Introduction
Feeding intolerance (FI) is a prevalent and
clinically significant complication in the intensive care unit
(ICU), adversely affecting a substantial proportion of critically
ill patients. Reported incidence rates range from 30 to 70%
globally, with a marked variation across geographic populations. A
study from European and North American cohorts have reported
incidences at the higher end of this range (50–70%) in adult
critically ill populations, whereas data from Asian populations
suggest relatively lower rates (30–50%) (1). A systematic review focusing on
critically ill children reported a prevalence range of 22–65%, with
similar geographic variations observed across the included studies
(2). This heterogeneity reflects
differences in patient populations, diagnostic criteria and
potentially underlying gut microbiota composition influenced by
geographic origin and dietary habits (3,4).
Most cases occur during enteral nutrition administration within the
first week of ICU admission or during the first 12 days following
ICU admission (1,2). Clinically, FI is characterized by
manifestations such as elevated gastric residual volumes (GRVs),
vomiting, abdominal distension and diarrhea, all of which
compromise the effective delivery of enteral nutrition (EN).
Failure to achieve prescribed caloric and protein targets
represents more than a nutritional deficiency; it is independently
associated with significant adverse clinical outcomes. A growing
body of evidence has demonstrated strong associations between FI
and increased rates of infectious complications, prolonged
mechanical ventilation, extended ICU and hospital lengths of stay,
increased health care costs and increased mortality (5–8). In
a systematic review, Gungabissoon et al (9) identified FI as a major contributor to
inadequate EN delivery and poorer clinical outcomes. Additionally,
Padar et al (6) reported
that gastrointestinal failure, considered a severe manifestation of
FI, independently predicts mortality in critically ill adults.
Historically, the pathophysiology of FI was
oversimplified and attributed primarily to impaired
gastrointestinal motility. However, the contemporary paradigm is
evolving to recognize FI as a complex, multifactorial disorder
intricately linked to the microbiome-gut-brain axis (10,11).
In a state of health, this sophisticated bidirectional
communication system ensures coordinated gastrointestinal function,
including motility, secretion and barrier integrity. Critical
illness, however, precipitates a profound collapse of this
homeostatic equilibrium. The ‘gut-critical illness nexus’ (defined
here as the complex bidirectional interplay between critical
illness pathophysiology and gut microbial ecology) is characterized
by a rapid and dramatic shift from a symbiotic, diverse microbiota
to a state of severe gut dysbiosis (12–14).
This dysbiosis is not a passive consequence but an active driver of
local and systemic organ dysfunction. A hallmark of this shift is
the marked depletion of commensal, obligate anaerobic bacteria,
particularly those belonging to the Clostridium clusters IV
and XIVa (such as Faecalibacterium prausnitzii, Roseburia
spp.), which are fundamental producers of beneficial metabolites
(13,15). This microbial collapse is driven by
the dual assault of the critical illness itself (e.g., systemic
inflammation, shock, sepsis) and ubiquitous iatrogenic insults,
most notably broad-spectrum antibiotics and proton pump inhibitors
(PPIs) (16–18). The functional consequence of this
dysbiosis is a critical deficit in microbial-derived metabolites,
with the short-chain fatty acid (SCFA) butyrate being of paramount
importance.
The critical illness-induced gut dysbiosis leads to
a functional deficit in microbial-derived butyrate, which serves as
a key pathophysiological predictor and mediator of FI. Butyrate,
produced primarily by the aforementioned bacterial clusters via the
fermentation of dietary fiber, is a keystone metabolite with
pleiotropic functions essential for gut health (14,19).
It serves as the primary energy source for colonocytes, thereby
reinforcing the intestinal epithelial barrier through the
regulation of tight junction proteins (20,21).
It exerts potent anti-inflammatory and immunomodulatory effects,
largely through the inhibition of histone deacetylases (HDAC) and
the induction of regulatory T cells (22,23).
Furthermore, butyrate directly modulates gastrointestinal motility
and blood flow through interactions with the enteric nervous system
and the promotion of hormonal secretion (24,25).
The loss of butyrate therefore creates a pathophysiological
trifecta of impaired barrier function, dysregulated immunity and
disrupted motility, a perfect storm that culminates in the clinical
syndrome of FI (26–28). Supporting this, studies have
demonstrated a quantitative loss of fecal and systemic butyrate in
critically ill patients, with this depletion correlating with
adverse gastrointestinal outcomes (19,29).
Deciphering this complex sequence of events requires
tools that move beyond traditional clinical observations. The
emergence of multi-omics technologies, including microbiomics (to
profile microbial community structure), metabolomics (to quantify
metabolites like butyrate) and metagenomics (to assess the
functional genetic potential for butyrogenesis), provides an
unprecedented lens through which to elucidate the etiology of FI
(30,31). These integrated approaches allow
for a systems-level understanding, providing evidence for strong
associations that may inform our understanding of potential causal
mechanisms. For example, Wijeyesekera et al (26) utilized multi-compartment
metabolomics in critically ill children to identify intestinal
dysbiosis and its functional consequences, directly linking
microbial metabolite perturbations to clinical status. Similarly,
longitudinal studies have shown that the loss of butyrate synthesis
pathways in the gut metagenome precedes and is strongly associated
with the development of FI and other complications (30,32).
Translating this mechanistic, multi-omics evidence
into improved patient outcomes hinges on effective bedside
implementation. In this context, critical care nurses are uniquely
positioned to lead this translational effort. Their continuous
presence at the bedside, primary responsibility for EN
administration and monitoring (including the controversial practice
of GRV measurement) (33–35) and holistic approach to patient care
make them the ideal agents for deploying microbiota-supportive
interventions (36–38). From advocating for antibiotic
stewardship and administering pre/pro/synbiotics to integrating
microbiome-informed metrics into gastrointestinal assessments,
nurses are the pivotal link between scientific discovery and
clinical practice. The move away from routine GRV monitoring, as
supported by recent meta-analyses (39,40)
and successfully implemented via nurse-driven protocols (34,38),
exemplifies how nursing practice can evolve based on evidence that
FI is more than a simple motility issue.
In comparison to existing published reviews on gut
microbiota and critical illness, the present review offers several
distinctive contributions. First, while previous reviews have
broadly described dysbiosis in the ICU, the present review uniquely
synthesizes recent multi-omics evidence, integrating microbiomic,
metabolomic and metagenomic datasets, to propose a specific,
testable pathogenic pathway whereby functional butyrate depletion
directly predicts the development of feeding intolerance (14,19).
Second, beyond mechanistic synthesis, the present review translates
ecological principles into a structured, nurse-driven
implementation framework that operationalizes microbiota support
across four actionable domains: Harm minimization, targeted
nourishment, direct restoration and innovative monitoring. This
pragmatic focus on nursing-led protocols addresses a significant
gap in the literature, where microbiome science has rarely been
integrated into clinical workflows with this level of specificity.
Third, unlike purely descriptive syntheses, the present review
critically appraises inconsistencies in the evidence base,
including geographic variations in microbiota resilience, host
genetic factors [e.g., monocarboxylate transporter-1 (MCT-1)
polymorphisms] and methodological heterogeneity across multi-omics
studies, thereby providing a balanced foundation for future
investigation.
The primary objective of the present review was to
synthesize the current multi-omics evidence that suggests a
plausible pathway from gut dysbiosis to butyrate loss and
subsequent FI. Furthermore, it aims to critically appraise this
evidence to propose and frame a practical, nurse-driven
intervention model designed to preserve and restore a healthy gut
microbiota, thereby predicting and preventing FI in the vulnerable
critically ill population. By integrating insights from microbial
ecology, molecular metabolism and clinical nursing science, the
present review sought to guide the development of targeted,
evidence-based protocols that address the potential root cause of
this common and debilitating condition.
The gut-critical illness interaction: From
homeostasis to dysbiosis
Under physiological conditions, the human gut
operates as a tightly regulated ecosystem whose metabolic output,
particularly microbiota-derived butyrate, underpins intestinal
barrier integrity, motility and systemic immune homeostasis.
Critical illness, however, precipitates a rapid and quantifiable
collapse of this symbiosis, converting a diverse, anaerobe-rich
community into a low-diversity, pathogen-laden consortium with
markedly impaired SCFA biosynthesis (Fig. 1). The following sections dissect
the temporal trajectory of this disruption, beginning with the
pre-morbid microbial blueprint, tracing the differential impact of
ICU-specific insults, and culminating in the functional and
clinical sequelae that set the stage for FI.
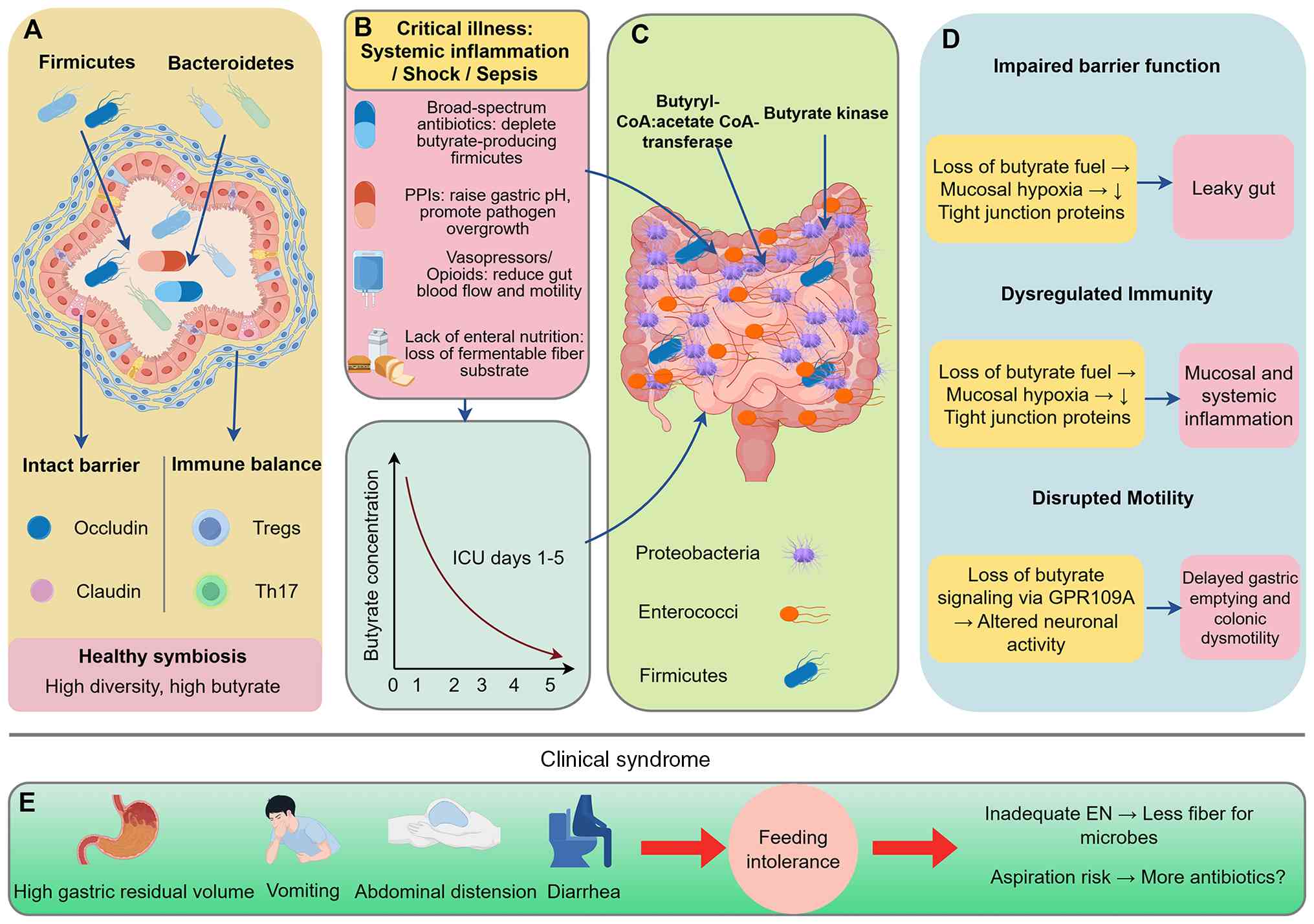 | Figure 1.The pathophysiological trajectory
linking critical illness-induced gut dysbiosis to FI. (A) In
health, a diverse, anaerobe-rich microbiota produces ample
butyrate, sustaining barrier integrity, immunotolerance and
motility. (B) ICU admission delivers a multifactorial assault
(illness severity, antibiotics, PPIs) that within days collapses
this ecosystem, (C) depleting butyrate producers and their genes.
(D) The resulting butyrate deficit drives a triad of barrier
breakdown, immune dysregulation and dysmotility, which (E)
clinically manifests as FI. This FI can then exacerbate dysbiosis,
creating a treatment-resistant vicious cycle. The image was
generated by Figdraw (version 2.0; www.figdraw.com; ID:PAWAR8a447). ICU, intensive care
unit; FI, feeding intolerance; PPIs, proton pump inhibitors; EN,
enteral nutrition. |
Pre-illness ecosystem: Composition and
metabolic signature
Healthy adults harbor a dense, anaerobe-dominated
community in which Firmicutes (notably Clostridium clusters
IV and XIVa) and Bacteroidetes predominate (15,41).
These taxa express a broad repertoire of butyrogenic genes that
convert dietary fiber into millimolar concentrations of butyrate,
propionate and acetate (26,42).
Butyrate fulfils three homeostatic tasks: It i) fuels colonocytes
via β-oxidation, thereby maintaining tight-junction integrity; ii)
acts as a histone-deacetylase inhibitor that expands peripheral
regulatory T cells; and iii) triggers enteric neuronal
5-hydroxytryptamine release that coordinates segmental motility
(43–45). A recent multi-center metagenomic
survey of 167 healthy volunteers showed that individuals in the
highest quartile of fecal butyrate (≥12 mmol/kg) displayed lower
systemic interleukin-6 (IL-6) and higher zonula-occludens-1
expression in rectal biopsies, underscoring the anti-inflammatory
and barrier-stabilizing properties of the metabolite (26).
ICU insults precipitating
dysbiosis
Within 24–48 h of critical illness, the symbiotic
architecture collapses. In three independent ICU cohorts (n=34-115
patients), a ≥30% reduction in observed species richness and a
≥10-fold drop in butyrate concentration during the first week of
admission were documented (15,41,46).
Proteobacteria (Escherichia, Klebsiella, Enterococcus)
expand from <5 to 30–50% of total reads, whereas in the study by
Ravi et al (15), butyrate
producers such as Faecalibacterium prausnitzii and
Roseburia spp. fell below the limit of detection in 60% of
subjects. Both prospective (41)
and retrospective (46) studies
identified identical predictors of this shift: Broad-spectrum
β-lactams, PPIs and vasopressor infusion. Notably, Lamarche et
al (41) demonstrated that
every additional day of meropenem correlated with a 2.3% loss in
Shannon diversity per day (P<0.001) and a 0.15 mmol/l decline in
serum butyrate.
Drug-specific mechanisms of microbe
loss
Antibiotics exert class-dependent collateral damage
on the gut ecosystem. Metagenomic analysis of 21 critically ill
adults revealed that meropenem selectively depletes Firmicutes
carrying the butyryl-CoA:acetate CoA-transferase gene, while
vancomycin eradicates commensal Gram-positive taxa without
compensatory butyrate recovery (47). PPIs amplify the effect by raising
gastric pH >4, permitting oral Streptococci and
Candida spp. to reach the colon and out-compete anaerobic
fiber degraders (48,49). In a double-blind randomized
controlled trial involving 48 healthy volunteers, 7-day
pantoprazole decreased fecal butyrate by 38% and increased fungal
internal transcribed spacer 1 copies 5-fold; these changes were
reversed within 4 weeks of drug cessation, indicating a transient
yet quantifiable perturbation (49).
Functional consequences beyond
butyrate depletion
The metabolic vacuum left by fiber-fermenting
bacteria is filled by opportunistic pathogens that exploit
ethanolamine and mucin-derived sugars. Elevated proteolytic
metabolites (p-cresol, indoxyl sulfate) and reduced SCFA synergize
to impair epithelial oxygen consumption, resulting in a ‘leaky’
phenotype (46,50). Using ex-vivo Using chambers,
Chernevskaya et al (46)
showed that serosal-to-mucosal permeability doubled in biopsy
samples collected from septic patients with low butyrate (<2
mmol/kg) compared with ICU controls with preserved levels.
Consistently, animal data confirm that antibiotic-induced dysbiosis
decreases claudin-1 and occludin expression via HDAC3-mediated
transcriptional repression, a defect rescued by oral butyrate
supplementation (45,51).
Clinical correlates: FI and
beyond
A total of three observational studies have linked
the magnitude of dysbiosis to subsequent gastrointestinal
complications. In 115 mechanically ventilated adults, the relative
abundance of butyrate producers on ICU day 3 predicted FI (GRV
>250 ml) with an area under the receiver operating
characteristic curve (AUROC) of 0.81 (95% CI 0.73–0.89) (15). Similarly, a pediatric critical-care
cohort study found that loss of microbial diversity preceded the
first episode of emesis by a median of 2 days, suggesting that
microbiota disruption is not merely associative but temporally
antecedent (26). Importantly, the
same datasets revealed that patients in the lowest quartile of
butyrate synthesis capacity had a 2.4-fold higher hazard of 28-day
mortality, implicating gut-derived metabolite deficiency as a
contributor to systemic decompensation (15,41).
Despite converging evidence, important
inconsistencies persist. Geographic origin, baseline diet and
ethnicity moderate the resilience of the microbiome; Asian
populations appear to retain higher levels of butyrate producers
under comparable antibiotic pressure (3). Furthermore, most reports are limited
to 16S rRNA surveys; only two studies to date have integrated
metagenomic and metabolomic layers to assign functional gene loss
to specific taxa (47,52). Finally, inter-individual variation
in host toll-like receptor signaling and bile-acid composition may
confound the speed and extent of dysbiosis, underscoring the need
for personalized profiling before implementing microbiota-targeted
interventions (52,53).
Collectively, these iatrogenic and physiological
stressors orchestrate a shift from homeostasis to a maladaptive
dysbiotic state, creating a vicious cycle of metabolite depletion
and epithelial dysfunction. Fig. 1
illustrates this cumulative pathophysiological trajectory from ICU
admission to the manifestation of FI, depicting the stepwise
transition from a healthy butyrate-producing microbiota through the
collapse of microbial ecology and butyrate deficit to clinical
feeding intolerance and a self-perpetuating vicious cycle.
Butyrate: A keystone metabolite in gut
health and systemic homeostasis
To date, accumulating multi-omics studies have
validated the correlation between specific microbial signatures,
particularly the depletion of butyrate producers, and clinical
feeding outcomes. Key observational studies establishing this link
are summarized in Table I.
Butyrate, a four-carbon SCFA generated by microbial fermentation of
dietary fiber, occupies a central position in the maintenance of
intestinal and systemic homeostasis. Its concentration in the
healthy colonic lumen (0.2–2 mmol/l) reflects the collective
butyrogenic capacity of a restricted group of Firmicutes, and even
modest reductions in this metabolite have been linked to increased
mucosal permeability, exaggerated inflammation and delayed gastric
emptying.
 | Table I.Key multi-omics studies linking
butyrate-producing microbiota and butyrate levels to gut health and
feeding intolerance. |
Table I.
Key multi-omics studies linking
butyrate-producing microbiota and butyrate levels to gut health and
feeding intolerance.
| Author(s),
year | Study type | Study
population/model | Key findings
related to butyrate producers or butyrate levels | Association with FI
or clinical gastrointestinal outcomes | Multi-omics methods
applied | (Refs.) |
|---|
| Wijeyesekera et
al, 2019 | Observational
cohort | Critically ill
children (ICU) | Multi-compartment
metabolomics identified depletion of Firmicutes (Clostridium
clusters IV/XIVa) and reduced fecal butyrate. Butyrate levels
correlated inversely with systemic inflammation and barrier
dysfunction. | Intestinal
dysbiosis and metabolite perturbations were directly linked to
clinical gastrointestinal dysfunction status. | Microbiomics (16S
rRNA sequencing), Metabolomics (NMR, MS) | (26) |
| Su et al,
2020 |
Cross-sectional | Patients with
Grave's disease | Gut dysbiosis and
reduced butyrate/propionate were associated with Treg/Th17
imbalance | Indirectly supports
the role of butyrate in immune regulation, though not directly
studied in FI. | Metagenomics,
Metabolomics | (55) |
| Lamarche et
al, 2018 | Prospective
observational | Mechanically
ventilated adults (ICU) | Each additional day
of broad-spectrum antibiotic (meropenem) correlated with a 2.3%
daily loss of microbial Shannon diversity and a 0.15 mmol/l decline
in serum butyrate. | Microbial diversity
loss and butyrate depletion were associated with increased
mortality. | Microbiomics (16S
rRNA sequencing), Serum metabolomics | (41) |
| Ravi et al,
2019 | Observational
cohort | Critically ill
adults (ICU) | Relative abundance
of butyrate producers on ICU day 3 predicted subsequent feeding
intolerance (GRV >250 ml) with an AUROC of 0.81. | Direct predictive
link between early loss of butyrate-producing taxa and later
development of FI. | Microbiomics (16S
rRNA sequencing) | (15) |
| Chernevskaya et
al, 2020 | Observational
pilot | Septic patients
(ICU) | Patients with low
serum butyrate (<2 mmol/kg) exhibited a two-fold increase in
intestinal permeability (ex-vivo) compared to ICU controls
with preserved butyrate. | Linked butyrate
depletion directly to impaired intestinal barrier function,a key
patho- physio logical feature of FI. | Serum & Fecal
Metabolomics, Functional assay (Using chamber) | (46) |
| Maier et al,
2021 |
Experimental/mechanistic | In vitro
& in vivo antibiotic exposure models | Metagenomic
analysis revealed that specific antibiotics (e.g., meropenem)
selectively depletedFirmicutes carrying the butyryl-CoA:acetate
CoA-transferase gene, a key butyrate synthesis pathway. | Demonstrated a
mechanism for antibiotic-induced functional loss of butyrogenic
capacity beyond taxonomic shifts. | Metagenomics,
Culturing | (47) |
Biosynthetic routes and principal
microbial producers
Butyrate is generated almost exclusively by
anaerobic fermentation of dietary fibers and resistant starch.
Metagenomic analyses of healthy adults consistently map >80% of
fecal butyrogenic potential to three Firmicutes lineages:
Faecalibacterium prausnitzii, Roseburia spp. and
Eubacterium rectale (54–56).
These taxa express either the butyryl-CoA:acetate-CoA-transferase
or the butyrate-kinase pathway; both routes converge on
butyryl-CoA, which is then converted to butyrate via phosphate
butyryl-transferase (56). In a
case-control study comparing 58 patients with Graves' disease and
63 healthy controls, Su et al (55) observed that the relative abundance
of butyrate-producing genera, including Faecalibacterium and
Roseburia, was significantly reduced in the patient group
and positively associated with fecal short-chain fatty acid
(including butyrate) concentrations. This correlation reinforces
the quantitative link between the density of these microbial
clusters and their metabolic output in the human gut, as a loss of
producer taxa directly paralleled a decline in metabolite
levels.
Enterocyte energy substrate and
barrier reinforcement
Once released into the colonic lumen, butyrate is
rapidly taken up by colonocytes through MCT-1 and sodium-coupled
MCT. Inside the cell it undergoes β-oxidation, supplying ≥70% of
basal oxygen consumption and maintaining hypoxic niche conditions
that suppress pathogen expansion (57,58).
Mechanistic work in Caco-2 monolayers demonstrated that 2 mmol/l
butyrate increases transepithelial electrical resistance by 35%
within 6 h, an effect mediated by AMP-activated protein
kinase-dependent phosphorylation of tight-junction proteins
occludin and claudin-1 (58).
Consistently, a randomized trial in antibiotic-induced dysbiosis
revealed that 4-week oral sodium butyrate (1 g/day) restored the
urinary lactulose-to-mannitol ratio to baseline values while
raising colonic zonula occludens-1 mRNA 2.3-fold (59). These observations position butyrate
as the principal metabolic fuel for epithelial renewal and
paracellular sealing.
Anti-inflammatory and immunoregulatory
actions
Beyond energetics, butyrate functions as a potent
epigenetic regulator. Its inhibition of HDAC1/2/3 promotes
acetylation of promoter regions for forkhead box P3 (FOXP3) and
IL-10, thereby expanding peripheral regulatory T cells (Treg) and
dampening type 17 T-helper cell polarization (54,60,61).
In a gnotobiotic mouse model colonized with butyrate-producing
Roseburia hominis, lamina propria FOXP3+ cells increased
from 8 to 22% and IL-17+ cells fell by 40% relative to germ-free
controls (62). Translationally,
with the caveat that these data derive from inflammatory bowel
disease rather than critical illness, a double-blind study in
ulcerative colitis showed that enema-delivered butyrate (100
mmol/l, 14 days) raised mucosal IL-10 concentration 3-fold and
decreased tumor necrosis factor α and IL-6 by 50%, paralleling
clinical remission in 65% of recipients (63). Collectively, these data indicate
that butyrate orchestrates a tolerogenic milieu through HDAC
inhibition and subsequent Treg expansion.
Modulation of gastrointestinal
motility and secretion
Butyrate also interacts with the enteric nervous
system to regulate motility. Electrophysiological recordings from
murine colonic segments revealed that 5 mmol/l butyrate depolarizes
cholinergic interneurons via G protein-coupled receptor 109A
signaling, increasing acetylcholine release and promoting
high-amplitude propagating contractions (60). Clinically, a pilot trial in
critically ill adults demonstrated that enteral infusion of sodium
butyrate (4 g/day for 7 days) shortened gastric emptying time (T½)
from 180 to 120 min (P=0.02) and reduced GRV >250 ml episodes by
45% (64). While these motility
benefits are encouraging, heterogeneity in dose (0.5–4 g), route
(oral vs. rectal) and patient phenotype precludes firm dosing
recommendations; nevertheless, the consistency across mechanistic
and early-phase studies supports a causal role for butyrate in
accelerating gut transit.
Despite converging evidence, several caveats should
be noted. First, most mechanistic insights derive from
supra-physiological concentrations (2–10 mmol/l) that exceed portal
levels recorded in healthy humans (0.2–1 mmol/l) (55,56).
Second, host genetics and diet modify responsiveness: Individuals
carrying MCT-1 loss-of-function variants achieve lower
intracellular butyrate and derive weaker barrier protection
(65). Third, comparative studies
reveal that propionate and acetate share certain immunomodulatory
properties, raising the possibility that synergistic SCFA mixtures,
not butyrate alone, mediate observed benefits (66). Future dose-response trials
integrating metagenomics, metabolomics and transcriptomics are
warranted to define minimal effective concentrations and to
identify responders most likely to benefit from butyrate-centric
therapeutics.
Multi-omics evidence linking dysbiosis,
butyrate depletion and FI
Multi-omics evidence published within the last
decade has begun to delineate a coherent axis linking gut
dysbiosis, functional loss of butyrate synthesis and the
development of FI in critically ill adults and children (Table II). Integrating microbiomic,
metabolomic and metagenomic data obtained from ICU cohorts, murine
sepsis models and randomized nutrition trials reveals a sequential
trajectory: Rapid contraction of butyrate-producing Firmicutes,
quantitative decline in luminal and systemic butyrate and
concomitant impairment of intestinal motility, barrier function and
local immunity. The following sections critically appraise these
independent yet complementary layers of evidence, highlight
methodological consistencies and discrepancies and evaluate their
collective robustness in establishing causality between
microbiota-derived butyrate depletion and FI.
| Table II.Multi-omics evidence linking gut
dysbiosis, butyrate depletion, and feeding intolerance in
critically ill patients. |
Table II.
Multi-omics evidence linking gut
dysbiosis, butyrate depletion, and feeding intolerance in
critically ill patients.
| Author(s),
year | Study design |
Population/model | Key
microbiome/metabolome findings | Functional genomics
findings (butyrate-related) | Clinical
association with feeding intolerance | (Refs.) |
|---|
| Wijeyesekera et
al, 2019 | Prospective
observational | Critically ill
children | Depletion of
Firmicutes, especially Faecalibacterium prausnitzii and
Roseburia spp.; reduced fecal SCFAs, particularly
butyrate. | Not explicitly
assessed. | Temporal link
between loss of butyrate-producing taxa and onset of
gastro-intestinal dysfunction. | (26) |
| Wu et al,
2020 | Experimental
(murine model) | Klebsiella
pneumoniae-induced pneumosepsis | Decreased cecal
butyrate; overgrowth of pathogenic taxa and reduction in
fiber-fermenting bacteria. | Not assessed. | Butyrate decline
preceded systemic inflammation and gastrointestinal
dysmotility. | (27) |
| Zhou et al,
2023 | Observational
cohort | Mechanically
ventilated adults | Decreased microbial
genes involved in SCFA metabolism, including butyrate production;
linked to gutlung axis disruption. | Metagenomic
dysfunction in butyrate pathways associated with 28-day
mortality. | Impaired microbial
metabolic capacity correlated with worse clinical outcomes,
including gastrointestinal intolerance. | (32) |
| Zhang et al,
2022 | Longitudinal
observational | COVID-19
patients | Prolonged reduction
in butyrate-producing bacteria; impaired butyrate biosynthesis
persisted post-acute phase. | Metagenomic
analysis indicated decreased butyrate synthesis potential. | Associated with
gastrointestinal symptoms and prolonged gut dysfunction. | (67) |
| Valdés-Duque et
al, 2020 | Case-control | Septic ICU
patients | Significantly lower
fecal butyrate levels vs. healthy controls; inverse correlation
with intestinal permeability markers. | Not assessed. | Butyrate depletion
correlated with increased intestinal barrier disruption and
systemic inflammation. | (69) |
| Haak et al,
2021 | Integrative
transkingdom analysis | ICU patients | Reduction in genes
encodingbutyryl-CoA:acetate CoA-transferase and butyrate kinase;
association with antibiotic exposure. | Functional loss of
butyrate synthesis pathways linked to dysbiosis and clinical
decline. | Predictive of
adverse clinical outcomes; supports functional gene loss as a
mediator of gastrointestinal dysmotility. | (70) |
Microbiomic signatures: Loss of
butyrate-producing taxa in critically ill patients
Recent microbiome analyses have consistently
revealed a significant reduction in butyrate-producing bacteria in
critically ill patients, particularly those developing FI.
Wijeyesekera et al (26)
conducted a multi-compartment metabolomic and microbiomic study in
pediatric ICU patients and found that the depletion of
Firmicutes, particularly Clostridium clusters IV and
XIVa (e.g., Faecalibacterium prausnitzii, Roseburia spp.),
was strongly associated with intestinal dysbiosis and systemic
metabolic disturbances. These taxa are known to harbor
butyryl-CoA:acetate CoA-transferase and butyrate kinase pathways,
which are essential for butyrate biosynthesis. Their reduction was
temporally linked with the onset of gastrointestinal dysfunction,
suggesting a potential causal relationship rather than mere
association.
Although COVID-19 pathophysiology differs from
general critical illness in its specific inflammatory and
immunological features, it provides proof-of-principle that
butyrate loss can persist beyond the acute phase; similarly, Zhang
et al (67) reported that
COVID-19 patients exhibited prolonged impairment in SCFA
biosynthesis, particularly butyrate, due to a persistent decline in
butyrogenic bacteria. This dysbiosis was not transient and
persisted beyond the acute phase of illness, implying that
microbiota disruption may have long-term consequences on gut
function and immunity. These findings align with earlier
observations in sepsis patients, where a significant reduction in
butyrate producers was associated with increased mucosal
inflammation and impaired gastrointestinal motility (68).
Metabolomic corroboration:
Quantitative decline in fecal and systemic butyrate
Metabolomic profiling has provided direct evidence
of butyrate depletion in critically ill populations. Valdés-Duque
et al (69) quantified
stool SCFAs in septic ICU patients and found significantly lower
levels of butyrate compared to healthy controls. This reduction was
inversely correlated with markers of intestinal permeability and
systemic inflammation, reinforcing the role of butyrate in
maintaining mucosal integrity and immune homeostasis.
In a murine model of Klebsiella
pneumoniae-induced pneumosepsis, Wu et al (27) observed a marked decrease in cecal
butyrate levels, coinciding with an overgrowth of pathogenic taxa
and a reduction in beneficial fiber-fermenting bacteria.
Importantly, this study also demonstrated that butyrate depletion
was closely associated with the onset of systemic inflammatory
responses, suggesting that microbial metabolite deficiency may be
an early driver rather than a consequence of critical illness.
However, the original study did not report a specific time interval
between these events (27). These
findings are consistent with human studies showing that fecal
butyrate levels are significantly reduced in patients with EN
intolerance and are predictive of worse clinical outcomes (26).
Functional genomics: Loss of
butyrogenic pathways in the gut metagenome
Beyond taxonomic shifts, metagenomic analyses have
revealed a functional collapse in butyrate synthesis pathways
during critical illness. Haak et al (70) performed an integrative transkingdom
analysis in ICU patients and identified a significant reduction in
genes encoding butyryl-CoA:acetate CoA-transferase and butyrate
kinase, key enzymes in butyrate biosynthesis. This functional gene
loss was most pronounced in patients exposed to broad-spectrum
antibiotics and those with prolonged ICU stays, indicating that
iatrogenic factors may exacerbate microbiota dysfunction.
Furthermore, Zhou et al (32) demonstrated that the gut-lung axis
disruption in mechanically ventilated patients was associated with
a reduction in microbial genes involved in SCFA metabolism,
including butyrate production. This metagenomic dysfunction was
predictive of 28-day mortality, underscoring the clinical relevance
of microbial metabolic capacity beyond taxonomic composition. These
data collectively support the hypothesis that the loss of
butyrogenic function, rather than simply the absence of specific
taxa, is a critical determinant of gastrointestinal dysmotility and
FI.
Integrative interpretation: From
dysbiosis to clinical phenotype
The convergence of microbiomic, metabolomic and
metagenomic evidence points to a coherent pathway wherein critical
illness-induced dysbiosis leads to butyrate depletion, which in
turn compromises intestinal barrier integrity, immune regulation
and motility, culminating in FI. However, some inconsistencies
merit discussion. For instance, geographic and dietary factors may
modulate microbiota resilience, with certain populations (e.g.,
Asian cohorts) retaining higher levels of butyrate producers
despite antibiotic exposure (3).
Additionally, host genetics, such as MCT-1 polymorphisms affecting
butyrate uptake, may influence individual susceptibility to FI
(65).
Furthermore, while most studies report a consistent
decline in butyrate and its producers, a small number of studies
have noted partial recovery of SCFA levels following probiotic or
synbiotic interventions, albeit with variable clinical efficacy
(71,72). This suggests that
microbiota-targeted therapies may hold promise, but their success
likely depends on the timing, baseline microbiota composition and
host metabolic context.
Nurse-driven microbiota-supportive
interventions: From evidence to action
Emerging multi-omics evidence identifies the
depletion of butyrate-producing gut flora as a key driver of FI in
critically ill patients. To bridge the gap between mechanistic
insights and bedside application, a structured, nurse-driven
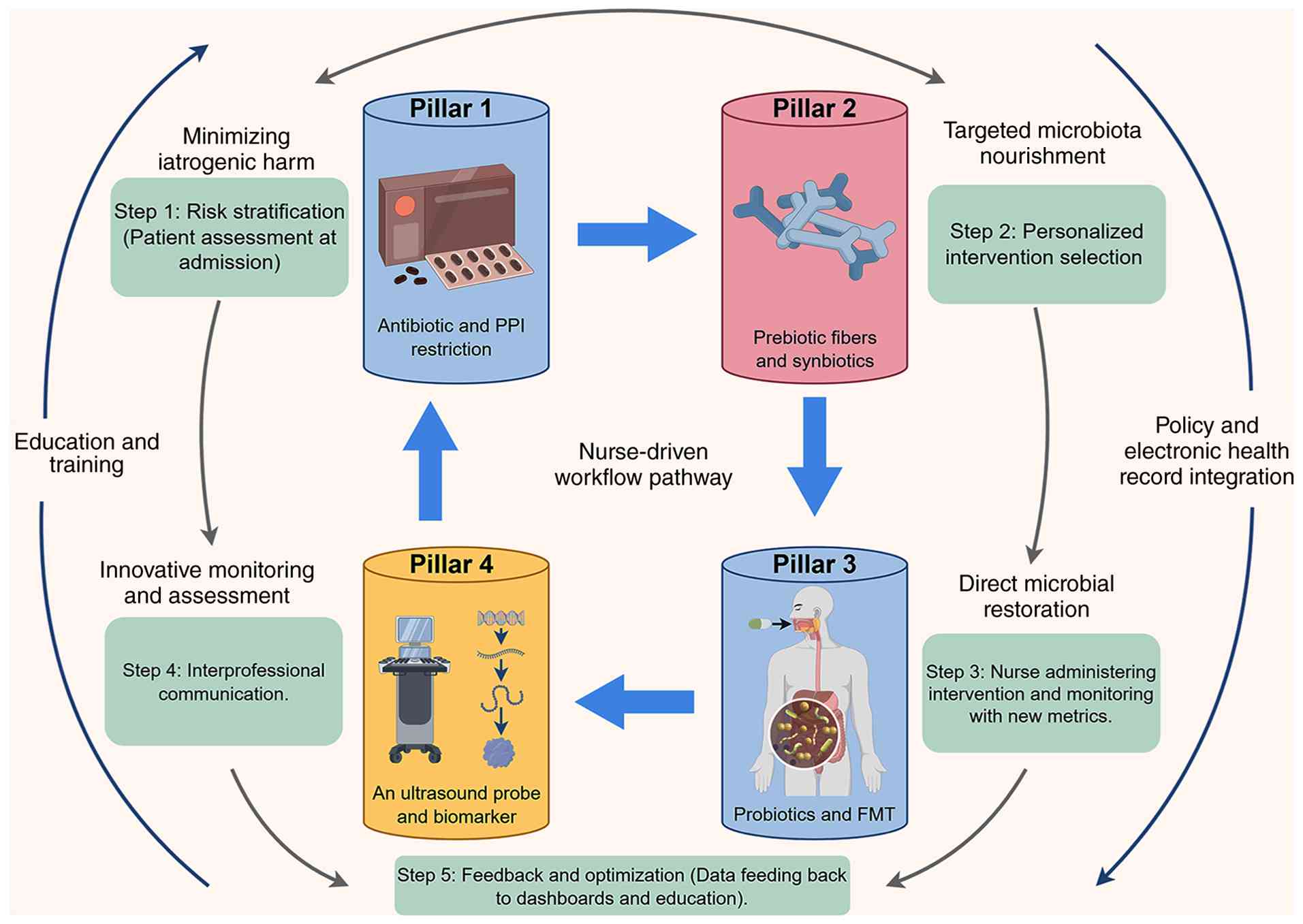
intervention framework (Fig. 2)
was provided in the present study, grounded in four core domains:
Harm minimization, targeted nourishment, microbial restoration and
dynamic monitoring (Table III).
As shown in Fig. 2, this framework
effects four core domains: Harm minimization (such as
antibiotic/PPI stewardship), targeted nourishment
(prebiotics/synbiotics), microbial restoration (probiotics/FMT),
and dynamic monitoring (such as deimplementation of routine GRV
measurement and point-of-care ultrasound).
| Table III.Evidence from nurse-implemented or
nurse-relevant microbiota-supportive intervention studies in
critical care. |
Table III.
Evidence from nurse-implemented or
nurse-relevant microbiota-supportive intervention studies in
critical care.
| Author(s),
year | Study design | Population (n) | Intervention
category & details | Key outcomes
related to microbiota & FI | (Refs.) |
|---|
| Bruen et al,
2020 | Retrospective
cohort | Adult critically
ill patients in a community hospital (n=200) | Monitoring &
assessment: Nurse-driven protocol eliminating routine GRV
monitoring. | Increased EN volume
delivered; no increase in vomiting, aspiration or mortality. | (34) |
| Shimizu et
al, 2018 | Double-blind
RCT | Septic adults
(n=165) | Targeted
nourishment/restoration: Synbiotics (Lactobacillus casei +
galacto-oligosaccharides) vs. placebo. | 27% reduction in
enteritis and VAP incidence; restoration of fecal butyrate to
control levels. Demonstrated efficacy and safety in a select ICU
population. | (71) |
| Mahmoodpoor et
al, 2019 | Double-blind
RCT | Critically ill,
ventilated adults (n=150) | Direct microbial
restoration: Probiotic (Lactobacillus plantarum ATCC 202195)
vs. placebo. | Decreased VAP rates
(11 vs. 27%, P=0.02); no probiotic bacteremia reported. | (75) |
| Landgrave,
2024 | QI study | Not specified (QI
project focus) | Monitoring &
assessment: Deimplementation of GRV monitoring in favor of
comprehensive gastrointestinal assessment. | Improved nutrition
delivery; highlighted nurse's role in changing practice based on
evidence. | (77) |
| Freedberg et
al, 2020 | Randomized pilot
trial | ICU patients on
broad-spectrum antibiotics (n=22) | Targeted
nourishment: Fiber-enriched EN formula (15 g mixed fermentable
substrate/day) vs. standard fiber-free formula. | Attenuated
antibiotic-associated diarrhea; doubled the abundance of
butyrate-producing bacteria in patients with preserved baseline
Firmicutes. | (81) |
| Wei et al,
2016 | Case series | ICU patients with
MODS & diarrhea post-sepsis (n=9) | Direct microbial
restoration: Single-dose FMT via nasoduodenal tube. | Resolution of
severe antibiotic-associated diarrhea; restoration of butyrate
synthesis pathways in metagenomic analysis. | (84) |
| Seifi et al,
2022 | RCT | Critically ill
adult patients (n=60) | Targeted
nourishment/restoration: Synbiotic supplementation vs.
placebo. | Significantly
improved enteral feeding tolerance and attenuated muscle wasting.
Nurses oversaw administration and monitoring, demonstrating
clinical feasibility. | (82) |
Overarching rationale for a
nurse-driven model
Before detailing the intervention components, the
operational scope of the term ‘nurse-driven’ must be clarified. A
nurse-driven model does not imply independent nursing performance
of interventions that legally or institutionally require physician
orders [such as fecal microbiota transplantation (FMT)
prescription, antibiotic de-escalation, probiotic selection,
diagnostic gastric ultrasound]. Rather, nurses function as protocol
initiators, continuous monitors and care coordinators operating
within validated institutional protocols and multidisciplinary team
structures. Final clinical decisions, including prescribing,
ordering of diagnostic tests and invasive procedures, remain with
physicians or pharmacists where required by law, hospital policy or
scope of practice. This distinction aligns with published
nurse-driven protocol models in critical care, including
nurse-driven sedation monitoring and early mobility protocols
(73,74).
Critical care nurses are uniquely positioned to
operationalize microbiota-targeted therapies. Compared with
physicians, pharmacists and dietitians, nurses offer the distinct
advantage of continuous bedside presence for real-time dynamic
monitoring, direct administration of EN and probiotics with
immediate adverse event detection and integration of gut health
assessment with overall patient status (hemodynamics, sedation,
infection). Continuous bedside presence, responsibility for EN
delivery and participation in infection-control bundles confer a
pivotal role in modulating antibiotic exposure, feeding substrates
and probiotic/synbiotic logistics. Recent meta-analyses demonstrate
that bundles coordinated by nurses reduce ventilator-associated
pneumonia (VAP) and antibiotic consumption, both of which
indirectly preserve butyrate-producing taxa (75,76).
Furthermore, nurse-driven withdrawal of routine gastric residual
aspiration increases EN volume delivered without increasing
infectious complications, thereby augmenting fermentable substrate
influx to the colon (34,77). Collectively, these observations
justify framing microbiota support as an integral component of
nursing quality indicators.
Minimizing iatrogenic harm: Antibiotic
stewardship and PPI restriction
Pharmacovigilance studies reveal that every
additional day of broad-spectrum β-lactam therapy correlates with a
2.3% daily loss in gut microbial Shannon diversity and a
quantifiable decline in fecal butyrate (78). Nurse-initiated daily ‘antibiotic
time-outs’ (i.e., a brief, structured daily review of antibiotic
necessity, indication, spectrum and duration, typically performed
at the bedside as part of routine care) embedded in the ICU bundle
do not grant nurses prescribing authority. Instead, nurses trigger
a multidisciplinary review by flagging patients who meet predefined
duration criteria (e.g., ≥72 h of broad-spectrum therapy). The
final decision to de-escalate, change or discontinue antibiotics
rests with the attending physician or infectious disease pharmacist
as per institutional antimicrobial stewardship policies. This
nurse-triggered mechanism has been shown to shorten median
antibiotic duration by 1.8 days and to lower the incidence of
subsequent multidrug-resistant gram-negative bacteremia (75). Likewise, restricting proton-pump
inhibitors to patients with overt upper gastrointestinal bleeding
reduces gastric pH-driven colonization of oral Streptococci and
Candida spp., taxa that out-compete fiber-fermenting
anaerobes (79,80). Although comparative trials
specifically measuring butyrate rebound after PPI restriction are
lacking, observational data report a 38% increase in fecal butyrate
within four weeks of drug cessation, supporting the biological
plausibility of this intervention (80).
Targeted microbiota nourishment:
Prebiotic fibers and synbiotics
Randomized work by Freedberg et al (81) demonstrated that a fiber-enriched
enteral formula (15 g mixed fermentable substrate/day) attenuated
antibiotic-associated diarrhea and doubled the abundance of
butyrate producers in ICU patients receiving broad-spectrum agents.
Of note, the benefit was restricted to individuals whose baseline
microbiota retained ≥5% relative abundance of Firmicutes. This
threshold was derived from the exploratory analysis of the pilot
trial by Freedberg et al (81). Notably, the benefit was restricted
to individuals whose baseline microbiota retained ≥5% relative
abundance of Firmicutes; complete ecological collapse precluded
fiber conversion, underscoring the need for early intervention.
When fiber alone is insufficient, synbiotics offer a pragmatic
escalation. In a double-blind trial, Shimizu et al (71) administered Lactobacillus
casei plus galacto-oligosaccharides to septic adults, achieving
a 27% reduction in enteritis and VAP incidence alongside
restoration of fecal butyrate to control levels. Consistent with
these observations, a randomized trial in enterally fed critically
ill patients showed that synbiotic supplementation significantly
improved feeding tolerance and attenuated muscle wasting,
reinforcing the clinical relevance of preserving or restoring a
functional microbiota (82).
Adverse-event monitoring revealed no probiotic bloodstream
isolates, but the authors excluded immunosuppressed and neutropenic
subjects, emphasizing the importance of careful patient selection,
a role that bedside nurses can operationalize through daily safety
checklists.
Direct microbial restoration:
Probiotics and FMT logistics
Probiotic selection and prescription are governed by
physician orders or multidisciplinary protocols; nurses are not
authorized to independently choose or prescribe probiotics. The
nurse-driven responsibilities include verifying the correct strain,
ensuring cold-chain integrity, administering the preparation via
enteral tube and monitoring for adverse events (e.g.,
probiotic-associated bacteraemia) using daily safety checklists
(82).
Probiotic monotherapy has yielded mixed results.
Mahmoodpoor et al (75)
reported that Lactobacillus plantarum (American Type Culture
Collection 202195) decreased VAP rates (11 vs. 27%, P=0.02) without
bacteremia, whereas Cohen et al (83) documented Lactobacillus
bloodstream infections genetically identical to the administered
strain in three hematology-oncology patients, prompting early trial
cessation. These discrepancies highlight the critical importance of
host immune status, strain selection and administration
route-variables that nurses can monitor in real time. For patients
with complete butyrogenic collapse, FMT represents an emerging
rescue strategy. Wei et al (84) described resolution of severe
antibiotic-associated diarrhea and restoration of butyrate
synthesis pathways in 9 ICU patients following single-dose FMT via
nasoduodenal tube. Nursing responsibilities included donor-stool
thawing under anaerobic conditions, 6-hourly stool-bank temperature
audits and post-FMT surveillance for fever or increased vasopressor
requirements. Although promising, FMT in critical care remains
experimental; stewardship committees should embed nursing protocols
for strain tracking and adverse-event reporting before wider
adoption.
Monitoring and assessment: Beyond
GRV
Traditional reliance on GRV has poor correlation
with true gastric emptying and inadvertently reduces EN delivery.
Two nurse-implemented protocols that replaced 4-hourly GRV
aspiration with abdominal distension assessment and bowel-sound
auscultation achieved a 22% increase in energy delivery without
raising aspiration events (34,77).
Integrating point-of-care ultrasound of the gastric antrum,
validated by Valla et al (85) in ventilated children, adds
objective quantification of gastric emptying and can guide
prokinetic timing, indirectly enhancing substrate availability for
microbial fermentation. Future quality metrics should therefore
incorporate the proportion of daily energy target achieved, days
without antibiotics and fecal or breath butyrate trends where
feasible, thereby converting microbiota health into measurable
nursing outcomes.
Implementation framework: Education,
inter-professional coordination and safety
Successful translation demands structured education.
A national survey revealed that only 34% of critical-care nurses
could correctly define ‘synbiotic’ and <20% were aware of
contraindications such as central venous catheter-related fungemia
(86). Interactive workshops
coupling microbiome science with practical skills (strain
reconstitution, aseptic tube-feeding connection, adverse-event
documentation) improved knowledge scores by 45% and increased
fiber-enriched formula prescribing 3-fold (87). Inter-professional daily ‘gut
rounds’ involving nurses, intensivists, pharmacists and dietitians
further facilitate antibiotic de-escalation, fiber optimization and
rapid response to probiotic-related bloodstream infection signals.
Finally, unit governance must mandate electronic capture of
probiotic batch numbers and linkage to infection-control databases
to enable real-time traceability, an essential safeguard now
recommended by European Society of Clinical Microbiology and
Infectious Diseases and European Society for Clinical Nutrition and
Metabolism guidelines (88,89).
Implementation framework and nursing
workflow integration
The integration of microbiota-supportive
interventions into critical care nursing requires a structured,
evidence-based framework that aligns mechanistic insights from
multi-omics studies with bedside feasibility. Although the
association between gut dysbiosis, butyrate depletion and FI is
increasingly supported by metabolomic and metagenomic data, the
translation of these findings into routine nursing workflows
remains inconsistent across institutions. To bridge this gap, a
standardized implementation model must address four core domains:
Protocol development, interprofessional coordination,
patient-family engagement and continuous outcome monitoring. This
section critically appraises recent literature to delineate a
pragmatic, nurse-driven pathway that operationalizes
microbiota-targeted care in the ICU.
Protocol development: Designing a ‘gut
bundle’ for ICU nurses
A nurse-driven ‘gut bundle’ should consolidate
evidence-based elements such as antibiotic stewardship,
fiber-enriched EN and probiotic/synbiotic administration into a
single, actionable protocol. For instance, the randomized trial by
Seifi et al (82)
demonstrated that synbiotic supplementation significantly improved
enteral feeding tolerance and attenuated muscle wasting in
critically ill adults, with nurses overseeing administration and
monitoring for adverse events. Similarly, the meta-analysis by Koch
et al (90) confirmed that
fiber-supplemented EN reduced diarrhea and improved caloric
delivery, particularly when initiated early. However, heterogeneity
in fiber type (soluble vs. mixed) and dose (10–30 g/day) across
trials limits generalizability. Notably, a recent meta-analysis by
Huang et al (91) of
pectin-supplemented formulas reported modest improvements in
gastrointestinal tolerance but emphasized the need for patient
stratification based on baseline microbiota profiles. Thus, bundle
design must allow for personalized adjustments, with nurses trained
to assess pre-illness dietary habits and antibiotic exposure
history.
Interprofessional coordination: The
nurse as microbiome ambassador
Effective implementation hinges on seamless
collaboration between nurses, intensivists, pharmacists and
dietitians. While nurses are pivotal in administering
interventions, pharmacists play a critical role in validating
probiotic strain selection and monitoring drug-microbe
interactions. This inconsistency underscores the need for nurses to
lead daily ‘gut rounds’ to reconcile discrepancies between
prescribed antibiotics and microbiota-supportive therapies. For
example, concurrent use of broad-spectrum β-lactams may negate
benefits of Lactobacillus probiotics, as demonstrated in an
adult ICU study where meropenem selectively depleted Firmicutes
carrying butyrate-synthesis genes (47). Conversely, Venegas-Borsellino and
Kwon (92) reported that soluble
fiber administration during antibiotic therapy preserved
butyrate-producing taxa, but only when paired with judicious
antibiotic de-escalation. The relevance of antibiotic-microbiota
interactions is further supported by metagenomic analyses in adult
ICU patients, demonstrating that specific antibiotic classes
differentially deplete butyrogenic taxa (47). These data advocate for
nurse-initiated ‘antibiotic time-outs’ to align antimicrobial
stewardship with microbiota preservation.
Patient-family education: Demystifying
the microbiome
Despite clinical efficacy, patient and family
acceptance of microbiota-targeted therapies remains low due to
misconceptions about probiotics and fiber safety. A cross-sectional
survey by O'Connor et al (93) found that 68% of UK parents viewed
blended tube feeds (containing prebiotic fibers) as ‘risky’, citing
fears of contamination and diarrhea. To counteract this, nurses
must employ teach-back techniques to explain the mechanistic
rationale: Fiber fermentation by gut commensals yields butyrate,
which strengthens intestinal tight junctions and reduces FI risk.
Notably, the ongoing LOME-PECT trial (NCT05923456), is evaluating
whether low-methoxy pectin formulas improve gastrointestinal
tolerance in ventilated adults. The trial protocol reported by
Kashiwagi et al (94)
included a preliminary, non-peer-reviewed finding that nurse-led
education sessions improved enrollment rates by 22%; however, this
result awaits confirmation in the final published trial. Visual
aids depicting the ‘fiber-butyrate-barrier’ axis may further
enhance comprehension, particularly when tailored to literacy
levels.
Overcoming barriers: Logistics,
knowledge and culture
Implementation is frequently hindered by logistical
constraints (e.g., cold-chain storage for probiotics), knowledge
deficits and institutional resistance to change. A 2024 survey of
Indian neonatologists revealed that 55% of ICUs lacked
refrigeration protocols for probiotic stocks, leading to 30%
viability loss at point-of-care (95). To mitigate this, nurses can
advocate for unit-based probiotic dispensing systems, akin to those
used for biologicals. Knowledge gaps are equally critical: Only 34%
of critical care nurses correctly identified synbiotic
contraindications (e.g., immunosuppression) in a national
assessment (86). Simulation-based
training, as piloted by Casavant et al (96), improved nurse confidence in
microbiome-informed care from 45 to 78% post-intervention. Finally,
cultural resistance persists where FI is viewed as inevitable.
Based on evidence from general ICU populations, Patel et al
(97) reframed FI as a
‘preventable iatrogenic injury’. This principle has been
successfully implemented in nurse-led studies conducted in general
ICU settings, without device-specific restrictions (34,77).
Monitoring and feedback: From gut
health to quality indicators
Traditional FI metrics (e.g., GRV) correlate poorly
with microbiota health. Instead, composite indicators such as ‘days
without antibiotics’, ‘percentage of energy target achieved’ and
fecal butyrate levels (via point-of-care testing) offer actionable
feedback. In a single-center quality improvement project,
integrating these metrics into nursing dashboards reduced FI
incidence by 18% over 6 months (98). However, a critical limitation of
the proposed 30-min butyrate assay is the absence of a clinically
validated decision threshold. Although one study reported that a
fecal butyrate concentration below 2 µmol/g within the first 72 h
of ICU admission predicted 30-day mortality (AUROC=0.87) (99), this cut-off has not been
prospectively validated to guide specific interventions for feeding
intolerance, such as probiotic selection, fiber escalation or
prokinetic therapy. Without such intervention-specific thresholds,
the clinical utility of real-time butyrate measurement remains
uncertain. However, the feasibility of fecal testing remains
contentious: While Green et al (100) validated a 30-min butyrate assay,
cost ($12/test), workflow disruption and the lack of actionable
cut-offs limit adoption. Therefore, widespread implementation of
point-of-care butyrate testing in routine ICU practice is currently
premature. Proxy markers (e.g., low fecal pH correlating with
butyrate abundance) may offer pragmatic alternatives pending
technological advances.
Future directions
Despite mounting associative data, the question of
whether butyrate depletion is a true driver of FI or merely a
bystander of critical illness remains unresolved. Longitudinal
multi-omics studies that repeatedly sample both the gut microbiome
and circulating metabolites from ICU admission to convalescence are
urgently needed. Haak et al (70) integrated longitudinal metagenomic
and metabolomic profiling in a cohort of 29 ventilated adults and
demonstrated that the loss of butyryl-CoA:acetate CoA-transferase
genes preceded the first episode of FI by a median of 48 h
(interquartile range not reported in the original study). However,
the lack of strain-level resolution limited the interpretation of
these findings, as the potential contribution of concomitant
pathogens, rather than the loss of butyrate-producing organisms
specifically, could not be excluded as precipitating dysmotility.
Conversely, Ivanova et al (101) employed high-throughput chromosome
conformation capture metagenomics to demonstrate the physical
linkage between butyrate synthesis genes and specific
Firmicutes-associated contigs in a cohort of 21 chronically
critically ill patients, thereby strengthening evidence for a
taxon-specific mechanistic relationship. Nevertheless, the lack of
daily nutritional intake data within this cohort limited the
ability to determine a causal association with FI onset. Future
investigations integrating strain-resolved metagenomics with
standardized EN protocols and high-frequency assessments of gastric
emptying are warranted to more definitively establish
causality.
Beyond causal inference, strain- and gene-level
specificity is required before microbiota-directed therapeutics can
be individualized. Most trials have relied on generic probiotics
whose genomes often lack the complete butyrate synthesis pathway. A
longitudinal analysis by Kitsios et al (102) revealed that only 6 of 21
commercially available probiotic strains carried
butyryl-CoA:acetate CoA-transferase and supplementation with these
strains yielded a 2.1-fold increase in fecal butyrate (no P-value
or confidence interval provided in the original report), whereas
strains without the pathway had no metabolomic benefit. Schlechte
et al (103) further
showed that carriage of the butyrate kinase route, dominant in
Roseburia spp., was associated with faster gastric emptying
(β=−0.34; P=0.02), whereas the butyryl-CoA:acetate CoA-transferase
route was not, implying that functional gene complement, rather
than taxonomic label, determines physiological efficacy. Future
trials should therefore pre-screen candidate strains for complete
butyrogenic cassettes and use metatranscriptomics to confirm
in-situ expression during critical illness.
Equally unsettled are the optimal timing, dose and
matrix of butyrate-enhancing interventions. Cho et al
(99) recently demonstrated that
fecal butyrate concentrations <2 µmol/g within the first 72 h of
ICU admission predicted 30-day mortality with an AUROC of 0.87,
suggesting an early ‘metabolomic window’ during which
microbiota-directed therapy might be most impactful. Yet
sequential-feeding trial by Yao et al (104) indicated that interrupting EN for
≥4 h abolished the butyrogenic effect of fiber, underscoring the
importance of continuous substrate delivery. Conversely, a
longitudinal multi-compartment study by Kitsios et al
(102) revealed that
pharmacologic sodium butyrate (4 g/day) shortened the gastric
emptying time only when administered after day 5 of illness,
implying that host responsiveness may hinge on the immune
trajectory. Dose-finding studies that integrate both prokinetic
endpoints and metabolomic read-outs across variable illness phases
are therefore warranted.
Finally, the bedside implementation of multi-omics
remains embryonic. Point-of-care sensors that measure breath or
fecal volatile organic compounds have been explored as non-invasive
surrogates for fecal butyrate, offering a potential tool amenable
to nurse-led monitoring (105).
Embedding such read-outs into electronic health records could
trigger closed-loop decision support: Automatic escalation of
fiber-enriched formulae, probiotic strain selection or antibiotic
de-escalation. Crucially, any future model must be evaluated in
nurse-driven, FI-primary randomized trials powered to link
metabolite restoration with hard clinical endpoints, such as
ventilator days and infection-free survival. Only through such
pragmatic, mechanism-grounded investigations will the promise of
multi-omics translate into tangible benefits for critically ill
patients.
Conclusions
Collectively, multi-omics evidence supports the
proposed pathway from critical illness-induced gut dysbiosis to
butyrate depletion and subsequent FI. This mechanistic
understanding reframes FI as a disorder of microbial ecology.
Consequently, the imperative is to translate this knowledge into
pragmatic bedside care. Nurses are uniquely positioned to lead this
paradigm shift by implementing microbiota-supportive interventions
that encompass stewardship, nourishment and restoration to preserve
gut health, prevent FI and improve outcomes in the vulnerable
critically ill.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
LW conceived and designed the review, acquired and
analyzed literature, and drafted the manuscript. XK contributed to
pathophysiological content and critically revised the manuscript.
YL focused on neonatal aspects and assisted in evidence synthesis.
HW developed the nursing framework and edited the manuscript. YG
and FS supervised the work, provided critical revisions and
approved the final version. Data authentication is not applicable.
All authors read, approved and are accountable for the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
FI
|
feeding intolerance
|
|
GRV
|
gastric residual volume
|
|
EN
|
enteral nutrition
|
|
ICU
|
intensive care unit
|
|
SCFA
|
short-chain fatty acid
|
|
PPIs
|
proton pump inhibitors
|
|
HDAC
|
histone deacetylase
|
|
Treg
|
regulatory T cell
|
|
FMT
|
fecal microbiota transplantation
|
|
AUROC
|
area under the receiver operating
characteristic curve
|
|
VAP
|
ventilator-associated pneumonia
|
|
MCT-1
|
monocarboxylate transporter 1
|
|
RCT
|
randomized controlled trial
|
|
IL
|
interleukin
|
|
QI
|
quality improvement
|
References
|
1
|
Jenkins B, Calder PC and Marino LV: A
systematic review of the definitions and prevalence of feeding
intolerance in critically ill adults. Clin Nutr ESPEN. 49:92–102.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Eveleens RD, Joosten KFM, de Koning BAE,
Hulst JM and Verbruggen SCAT: Definitions, predictors and outcomes
of feeding intolerance in critically ill children: A systematic
review. Clin Nutr. 39:685–693. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Guo Y, Zhang Y, Gerhard M, Gao JJ,
Mejias-Luque R, Zhang L, Vieth M, Ma JL, Bajbouj M, Suchanek S, et
al: Effect of Helicobacter pylori on gastrointestinal
microbiota: A population-based study in Linqu, a high-risk area of
gastric cancer. Gut. 69:1598–1607. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Heyland DK, Ortiz A, Stoppe C, Patel JJ,
Yeh DD, Dukes G, Chen YJ, Almansa C and Day AG: Incidence, risk
factors, and clinical consequence of enteral feeding intolerance in
the mechanically ventilated Critically Ill: An analysis of a
multicenter, multiyear database. Crit Care Med. 49:49–59. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lavrentieva A, Kontakiotis T and Bitzani
M: Enteral nutrition intolerance in critically ill septic burn
patients. J Burn Care Res. 35:313–318. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Padar M, Starkopf J, Uusvel G and Reintam
Blaser A: Gastrointestinal failure affects outcome of intensive
care. J Crit Care. 52:103–108. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sun Y, Li S, Wang S, Li C, Li G, Xu J,
Wang H, Liu F, Yao G, Chang Z, et al: Predictors of 1-year
mortality in patients on prolonged mechanical ventilation after
surgery in intensive care unit: A multicenter, retrospective cohort
study. BMC Anesthesiol. 20:442020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Vijayaraghavan R, Maiwall R, Arora V,
Choudhary A, Benjamin J, Aggarwal P, Jamwal KD, Kumar G, Joshi YK
and Sarin SK: Reversal of feed intolerance by prokinetics improves
survival in critically Ill cirrhosis patients. Dig Dis Sci.
67:4223–4233. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gungabissoon U, Hacquoil K, Bains C,
Irizarry M, Dukes G, Williamson R, Deane AM and Heyland DK:
Prevalence, risk factors, clinical consequences, and treatment of
enteral feed intolerance during critical illness. JPEN J Parenter
Enteral Nutr. 39:441–448. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Martinez EE, Douglas K, Nurko S and Mehta
NM: Gastric dysmotility in critically Ill children:
Pathophysiology, diagnosis, and management. Pediatr Crit Care Med.
16:828–836. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Robles-Vera I, Toral M and Duarte J:
Microbiota and hypertension: Role of the sympathetic nervous system
and the immune system. Am J Hypertens. 33:890–901. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Latorre M, Krishnareddy S and Freedberg
DE: Microbiome as mediator: Do systemic infections start in the
gut? World J Gastroenterol. 21:10487–10492. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shimizu K, Ogura H, Asahara T, Nomoto K,
Matsushima A, Hayakawa K, Ikegawa H, Tasaki O, Kuwagata Y and
Shimazu T: Gut microbiota and environment in patients with major
burns-a preliminary report. Burns. 41:e28–e33. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yamashiro Y: Gut microbiota in health and
disease. Ann Nutr Metab. 71:242–246. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ravi A, Halstead FD, Bamford A, Casey A,
Thomson NM, van Schaik W, Snelson C, Goulden R, Foster-Nyarko E,
Savva GM, et al: Loss of microbial diversity and pathogen
domination of the gut microbiota in critically ill patients. Microb
Genom. 5:e0002932019.PubMed/NCBI
|
|
16
|
Buelow E, Bello González TDJ, Fuentes S,
de Steenhuijsen Piters WAA, Lahti L, Bayjanov JR, Majoor EAM, Braat
JC, van Mourik MSM, Oostdijk EAN, et al: Comparative gut microbiota
and resistome profiling of intensive care patients receiving
selective digestive tract decontamination and healthy subjects.
Microbiome. 5:882017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Freedberg DE, Zhou MJ, Cohen ME,
Annavajhala MK, Khan S, Moscoso DI, Brooks C, Whittier S, Chong DH,
Uhlemann AC and Abrams JA: Pathogen colonization of the
gastrointestinal microbiome at intensive care unit admission and
risk for subsequent death or infection. Intensive Care Med.
44:1203–1211. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Marfil-Sánchez A, Zhang L, Alonso-Pernas
P, Mirhakkak M, Mueller M, Seelbinder B, Ni Y, Santhanam R, Busch
A, Beemelmanns C, et al: An integrative understanding of the large
metabolic shifts induced by antibiotics in critical illness. Gut
Microbes. 13:19935982021. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ney LM, Wipplinger M, Grossmann M, Engert
N, Wegner VD and Mosig AS: Short chain fatty acids: Key regulators
of the local and systemic immune response in inflammatory diseases
and infections. Open Biol. 13:2300142023. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang XY, Chen J, Yi K, Peng L, Xie J, Gou
X, Peng T and Tang L: Phlorizin ameliorates obesity-associated
endotoxemia and insulin resistance in high-fat diet-fed mice by
targeting the gut microbiota and intestinal barrier integrity. Gut
Microbes. 12:1–18. 2020. View Article : Google Scholar
|
|
21
|
Du L and Jiang W, Zhu X, Zhu L, Fan Y and
Jiang W: Rifaximin alleviates intestinal barrier disruption and
systemic inflammation via the PXR/NFκB/MLCK pathway and modulates
intestinal Lachnospiraceae abundance in heat-stroke mice. Int
Immunopharmacol. 143:1134622024. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Al-Harbi NO, Nadeem A, Ahmad SF, Alotaibi
MR, AlAsmari AF, Alanazi WA, Al-Harbi MM, El-Sherbeeny AM and
Ibrahim KE: Short chain fatty acid, acetate ameliorates
sepsis-induced acute kidney injury by inhibition of NADPH oxidase
signaling in T cells. Int Immunopharmacol. 58:24–31. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang T, Xie S, Cao L, Li M, Ding L, Wang
L, Pang S, Wang Z and Geng L: Astragaloside IV Modulates gut
macrophages M1/M2 polarization by reshaping Gut microbiota and
short chain fatty acids in sepsis. Shock. 61:120–131. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yagmurdur H and Leblebici F: Enteral
nutrition preference in critical care: Fibre-enriched or
fibre-free? Asia Pac J Clin Nutr. 25:740–746. 2016.PubMed/NCBI
|
|
25
|
Chen Y, A S, Liu C, Zhang T, Yang J and
Tian X: A randomized controlled trial assessing the impact of
transcutaneous electrical acupoint stimulation on gastrointestinal
motility, nutritional status, and immune function in patients
following cerebrovascular accident surgery. J Invest Surg.
37:24340932024. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wijeyesekera A, Wagner J, De Goffau M,
Thurston S, Rodrigues Sabino A, Zaher S, White D, Ridout J, Peters
MJ, Ramnarayan P, et al: Multi-compartment profiling of bacterial
and host metabolites identifies intestinal dysbiosis and its
functional consequences in the Critically Ill child. Crit Care Med.
47:e727–e734. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wu T, Xu F, Su C, Li H, Lv N, Liu Y, Gao
Y, Lan Y and Li J: Alterations in the gut microbiome and cecal
metabolome during klebsiella pneumoniae-induced pneumosepsis. Front
Immunol. 11:13312020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fang H, Fang M, Wang Y, Zhang H, Li J,
Chen J, Wu Q, He L, Xu J, Deng J, et al: Indole-3-Propionic acid as
a potential therapeutic agent for Sepsis-induced gut microbiota
disturbance. Microbiol Spectr. 10:e00125222022. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pauline M, Fouhse J, Hinchliffe T, Wizzard
P, Nation P, Huynh H, Wales P, Willing B and Turner J: Probiotic
treatment vs empiric oral antibiotics for managing dysbiosis in
short bowel syndrome: Impact on the mucosal and stool microbiota,
short-chain fatty acids, and adaptation. JPEN J Parenter Enteral
Nutr. 46:1828–1838. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Stewart CJ, Embleton ND, Marrs ECL, Smith
DP, Fofanova T, Nelson A, Skeath T, Perry JD, Petrosino JF,
Berrington JE and Cummings SP: Longitudinal development of the gut
microbiome and metabolome in preterm neonates with late onset
sepsis and healthy controls. Microbiome. 5:752017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vaitkute G, Panic G, Alber DG,
Faizura-Yeop I, Cloutman-Green E, Swann J, Veys P, Standing JF,
Klein N and Bajaj-Elliott M: Linking gastrointestinal microbiota
and metabolome dynamics to clinical outcomes in paediatric
haematopoietic stem cell transplantation. Microbiome. 10:892022.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou P, Zou Z, Wu W, Zhang H, Wang S, Tu
X, Huang W, Chen C, Zhu S, Weng Q and Zheng S: The gut-lung axis in
critical illness: Microbiome composition as a predictor of
mortality at day 28 in mechanically ventilated patients. BMC
Microbiol. 23:3992023. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ozen N, Blot S, Ozen V, Arikan Donmez A,
Gurun P, Cinar FI and Labeau S: Gastric residual volume measurement
in the intensive care unit: An international survey reporting
nursing practice. Nurs Crit Care. 23:263–269. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bruen T, Rawal S, Tomesko J and Byham-Gray
L: Elimination of routine gastric residual volume monitoring
improves patient outcomes in adult Critically Ill patients in a
community hospital setting. Nutr Clin Pract. 35:522–532. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yasuda H, Kondo N, Yamamoto R, Asami S,
Abe T, Tsujimoto H, Tsujimoto Y and Kataoka Y: Monitoring of
gastric residual volume during enteral nutrition. Cochrane Database
Syst Rev. 9:CD0133352021.PubMed/NCBI
|
|
36
|
Alexander JL and Mullish BH: A guide to
the gut microbiome and its relevance to critical care. Br J Nurs.
29:1106–1112. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hamilton LA and Behal ML: Altering routine
intensive care unit practices to support commensalism. Nutr Clin
Pract. 35:433–441. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Smith M, Smith M and Robinson KN: Using
Nurse-driven protocols to eliminate routine gastric residual volume
measurements: A retrospective study. Crit Care Nurse. 42:e1–e10.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Feng L, Chen J and Xu Q: Is monitoring of
gastric residual volume for critically ill patients with enteral
nutrition necessary? A meta-analysis and systematic review. Int J
Nurs Pract. 29:e131242023. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lindner M, Padar M, Mändul M, Christopher
KB, Reintam Blaser A, Gratz HC, Elke G and Bachmann KF: Current
practice of gastric residual volume measurements and related
outcomes of critically ill patients: A secondary analysis of the
intestinal-specific organ function assessment study. JPEN J
Parenter Enteral Nutr. 47:614–623. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lamarche D, Johnstone J, Zytaruk N, Clarke
F, Hand L, Loukov D, Szamosi JC, Rossi L, Schenck LP, Verschoor CP,
et al: Microbial dysbiosis and mortality during mechanical
ventilation: A prospective observational study. Respir Res.
19:2452018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yang W, Yu T, Huang X, Bilotta AJ, Xu L,
Lu Y, Sun J, Pan F, Zhou J, Zhang W, et al: Intestinal
microbiota-derived short-chain fatty acids regulation of immune
cell IL-22 production and gut immunity. Nat Commun. 11:44572020.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yildiz SS, Yalinay M and Karakan T:
Bismuth-based quadruple Helicobacter pylori eradication regimen
alters the composition of gut microbiota. Infez Med. 26:115–121.
2018.PubMed/NCBI
|
|
44
|
Annavajhala MK, Gomez-Simmonds A, Macesic
N, Sullivan SB, Kress A, Khan SD, Giddins MJ, Stump S, Kim GI,
Narain R, et al: Colonizing multidrug-resistant bacteria and the
longitudinal evolution of the intestinal microbiome after liver
transplantation. Nat Commun. 10:47152019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Feng Y, Huang Y, Wang Y, Wang P, Song H
and Wang F: Antibiotics induced intestinal tight junction barrier
dysfunction is associated with microbiota dysbiosis, activated
NLRP3 inflammasome and autophagy. PLoS One. 14:e02183842019.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chernevskaya E, Beloborodova N, Klimenko
N, Pautova A, Shilkin D, Gusarov V and Tyakht A: Serum and fecal
profiles of aromatic microbial metabolites reflect gut microbiota
disruption in critically ill patients: A prospective observational
pilot study. Crit Care. 24:3122020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Maier L, Goemans CV, Wirbel J, Kuhn M,
Eberl C, Pruteanu M, Müller P, Garcia-Santamarina S, Cacace E,
Zhang B, et al: Unravelling the collateral damage of antibiotics on
gut bacteria. Nature. 599:120–124. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Hojo M, Asahara T, Nagahara A, Takeda T,
Matsumoto K, Ueyama H, Matsumoto K, Asaoka D, Takahashi T, Nomoto
K, et al: Gut microbiota composition before and after use of proton
pump inhibitors. Dig Dis Sci. 63:2940–2949. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Horvath A, Leber B, Feldbacher N,
Steinwender M, Komarova I, Rainer F, Blesl A and Stadlbauer V: The
effects of a multispecies synbiotic on microbiome-related side
effects of long-term proton pump inhibitor use: A pilot study. Sci
Rep. 10:27232020. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Leclercq S, Le Roy T, Furgiuele S, Coste
V, Bindels LB, Leyrolle Q, Neyrinck AM, Quoilin C, Amadieu C, Petit
G, et al: Gut Microbiota-induced changes in β-Hydroxybutyrate
metabolism are linked to altered sociability and depression in
alcohol use disorder. Cell Rep. 33:1082382020. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Lama A, Annunziata C, Coretti L, Pirozzi
C, Di Guida F, Nitrato Izzo A, Cristiano C, Mollica MP, Chiariotti
L, Pelagalli A, et al: N-(1-carbamoyl-2-phenylethyl) butyramide
reduces antibiotic-induced intestinal injury, innate immune
activation and modulates microbiota composition. Sci Rep.
9:48322019. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Rashidi A, Ebadi M, Rehman TU, Elhusseini
H, Nalluri H, Kaiser T, Holtan SG, Khoruts A, Weisdorf DJ, Staley
C, et al: Gut microbiota response to antibiotics is personalized
and depends on baseline microbiota. Microbiome. 9:2112021.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yang J, Wei H, Zhou Y, Szeto CH, Li C, Lin
Y, Coker OO, Lau HCH, Chan AWH, Sung JJY and Yu J: High-Fat diet
promotes colorectal tumorigenesis through modulating gut microbiota
and metabolites. Gastroenterology. 162:135–149.e2. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yuille S, Reichardt N, Panda S, Dunbar H
and Mulder IE: Human gut bacteria as potent class I histone
deacetylase inhibitors in vitro through production of butyric acid
and valeric acid. PLoS One. 13:e02010732018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Su X, Yin X, Liu Y, Yan X, Zhang S, Wang
X, Lin Z, Zhou X, Gao J, Wang Z and Zhang Q: Gut dysbiosis
contributes to the imbalance of treg and Th17 cells in Graves'
disease patients by propionic acid. J Clin Endocrinol Metab.
105:dgaa5112020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Liu C, Liu J, Wang W, Yang M, Chi K, Xu Y
and Guo N: Epigallocatechin gallate alleviates staphylococcal
enterotoxin A-Induced intestinal barrier damage by regulating gut
microbiota and inhibiting the TLR4-NF-κB/MAPKs-NLRP3 inflammatory
cascade. J Agric Food Chem. 71:16286–16302. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Karunaratne TB, Okereke C, Seamon M,
Purohit S, Wakade C and Sharma A: Niacin and Butyrate:
Nutraceuticals targeting dysbiosis and intestinal permeability in
Parkinson's disease. Nutrients. 13:282020. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
He KY, Lei XY, Wu DH, Zhang L, Li JQ, Li
QT, Yin WT, Zhao ZL, Liu H, Xiang XY, et al: Akkermansia
muciniphila protects the intestine from irradiation-induced
injury by secretion of propionic acid. Gut Microbes.
15:22933122023. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zaccaria E, Klaassen T, Alleleyn AME,
Boekhorst J, Smokvina T, Kleerebezem M and Troost FJ: Endogenous
small intestinal microbiome determinants of transient colonisation
efficiency by bacteria from fermented dairy products: A randomised
controlled trial. Microbiome. 11:432023. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Stilling RM, van de Wouw M, Clarke G,
Stanton C, Dinan TG and Cryan JF: The neuropharmacology of
butyrate: The bread and butter of the microbiota-gut-brain axis?
Neurochem Int. 99:110–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kennedy KV, Fawad JA, Neugut YD,
Valenzuela-Araujo D, Coe A, Bhatti TR, Acord M, Manfredi MD, Mamula
P, Loomes KM and Brownell JN: Histone deacetylase inhibition by gut
Microbe-generated short-chain fatty acids entrains intestinal
epithelial circadian rhythms. Gastroenterology. 163:1377–1390.e11.
2022. View Article : Google Scholar
|
|
62
|
Hoffmann TW, Pham HP, Bridonneau C, Aubry
C, Lamas B, Martin-Gallausiaux C, Moroldo M, Rainteau D, Lapaque N,
Six A, et al: Microorganisms linked to inflammatory bowel
disease-associated dysbiosis differentially impact host physiology
in gnotobiotic mice. ISME J. 10:460–477. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Ventura I, Chomon-García M, Tomás-Aguirre
F, Palau-Ferré A, Legidos-García ME, Murillo-Llorente MT and
Pérez-Bermejo M: Therapeutic and immunologic effects of Short-chain
fatty acids in inflammatory bowel disease: A systematic review. Int
J Mol Sci. 25:108792024. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Doifode T, Giridharan VV, Generoso JS,
Bhatti G, Collodel A, Schulz PE, Forlenza OV and Barichello T: The
impact of the microbiota-gut-brain axis on Alzheimer's disease
pathophysiology. Pharmacol Res. 164:1053142021. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Chen C, Liu C, Mu K and Xue W:
Lactobacillus paracasei AH2 isolated from Chinese sourdough
alleviated gluten-induced food allergy through modulating gut
microbiota and promoting short-chain fatty acid accumulation in a
BALB/c mouse model. J Sci Food Agric. 104:664–674. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Blot F, Marchix J, Ejarque M, Jimenez S,
Meunier A, Keime C, Trottier C, Croyal M, Lapp C, Mahe MM, et al:
Gut microbiota remodeling and intestinal adaptation to lipid
malabsorption after enteroendocrine cell loss in adult mice. Cell
Mol Gastroenterol Hepatol. 15:1443–1461. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Zhang F, Wan Y, Zuo T, Yeoh YK, Liu Q,
Zhang L, Zhan H, Lu W, Xu W, Lui GCY, et al: Prolonged impairment
of Short-chain fatty acid and L-Isoleucine biosynthesis in gut
microbiome in patients with COVID-19. Gastroenterology.
162:548–561.e4. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Nabizadeh E, Sadeghi J, Rezaee MA,
Hamishehkar H, Hasani A, Kafil HS, Sharifi Y, Asnaashari S,
Kadkhoda H and Ghotaslou R: The profile of key gut microbiota
members and short-chain fatty acids in patients with sepsis.
Heliyon. 9:e178802023. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Valdés-Duque BE, Giraldo-Giraldo NA,
Jaillier-Ramírez AM, Giraldo-Villa A, Acevedo-Castaño I,
Yepes-Molina MA, Barbosa-Barbosa J, Barrera-Causil CJ and
Agudelo-Ochoa GM: Stool short-chain fatty acids in critically Ill
patients with sepsis. J Am Coll Nutr. 39:706–712. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Haak BW, Argelaguet R, Kinsella CM,
Kullberg RFJ, Lankelma JM, Deijs M, Klein M, Jebbink MF, Hugenholtz
F, Kostidis S, et al: Integrative transkingdom analysis of the gut
microbiome in antibiotic perturbation and critical Illness.
mSystems. 6:e01148–20. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Shimizu K, Yamada T, Ogura H, Mohri T,
Kiguchi T, Fujimi S, Asahara T, Yamada T, Ojima M, Ikeda M and
Shimazu T: Synbiotics modulate gut microbiota and reduce enteritis
and ventilator-associated pneumonia in patients with sepsis: A
randomized controlled trial. Crit Care. 22:2392018. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Chu JR, Kang SY, Kim SE, Lee SJ, Lee YC
and Sung MK: Prebiotic UG1601 mitigates constipation-related events
in association with gut microbiota: A randomized placebo-controlled
intervention study. World J Gastroenterol. 25:6129–6144. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Neunhoeffer F, Kumpf M, Renk H, Hanelt M,
Berneck N, Bosk A, Gerbig I, Heimberg E and Hofbeck M: Nurse-driven
pediatric analgesia and sedation protocol reduces withdrawal
symptoms in critically ill medical pediatric patients. Paediatr
Anaesth. 25:786–794. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Black K, Smith S, Frotan M, Vandertulip K
and Miller A: Safety of a nurse-driven mobility protocol in a
surgical trauma intensive care unit. J Acute Care Phys Ther.
12:51–56. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Mahmoodpoor A, Hamishehkar H, Asghari R,
Abri R, Shadvar K and Sanaie S: Effect of a probiotic preparation
on ventilator-associated pneumonia in critically Ill patients
admitted to the intensive care unit: A prospective double-blind
randomized controlled trial. Nutr Clin Pract. 34:156–162. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Tsilika M, Thoma G, Aidoni Z, Tsaousi G,
Fotiadis K, Stavrou G, Malliou P, Chorti A, Massa H, Antypa E, et
al: A four-probiotic preparation for ventilator-associated
pneumonia in multi-trauma patients: Results of a randomized
clinical trial. Int J Antimicrob Agents. 59:1064712022. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Landgrave HE: Deimplementation of gastric
residual volume monitoring to enhance patient nutrition. Crit Care
Nurse. 44:34–44. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Wan CM, Yu H, Liu G, Xu HM, Mao ZQ, Xu Y,
Jin Y, Luo RP, Wang WJ and Fang F: A multicenter randomized
controlled study of Saccharomyces boulardii in the
prevention of antibiotic-associated diarrhea in infants and young
children. Zhonghua Er Ke Za Zhi. 55:349–354. 2017.(In Chinese).
PubMed/NCBI
|
|
79
|
Tuncay P, Arpaci F, Doganay M, Erdem D,
Sahna A, Ergun H and Atabey D: Use of standard enteral formula
versus enteric formula with prebiotic content in nutrition therapy:
A randomized controlled study among neuro-critical care patients.
Clin Nutr ESPEN. 25:26–36. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Vahdat Shariatpanahi Z, Jamshidi F,
Nasrollahzadeh J, Amiri Z and Teymourian H: Effect of honey on
diarrhea and fecal microbiotain in critically Ill Tube-fed
patients: A single center randomized controlled study. Anesth Pain
Med. 8:e628892018.PubMed/NCBI
|
|
81
|
Freedberg DE, Messina M, Lynch E, Tess M,
Miracle E, Chong DH, Wahab R, Abrams JA, Wang HH and Munck C:
Impact of Fiber-based enteral nutrition on the gut microbiome of
ICU patients receiving broad-spectrum antibiotics: A randomized
pilot trial. Crit Care Explor. 2:e01352020.PubMed/NCBI
|
|
82
|
Seifi N, Rezvani R, Sedaghat A, Nematy M,
Khadem-Rezaiyan M and Safarian M: The effects of synbiotic
supplementation on enteral feeding tolerance, protein homeostasis,
and muscle wasting of critically ill adult patients: A randomized
controlled trial. Trials. 23:8462022. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Cohen SA, Woodfield MC, Boyle N, Stednick
Z, Boeckh M and Pergam SA: Incidence and outcomes of bloodstream
infections among hematopoietic cell transplant recipients from
species commonly reported to be in over-the-counter probiotic
formulations. Transpl Infect Dis. 18:699–705. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Wei Y, Yang J, Wang J, Yang Y, Huang J,
Gong H, Cui H and Chen D: Successful treatment with fecal
microbiota transplantation in patients with multiple organ
dysfunction syndrome and diarrhea following severe sepsis. Crit
Care. 20:3322016. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Valla FV, Cercueil E, Morice C, Tume LN
and Bouvet L: Point-of-care gastric ultrasound confirms the
inaccuracy of gastric residual volume measurement by aspiration in
Critically Ill children: GastriPed study. Front Pediatr.
10:9039442022. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Powers J, Bourgault AM and Carroll Simmons
JS: Assessment for enteral feeding intolerance by critical care
nurses: A National Survey. Dimens Crit Care Nurs. 44:69–76. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Wheeler KE, Cook DJ, Mehta S, Calce A,
Guenette M, Perreault MM, Thiboutot Z, Duffett M and Burry L: Use
of probiotics to prevent ventilator-associated pneumonia: A survey
of pharmacists' attitudes. J Crit Care. 31:221–226. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Singer P, Blaser AR, Berger MM, Calder PC,
Casaer M, Hiesmayr M, Mayer K, Montejo-Gonzalez JC, Pichard C,
Preiser JC, et al: ESPEN practical and partially revised guideline:
Clinical nutrition in the intensive care unit. Clin Nutr.
42:1671–1689. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
van den Akker CHP, van Goudoever JB,
Shamir R, Domellöf M, Embleton ND, Hojsak I, Lapillonne A, Mihatsch
WA, Berni Canani R, Bronsky J, et al: Probiotics and preterm
infants: A position paper by the European Society for Paediatric
gastroenterology hepatology and nutrition committee on nutrition
and the European Society for Paediatric Gastroenterology Hepatology
and nutrition working group for probiotics and prebiotics. J
Pediatr Gastroenterol Nutr. 70:664–680. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Koch JL, Lew CCH, Kork F, Koch A, Stoppe
C, Heyland DK, Dresen E, Lee ZY and Hill A: The efficacy of
fiber-supplemented enteral nutrition in critically ill patients: A
systematic review and meta-analysis of randomized controlled trials
with trial sequential analysis. Crit Care. 28:3592024. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Huang HB, Zhu YB and Yu DX: Use of
pectin-supplemented enteral nutrition in intensive care: A
systematic review and meta-analysis. Clin Nutr ESPEN. 68:62–70.
2025. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Venegas-Borsellino C and Kwon M: Impact of
soluble fiber in the microbiome and outcomes in Critically Ill
patients. Curr Nutr Rep. 8:347–355. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
O'Connor G, Cleghorn S, Salam M and Watson
K: Exploring dietitians' Experience of blended tube feed in
paediatric inpatient settings: National Cross-sectional survey,
United Kingdom. J Hum Nutr Diet. 38:e700162025. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Kashiwagi S, Takaki S, Oi Y, Honzawa H,
Yamamoto R, Yamashita I, Ohki I, Fujitani S, Nagatomi A, Ohshima Y,
et al: Low-methoxy pectin-containing enteral nutrition in critical
care for intestinal tolerance (LOME-PECT): Study protocol for a
randomized controlled trial. PLoS One. 20:e03265822025. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
More K, Hanumantharaju A, Amrit A,
Nimbalkar SM and Patole S: Use of probiotics for preventing
necrotizing enterocolitis in preterm infants: A survey of current
practices among Indian neonatologists. Cureus.
16:e739232024.PubMed/NCBI
|
|
96
|
Casavant SG, Chen J, Xu W, Lainwala S,
Matson A, Chen MH, Starkweather A, Maas K and Cong XS: Multi-Omics
analysis on neurodevelopment in preterm neonates: A protocol paper.
Nurs Res. 70:462–468. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Patel JJ, Martindale RG and McClave SA:
Contemporary rationale for delivering enteral nutrition in
Critically Ill adults. Crit Care Med. 53:e1481–e1490. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Faust KB, Lupatsii M, Römer F,
Graspeuntner S, Waschina S, Zimmermann S, Humberg A, Fortmann MI,
Hanke K, Böckenholt K, et al: Use of macrogol to accelerate feeding
advancement in extremely preterm infants. Neonatology. 122:350–359.
2025. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Cho NA, Strayer K, Dobson B and McDonald
B: Pathogenesis and therapeutic opportunities of gut microbiome
dysbiosis in critical illness. Gut Microbes. 16:23514782024.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Green CH, Busch RA and Patel JJ: Fiber in
the ICU: Should it be a regular part of feeding? Curr Gastroenterol
Rep. 23:142021. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Ivanova V, Chernevskaya E, Vasiluev P,
Ivanov A, Tolstoganov I, Shafranskaya D, Ulyantsev V, Korobeynikov
A, Razin SV, Beloborodova N, et al: Exploring clinically relevant
features of gut microbiome in chronically Critically Ill patients.
Front Microbiol. 12:7703232021. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Kitsios GD, Sayed K, Fitch A, Yang H,
Britton N, Shah F, Bain W, Evankovich JW, Qin S, Wang X, et al:
Longitudinal multicompartment characterization of host-microbiota
interactions in patients with acute respiratory failure. Nat
Commun. 15:47082024. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Schlechte J, Zucoloto AZ, Yu IL, Doig CJ,
Dunbar MJ, McCoy KD and McDonald B: Dysbiosis of a
microbiota-immune metasystem in critical illness is associated with
nosocomial infections. Nat Med. 29:1017–1027. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Yao B, Liu JY, Liu Y, Song XX, Wang SB,
Liu N, Dong ZH, Yuan ZY, Han XN and Xing JY: Sequential versus
continuous feeding and its effect on the gut microbiota in
critically ill patients: A randomized controlled trial. Clin Nutr
ESPEN. 66:245–254. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Mick E, Tsitsiklis A, Kamm J, Kalantar KL,
Caldera S, Lyden A, Tan M, Detweiler AM, Neff N, Osborne CM, et al:
Integrated host/microbe metagenomics enables accurate lower
respiratory tract infection diagnosis in critically ill children. J
Clin Invest. 133:e1659042023. View Article : Google Scholar : PubMed/NCBI
|