the Wilcoxon
rank-sum test and Kruskal-Wallis test with Dunn's multiple
comparisons were applied. P<0.05 was considered to indicate a
statistically significant difference.
Results
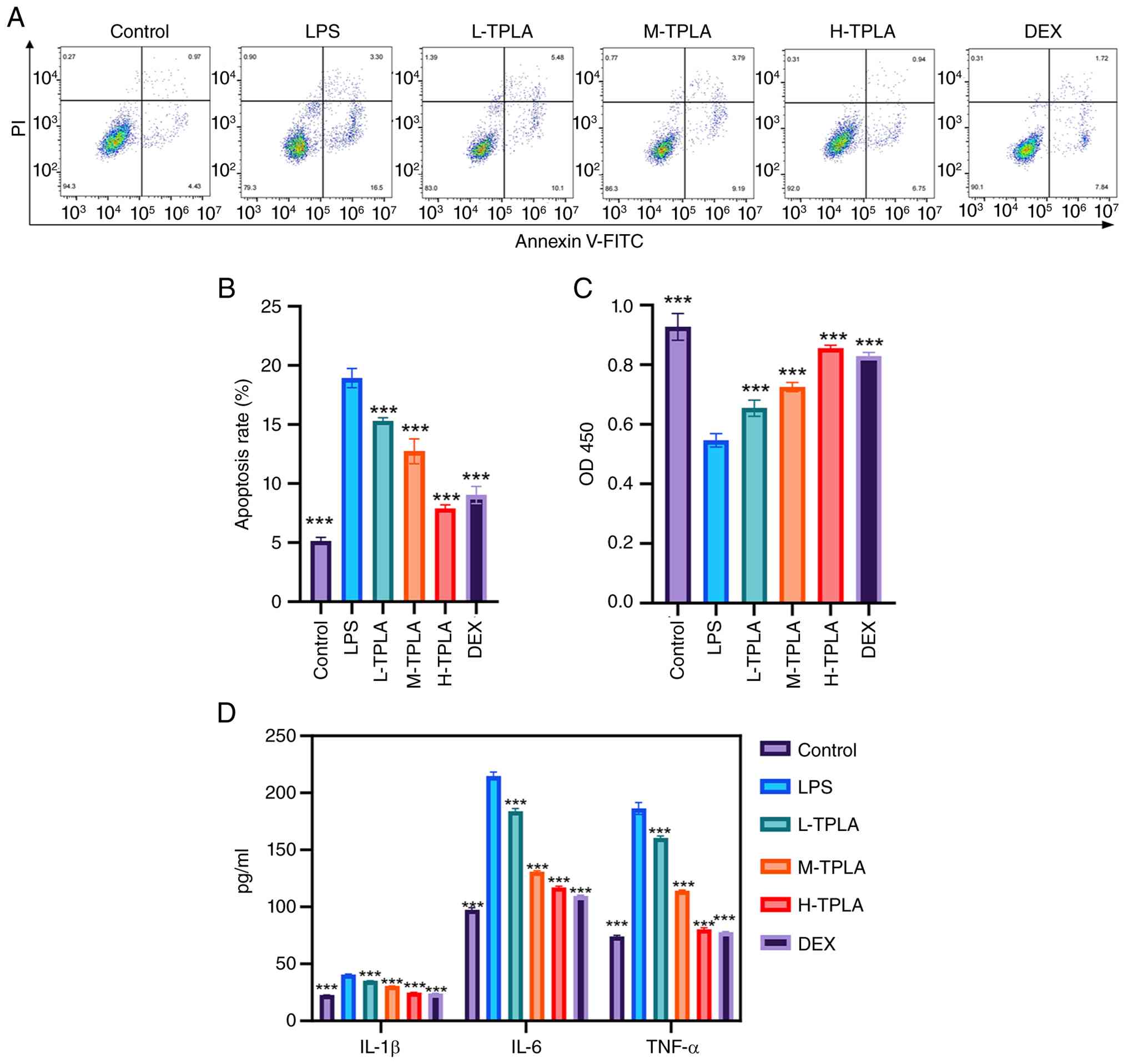
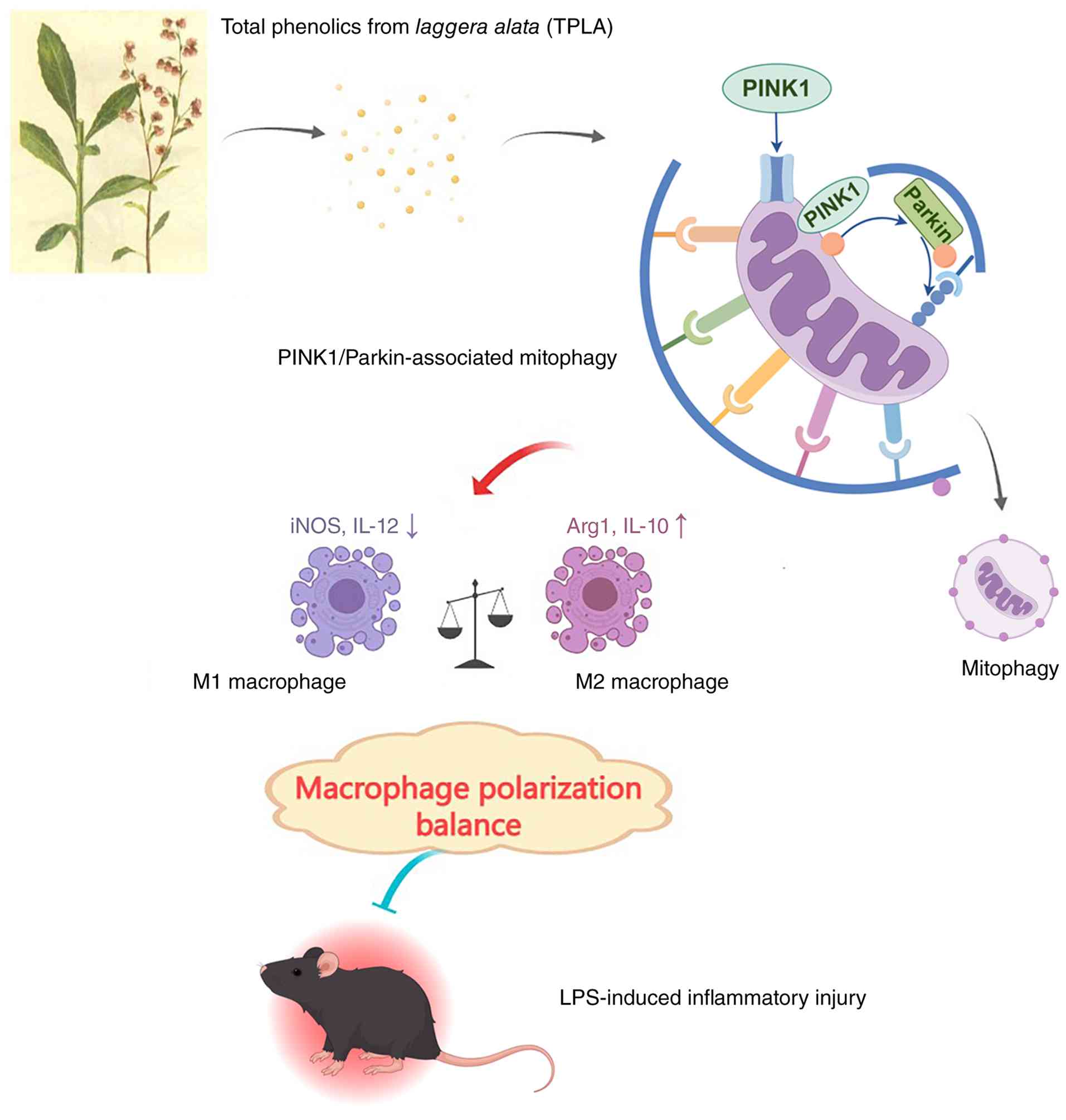
TPLA significantly alleviates
LPS-induced macrophage apoptosis and pro-inflammatory cytokine
release
Flow cytometric analysis revealed a significant
increase in both early and late apoptotic rates in macrophages
treated with LPS, whereas treatment with low, medium and high doses
of TPLA reduced apoptosis in a dose-dependent manner, with the
high-dose group showing an effect similar to that of DEX (Fig. 1A). Quantitative analysis confirmed
that the apoptotic rate in the LPS group increased to ~20%, while
treatment with TPLA reduced this rate to ~15, 13 and 8%,
respectively, and these reductions were statistically significant
compared with the LPS group (Fig.
1B). Cell viability assessed by CCK-8 assay showed a
significant reduction following LPS stimulation, which was restored
in a dose-dependent manner by TPLA treatment to levels approaching
those of the control group (Fig.
1C). ELISA results demonstrated that LPS significantly
increased the levels of IL-1β, IL-6 and TNF-α, while TPLA treatment
significantly suppressed the secretion of these cytokines,
especially in the high-dose group, which exhibited cytokine levels
comparable to the control (Fig.
1D).
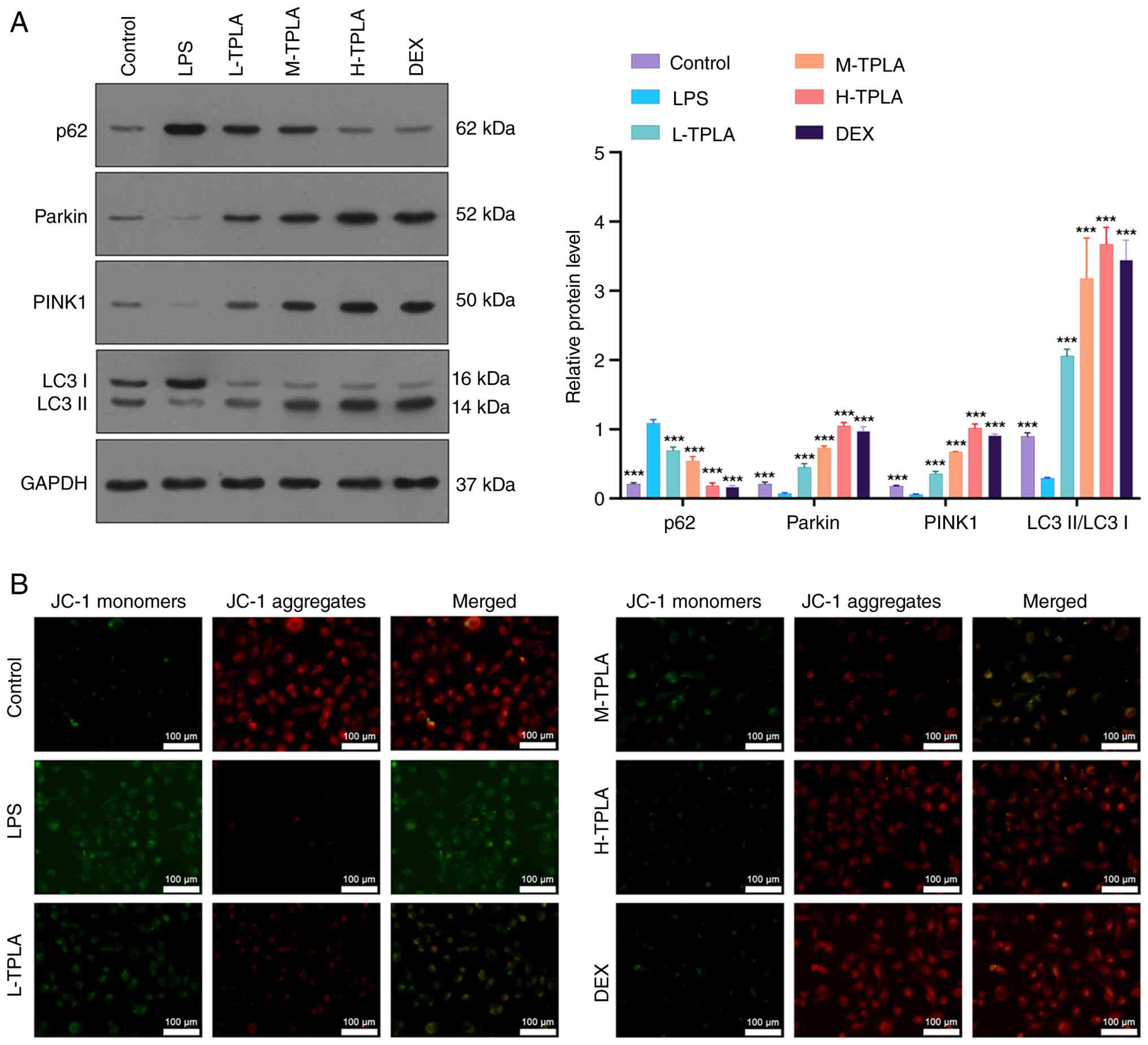
TPLA restores LPS-suppressed
PINK1/Parkin-associated mitophagy-related signaling in
macrophages
Western blot analysis indicated that LPS treatment
significantly reduced LC3-II expression and decreased the
LC3-II/LC3-I ratio, suggesting suppression of autophagy-related
activity. This impairment was reversed by medium and high doses of
TPLA, which restored the LC3-II/LC3-I ratio to levels comparable to
those in the DEX group. In addition, LPS elevated expression of the
autophagy substrate p62 while downregulating the mitophagy-related
proteins Parkin and PINK1. TPLA treatment dose-dependently reduced
p62 expression and significantly increased Parkin and PINK1 levels,
consistent with activation of PINK1/Parkin-associated
mitophagy-related signaling (Fig.
2A). JC-1 staining revealed that LPS reduced MMP, as evidenced
by increased green monomer fluorescence and decreased red aggregate
fluorescence. This mitochondrial dysfunction was reversed by TPLA
treatment, particularly at higher doses (Fig. 2B).
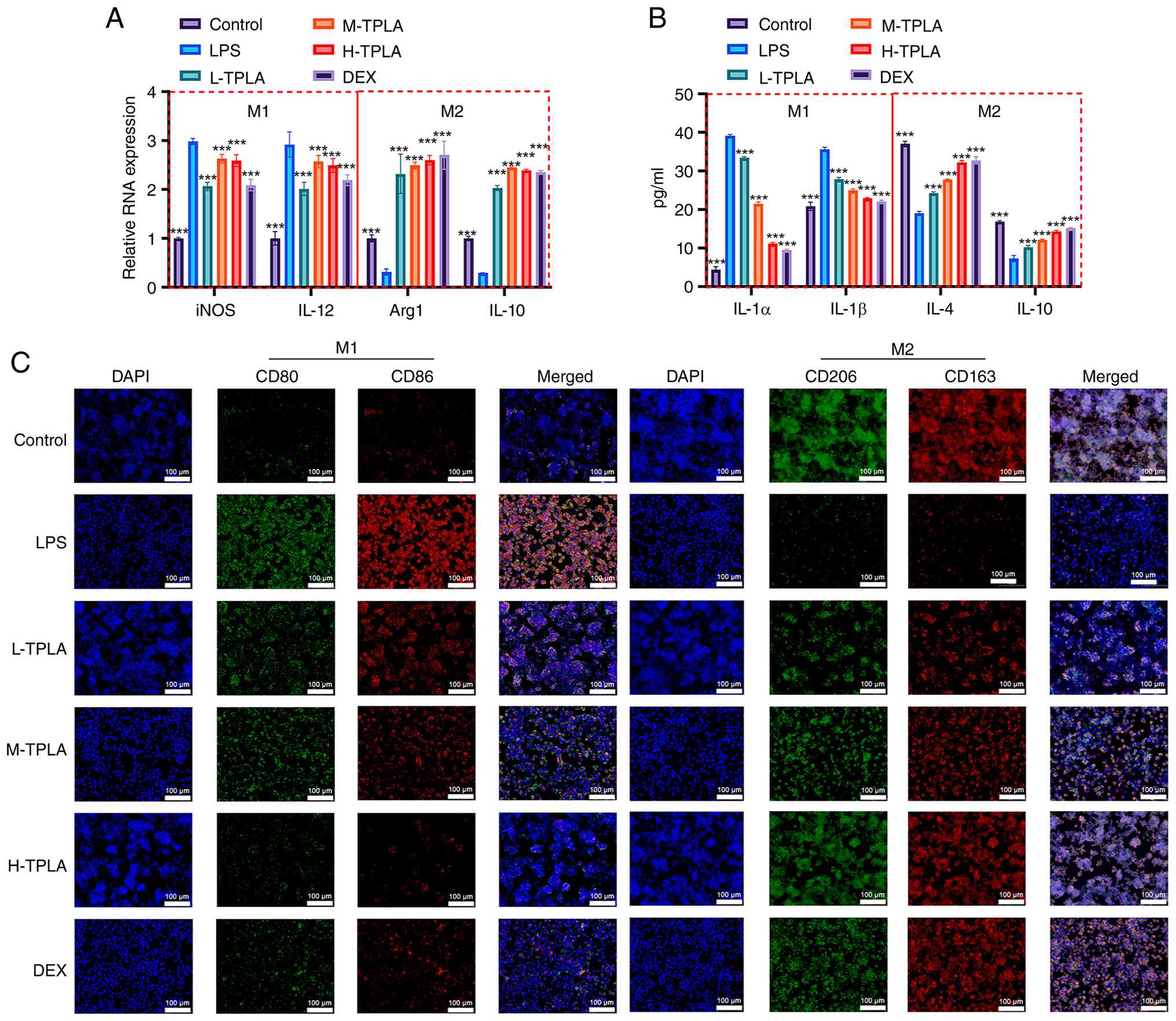
TPLA suppresses LPS-induced M1
macrophage polarization and promotes M2 phenotype transition
RT-qPCR analysis showed that LPS stimulation
significantly increased M1-associated genes iNOS and IL-12, while
decreasing M2-associated genes Arg1 and IL-10. TPLA treatment
dose-dependently reversed this trend, decreasing iNOS and IL-12
expression and enhancing Arg1 and IL-10 expression (Fig. 3A). ELISA results demonstrated that
LPS significantly elevated levels of M1-associated cytokines IL-1α
and IL-1β and suppressed M2-associated cytokines IL-4 and IL-10.
TPLA treatment decreased M1 cytokine levels and increased M2
cytokine levels in a dose-dependent manner (Fig. 3B). IF analysis further confirmed
these results. CD80/CD86 expression (M1 markers) was increased in
the LPS group, while CD206/CD163 expression (M2 markers) was
reduced. TPLA treatment reversed these changes, with high-dose TPLA
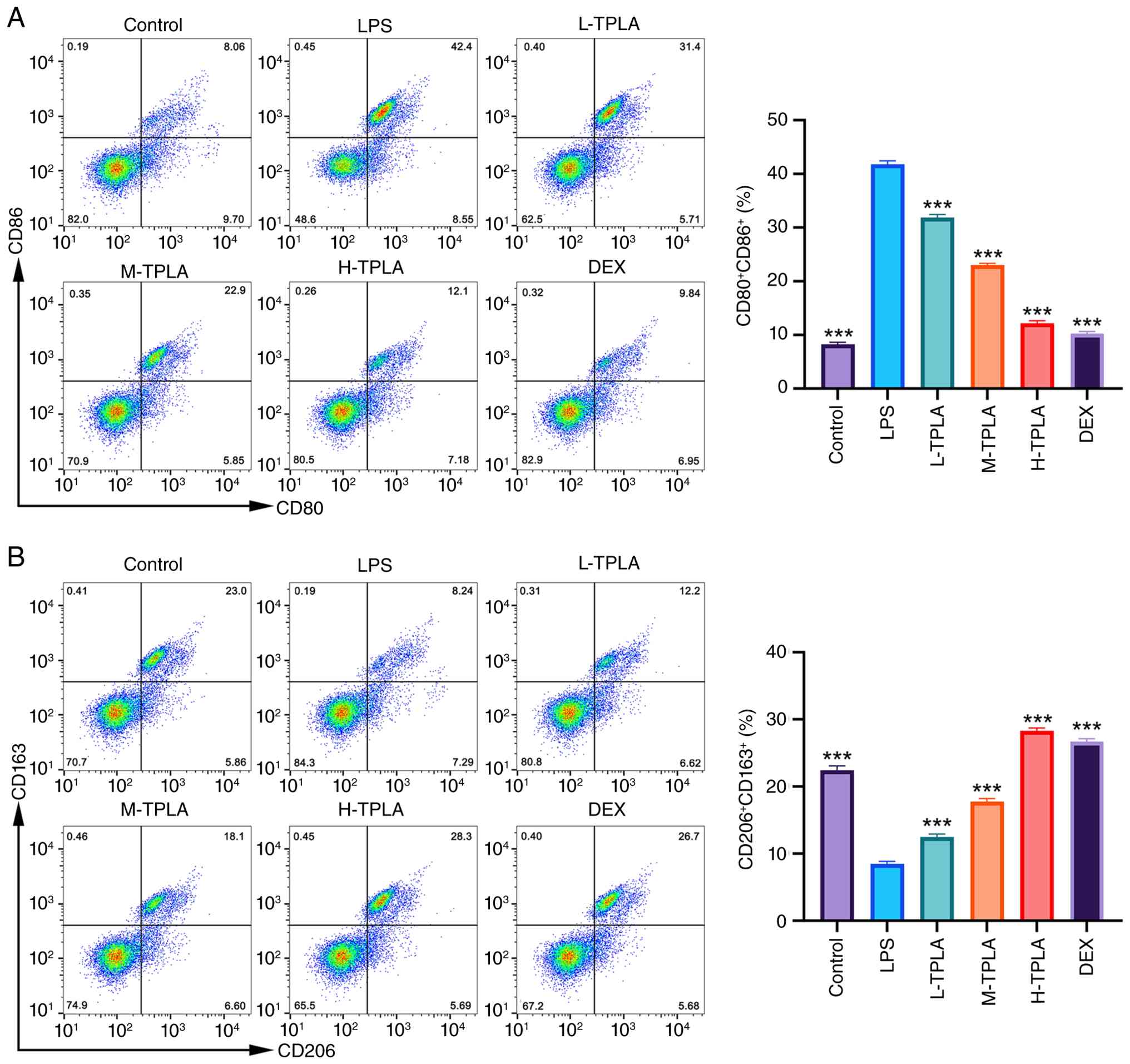
showing effects comparable to DEX (Fig. 3C). Flow cytometric analysis further
confirmed the regulatory effect of TPLA on macrophage polarization.
LPS stimulation significantly increased the proportion of
CD80/CD86-positive M1 macrophages compared with the control group,
whereas TPLA treatment reduced the CD80/CD86-positive population in
a dose-dependent manner. In particular, high-dose TPLA decreased
the proportion of CD80/CD86-positive macrophages to a level close
to that observed in the DEX group (Fig. 4A). Conversely, LPS significantly
decreased the proportion of CD206/CD163-positive M2 macrophages,
while TPLA treatment dose-dependently restored the
CD206/CD163-positive population. The effect of high-dose TPLA was
comparable to that of DEX (Fig.
4B).
TPLA shifts macrophage polarization
toward an M2-like phenotype and enhances PINK1/Parkin-associated
mitophagy-related signaling
LPS stimulation significantly increased the
proportion of CD80/CD86-positive M1 macrophages compared with the
control group, whereas high-dose TPLA treatment reduced the
CD80/CD86-positive population (Fig.
5A). Conversely, LPS significantly decreased the proportion of
CD206/CD163-positive M2 macrophages, while high-dose TPLA treatment
restored the CD206/CD163-positive population (Fig. 5B). However, co-treatment with
Mdivi-1 partially reversed the effects of high-dose TPLA. To
further examine whether the mitophagy-related effects of TPLA were
associated with the PINK1/Parkin pathway, Mdivi-1 was used to
inhibit mitochondrial fission- and mitophagy-related responses.
Western blot analysis demonstrated that high-dose TPLA increased
the expression levels of Parkin and PINK1, enhanced the
LC3-II/LC3-I ratio, and reduced p62 accumulation compared with the
LPS group. However, co-treatment with Mdivi-1 partially reversed
these changes, as evidenced by decreased Parkin and PINK1
expression, reduced LC3-II/LC3-I ratio, and increased p62
expression (Fig. 5C). These
results indicate that TPLA-induced activation of the
PINK1/Parkin-associated mitophagy pathway was partially attenuated
by Mdivi-1.
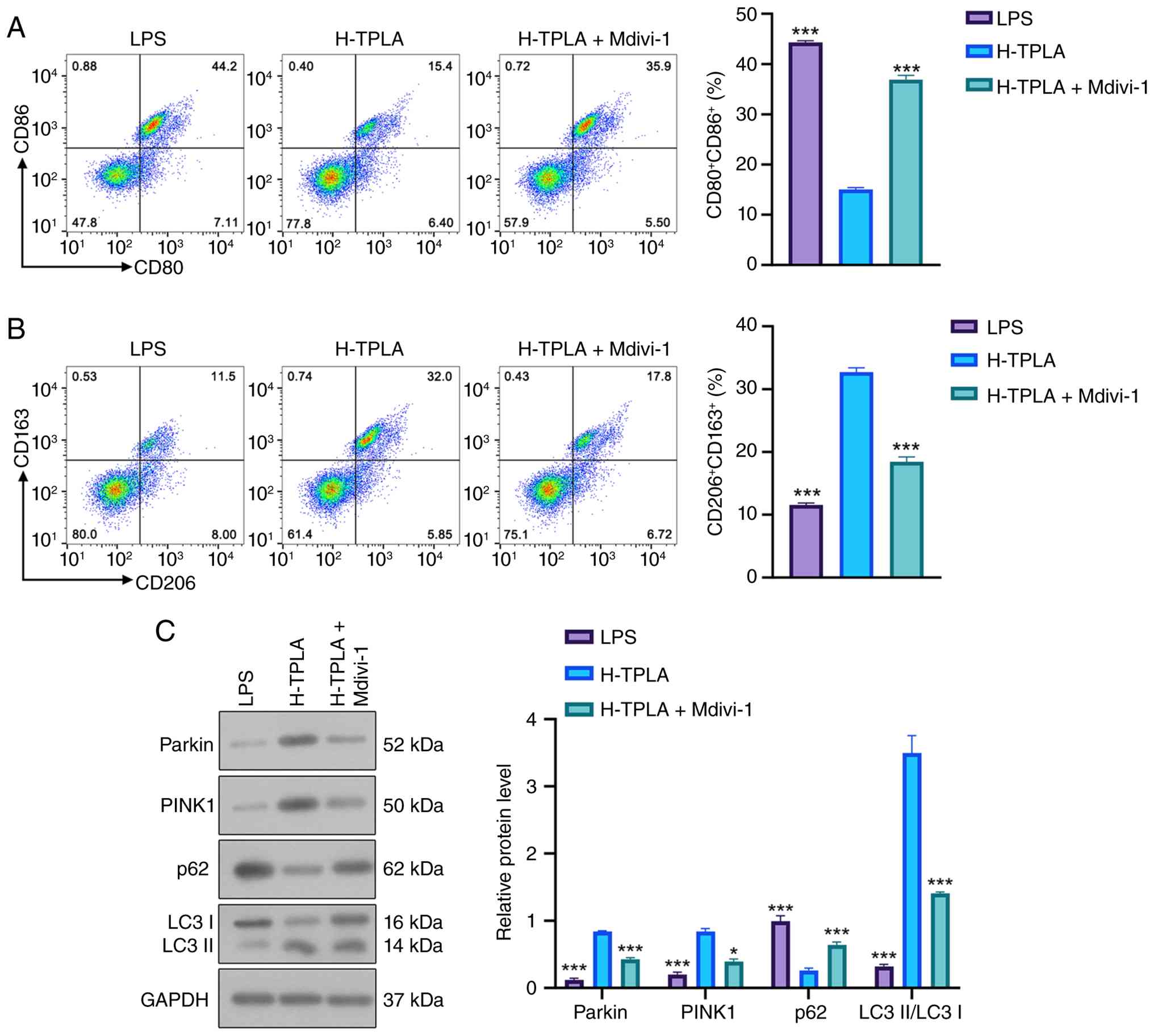 | Figure 5.TPLA shifts macrophage polarization
toward an M2-like phenotype and enhances PINK1/Parkin-associated
mitophagy-related signaling. (A) Representative flow cytometry
plots and quantification of CD80/CD86-positive M1 macrophages. (B)
Representative flow cytometry plots and quantification of
CD206/CD163-positive M2 macrophages. (C) Western blot analysis and
quantitative analysis of Parkin, PINK1, p62 and the LC3-II/LC3-I
ratio in macrophages treated with LPS, H-TPLA, or H-TPLA + Mdivi-1.
Data were analyzed by one-way ANOVA followed by Tukey's post hoc
test. Data are presented as the mean ± SD (n=3). *P<0.05 and
***P<0.001 compared with the H-TPLA group. The protein levels of
p62, Parkin and PINK1 were normalized to GAPDH, whereas LC3 was
expressed as the LC3-II/LC3-I ratio. TPLA, total phenolics from
Laggera alata; LPS, lipopolysaccharide; H-TPLA, high-dose
TPLA; Mdivi-1, mitochondrial division inhibitor 1. |
TPLA alleviates lung and liver tissue
injury in an LPS-induced systemic inflammation mouse model
H&E staining of lung tissues revealed that the
control group exhibited normal alveolar architecture with thin
alveolar septa and minimal inflammatory cell infiltration. By
contrast, the LPS group displayed marked pathological injury,
characterized by alveolar collapse, interstitial thickening,
congestion, and marked inflammatory cell infiltration.
Semi-quantitative histopathological scoring further confirmed that
the lung injury score was markedly increased in the LPS group
compared with the control group. TPLA treatment reduced lung
pathological injury in a dose-dependent manner, with the medium-
and high-dose groups showing clear attenuation of alveolar
structural disruption and inflammatory infiltration. The effect of
high-dose TPLA was similar to that observed in the DEX group
(Fig. 6A and B). In liver tissues,
the control group demonstrated orderly hepatic cords, clear hepatic
sinusoidal structures and intact hepatocyte morphology. The LPS
group exhibited pronounced hepatocellular swelling, cytoplasmic
rarefaction, inflammatory infiltration and nuclear pyknosis.
Semi-quantitative scoring showed that LPS markedly increased the
liver injury score, whereas TPLA treatment reduced hepatic
pathological damage in a dose-dependent manner. In particular,
medium- and high-dose TPLA significantly improved hepatocellular
architecture and reduced inflammatory changes, with high-dose TPLA
showing an effect comparable to DEX (Fig. 6C and D).
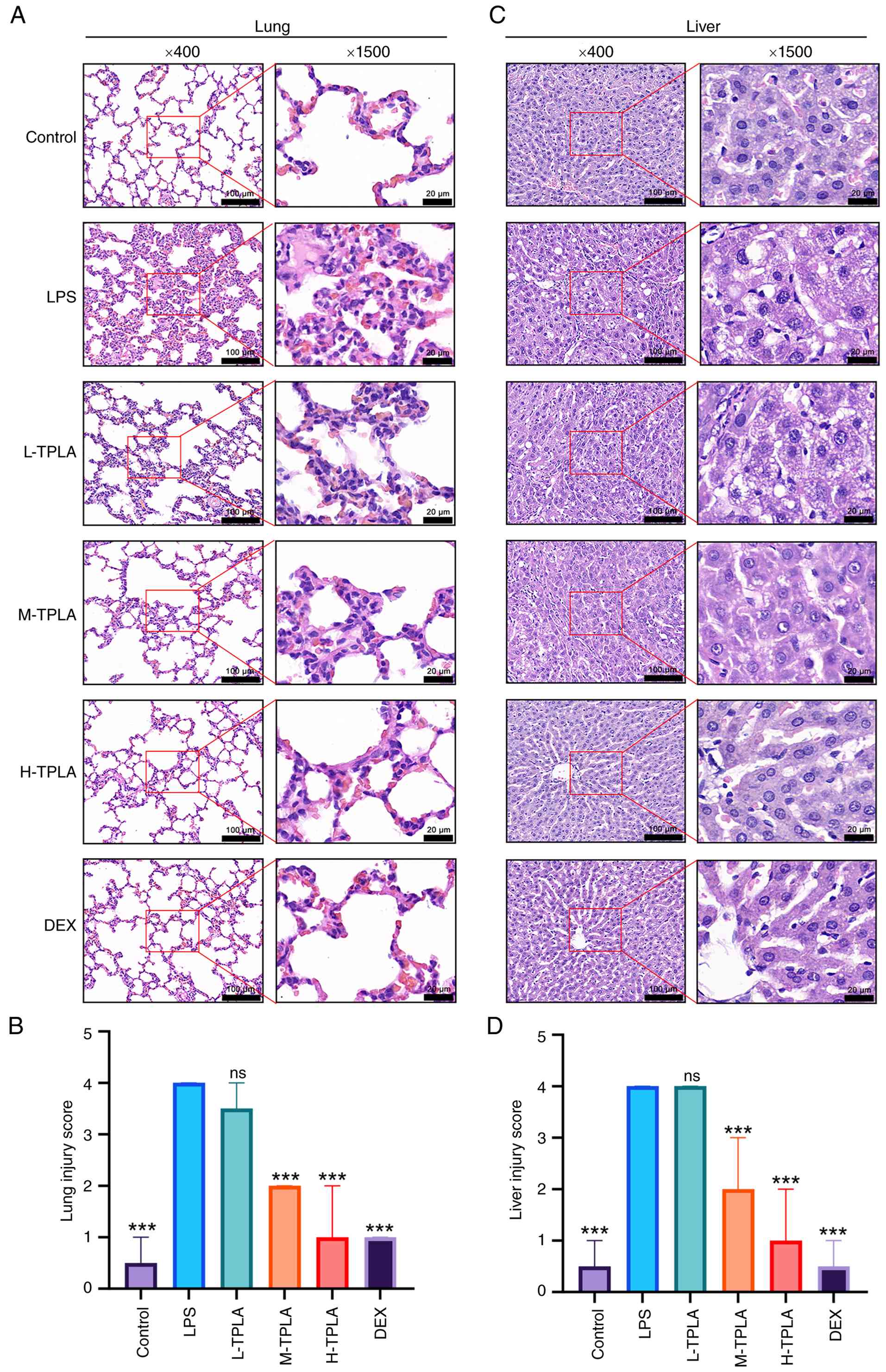 | Figure 6.TPLA alleviates lung and liver tissue
damage in an LPS-induced systemic inflammation mouse model. (A)
Representative H&E-stained images of lung tissues. (B)
Semi-quantitative histopathological injury scores of lung tissues.
(C) Representative H&E-stained images of liver tissues. (D)
Semi-quantitative histopathological injury scores of liver tissues.
Scale bar, 100 µm; enlarged, 20 µm. Data are presented as median
with interquartile range (n=6). Statistical analysis was performed
using the Kruskal-Wallis test followed by Dunn's multiple
comparisons test. ***P<0.001 compared with the LPS group. TPLA,
total phenolics from Laggera alata; LPS, lipopolysaccharide;
L-TPLA, low-dose TPLA; M-TPLA, medium-dose TPLA; H-TPLA, high-dose
TPLA; DEX, dexamethasone; H&E, hematoxylin and eosin; ns, not
significant. |
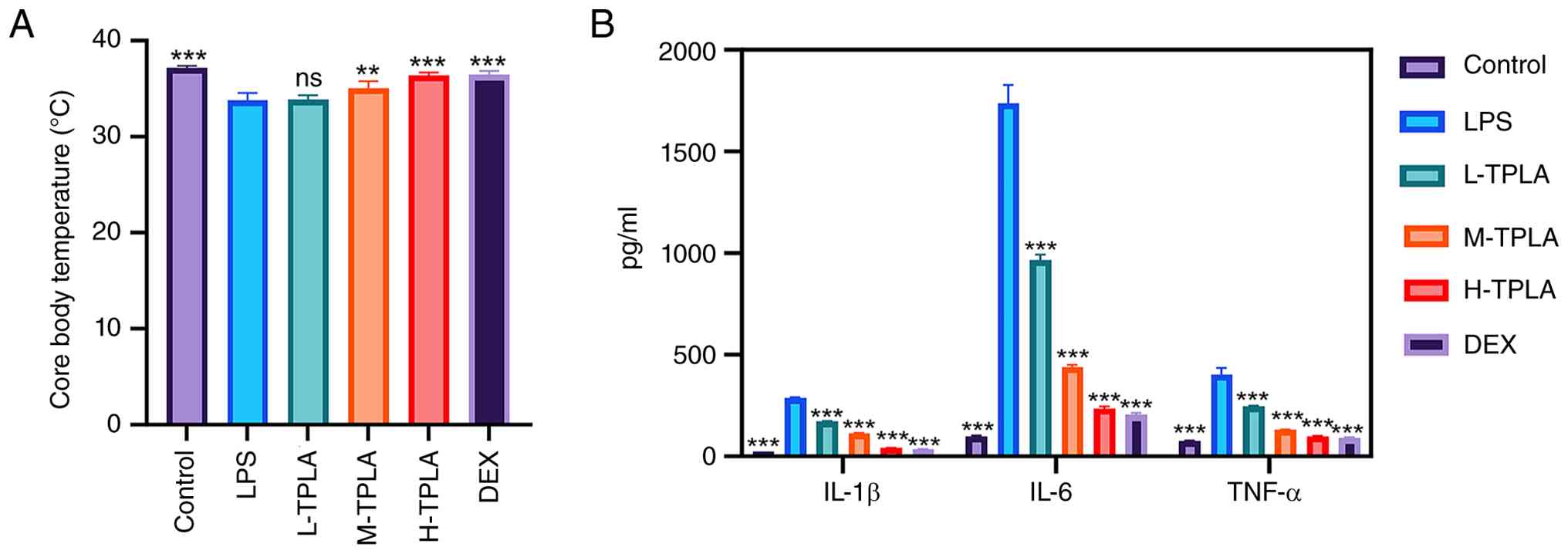
TPLA mitigates LPS-induced hypothermia
and systemic inflammatory response
LPS administration markedly reduced rectal
temperature in mice by ~2°C compared with the control group.
Medium- and high-dose TPLA treatment significantly restored body
temperature to levels comparable with the control and DEX groups,
while the low-dose group showed no significant improvement
(Fig. 7A). ELISA analysis revealed
that LPS markedly increased serum levels of IL-1β, IL-6 and TNF-α,
reaching ~400, 1,800 and 800 pg/ml, respectively. TPLA treatment
reduced serum levels of these inflammatory cytokines in a
dose-dependent manner, with the high-dose group showing the most
pronounced effect, comparable to that of DEX (Fig. 7B).
Discussion
Sepsis is a life-threatening systemic inflammatory
syndrome caused by infection, often accompanied by immune
dysregulation and multi-organ failure (31). In TCM, the pathogenesis of sepsis
is attributed to disruptions in qi and blood as well as dysfunction
of the internal organs. TCM interventions are considered to
alleviate inflammatory damage by modulating systemic immune
responses, restoring internal homeostasis and rebalancing the
microbiota (32). The present
study provides evidence that TPLA attenuates LPS-induced acute
inflammatory injury through modulation of mitophagy-related
signaling and macrophage polarization (Fig. 8).
Previous studies have demonstrated the
immunomodulatory effects of TPLA. Its bioactive constituents have
been reported to suppress M1 macrophage markers and enhance M2
polarization, thereby exerting anti-inflammatory effects (11). Other compounds derived from
Laggera alata, such as isochlorogenic acid A, have shown
hepatoprotective and anti-inflammatory activities, particularly by
targeting pathways like HMGB1/TLR4/NF-κB in hepatic fibrosis
(10). These findings provide a
pharmacological foundation for the potential application of TPLA in
inflammatory disorders.
Macrophages play a central role in the progression
and resolution of sepsis. Although the M1/M2 classification
provides a useful framework, recent reviews emphasize that
macrophage polarization represents a highly plastic and dynamic
spectrum regulated by inflammatory stimuli, metabolic
reprogramming, tissue microenvironment and disease stage (33–35).
In LPS-induced models, macrophages are driven toward the M1
phenotype, amplifying the inflammatory response (36). Consistently, recent studies on
sepsis-associated acute lung injury have shown that macrophage
polarization is closely coupled with mitochondrial dysfunction,
oxidative stress and metabolic remodeling, further supporting
mitochondria-centered regulation as a potential strategy to
rebalance inflammatory macrophage responses (17,18).
In the present study, TPLA shifted macrophages away from an M1-like
phenotype and toward an M2-like phenotype. This immunoregulatory
role is consistent with recent evidence showing that natural
products or bioactive small molecules can regulate inflammatory
injury by targeting mitochondrial dysfunction, mitophagy and
macrophage polarization. For example, urolithin A was recently
reported to alleviate sepsis-induced acute lung injury by reducing
mitochondrial dysfunction, enhancing PINK1/Parkin-associated
mitophagy, and modulating macrophage polarization (37). In addition to regulating macrophage
polarization, TPLA also mitigated the systemic inflammatory
response induced by LPS. One of the early indicators of severe
sepsis is hypothermia, which reflects dysregulation of
thermoregulation. In the present study, LPS administration
significantly reduced the core body temperature of mice, consistent
with previous models of sepsis (38,39).
TPLA treatment, particularly at medium and high doses, restored
body temperature to near-normal levels, comparable to the effect of
DEX. This thermoregulatory protection is likely linked to TPLA's
ability to inhibit pro-inflammatory cytokines such as IL-1β, IL-6
and TNF-α. As hypothermia in sepsis is associated with poor
prognosis, these findings suggest that TPLA may help restore
inflammatory and systemic homeostasis in this model.
A key mechanistic insight from the present study is
the involvement of mitophagy in TPLA's anti-inflammatory effects.
Mitophagy is essential for maintaining mitochondrial integrity,
limiting mitochondrial ROS accumulation, and preserving cellular
metabolic homeostasis. Recent reviews have highlighted that
mitophagy can be mediated by ubiquitin-dependent pathways,
particularly the PINK1/Parkin pathway, as well as receptor-mediated
and other ubiquitin-independent mechanisms (14,40).
In macrophages, mitophagy not only affects mitochondrial quality
control and energy metabolism but also participates in phenotype
remodeling, with recent studies suggesting that activation of
mitophagy can facilitate the transition from a pro-inflammatory M1
phenotype toward a reparative or anti-inflammatory M2-like
phenotype under certain inflammatory contexts (17). During mitophagy, PINK1 cooperates
with the Parkin RBR E3 ubiquitin protein ligase to target damaged
mitochondria for lysosomal degradation (41). It has been shown that
PINK1/Parkin-mediated mitophagy alleviates inflammatory responses
by removing dysfunctional mitochondria and restoring macrophage
function (42). In the present
study, TPLA upregulated PINK1 expression and was associated with
activation of mitophagy-related signaling. This process was
accompanied by downregulation of M1 markers such as iNOS and IL-12
and upregulation of M2 markers including Arg1 and IL-10, thereby
modulating macrophage polarization and reducing LPS-induced
inflammatory responses. Importantly, the additional Mdivi-1
experiment further supported the involvement of
PINK1/Parkin-associated mitophagy in the action of TPLA. Mdivi-1
treatment partially reversed the high-dose TPLA-induced increases
in Parkin, PINK1 and the LC3-II/LC3-I ratio, and increased p62
accumulation. Together with the flow cytometric evidence showing
that TPLA reduced CD80/CD86-positive macrophages and increased
CD206/CD163-positive macrophages, these findings suggest that
TPLA-mediated macrophage phenotype remodeling is closely associated
with activation of the PINK1/Parkin-associated mitophagy
pathway.
Following accumulation on damaged mitochondria,
PINK1 recruits downstream effectors such as Parkin to initiate
mitochondrial ubiquitination and degradation, thus maintaining
mitochondrial function (42,43).
Dysfunction of mitophagy leads to cellular damage through
accumulation of ROS, dysregulated LC3/p62 expression, and reduced
adaptability to inflammatory stress (44). The present findings suggest that
TPLA alleviated these alterations by enhancing mitophagy-related
signaling and improving MMP. PINK1/Parkin-mediated mitophagy has
been reported to reduce renal injury by mitigating mitochondrial
damage (45), and macrophage
mitophagy also protects against renal fibrosis via the
PINK1/MFN2/Parkin axis (46).
Furthermore, PINK1/Parkin-dependent mitophagy is activated during
sepsis and has been shown to exert anti-apoptotic and
immunoregulatory effects in dendritic cells (47). The present study further supports
the possibility that TPLA mitigates inflammatory responses through
activation of the PINK1/Parkin-associated mitophagy pathway.
Dysregulated mitophagy not only impairs
mitochondrial function but also exacerbates macrophage-driven
inflammation. It has been previously shown that in the absence of
functional mitophagy, macrophages exhibit enhanced inflammatory
responses and reduced adaptability to repeated inflammatory stimuli
(48). Interestingly, the role of
mitophagy in immune regulation is bidirectional. Patoli et
al (49) found that inhibition
of PINK1-dependent mitophagy enhanced macrophage bactericidal
activity via mitochondrial ROS and caspase signaling, suggesting
context-dependent effects. Qi et al (21) proposed that stage-specific
modulation of mitophagy may help manage cytokine storms in early
sepsis while supporting immune recovery later. The current findings
contribute to this framework by highlighting how TPLA modulates
mitophagy-related signaling to balance macrophage function and
inflammation. Thus, TPLA may exert both anti-inflammatory and
immunomodulatory effects through regulation of the
PINK1/Parkin-associated pathway.
Nevertheless, although the present study supports
the protective potential of TPLA against LPS-induced systemic
inflammatory injury, several limitations should be acknowledged.
Although the present study provides pharmacological and
protein-expression evidence supporting the involvement of
PINK1/Parkin-associated mitophagy, direct ultrastructural evidence
of mitophagy was not obtained, as transmission electron microscopy
was not performed to visualize mitophagosomes or
mitochondria-containing autophagic structures. In addition, further
studies using genetic loss-of-function approaches, such as PINK1 or
Parkin knockdown, will be required to more definitively establish
the causal relationship between mitophagy and macrophage
polarization. Future studies should integrate morphological
validation, mitophagy flux assessment, and genetic intervention to
further clarify the mechanism by which TPLA regulates macrophage
function in sepsis-related inflammatory injury. In addition, the
27-day oral administration of TPLA was designed as a pretreatment
regimen and does not mimic the clinical therapeutic window of
established acute sepsis; therefore, the in vivo findings
should be interpreted as protective effects against LPS-induced
systemic inflammatory injury rather than direct therapeutic
efficacy in established sepsis. In vivo validation of
PINK1/Parkin-associated mitophagy and macrophage polarization in
lung and liver tissues was not performed and should be addressed in
future studies. Another limitation is that only male mice were used
in the in vivo experiment. Although this design was intended
to reduce variability associated with sex-dependent hormonal and
immune differences, it may introduce sex-related bias and limit the
generalizability of the findings to female animals. Future studies
should include both male and female mice and evaluate sex as a
biological variable to determine whether the protective effects of
TPLA are sex-dependent.
Acknowledgements
Not applicable.
Funding
The present study was supported by Guangxi Natural Science
Foundation for Young Scholars (grant no. 2024GXNSFBA010302), the
Key Discipline Construction Project of High-Level Traditional
Chinese Medicine under the National Administration of Traditional
Chinese Medicine-Ethnic Minority Medicine (Zhuang Medicine) (grant
no. zyyzdxk-2023165), the Qihuang Project-High-level Talent Team
Training Program of Guangxi University of Chinese Medicine (grant
no. 202414), the Interdisciplinary Innovation Team Project of
Traditional Chinese Medicine in Guangxi (grant no. GZKJ2309), the
Three-Year Action Plan for High-Level Talent Team Construction
(2023) of Guangxi International Zhuang Medicine Hospital (grant no.
GZCX20231203), the ‘Qingmiao Project’ Talent Cultivation Program of
Guangxi International Zhuang Medicine Hospital (grant no. 2022001),
the Research Initiation Project of Guangxi International Zhuang
Medicine Hospital (grant no. 2023GZYJKT008), the National Veteran
Pharmacist Inheritance Studio Construction Project [Issued by the
Department of Human Resources and Education, National
Administration of Traditional Chinese Medicine (2024); grant no.
255].
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
JW performed the in vivo and in vitro
experiments, developed the methodology, organized the raw
experimental data, checked data consistency, conducted the formal
analysis and wrote the original draft. WZ developed the
methodology, performed the in vivo and in vitro
experiments, and performed data validation. GZ performed software
analysis and data visualization, organized and checked the datasets
used for statistical analysis and figure preparation, contributed
to the interpretation of the analytical results, and critically
revised the manuscript for important intellectual content. MH
provided resources, assisted with the in vivo and in
vitro experiments, contributed to data acquisition and
interpretation, and critically revised the manuscript for important
intellectual content. XH conceptualized and supervised the study,
conducted project administration, and wrote, reviewed and edited
the manuscript. JL acquired funding, conceptualized and supervised
the study, developed the methodology, and wrote, reviewed and
edited the manuscript. XH and JL confirm the authenticity of all
the raw data. All authors have read and approved the final version
of the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Animal Ethics
Committee of Guangxi University of Chinese Medicine (approval no.
DW20240923-01; Nanning, China). All animal experiments complied
with the ARRIVE guidelines and the National Institutes of Health
Guide for the Care and Use of Laboratory Animals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
TPLA
|
total phenolics from Laggera
alata
|
|
LPS
|
lipopolysaccharide
|
|
iNOS
|
inducible nitric oxide synthase
|
|
Arg1
|
arginase 1
|
|
PINK1
|
PTEN-induced putative kinase 1
|
References
|
1
|
Modugula S, Altenbaugh M, Ivanova M,
DuMont T and Arshad H: Sepsis epidemiology, definitions, scoring
systems, and diagnostic markers. Crit Care Nurs Q. 48:229–236.
2025. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cheng L, Cao Y, Liu S, Lv L, Zhang J, Bao
J, Wang G and Xu P: Unveiling the research advances of sepsis:
Pathogenesis, precise intervention and clinical perspective. Int J
Surg. 111:6260–6289. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Martin-Loeches I, Singer M and Leone M:
Sepsis: Key insights, future directions, and immediate goals. A
review and expert opinion. Intensive Care Med. 50:2043–2049. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chanderraj R, Admon AJ, He Y, Nuppnau M,
Albin OR, Prescott HC, Dickson RP and Sjoding MW: Mortality of
patients with sepsis administered Piperacillin-tazobactam vs
cefepime. JAMA Intern Med. 184:769–777. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lin SJ, Cheng YY, Chang CH, Lee CH, Huang
YC and Su YC: Traditional Chinese medicine diagnosis ‘Yang-Xu
Zheng’: Significant prognostic predictor for patients with severe
sepsis and septic shock. Evid Based Complement Alternat Med.
2013:7597482013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang GC, Li GQ, Geng HW, Li T, Xu JJ, Ma
F, Wu X, Ye WC and Li YL: Eudesmane-type sesquiterpene derivatives
from Laggera alata. Phytochemistry. 96:201–207. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu YQ, Li N and Wang MW: Research progress
on chemical constituents of Laggera plants in China. Zhongguo Zhong
Yao Za Zhi. 31:181–184. 2006.(In Chinese). PubMed/NCBI
|
|
8
|
Zhou CX, Wu DY, Li XP, Wu YH, Zhao J, Dong
N, Yu RM, Wei W, Zheng QX, Sun HD, et al: Research progress in
Laggera medicinal plants. Zhongguo Zhong Yao Za Zhi. 31:1133–1140.
2006.(In Chinese). PubMed/NCBI
|
|
9
|
Getahun T, Sharma V and Gupta N: The genus
laggera (Asteraceae)-Ethnobotanical and ethnopharmacological
information, chemical composition as well as biological activities
of its essential oils and extracts: A Review. Chem Biodivers.
16:e19001312019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu X, Huang K, Zhang RJ, Mei D and Zhang
B: Isochlorogenic Acid A attenuates the progression of liver
fibrosis through regulating HMGB1/TLR4/NF-κB signaling pathway.
Front Pharmacol. 11:5822020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wei J, Tang Y, Qin S, Ma X, Zhong W, Yang
P, Deng Q and Ma J: Laggera alata attenuates inflammatory
response by regulating macrophage polarization in rheumatoid
arthritis mice. Mol Biotechnol. 66:1934–1941. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li Q, Shi X, Huang H, Gao Q, Sun Q, Meng
Y, Niu L, Xie C and Yang C: 5β-hydroxycostic acid from Laggera
alata ameliorates sepsis-associated acute kidney injury through
its anti-inflammatory and anti-ferroptosis effects via NF-κB and
MAPK pathways. J Ethnopharmacol. 341:1193592025. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
van der Slikke EC, Star BS, van Meurs M,
Henning RH, Moser J and Bouma HR: Sepsis is associated with
mitochondrial DNA damage and a reduced mitochondrial mass in the
kidney of patients with sepsis-AKI. Crit Care. 25:362021.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang S, Long H, Hou L, Feng B, Ma Z, Wu Y,
Zeng Y, Cai J, Zhang DW and Zhao G: The mitophagy pathway and its
implications in human diseases. Signal Transduct Target Ther.
8:3042023. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Picca A, Faitg J, Auwerx J, Ferrucci L and
D'Amico D: Mitophagy in human health, ageing and disease. Nat
Metab. 5:2047–2061. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lu Y, Li Z, Zhang S, Zhang T, Liu Y and
Zhang L: Cellular mitophagy: Mechanism, roles in diseases and small
molecule pharmacological regulation. Theranostics. 13:736–766.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang K, Zhao Q, Sun Y, Lin L and Han X:
Mitochondrial immunometabolism in sepsis: Orchestrating macrophage
polarization and dysfunction. Eur J Med Res. 31:362025. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang W, Wang L, Huang Z, Sun Z and Zheng
B: Peroxiredoxin 3 has a crucial role in the macrophage
polarization by regulating mitochondrial homeostasis. Respir Res.
25:1102024. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang Y, Zhu J, Liu Z, Shu S, Fu Y, Liu Y,
Cai J, Tang C, Liu Y, Yin X and Dong Z: The PINK1/PARK2/optineurin
pathway of mitophagy is activated for protection in septic acute
kidney injury. Redox Biol. 38:1017672021. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xie F, Zhou J, Liu B, Zhao L, Lv C, Zhang
Q, Yuan L, Sun D and Wei W: Low fluoride regulates macrophage
polarization through mitochondrial autophagy mediated by
PINK1/Parkin axis. Biomolecules. 15:6472025. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Qi LY, Xing JX, Ouyang BQ, Li YF and Lei
M: Role of mitophagy affecting macrophage polarization in
immunomodulationin sepsis and traditional Chinese medicine
intervention: A review. Zhongguo Yi Xue Ke Xue Yuan Xue Bao.
46:720–731. 2024.(In Chinese). PubMed/NCBI
|
|
22
|
Reilly B, Tan C, Murao A, Nofi C, Jha A,
Aziz M and Wang P: Necroptosis-mediated eCIRP release in sepsis. J
Inflamm Res. 15:4047–4059. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bojalil R, Ruíz-Hernández A,
Villanueva-Arias A, Amezcua-Guerra LM, Cásarez-Alvarado S,
Hernández-Dueñas AM, Rodríguez-Galicia V, Pavón L, Marquina B,
Becerril-Villanueva E, et al: Two murine models of sepsis:
Immunopathological differences between the sexes-possible role of
TGFβ1 in female resistance to endotoxemia. Biol Res. 56:542023.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Percie du Sert N, Hurst V, Ahluwalia A,
Alam S, Avey MT, Baker M, Browne WJ, Clark A, Cuthill IC, Dirnagl
U, et al: The ARRIVE guidelines 2.0: Updated guidelines for
reporting animal research. PLoS Biol. 18:e30004102020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen Y, Chen X and Zhou Q: Different
effects of a perioperative single dose of dexamethasone on wound
healing in mice with or without sepsis. Front Surg. 10:9271682023.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fang H, Wang X, Damarla M, Sun R, He Q, Li
R, Luo P, Liu JO and Xia Z: Dimethyl Fumarate protects against
lipopolysaccharide-(LPS-) Induced sepsis through inhibition of
NF-κB pathway in mice. Mediators Inflamm. 2023:51335052023.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Davis FM, Schaller MA, Dendekker A, Joshi
AD, Kimball AS, Evanoff H, Wilke C, Obi AT, Melvin WJ, Cavassani K,
et al: Sepsis induces prolonged epigenetic modifications in bone
marrow and peripheral macrophages impairing inflammation and wound
healing. Arterioscler Thromb Vasc Biol. 39:2353–2366. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pang D and Laferriere C: Review of
intraperitoneal injection of sodium pentobarbital as a method of
euthanasia in laboratory rodents. J Am Assoc Lab Anim Sci.
59:3462020.PubMed/NCBI
|
|
30
|
Shomer NH, Allen-Worthington KH, Hickman
DL, Jonnalagadda M, Newsome JT, Slate AR, Valentine H, Williams AM
and Wilkinson M: Review of rodent euthanasia methods. J Am Assoc
Lab Anim Sci. 59:242–253. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang XH, Xu DQ, Chen YY, Yue SJ, Fu RJ,
Huang L and Tang YP: Traditional Chinese Medicine: A promising
strategy to regulate inflammation, intestinal disorders and
impaired immune function due to sepsis. Front Pharmacol.
13:9529382022. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wen Y, Feng C, Chen W, Chen C, Kuang S,
Liu F, Tang Q and Chen M: Effect of traditional Chinese medicine on
serum inflammation and efficacy in patients with sepsis: A
systematic review and meta-analysis. Ann Palliat Med.
10:12456–12466. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen S, Saeed A, Liu Q, Jiang Q, Xu H,
Xiao GG, Rao L and Duo Y: Macrophages in immunoregulation and
therapeutics. Signal Transduct Target Ther. 8:2072023. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Luo M, Zhao F, Cheng H, Su M and Wang Y:
Macrophage polarization: An important role in inflammatory
diseases. Front Immunol. 15:13529462024. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ji Y, Li X, Yao X, Sun J, Yi J, Shen Y,
Chen B and Sun H: Macrophage polarization: Molecular mechanisms,
disease implications, and targeted therapeutic strategies. Front
Immunol. 16:17327182025. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li JM, Li X, Chan LWC, Hu R, Zheng T, Li H
and Yang S: Lipotoxicity-polarised macrophage-derived exosomes
regulate mitochondrial fitness through Miro1-mediated mitophagy
inhibition and contribute to type 2 diabetes development in mice.
Diabetologia. 66:2368–2386. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mohsin M, Zaki A, Tabassum G, Khan S, Ali
S, Ahmad T and Syed MA: Urolithin-A supplementation alleviates
sepsis-induced acute lung injury by reducing mitochondrial
dysfunction and modulating macrophage polarization. Mitochondrion.
84:1020472025. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tang Z, Ning Z and Li Z: The beneficial
effects of Rosuvastatin in inhibiting inflammation in sepsis. Aging
(Albany NY). 16:10424–10434. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Onishi K, Fu HY, Sofue T, Tobiume A,
Moritoki M, Saiga H, Ohmura-Hoshino M, Hoshino K and Minamino T:
Galectin-9 deficiency exacerbates lipopolysaccharide-induced
hypothermia and kidney injury. Clin Exp Nephrol. 26:226–233. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yang M, Wei X, Yi X and Jiang DS:
Mitophagy-related regulated cell death: Molecular mechanisms and
disease implications. Cell Death Dis. 15:5052024. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Han R, Liu Y, Li S, Li XJ and Yang W:
PINK1-PRKN mediated mitophagy: Differences between in vitro and in
vivo models. Autophagy. 19:1396–1405. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Su L, Zhang J, Gomez H, Kellum JA and Peng
Z: Mitochondria ROS and mitophagy in acute kidney injury.
Autophagy. 19:401–414. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Choudhuri S, Chowdhury IH and Garg NJ:
Mitochondrial regulation of macrophage response against pathogens.
Front Immunol. 11:6226022020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Liu B, Cao Y, Wang D, Zhou Y, Zhang P, Wu
J, Chen J, Qiu J and Zhou J: Zhen-Wu-Tang Induced mitophagy to
protect mitochondrial function in chronic glomerulonephritis via
PI3K/AKT/mTOR and AMPK pathways. Front Pharmacol. 12:7776702021.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang X, Du J, Li B, Huo S, Zhang J, Cui
Y, Song M, Shao B and Li Y: PINK1/Parkin-mediated mitophagy
mitigates T-2 toxin-induced nephrotoxicity. Food Chem Toxicol.
164:1130782022. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bhatia D, Chung KP, Nakahira K, Patino E,
Rice MC, Torres LK, Muthukumar T, Choi AM, Akchurin OM and Choi ME:
Mitophagy-dependent macrophage reprogramming protects against
kidney fibrosis. JCI Insight. 4:e1328262019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhang Y, Chen L, Luo Y, Wang K, Liu X,
Xiao Z, Zhao G, Yao Y and Lu Z: Pink1/Parkin-mediated mitophagy
regulated the apoptosis of dendritic cells in sepsis. Inflammation.
45:1374–1387. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Orekhov AN, Zhuravlev AD, Vinokurov AY,
Nikiforov NG, Omelchenko AV, Sukhorukov VN, Sinyov VV and Sobenin
IA: Defective mitophagy impairs response to inflammatory activation
of Macrophage-Like cells. Curr Med Chem. 32:111–122. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Patoli D, Mignotte F, Deckert V, Dusuel A,
Dumont A, Rieu A, Jalil A, Van Dongen K, Bourgeois T, Gautier T, et
al: Inhibition of mitophagy drives macrophage activation and
antibacterial defense during sepsis. J Clin Invest. 130:5858–5874.
2020. View Article : Google Scholar : PubMed/NCBI
|