Introduction
Breast cancer is the most common malignant tumor
among women and, of all cancers, it is the second leading cause of
mortality in women in the United States. In 2010, an estimated
207,090 women are likely to be diagnosed with invasive breast
cancer and 39,840 women are likely to succumb to this disease
(1). Obesity has been epidemic in
the United States for more than two decades and the proportion of
overweight and obese adults in the population continues to rise. In
an investigation of the role of overweight and obesity in
carcinogenesis, Calle et al documented not only an
association between body mass index (BMI) and mortality from
various types of cancer, but also provided a reliable estimate of
the contribution of overweight and obesity to the total mortality
from cancer (2). The authors
reported that women with the highest BMI (40 kg/m2) had
mortality rates from all types of cancer combined that were 62%
higher (with a relative risk of death of 1.62) than the rates of
women of normal weight. These authors reported a significant trend
whereby individuals with a higher BMI exhibited an increased risk
of succumbing to cancers of the breast, uterus, cervix and
ovary.
Numerous studies suggest health benefits from soy
consumption, including its role in the reduction of cardiovascular
disease and certain types of cancer (3,4).
Soybeans and soy protein products are a major source of
phytoestrogen, plant-based estrogen-like substances. Soybeans
contain isoflavones, which are structurally similar to mammalian
estrogens, have estrogenic properties and are potential
anticarcinogens (5). The most
common dietary isoflavones are genistein, daidzein and glycitein,
also known as isoflavonoids (6).
Gut flora metabolizes daidzein into equol, a non-steroidal estrogen
(6). It has been proposed that a
high consumption of soy foods is partially responsible for the
observed lower breast cancer rate in Asian women (7). However, results of these studies
conflict with those of studies concluding that certain components
of soy are estrogenic and therefore increase the breast cancer
risk.
Certain studies are available regarding the
protective effects of soy on breast cancer, while findings of other
studies report the adverse effects of soy. For example, soy intake
has been found to increase spontaneous mammary gland tumors in mice
(8). Moreover, the use of a soy
protein supplement for 2 weeks by women with benign or malignant
breast disease was found to have estrogenic effects on the breasts
of these individuals (9). On the
other hand, in a large cohort study of British women no effects of
dietary isoflavone intake and breast cancer risk were found among
either pre- or post-menopausal women (10). Experimental studies showed variable
results on breast cancer reduction by soy or the soy isoflavone
genistein (11–15). For example, it was reported that
genistein reduces the growth of tumors that arise from inoculated
human or mouse breast cancer carcinoma cells in pre-menopausal
mouse models (16,17). On the other hand, isolated genistein
was found to promote MNU-induced estrogen-dependent mammary
tumorigenesis and growth of the MCF-7 estrogen receptor
(ER)-positive human breast cancer cell line implanted in
ovariectomized rats and mice as a model for postmenopausal status
(18,19). Data on the effects of soy
consumption on breast health, particularly in obese women, are
lacking.
A number of epidemiological studies showed that
elevated levels of serum insulin-like growth factor-1 (IGF-1), a
mitogenic and antiapoptotic protein, are related to an increased
risk of breast cancer (20–27). IGF-1 levels are increased in obese
individuals and at least part of the greater risk of breast cancer
in obese women may be due to IGF-1 growth stimulation of breast
cells. IGF-1 activity is modulated by a family of IGF-binding
proteins, IGFBPs 1–6. These proteins bind and reduce the
availability of IGF-1 in serum. Increased IGFBP levels are
associated with lower bioavailable IGF-1 and are involved in the
antiproliferative effects of antiestrogens in human breast cancer
cell lines (28,29). IGF-1 and estrogen function as growth
factors for mammary epithelial cells and for mammary tumor cell
lines, operating to increase proliferation in breast cancer cell
lines. Additionally, IGF-1 is required for maximum ER activation in
these cells (28,29). Higher insulin levels are also
associated with obesity and may enhance cellular proliferation
leading to the development of breast cancer (30).
Of the IGFBPs, IGFBP-3 exhibits the highest
concentration in serum and the majority of IGF-1 in serum are bound
by IGFBP-3. IGFBP-3 with bound IGF-1 is thought to be inactive, but
IGFBP-3 can be degraded by proteases in tumor cells, thereby
releasing IGF-1 and increasing local IGF-1 bioavailability. Human
data regarding IGFBP-3 levels and breast cancer risk have been
inconclusive. Some studies show a relationship between breast
cancer risk and IGFBP-3 concentrations (23,24,26),
whereas in other studies a protective effect (22) or no association at all (31) was noted.
The Zucker rat (fa/fa) is the best known,
most widely used rat model for genetic obesity. Obesity in the
Zucker rat is inherited as an autosomal recessive trait caused by a
mutation (fa) in the leptin receptor gene (32,33),
discovered by Zucker and Zucker (34,35).
Animals homozygous for the fa allele were notably obese by
3–5 weeks of age and by 14 weeks of age their body composition was
>40% lipid (36). A number of
investigators have used this model to study the development,
etiology, associated pathogenesis, possible treatment and putative
mechanisms of severe obesity (37).
Obese Zucker rats develop hyperinsulinemia and insulin resistance
prior to the development of obesity-associated
non-insulin-dependent diabetes mellitus (NIDDM) in a manner similar
to that in humans (38). Lean
Zucker rats, by contrast, exhibit normal metabolic function and are
considered ideal controls. Consequently, this model can be used to
investigate the impact of soy consumption on mammary tumor
development in an obese animal model. No published data are
currently available regarding the effects of soy diet and obesity
on mammary tumor development in obese animals.
Material and methods
Experimental design
The animal protocols were approved by the
Institutional Animal Care and Use Committee (IACUC) at the
University of Arkansas for Medical Sciences. A total of 99
five-week-old female Zucker rats (45 obese fa/fa and 54
lean) were purchased from Harlan Industries (Indianapolis, IN,
USA). Harlan Industries performed genotyping to identify
fa/fa and lean/lean rats at the age of 24 days. After 1 week
of acclimation (age 42 days), rats were randomly assigned to
groups: i) lean, casein diet; ii) obese, casein diet; iii) lean,
soy protein diet; and iv) obese, soy protein diet. The rats were
housed 2 per cage with ad libitum access to water and a
semi-purified diet similar to the AIN-93G diet (Harlan Teklad,
Madison, WI, USA) prepared with dietary protein, either casein
(control) or an enzymatically treated soy protein isolate with high
isoflavones [3.24 mg total isoflavones/g protein (1.88 aglycone
equivalents/g protein), Lot no. M330024462; Solae LLC, St. Louis,
MO, USA]. The compositions of the two diets are described in
Table I. At age 50 days the rats
received 65 mg DMBA/kg body weight (Sigma Chemical Co., St. Louis,
MO, USA) in sesame oil, via gavage. Blood samples were drawn from
the tail vein at various ages (43, 99 and 197 days) to measure
serum IGF-1 using ELISA kit (IDS, Fountain Hills, AZ, USA) and
IGFBP-3 ELISA kit (DSL, Webster, TX, USA). Beginning 4 weeks after
DMBA treatment, the rats were palpated twice weekly for mammary
tumor detection. The detection date and location of each mammary
tumor were recorded for each rat. The rats were sacrificed 147 days
post-DMBA treatment. The mammary tumors were excised, counted and
weighed. Three rats with tumor masses >2.5 cm in diameter were
sacrificed early according to our IACUC-approved animal protocol.
The tumor sections were placed in 10% neutral-buffered formalin for
histopathologic analysis. Sections (4 μm) of the paraffin-embedded
tumors were stained with hematoxylin and eosin for light
microscopic evaluation.
 | Table IComposition of study diets. |
Table I
Composition of study diets.
| Ingredients | Casein
(g/kg)a | Soy protein
(g/kg) |
|---|
| Casein | 200.0 | - |
| Isolated soy
protein | - | 202.0 |
| L-Cystine | 3.0 | - |
| L-Methionine | - | 2.5 |
| L-Cystine | - | 1.3 |
| L-Tryptophan | - | 0.4 |
| L-Threonine | - | 0.3 |
| Corn starch | 397.5 | 409.0 |
| Maltodextrin | 132.0 | 132.0 |
| Sucrose | 100.0 | 108.0 |
| Corn oilb | 70.0 | 63.0 |
| Cellulose | 50.0 | 50.0 |
| AIN-93G mineral
mix | 35.0 | 35.0 |
| AIN-93G vitamin
mix | 10.0 | 10.0 |
| Choline
bitartrate | 2.5 | 2.5 |
| TBHQ,
antioxidant | 0.014 | 0.014 |
Pathology
A board-certified anatomic pathologist (S.K.)
evaluated the tumors in a blinded protocol. The tumors were
classified as benign (essentially unremarkable breast parenchyma
with non-proliferative fibrocystic changes); intraductal
proliferation (IDP), characterized by epithelial proliferation or
presence or multiple papillomas; ductal carcinoma in situ
(DCIS), uniform proliferation of tumor cells within pre-existing
structures without evidence of invasion to the surrounding tissue;
invasive ductal and lobular carcinoma (IDC), showing an invasive
(infiltrative) pattern of neoplastic cells; or Phyllodes,
characterized by a relatively well-demarcated fibroepithelial mass
showing marked stromal cellularity and numerous stromal mitosis,
similar to the pattern of the Phyllodes tumor observed in female
individuals.
Statistical analysis
A Kruskal-Wallis test was used to analyze the four
treatment groups, followed by between-group comparisons using the
Mann-Whitney U test due to unequal variances in the groups. A
two-way ANOVA was used to measure serum IGF-1 and IGFBP-3 levels,
followed by Tukey’s post-hoc test. A Kaplan-Meier analysis
of tumor latency was performed. A log-rank test was used to compare
the median tumor-free times. Fisher’s exact test was used to
compare the percentage of rats with tumors and tumor histology in
each group. The median number of tumors per tumor-bearing rat
(multiplicity) for each group was compared using the non-parametric
Kruskal-Wallis test for overall differences and the Mann-Whitney U
test for group comparisons. Statistical significance was set at
P<0.05 and the P-values were not adjusted for multiple
comparisons. For the rats that were sacrificed early due to tumor
burden, the number of tumors was assumed to have remained constant
until the end of the study. Data analyses were generated and the
plots were constructed using SPSS© version 17.0 for
Windows (SPSS Inc., Chicago, IL, USA) and SAS version 9.0 (SAS
Inc., Cary, NC, USA).
Results
Body weight
All of the rats gained weight during the course of
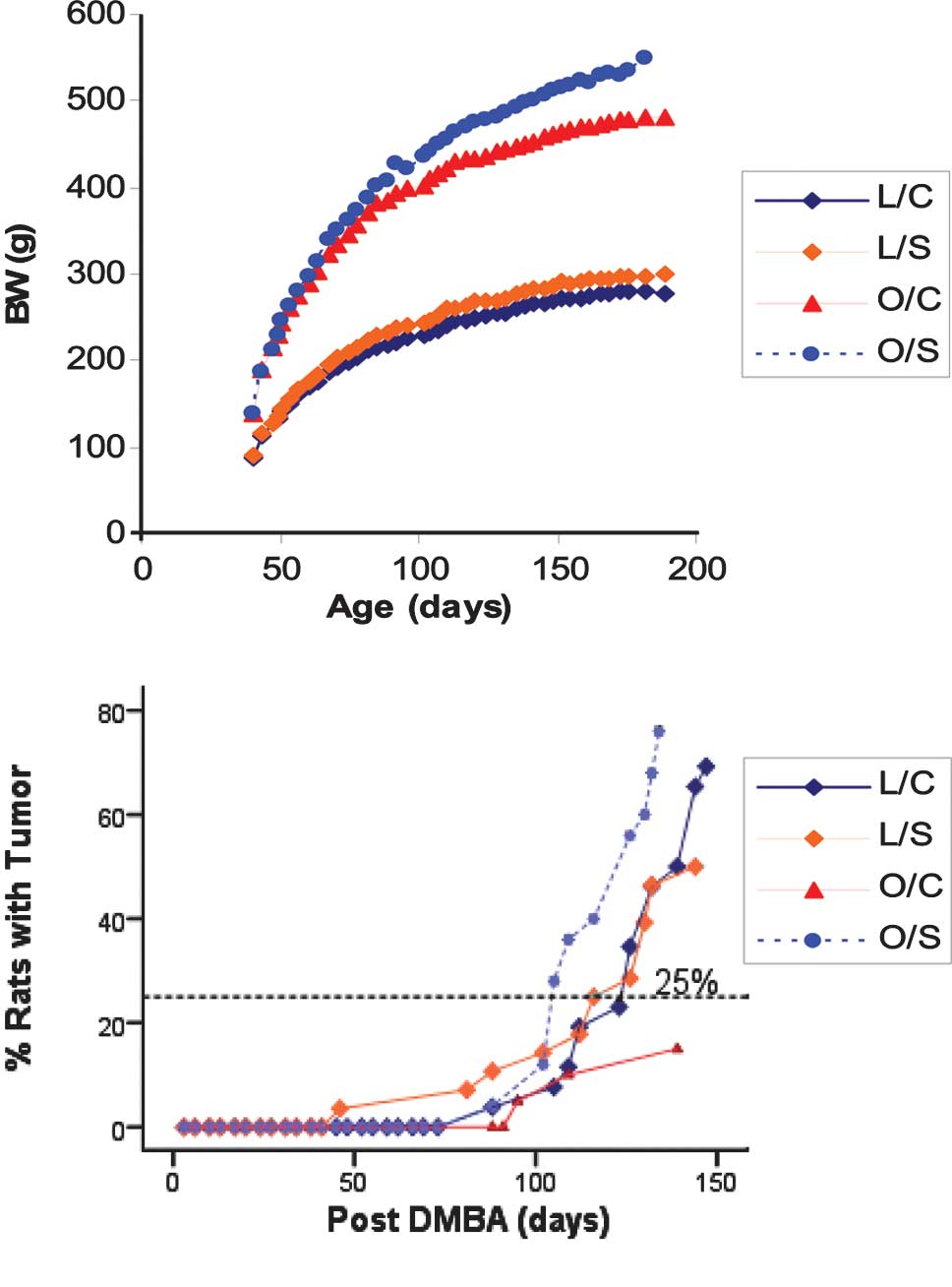
the experiment (Fig. 1A). Obese
rats gained significantly (P<0.001) more weight than lean rats.
Obese soy protein-fed rats were significantly heavier than obese
casein rats (543±11 vs. 484±8) (P<0.001). The lean soy
protein-fed rats also exhibited a higher body weight than the lean
casein-fed rats (300±5 vs. 283±4) (P=0.024).
Serum IGF and IGFBP-3 levels
At age 43 days, no differences were found in the
serum IGF-1 levels among the groups, but IGFBP-3 was increased in
the high-isoflavone soy protein-fed lean group compared to the
casein-fed lean rats (Table II).
At age 99 days, obese rats fed a casein diet exhibited increased
IGFBP-3 levels (P<0.001), but obese rats fed soy protein
exhibited a significant decrease in IGFBP-3 levels compared to the
lean soy protein-fed group (P<0.001). Moreover, obese soy
protein-fed rats exhibited a significant decrease in IGFBP-3 levels
compared to obese casein-fed rats (P<0.001). At the end of the
experiment, IGFBP-3 levels were increased in the obese casein- and
soy protein-fed rats (P<0.001).
| Table IISerum levels of IGF-1 and IGFBP-3
(mean ± standard error). |
Table II
Serum levels of IGF-1 and IGFBP-3
(mean ± standard error).
| | L/C (n=26) | L/S (n=28) | O/C (n=20) | O/S (n=25) |
|---|
| IGF-1 |
| Age (days) | 43 | 1.327±54 | 1.422±57 | 1.309±52 | 1.472±65 |
| 99 | 1.129±27a,c | 1.213±32 | 1.705±76b,d | 1.528±67b,e |
| 197 | 816±18 | 834±20 | 1.652±67b,d | 1.433±84b,e |
| IGFBP-3 |
| Age (days) | 43 | 69±2b,c | 79±1 | 78±2b,d | 78±1 |
| 99 | 44±2 | 46±1 | 59±3b,d | 40±1b,e,f |
| 197 | 16±2 | 19±2 | 40±5b,d | 30±3b,e |
Time course for tumor formation, latency
and multiplicity
The time course of palpable mammary tumor detection
is shown in Fig. 1B and the data
are presented in Table III.
| Table IIICharacteristics of DMBA-induced
mammary tumors. |
Table III
Characteristics of DMBA-induced
mammary tumors.
| L/C (n=26) | L/S (n=28) | O/C (n=20) | O/S (n=25) |
|---|
| Tumor onset |
| Day of first
tumora | 88 | 46 | 95 | 88 |
| Day at 25%
tumorsb | 124 | 116 | N/A | 106 |
| Tumor
incidence |
| Rats with tumors
(%)c | 69f,g | 50 | 15 | 76f,h |
|
Multiplicityd | 1 (1–4) | 1 (1–2) | 1 | 1 (1–3) |
| Total tumors
(n=82)e | 30 (37%) | 18 (21%) | 3 (4%) | 31 (38%) |
| Benign | 2 (7%) | 0 (0%) | 0 (0%) | 2 (6%) |
| IDP | 6 (20%) | 0 (0%) | 0 (0%) | 0 (0%) |
| DCIS | 21 (70%) | 9 (50%) | 2 (67%) | 16 (52%) |
| IDC | 0 (0%) | 6 (33%) | 1 (33%) | 11 (35%) |
| Phyllodes | 1 (3%) | 3 (17%) | 0 (0%) | 2 (7%) |
Tumor latency (the number of days post-DMBA
treatment until the detection of the first mammary tumor) was
longer in the obese casein-fed rats than the remaining three
groups. The first mammary tumor detected in lean soy protein-fed
rats was 46 days post-DMBA treatment compared to 88 days in lean
casein-fed rats. For the obese soy protein-fed rats, the first
mammary tumor was detected 88 days post-DMBA treatment, compared to
95 days in obese casein-fed rats. In addition, 25% of the lean soy
protein-fed rats developed mammary tumors 116 days post-DMBA
treatment, compared to 124 days for the lean casein-fed rats and
106 days for the obese soy protein-fed rats. By the end of the
experiment, 69% of the lean casein rats developed mammary tumors,
compared to 50% in the lean high-isoflavone soy protein group.
However, this difference was not statistically significant
(P=0.176). In the obese group, 76% of the high-isoflavone soy
protein-fed rats developed mammary tumors, compared to 15% of the
obese casein-fed rats (P<0.001). The lean casein-fed group had a
higher incidence of mammary tumor development than that of the
obese casein rats (69 vs. 15%) (P<0.001). A higher incidence of
mammary tumors was noted in the obese soy protein-fed rats than in
the lean soy protein-fed rats. However, this result was not
statistically significant (76 vs. 50%) (P=0.088). The median number
of mammary tumors per tumor-bearing rat (multiplicity) was
compared. No statistically significant difference was found among
the four groups. In regards to multiplicity, the lean soy
protein-fed rats had a range of 1–2 tumors per rat, compared to a
range of 1–4 tumors per rat in the lean casein-fed group. The tumor
multiplicity for obese casein-fed rats (1 tumor per rat) was lower
compared to the obese soy protein-fed rats (1–3 tumors per rat)
(Table III).
Mammary tumor characteristics
Mammary tumor histology data are presented in
Table III. A total of 82 mammary
tumors were detected in the study. Table III shows the light microscopic
classification of these mammary tumors in the four groups. A total
of 30 masses were detected (37% of the total tumors) in the lean
casein-fed group compared to 18 tumors (21%) in the lean soy
protein-fed group, 31 tumors (38%) in the obese soy protein-fed
rats and only 3 tumors (4%) in the obese casein-fed rats.
Pathological analysis of the 30 tumors in the lean casein-fed group
showed the following classification: 2 (7%) benign, 6 (20%) IDP, 21
(70%) DCIS and 1 (3%) Phyllodes. Of the 18 mammary tumors in the
lean soy protein-fed group 9 were DCIS (50%), 6 IDC (33%) and 3
Phyllodes (17%). For the obese casein-fed group, 2 tumors were
classified as DCIS and 1 tumor as IDC. Mammary tumors in the obese
soy protein-fed group included 2 (6%) benign, 16 (52%) DCIS, 11
(35%) IDC and 2 (7%) Phyllodes. While tumors classified as benign,
IDP, DCIS and IDC were observed in the author’s previously
published study on obesity and breast cancer in female Zucker rats
(39,40), tumors with the morphologic
characteristics of Phyllodes tumors have not been observed in prior
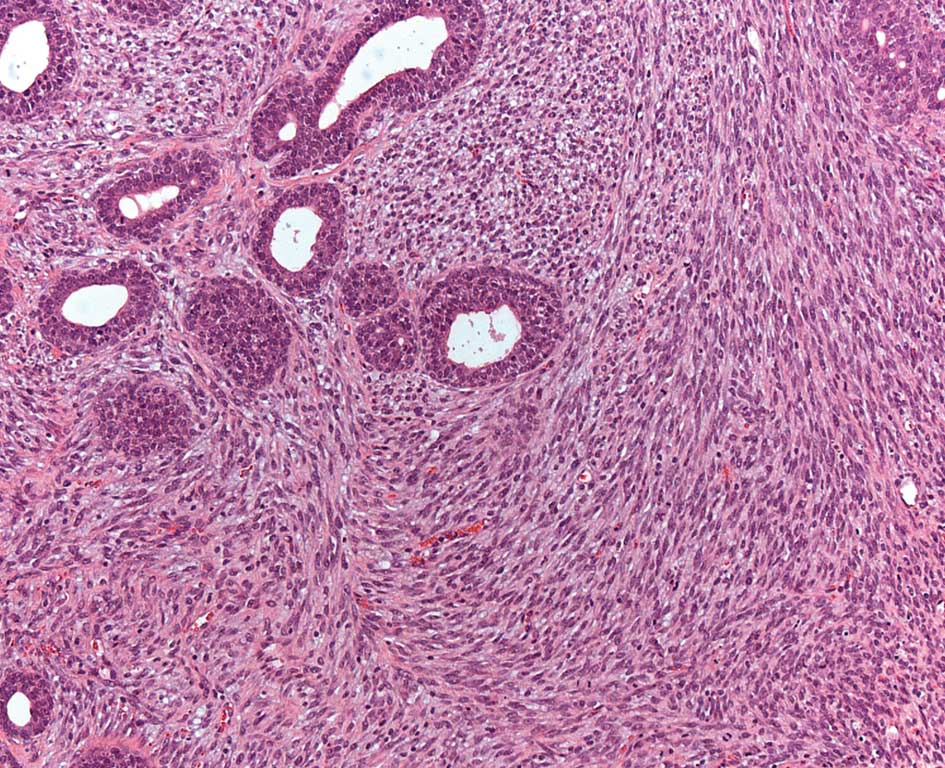
studies. A tumor classified as Phyllodes from this study is shown
in Fig. 2.
Discussion
This study aimed to compare the effects of long-term
high-isoflavone soy protein to those of long-term casein protein
intake and obesity on DMBA-induced mammary tumor development, as
well as to investigate the effects of high-isoflavone soy protein
and obesity on serum IGF-1 and IGFBP-3 levels.
Results of the present study showed that obese soy
protein-fed rats gained significantly more weight than obese
casein-fed rats (P<0.001) and the lean soy protein-fed rats
exhibited a slightly higher body weight than their lean casein-fed
counterparts (P=0.057). In the lean groups, 69% of the casein-fed
rats developed mammary tumors compared to 50% (P<0.08) of the
soy protein-fed rats. In the obese groups, 76% of the soy
protein-fed rats developed mammary tumors compared to 15%
(P<0.05) of the obese casein-fed rats. At age 99 days, obese
rats fed a casein diet exhibited increased IGFBP-3 levels
(P<0.001), but obese rats fed soy protein exhibited a
significant decrease in IGFBP-3 levels compared to the lean soy
protein-fed group (P<0.001). Additionally, IGFBP-3 levels in
obese soy protein-fed rats were significantly decreased compared to
obese casein-fed rats (P<0.001). At the end of the experiment,
the IGFBP-3 levels were found to have increased in obese casein-
and soy protein-fed rats (P<0.001).
Our observations regarding weight increase of obese
soy protein-fed compared to obese casein-fed rats (P<0.001) are
agreement with those of Tovar et al (41). These authors found that obese ZDF
fa/fa rats fed either a casein or soy protein diet gained
weight at a similar rate until day 140, but after day 150, rats fed
the casein diet began to lose weight compared to rats fed the soy
protein diet (P=0.004). By day 160 post-treatment, Tovar et
al observed that the casein-fed rats weighed significantly less
than the soy-fed rats. These authors concluded that the difference
in weight gain between groups is explained by a physical
deterioration observed in rats fed the casein diet in the last days
of the study. Another study reported that among obese rats fed
either casein or soy diets, casein-fed rats gained significantly
less weight than soy-fed rats, probably due to their physical
deterioration (42). In their
study, Mezei et al reported that when female obese Zucker
rats were fed with high- or low-isoflavone soy diets, the
high-isoflavone soy-fed rats gained more body weight than the
low-isoflavone soy-fed and casein-fed rats (P<0.05) (43).
Our results for casein-fed obese rats showing that
obesity protected against DMBA-induced mammary tumor development
are in agreement with epidemiological data showing that obesity
protects against breast cancer development in obese pre-menopausal
women (44–46).
Our results show that in obese Zucker rats, a soy
protein diet containing high isoflavone levels promoted
DMBA-induced mammary tumor development compared to a casein-based
diet. We found that lean and obese rats fed soy protein diets
developed more advanced mammary tumors, graded IDC, than their
respective controls in the casein group. Simmen et al
reported that when Sprague-Dawley rats were fed AIN-93G diets with
either casein or soy protein as the source of protein and treated
with N-methyl nitrosourea, rats on soy protein diets developed more
advanced grade tumors (47). The
mechanism underlying the higher grade tumors elicited by soy diet
has yet to be determined.
The presence of dietary isoflavones may affect the
development of mammary tumors, as previously reported (19,48).
For this reason, we used the purified diet AIN-93G, which contains
no soy or soy isoflavones. In our previous study (39), the standard natural ingredient diet
2018 was employed. The 2018 diet contains soybean meal with the
isoflavone levels ranging from 150 to 250 mg/kg diet, depending on
the batch. The presence of soy isoflavones in this diet may explain
the differences in mammary tumor induction observed in other
sutdies (39).
Meta-analyses on soy intake and breast cancer risk
have been reported. Epidemiological studies have showed that soy
consumption was associated with a reduction in breast cancer risk
in pre-but not postmenopausal women in Singapore (49). Another meta-analysis found an
inverse association between soy intake and breast cancer risk among
pre-menopausal women, but no effect was noted among postmenopausal
women (50). A meta-analysis of 18
epidemiological studies has shown that a high soy intake may be
associated with a small reduction of breast cancer risk, but this
association was not significant among women in Asian countries and
the reduction was more significant in pre-compared to
postmenopausal women (51). The
inverse association between soy food intake and breast cancer risk
was mainly noted in Asian women, with only one study reporting such
a correlation in women of Caucasian descent (52). The correlation of adolescent and
adult soy food intake with breast cancer risk was evaluated in a
cohort of 73,223 Chinese women who participated in the Shanghai
Women’s Health Study. A protective effect of soy food intake
against pre-menopausal breast cancer was noted (53). A recent meta-analysis of the
literature (a total of 34 studies) on the relationship between
dietary soy intake and breast cancer risk has concluded that the
intake of naturally occurring dietary soy food or its components
appears to be safe for women without breast cancer. However, the
safety of high levels of consumption of soy containing supplements
or soy components was less certain (54).
Experimental studies in non-obese animal models have
shown an association between serum IGF-1 concentrations and mammary
cancer risk. For example, liver IGF-1 deficient mice, which have
hepatic IGF-1 gene deletion, show a 75% reduction in serum IGF-1
levels, but normal growth and development. These mice have shown
delays in the onset of chemically-and genetically-induced mammary
tumors. Wu et al (55) used
an IGF-1 gene-deleted mouse model to show that the lack of IGF-1 in
these mice resulted in a significant delay in the onset of
DMBA-induced mammary tumors. These data indicate that tumor
development can be induced by IGF-1. Thus, modulation of IGF-1 and
IGFPB-3 concentration is a crucial strategy for determining the
effects of the incidence of breast cancer in obese Zucker rats.
Clinical studies on the effects of soy on IGF-1 levels were found
to be inconclusive, with some studies showing that soy increases
IGF-1 levels (56,57). However, these studies used subjects
with normal body weight and not obese ones. By contrast, other
studies have shown decreases in serum IGF-1 levels (58,59).
Our preliminary data using obese Zucker rats have shown that
obesity increases the serum IGF-1 levels (60). The impact of dietary factors on
IGF-1 levels in humans is limited to a number of observational
epidemiological studies (61–72).
Thus, no data regarding the effect of specific factors, such as soy
or isoflavones, are available. Since obesity is likely to increase
serum IGF-1 levels, and a soy diet with a high isoflavone content
modulates serum IGF-1 levels, we investigated the effects of a
soy-based diet with isoflavones on serum IGF-1 and IGFBP-3 levels
and its effect on DMBA-induced mammary tumors. Elevated IGF-1 has
been shown to be a risk factor for breast cancer in women (22,26,73).
It is thought that increased IGFBP-3 levels may be play a
protective role in breast cancer by decreasing the levels of
bioavailable IGF-1 (24,74).
For these reasons, we measured IGF-1 and IGFBP-3
serum levels at three time points in lean and obese casein- and soy
protein-fed rats. Lean soy protein-fed rats exhibited increased
IGFBP-3 levels compared to casein-fed rats. After 1 day on the
diets (Day 43), the lean soy protein-fed rats exhibited higher
IGFBP-3 levels compared to their lean casein-fed counterparts.
Short-term dietary intervention with soy isoflavones is known to
result in rapid changes in serum IGFBP-3 levels (75). Increased IGFBP-3 may reduce the
bioavailability of IGF-1 and be partially responsible for the
observed lower incidence (non-significant) of tumors in lean soy
protein-fed rats. However, in the obese groups, IGFBP-3 levels were
increased in casein-fed rats, while much lower levels were noted in
soy-fed rats. Our results clearly show that obesity increased serum
IGF-1 levels for casein-and soy protein-fed rats compared to lean
rats fed with either diet. By age 99 days, the obese casein-fed
rats exhibited increased IGFBP-3 levels compared to their soy
protein-fed counterparts. This increase in IGFBP-3 and the
resultant decrease in bioavailable IGF-1 may be the mechanism
responsible for the lower tumor numbers observed in obese
casein-fed compared to obese soy protein-fed animals.
To the best of our knowledge, this report is the
first to show that intact obese vs. lean Zucker female rats fed
semi-purified diets are protected against DMBA-induced mammary
tumors for a longer period of time while on casein vs.
high-isoflavone soy protein diets. The majority of the observed
tumors were epithelial in nature resembling the invasive and in
situ carcinoma observed in female individuals. However, we have
observed another rare tumor which mimics the Phyllodes tumors found
in women. Phyllodes are in general rare tumors and their etiology
has yet to be elucidated.
Our results suggest that the dietary protein source
affects serum IGFBP-3 levels in obese female Zucker rats.
Furthermore, changes in the concentration of IGFBP-3 may contribute
to the reduction of the incidence of DMBA-induced mammary
tumors.
Acknowledgements
The authors wish to thank John Gregan and Phaedra
Yount for valuable assistance in the preparation of this manuscript
on behalf of the Arkansas Children’s Hospital Research Institute.
This study was supported by the U.S. Department of Agriculture and
the Arkansas Biosciences Institute. R.H. applied for and received
funding, designed and supervised the study, and participated in the
collection of tumor data and the writing of the manuscript. S.S.
coordinated and performed the daily activities regarding overseeing
the animals, recording data and processing the samples. S.M.
participated in the study design, interpretation of study results
and participated in discussions involving the findings. C.H.J.
participated in the study design and performed the statistical
analyses. S.K. provided expertise in classifying and analyzing all
of the mammary tumors and participated in the interpretation of
study results and writing of this manuscript. The authors read and
approved the final manuscript.
Abbreviations:
|
DMBA
|
7,12-dimethylbenz(a)anthracene
|
|
BMI
|
body mass index
|
|
ER
|
estrogen receptor
|
|
IGF-1
|
insulin growth factor-1
|
|
IGFBP-3
|
insulin growth factor binding
protein-3
|
|
NIDDM
|
non-insulin-dependent diabetes
mellitus
|
|
IACUC
|
Institutional Animal Care and Use
Committee
|
References
|
1
|
American Cancer Society. Cancer Facts and
Figures. 2010.
|
|
2
|
Calle EE, Rodriguez C, Walker-Thurmond K
and Thun MJ: Overweight, obesity, and mortality from cancer in a
prospectively studied cohort of U.S. adults. N Engl J Med.
348:1625–1638. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hakkak R, Korourian S, Shelnutt SR,
Lensing S, Ronis MJ and Badger TM: Diets containing whey proteins
or soy protein isolate protect against
7,12-dimethylbenz(a)anthracene-induced mammary tumors in female
rats. Cancer Epidemiol Biomarkers Prev. 9:113–117. 2000.PubMed/NCBI
|
|
4
|
Kris-Etherton PM, Hecker KD, Bonanome A,
et al: Bioactive compounds in foods: their role in the prevention
of cardiovascular disease and cancer. Am J Med. 113(Suppl 9):
71–88. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Peeters PH, Keinan-Boker L, van der Schouw
YT and Grobbee DE: Phytoestrogens and breast cancer risk. Review of
the epidemiological evidence. Breast Cancer Res Treat. 77:171–183.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kelly GE, Nelson C, Waring MA, Joannou GE
and Reeder AY: Metabolites of dietary (soya) isoflavones in human
urine. Clin Chim Acta. 223:9–22. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Adlercreutz CH, Goldin BR, Gorbach SL, et
al: Soybean phytoestrogen intake and cancer risk. J Nutr.
125:S757–S770. 1995.
|
|
8
|
Gridley DS, Kettering JD, Slater JM and
Nutter RL: Modification of spontaneous mammary tumors in mice fed
different sources of protein, fat and carbohydrate. Cancer Lett.
19:133–146. 1983.PubMed/NCBI
|
|
9
|
Hargreaves DF, Potten CS, Harding C, et
al: Two-week dietary soy supplementation has an estrogenic effect
on normal premenopausal breast. J Clin Endocrinol Metab.
84:4017–4024. 1999.PubMed/NCBI
|
|
10
|
Travis RC, Allen NE, Appleby PN, Spencer
EA, Roddam AW and Key TJ: A prospective study of vegetarianism and
isoflavone intake in relation to breast cancer risk in British
women. Int J Cancer. 122:705–710. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cohen LA, Zhao Z, Pittman B and Scimeca
JA: Effect of intact and isoflavone-depleted soy protein on
NMU-induced rat mammary tumorigenesis. Carcinogenesis. 21:929–935.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Constantinou AI, Lantvit D, Hawthorne M,
Xu X, van Breemen RB and Pezzuto JM: Chemopreventive effects of soy
protein and purified soy isoflavones on DMBA-induced mammary tumors
in female Sprague-Dawley rats. Nutr Cancer. 41:75–81. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gallo D, Giacomelli S, Cantelmo F, et al:
Chemoprevention of DMBA-induced mammary cancer in rats by dietary
soy. Breast Cancer Res Treat. 69:153–164. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jin Z and MacDonald RS: Soy isoflavones
increase latency of spontaneous mammary tumors in mice. J Nutr.
132:3186–3190. 2002.PubMed/NCBI
|
|
15
|
Ueda M, Niho N, Imai T, et al: Lack of
significant effects of genistein on the progression of
7,12-dimethylbenz(a)anthracene-induced mammary tumors in
ovariectomized Sprague-Dawley rats. Nutr Cancer. 47:141–147. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hewitt AL and Singletary KW: Soy extract
inhibits mammary adenocarcinoma growth in a syngeneic mouse model.
Cancer Lett. 192:133–143. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shao ZM, Wu J, Shen ZZ and Barsky SH:
Genistein exerts multiple suppressive effects on human breast
carcinoma cells. Cancer Res. 58:4851–4857. 1998.PubMed/NCBI
|
|
18
|
Allred CD, Allred KF, Ju YH, et al:
Dietary genistein results in larger MNU-induced, estrogen-dependent
mammary tumors following ovariectomy of Sprague-Dawley rats.
Carcinogenesis. 25:211–218. 2004. View Article : Google Scholar
|
|
19
|
Ju YH, Allred CD, Allred KF, Karko KL,
Doerge DR and Helferich WG: Physiological concentrations of dietary
genistein dose-dependently stimulate growth of estrogen-dependent
human breast cancer (MCF-7) tumors implanted in athymic nude mice.
J Nutr. 131:2957–2962. 2001.
|
|
20
|
Bruning PF, van Doorn J, Bonfrer JM, et
al: Insulin-like growth-factor-binding protein 3 is decreased in
early-stage operable pre-menopausal breast cancer. Int J Cancer.
62:266–270. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Del Giudice ME, Fantus IG, Ezzat S,
McKeown-Eyssen G, Page D and Goodwin PJ: Insulin and related
factors in premenopausal breast cancer risk. Cancer Res Treat.
47:111–120. 1998.PubMed/NCBI
|
|
22
|
Hankinson SE, Willett WC, Colditz GA, et
al: Circulating concentrations of insulin-like growth factor-I and
risk of breast cancer. Lancet. 351:1393–1396. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Krajcik RA, Borofsky ND, Massardo S and
Orentreich NI: Insulin-like growth factor I (IGF-I), IGF-binding
proteins, and breast cancer. Cancer Epidemiol Biomarkers Prev.
11:1566–1573. 2002.PubMed/NCBI
|
|
24
|
Muti P, Quattrin T, Grant BJ, et al:
Fasting glucose is a risk factor for breast cancer: a prospective
study. Cancer Epidemiol Biomarkers Prev. 11:1361–1368.
2002.PubMed/NCBI
|
|
25
|
Peyrat JP, Bonneterre J, Hecquet B, et al:
Plasma insulin-like growth factor-1 (IGF-1) concentrations in human
breast cancer. Eur J Cancer Prev. 29A:492–497. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Toniolo P, Bruning PF, Akhmedkhanov A, et
al: Serum insulin-like growth factor-I and breast cancer. Int J
Cancer. 88:828–832. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yu H, Jin F, Shu XO, et al: Insulin-like
growth factors and breast cancer risk in Chinese women. Cancer
Epidemiol Biomarkers Prev. 11:705–712. 2002.PubMed/NCBI
|
|
28
|
Khandwala HM, McCutcheon IE, Flyvbjerg A
and Friend KE: The effects of insulin-like growth factors on
tumorigenesis and neoplastic growth. Endocr Rev. 21:215–244. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yu H and Rohan T: Role of the insulin-like
growth factor family in cancer development and progression. J Natl
Cancer Inst. 92:1472–1489. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bruning PF, Bonfrer JM, van Noord PA, Hart
AA, de Jong-Bakker M and Nooijen WJ: Insulin resistance and
breast-cancer risk. Int J Cancer. 52:511–516. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kaaks R, Lundin E, Rinaldi S, et al:
Prospective study of IGF-I, IGF-binding proteins, and breast cancer
risk, in northern and southern Sweden. Cancer Causes Control.
13:307–316. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tartaglia LA, Dembski M, Weng X, et al:
Identification and expression cloning of a leptin receptor, OB-R.
Cell. 83:1263–1271. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chua SC Jr, Chung WK, Wu-Peng XS, et al:
Phenotypes of mouse diabetes and rat fatty due to mutations in the
OB (leptin) receptor. Science. 271:994–996. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zucker L and Zucker TF: Fatty, a new
mutation in the rat. J Heredity. 52:275–278. 1961.
|
|
35
|
Zucker TF and Zucker LM: Fat accretion and
growth in the rat. J Nutr. 80:6–19. 1963.
|
|
36
|
Zucker LM: Fat mobilization in vitro and
in vivo in the genetically obese Zucker rat ‘Fatty’. J Lipid Res.
13:234–243. 1972.
|
|
37
|
Bray GA, York DA and Fisler JS:
Experimental obesity: a homeostatic failure due to defective
nutrient stimulation of the sympathetic nervous system. Vitam Horm.
45:1–125. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bray GA: The Zucker-fatty rat: a review.
Fed Proc. 36:148–153. 1977.
|
|
39
|
Hakkak R, Holley AW, MacLeod S, et al:
Obesity promotes 7,12-dimethylbenz(a)anthracene-induced mammary
tumor development in female Zucker rats. Breast Cancer Research.
7:R627–R633. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hakkak R, MacLeod S, Shaaf S, et al:
Obesity increases the incidence of
7,12-dimethylbenz(a)anthracene-induced mammary tumors in an
ovariectomized Zucker rat model. Int J Oncol. 30:557–563.
2007.PubMed/NCBI
|
|
41
|
Tovar AR, Torre-Villavazo I, Ochoa M, et
al: Soy protein reduces hepatic lipotoxicity in hyperinsulinemic
obese Zucker fa/fa rats. J Lipid Res. 46:1823–1832. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Trujillo J, Ramirez V, Perez J, et al:
Renal protection by a soy diet in obese Zucker rats is associated
with restoration of nitric oxide generation. Am J Physiol Renal
Physiol. 288:F108–F116. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Mezei O, Banz WJ, Steger RW, Peluso MR,
Winters TA and Shay N: Soy isoflavones exert antidiabetic and
hypolipidemic effects through the PPAR pathways in obese Zucker
rats and murine raw 264.7 cells. J Nutr. 133:1238–1243.
2003.PubMed/NCBI
|
|
44
|
Pathak DR and Whittemore AS: Combined
effects of body size, parity, and menstrual events on breast cancer
incidence in seven countries. Am J Epidemiol. 135:153–168.
1992.PubMed/NCBI
|
|
45
|
Peacock SL, White E, Daling JR, Voigt LF
and Malone KE: Relation between obesity and breast cancer in young
women. Am J Epidemiol. 149:339–346. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Weiderpass E, Braaten T, Magnusson C, et
al: A prospective study of body size in different periods of life
and risk of premenopausal breast cancer. Cancer Epidemiol
Biomarkers Prev. 13:1121–1127. 2004.PubMed/NCBI
|
|
47
|
Simmen RC, Eason RR, Till SR, et al:
Inhibition of NMU-induced mammary tumorigenesis by dietary soy.
Cancer Lett. 224:45–52. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Hsieh CY, Santell RC, Haslam SZ and
Helferich WG: Estrogenic effects of genistein on the growth of
estrogen receptor-positive human breast cancer (MCF-7) cells in
vitro and in vivo. Cancer Res. 58:3833–3838. 1998.PubMed/NCBI
|
|
49
|
Lee HP, Gourley L, Duffy SW, Esteve J, Lee
J and Day NE: Dietary effects on breast-cancer risk in singapore.
Lancet. 337:1197–1200. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wu AH, Yu MC, Tseng CC and Pike MC:
Epidemiology of soy exposures and breast cancer risk. Br J Cancer.
98:9–14. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Trock BJ, Hilakivi-Clarke L and Clarke R:
Meta-analysis of soy intake and breast cancer risk. J Natl Cancer
Inst. 98:459–471. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Messina MJ: Emerging evidence on the role
of soy in reducing prostate cancer risk. Nutr Rev. 61:117–131.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Lee SA, Shu XO, Li H, et al: Adolescent
and adult soy food intake and breast cancer risk: results from the
Shanghai Women’s Health Study. Am J Clin Nutr. 89:1920–1926.
2009.PubMed/NCBI
|
|
54
|
Enderlin CA, Coleman EA, Stewart CB and
Hakkak R: Dietary soy intake and breast cancer risk. Oncol Nurs
Forum. 36:531–539. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wu Y, Cui K, Miyoshi K, et al: Reduced
circulating insulin-like growth factor I levels delay the onset of
chemically and genetically induced mammary tumors. Cancer Res.
63:4384–4388. 2003.PubMed/NCBI
|
|
56
|
Arjmandi BH, Khalil DA, Smith BJ, et al:
Soy protein has a greater effect on bone in postmenopausal women
not on hormone replacement therapy, as evidenced by reducing bone
resorption and urinary calcium excretion. J Clin Endocrinol Metab.
88:1048–1054. 2003. View Article : Google Scholar
|
|
57
|
Khalil DA, Lucas EA, Juma S, Smith BJ,
Payton ME and Arjmandi BH: Soy protein supplementation increases
serum insulin-like growth factor-I in young and old men but does
not affect markers of bone metabolism. J Nutr. 132:2605–2608.
2002.PubMed/NCBI
|
|
58
|
Sanderson M, Shu XO, Yu H, et al:
Insulin-like growth factor-I, soy protein intake, and breast cancer
risk. Nutr Cancer. 50:8–15. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Wangen KE, Duncan AM, Merz-Demlow BE, et
al: Effects of soy isoflavones on markers of bone turnover in
premenopausal and postmenopausal women. J Clin Endocr Metab.
85:3043–3048. 2000.PubMed/NCBI
|
|
60
|
Hakkak R, Holley AW, Bunn RC, Winters A
and MacLeod S: Effects of obesity on serum insulin growth factor 1
(IGF-1) levels in lean and obese female Zucker rats following DMBA
treatment. FASEB J. 19:A9932005.
|
|
61
|
Allen NE, Appleby PN, Davey GK, Kaaks R,
Rinaldi S and Key TJ: The associations of diet with serum
insulin-like growth factor I and its main binding proteins in 292
women meat-eaters, vegetarians, and vegans. Cancer Epidemiol
Biomarkers Prev. 11:1441–1448. 2002.PubMed/NCBI
|
|
62
|
Devine A, Rosen C, Mohan S, Baylink D and
Prince RL: Effects of zinc and other nutritional factors on
insulin-like growth factor I and insulin-like growth factor binding
proteins in postmenopausal women. Am J Clin Nutr. 68:200–206.
1998.PubMed/NCBI
|
|
63
|
Giovannucci E, Pollak M, Liu Y, et al:
Nutritional predictors of insulin-like growth factor I and their
relationships to cancer in men. Cancer Epidemiol Biomarkers Prev.
12:84–89. 2003.PubMed/NCBI
|
|
64
|
Gunnell D, Oliver SE, Peters TJ, et al:
Are diet-prostate cancer associations mediated by the IGF axis? A
cross-sectional analysis of diet, IGF-I and IGFBP-3 in healthy
middle-aged men. Br J Cancer. 88:1682–1686. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Heald AH, Cade JE, Cruickshank JK,
Anderson S, White A and Gibson JM: The influence of dietary intake
on the insulin-like growth factor (IGF) system across three ethnic
groups: a population-based study. Public Health Nutr. 6:175–180.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Holmes MD, Pollak MN, Willett WC and
Hankinson SE: Dietary correlates of plasma insulin-like growth
factor I and insulin-like growth factor binding protein 3
concentrations. Cancer Epidemiol Biomarkers Prev. 11:852–861.
2002.
|
|
67
|
Ma J, Giovannucci E, Pollak M, et al: Milk
intake, circulating levels of insulin-like growth factor-I, and
risk of colorectal cancer in men. J Natl Cancer Inst. 93:1330–1336.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Mucci LA, Tamimi R, Lagiou P, et al: Are
dietary influences on the risk of prostate cancer mediated through
the insulin-like growth factor system? BJU Int. 87:814–820. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Nagata C, Shimizu H, Takami R, Hayashi M,
Takeda N and Yasuda K: Dietary soy and fats in relation to serum
insulin-like growth factor-1 and insulin-like growth factor-binding
protein-3 levels in premenopausal Japanese women. Nutr Cancer.
45:185–189. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Probst-Hensch NM, Wang H, Goh VH, Seow A,
Lee HP and Yu MC: Determinants of circulating insulin-like growth
factor I and insulin-like growth factor binding protein 3
concentrations in a cohort of Singapore men and women. Cancer
Epidemiol Biomarkers Prev. 12:739–746. 2003.PubMed/NCBI
|
|
71
|
Signorello LB, Kuper H, Lagiou P, et al:
Lifestyle factors and insulin-like growth factor 1 levels among
elderly men. Eur J Cancer Prev. 9:173–178. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Vrieling A, Voskuil DW, Bueno-de-Mesquita
HB, et al: Dietary determinants of circulating insulin-like growth
factor (IGF)-I and IGF binding proteins 1, -2 and -3 in women in
the Netherlands. Cancer Causes Control. 15:787–796. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Rinaldi S, Toniolo P, Muti P, et al:
IGF-I, IGFBP-3 and breast cancer in young women: a pooled
re-analysis of three prospective studies. Eur J Cancer Prev.
14:493–496. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Bohlke K, Cramer DW, Trichopoulos D and
Mantzoros CS: Insulin-like growth factor-I in relation to
premenopausal ductal carcinoma in situ of the breast. Epidemiology.
9:570–573. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Woodside JV, Campbell MJ, Denholm EE, et
al: Short-term phytoestrogen supplementation alters insulin-like
growth factor profile but not lipid or antioxidant status. J Nutr
Biochem. 17:211–215. 2006. View Article : Google Scholar : PubMed/NCBI
|