Introduction
NAD+ glycohydrolases (EC 3.2.2.5)
catalyze the hydrolysis of NAD+ into ADP-ribose and
nicotinamide. Most eukaryotic NAD+ hydrolases are
localised at the outer surface of the cell membrane, and
erythrocytes are particularly rich in such ecto-NAD+
glycohydrolases (1).
NAD+ glycohydrolase and closely associated ADP-ribosyl
cyclase activities pertain to a protein that has been identified
immunologically as the lymphocyte surface antigen CD38 (2–4) and
purified to homogeneity (2). CD38
expression is also widespread beyond the hematopoietic system, with
CD38 having been found in different cell systems and tissues
(5).
Erythrocyte as well as serum NAD+
glycohydrolase and ADP-ribosyl cyclase activities were found to be
elevated in cancer, as were the carcinoembryonic antigen (CEA)
values (6,7). These results suggested that CD38
expression provides a prognostic outlook on tumor progression, as
is the case in monitoring HIV infection and recovery (8).
Subsequently, an animal model was used in this study
that would provide additional data to confirm the results obtained
with blood samples from cancer patients and to provide insight into
the mechanisms underlying these results. To that effect, BALB/c
mice to which Ehrlich ascites tumor cells (EATC) were applied by
intraperitoneal injections were used as the experimental model. In
these mice, the development of ascites was accompanied by a
considerable elevation in CD38 expression. A similar effect was
achieved by administration of the serum samples from mice with
fully-developed tumors or from cancer patients as well as EATC
culture medium (EATC supernatant) to normal mice or by their
addition to lymphocyte culture. The biological relevance of these
results in terms of a response of the organism to cell
proliferation processes is discussed, as well as their relationship
to tumor- and immune system-generated cytokines.
Materials and methods
Materials
[Carbonyl-14C] NAD+ (specific
activity 53 Ci/mol) was purchased from Amersham International.
Chemicals of analytical grade were obtained from Sigma Chemical Co.
(St. Louis, MO, USA) and AG-X4 resin from Bio-Rad Laboratories
(Hercules, CA, USA). Amerlite CEA-60 assay kit was purchased from
Kodak Clinical Diagnostics (Rochester, NY, USA). EATC were obtained
from the Research Institute for Experimental Medicine of Istanbul
University where the cells had been maintained through successive
passages in BALB/c mice over a long period of time. Mouse
CD38-specific goat polyclonal IgG, CD-38 (M-19), which was
purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA), was
used in the experiments.
Blood samples were obtained from apparently healthy
individuals (controls) and from one of the Tumor Marker
Laboratories of the Istanbul Faculty of Medicine. Erythrocytes were
purified from anticoagulated blood samples by centrifugation for 20
min at 3,000 × g through a Histopaque gradient (Histopaque-1077;
Sigma, St. Louis, MO, USA) and resuspended in phosphate-buffered
saline (PBS). Erythrocyte numbers were determined after
spectroscopic measurements were obtained using calibration curves
where 106 erythrocytes corresponded to 0.6
A400. Erythrocyte ghosts were prepared as previously
described (9). Erythrocytes were
briefly suspended in H2O and lysed by repeated drawing
and pressing of the suspension through the outlet of a
micropipette. The lysate was centrifuged for 10 min at 10,000 × g
and the pellet was washed repeatedly by resuspension and
centrifugation in PBS.
Lymphocytes were isolated by layering the blood on a
Histopaque gradient and undergoing centrifugation for 30 min at 400
× g. Interfaced cells were harvested, resuspended in PBS and washed
twice in PBS by centrifugation for 10 min at 3000 × g in order to
separate lymphocytes from thrombocytes (10). The lymphocyte pellet was finally
resuspended in PBS. For membrane isolation, the lymphocytes were
lysed by 3 freeze/thaw cycles and the supernatants, obtained by
centrifugation of lysates for 10 min at 3000 × g, were used as the
lymphocyte membrane fraction (11).
The total cell counts were determined on aliquots using a
hemocytometer. Cell viability was determined by trypan blue
exclusion by mixing one drop of trypan blue with an aliquot of cell
suspension and examining the cells under a microscope for dye
exclusion (12).
Animal model
EATC (n=8×105) was administered in 0.2 ml
PBS to BALB/c mice by intraperitoneal injections. For each group,
four mice were used. Alternatively, 50 μl of ascites fluid or serum
from mice with developed EAT, serum from cancer patients or EATC
supernatant were administered intraperitoneally to new groups of
mice. Where indicated, these injections were repeated on day 4.
Control groups of mice received PBS, irradiated EATC (50 Gy total
dose) or control serum. As an alternative to irradiated EATC, EATC
incubated with 1 mM cycloheximide were used, and similar results
were obtained. If not otherwise indicated, blood samples were
obtained from such mice on every fourth day and subjected to assays
as described below.
Lymphocyte culture
Peripheric mouse lymphocytes isolated as described
above by centrifugation in Histopaque gradients were cultured in a
humidified atmosphere of 5% CO2 in 2 ml Dulbecco’s
modified Eagle’s medium (DMEM) in 24-well plates at 37°C for 7
days. The lymphocytes were supplemented with 10% fetal calf serum
(FCS), serum from normal BALB/c mice, serum from BALB/c mice with
developed EAT (Day 8), serum from normal individuals, serum from
cancer patients with high CEA values, ascites fluid or EATC
supernatant, obtained as described below.
Ehrlich ascites tumor cell culture
EATC were propagated in RPMI-1640, supplemented with
10% FCS, then with 1% FCS and finally with serum-free medium with
no supplements. EATC culture was maintained in this manner over
several passages and for several weeks. The resulting cell culture
medium (EATC supernatant) was then assayed either directly or after
concentration, using IVSS vivaspin 2 centrifugal concentrator
(Sartorius Stedim Biotech, Aubagne, France).
Assay for NAD+ glycohydrolase
activity
NAD+ glycohydrolase activity in serum
samples was determined by separation of [carbonyl-14C]
nicotinamide released from [carbonyl-14C]
NAD+ on Bio-Rad AG1×4 anion exchange resin (13). Reaction mixtures (20 μl) containing
12 μl serum, 10 mM NaCl, 500 μM ZnCl2, 50 μM
CaCl2, 20 mM Tris-HCl, pH 9.0 and 5 μM
[carbonyl-14C] NAD+ were incubated for 30 min
at 37°C (7). Reactions were stopped
with 1 ml 0.1% SDS. The samples were then applied to the Bio-Rad
AG1×4 column and [carbonyl-14C] nicotinamide was eluted
with H2O. Unhydrolised [carbonyl-14C]
NAD+ was retained on the column and then eluted with 0.5
M NaCl. Radioactivity was determined by counting aliquots from the
eluate in a liquid scintillation counter (Packard Tri-Carb 1000 TK,
Meriden, CT, USA); counting efficiency for 14C was 90%.
Ecto-NAD+ glycohydrolase activity was assayed in 20 μl
reaction mixtures containing 5×106 erythrocytes (or
5×105 lymphocytes) in PBS. After incubation for 30 min
at 37°C and subsequent centrifugation of cells, the supernatants
were subjected to analysis as above (14).
SDS-PAGE and Western blotting
SDS-PAGE and Western blotting using the
CD38-specific goat polyclonal IgG antibody (M-19) were performed as
previously described (15,16). The erythrocyte ghost or lymphocyte
membrane fractions were solubilised in 1 M Tris-HCl, pH 6.8, 2%
glycerin, 10% SDS, 5% 2-mercaptoethanol, 0.1% bromphenol blue, and
subjected to SDS-PAGE. The separated proteins were then transferred
electrophoretically onto nitrocellulose membranes (Schleicher &
Schuell BioScience, Dassel, Germany). The blots were blocked by 1 h
incubation with 0.5% BSA in TBST (10 mM Tris-HCl, pH 8.0, 150 mM
NaCl and 0.05% Tween-20) followed by successive 1 h incubations
with CD38-specific goat polyclonal IgG (M-19). Detection of
immunocomplexes was achieved using alkaline phosphatase-conjugated
bovine anti-goat antibody (Sigma) (1:1000 in TBST) and BCIP/NBP as
a substrate (7).
The procedure described by the producing company was
followed for the determination of CEA.
Flow cytometry
Mouse peripheric lymphocytes (n=10,000), which were
prepared as described above were resuspended in 100 μl PBS and
incubated with anti-mouse CD38 monoclonal antibody, fluorescein
(FITC) conjugated (Cedarlane), at a concentration of 1 μg/ml for 30
min at 4°C. Lymphocytes were then washed three times in PBS and
analyzed on a FACSCalibur flow cytometer (Becton-Dickinson, San
Jose, CA, USA), equipped with a 5-W argon ion laser. FITC
excitation occured at 488 nm, and forward and side scatter
properties of lymphocytes were used to establish sorting gates.
Data acquisition and analysis were carried out using CellQuest Pro
software (Becton-Dickinson).
Statistical analysis
Each experiment was repeated at least four times and
in each experiment each value was the mean of 4 mice. Statistical
analysis was performed according to the Kruskal-Wallis ANOVA test
and Kruskal-Wallis multiple comparison z-value test.
The investigation was carried out in line with the
ethical principles and approval of the Ethics Committee of the
Istanbul Faculty of Medicine.
Results
Effect of administration of live EATC to
BALB/c mice on CD38-expression
The objective of this study was to reproduce, in an
appropriate animal model, the increases observed in serum and
erythrocyte NAD+ glycohydrolase in blood samples from
cancer patients. To that effect, EATC were administered to BALB/c
mice by intraperitoneal injections and enzyme activities were then
followed up concomitantly to ascites tumor development. In general,
a complete image of a developed ascites tumor was observed in these
animals in the course of 10–12 days after the injection of EATC. A
leukocytosis of 18,000–20,000 (cells/μl) appeared to precede the
onset of ascites symptoms. It was already detected on day 4 after
the application of EATC and remained at this level until the
termination of the experiment.
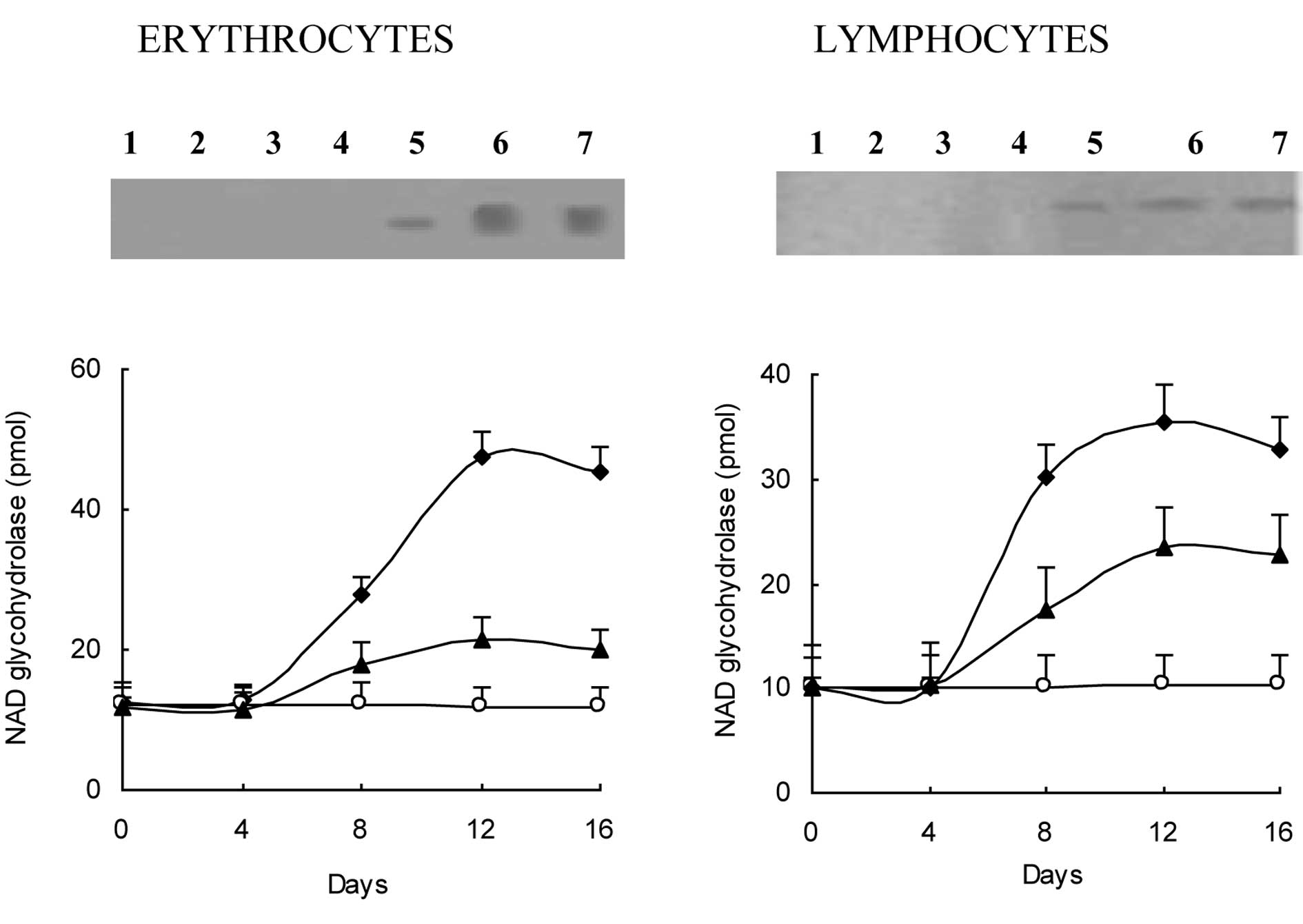
The development of ascites tumors was accompanied by
a considerable enhancement in CD38 expression, as revealed by a
comparison of anti-CD38 reactive proteins in erythrocyte ghost and
lymphocyte membrane fractions in mice with developed ascites tumors
versus the controls (Fig. 1A). In
line with recent findings of studies pertaining to cancer cases
(7), an anti-CD38 reactive protein
band of 45 kDa was detected in the Western blot analyses of
erythrocyte ghost proteins from mice to which live EATC had been
administered. In these mice, the 45 kDa band became visible 8 days
after the EATC injection and reached a maximum intensity by day 12
after the injection. Western blot analyses of lymphocyte membrane
proteins revealed similar results. No anti-CD38 reactive protein
band was found in the controls. Western blot analysis findings were
supported by the increases observed in erythrocyte and lymphocyte
NAD+ glycohydrolase (Fig.
1B). Elevations in these activities were observed 8 days after
the application of EATC and reached maximum values after 12 days.
These values were maintained until day 20 when the experiment was
terminated. The increases in erythro- cyte and lymphocyte
NAD+ glycohydrolase activities were on average not less
than approximately 3-fold higher than the controls. However, in
this case, the increases caused by tumor development were 2- to
3-fold higher than the controls.
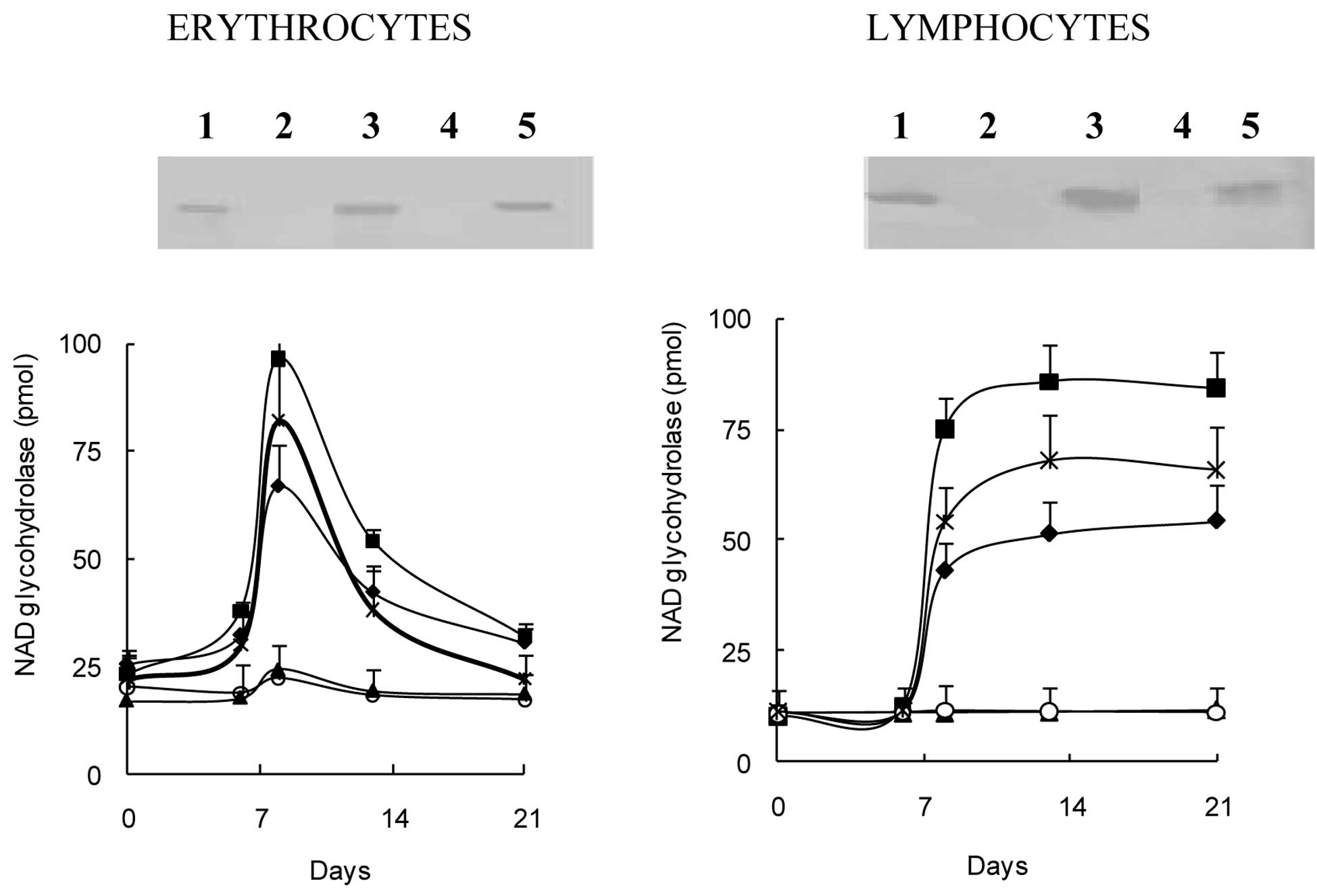
Stimulatory effect of of serum and
ascites samples on CD38-expression
The results suggest the existence of certain serum
factors as mediators of the stimulatory effect on CD38 expression
and related enzymatic activities. Thus, the experiments were
repeated with serum and ascites fluid samples from mice with fully
developed tumors as well as serum samples from cancer patients. The
administration of these samples was carried out twice with a 4-day
interval was almost as effective as that of live Ehrlich cells in
inducing CD38-expression. As shown in Fig. 2A, the injection of serum and ascites
samples from mice with developed EATC, as well as serum samples
from cancer patients with high (>100 ng/ml) CEA values again
gave rise to the appearance of the anti-CD38 reactive band of 45
kDa in Western blot analyses of erythrocyte ghost and lymphocyte
membrane proteins. The administration of serum samples from control
mice and normal individuals failed to exhihibit a similar inducing
effect. The serum samples from mice with tumors also gave rise to
increases in NAD+ glycohydrolase (Fig. 2B). Ascites fluid had a similar and
even a higher stimulatory impact. The highest stimulation was
achieved with intraperitoneal injections of serum samples from
cancer patients. The elevated activity declined gradually in
erythrocytes from day 12 to reach control levels by day 20, but
activity persisted in the lymphocytes until day 20.
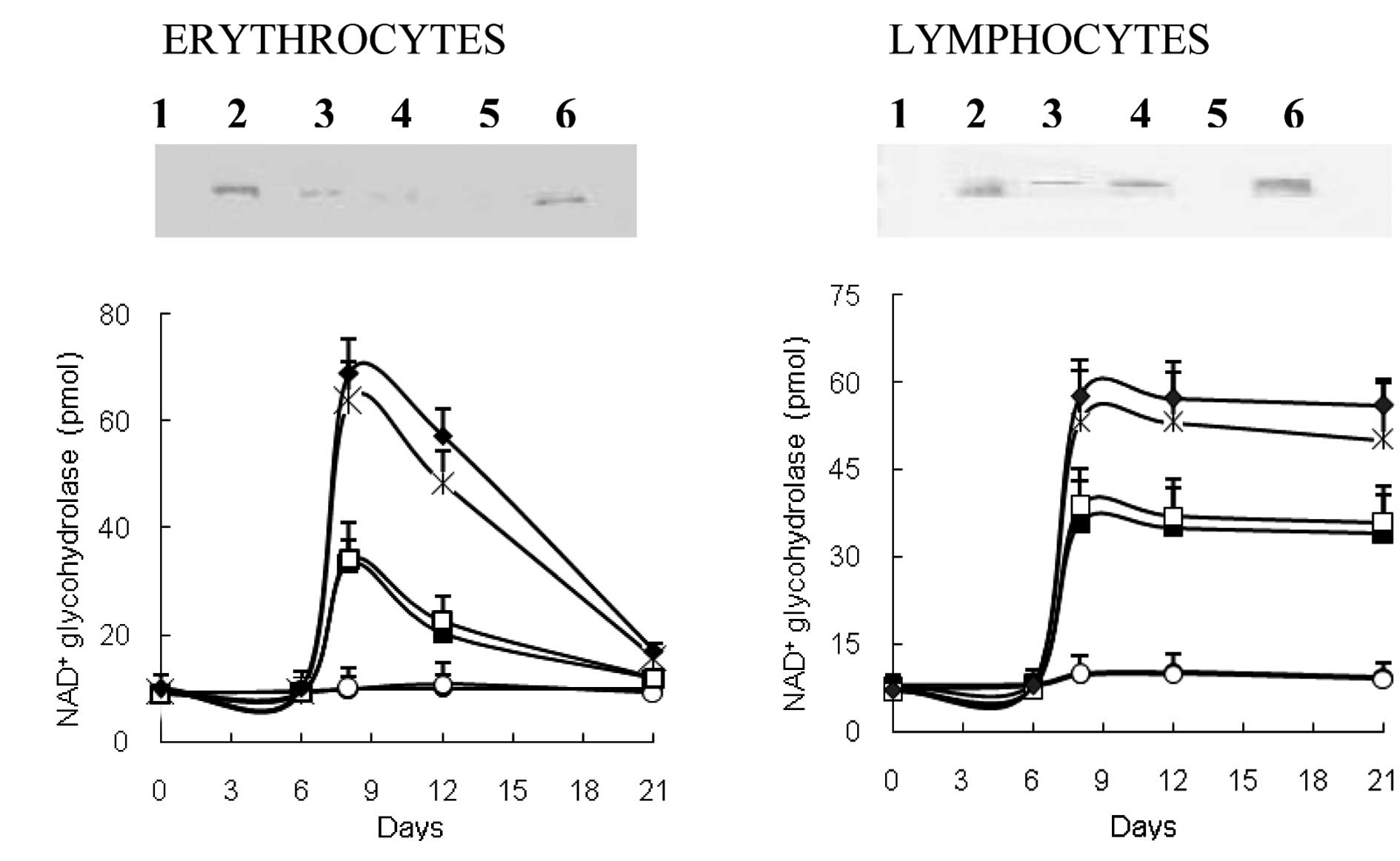
EATC supernatant alone is capable of
inducing CD38 expression
Of note is that the injection of EATC culture medium
propagated either with or without FCS supplement, i.e., EATC
supernatant with or without FCS supplement, also resulted in the
expression of CD38. In this case, the immunoreactive band, although
sharp and distinct, was not as prominent as the bands obtained
after injection of serum or ascites fluid samples. However, since
this was achieved with trace amounts of protein present in
serum-free culture medium, the finding indicated a high level of
inducing activity (Fig. 3A). The
increase in NAD+ glycohydrolase activity promoted by
EATC supernatants was also less than that obtained with serum
samples or ascites fluid, but significant in comparison to the
controls (Fig. 3B). Moreover, the
inducing effect on CD38 expression may be shown by flow cytometry
(Fig. 3C-J).
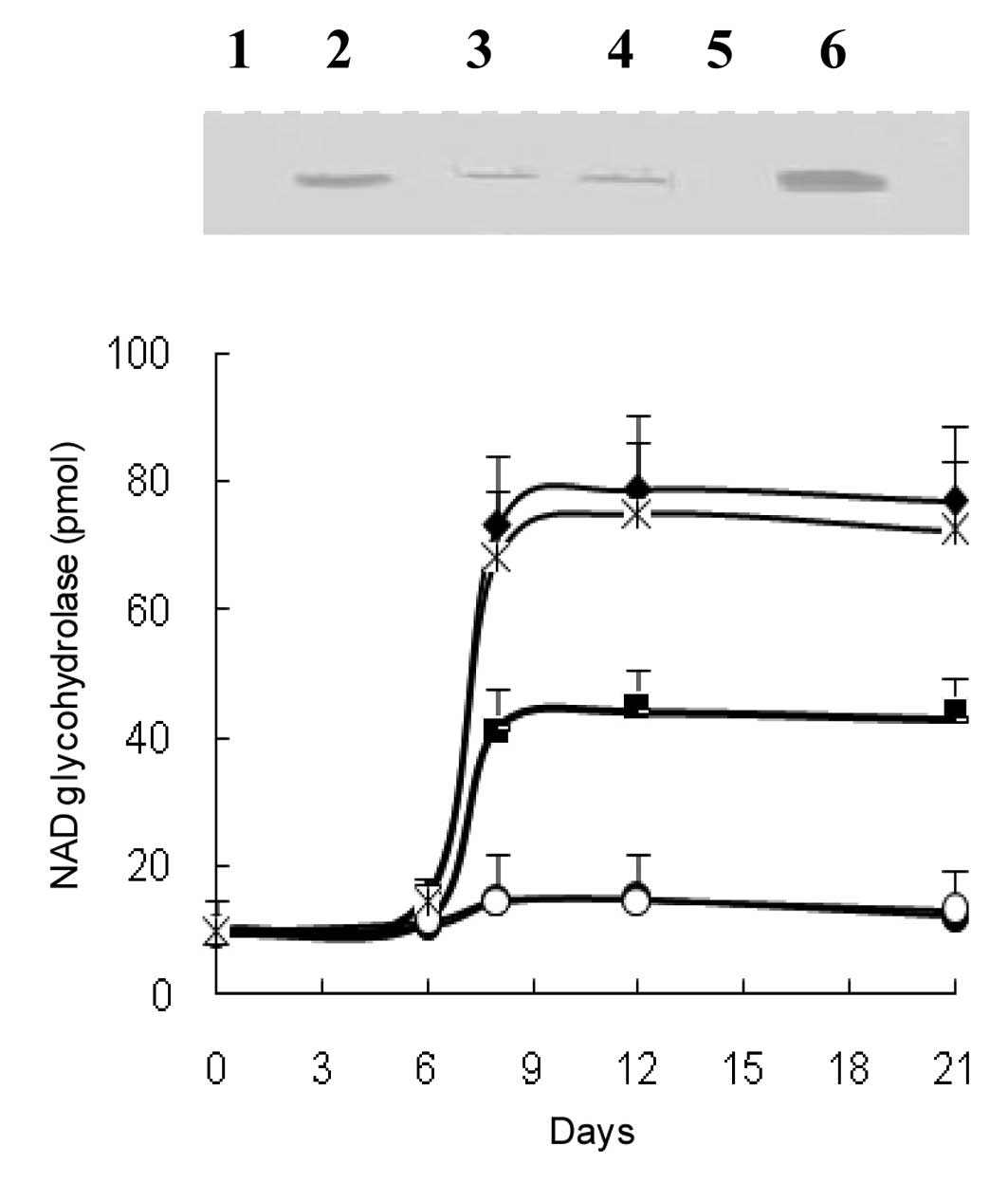
CD38 expression in peripheric mouse
lymphocytes in culture
Finally, CD38 expression was investigated in a
peripheric lymphocyte culture system. The findings obtained in
whole mice were reproduced in this system by the addition of serum
samples from mice with developed EAT, patients with high CEA
values, ascites fluid and, in particular, EATC (Fig. 4).
Discussion
Recent investigations have shown that considerable
changes in NAD+/ADP-ribose metabolism (17) and increases in CD38 expression and
related enzymatic activities (6,7) occur
concomitantly to neoplastic development. CD38 expression has proven
to be highly useful in clinics in monitoring HIV-1 infection and
recovery (8). Thus, with its
pivotal position in lymphocyte activation, CD38 expression appears
to be a sensitive and rather specific indicator of immune reaction
against variant forms of progressive disease, although its role in
this process remains to be clarified. CD38 appears to exert and/or
mediate pleiotropic effects through its multifaceted identity as an
enzyme, receptor and adhesion molecule.
This study utilized clinical blood samples
suggesting the use of an animal model for further and extended
studies, and addressed the issue of identifying the primary source
of the factors and/or mechanisms that control CD38 up-regulation.
The present investigation has been initiated with this
consideration in mind, and has used BALB/c mice to which EATC were
administered as the experimental model. The results obtained in
this conventional system appear to support and extend the data
obtained by analyzing blood samples from cancer patients. Thus,
increases in CD38 expression and CD38-associated enzymatic
activities found in cancer patients were successfully reproduced in
mice after the application of live Ehrlich cells, concomitantly
with the development of ascites tumors. The increases were also
achieved after injections of serum samples from mice with fully
developed ascites tumors, cancer patients with high CEA values and
ascites fluid. Moreover, the addition of these serum samples
resulted in similar increases in CD38 expression and
CD38-associated enzymatic activities in peripheric lymphocytes in
culture. Thus, CD38 induction was observed both in erythrocytes and
lymphocytes in whole animals as well as peripheric lymphocytes in
culture. This observation suggests that induction occurs in various
hematopoietic cell lineages and at different levels of
differentiation.
EATC supernatant exhibited a similar, albeit weaker
inducing effect on CD38 expression. This finding suggests that the
observed up-regulation of CD38 is partially triggered by factors
originating from the tumor. This effect is likely accompanied and
amplified by factors/cytokines produced by the host in response to
the tumor. Recent studies provide evidence for the involvement of
proinflammatory cytokines in tumor vs. host interactions in cancer
disease. Various neoplastic cell lines are known for the production
of such proinflammatory cytokines (18–20).
Thus, these cytokines are finding increasing use as markers in the
detection and prognostic assessment of cancer (21–29).
Moreover, cytokines have been shown to enhance CD38 expression in
different cell types (26).
The biological relevance of the increases in the
reported activities associated with CD38 induction as a response to
a proliferative process remains to be clarified. The product of
ADP-ribosyl cyclase activity, cyclic ADP-ribose, is a
Ca2+-mobilizing second messenger that plays a role in
cell differentiation and/or proliferation (27). On the other hand, the ligation of
CD38 by selected agonistic monoclonal antibodies is followed via
its receptorial properties by signals, resulting in proliferative
or apoptotic effects and cytokine induction in various members of
the immune system (28). CD38 is
also involved in the binding of lymphocytes to endothelial cells
via interaction with CD31, its counter-receptor. Finally, CD38
activates, upon ligation with agonistic monoclonal antibody or
interaction with CD31, pathways that lead to the secretion of
proinflammatory cytokines from human monocytes (28). Thus, CD38-induction possibly
triggered in the course of proliferative processes via certain
proinflammatory cytokines may be an event in which proliferative or
apoptotic processes are involved, but also with a positive feedback
effect on the tumor vs. host reactions.
CD38 expression has gained interest as a prognostic
marker in human chronic lymphocytic leukemia (CLL) following the
finding that CD38 expression is associated with the absence of
mutations in immunoglobulin variable (IgV) genes in CLL patients
(29). With an increasing
percentage of clonal cells expressing CD38, together with ZAP-70
activation as well as absence of mutations in IgV genes, the
prognosis of cases becomes poorer with a shorter survival span and
enhanced and repeated requirement for chemotherapy (5,30).
CD38 expression appears to reflect the involvement of a key element
in the pathogenetic network underlying the disease (5). Through its receptorial properties
and/or association with various signal pathways, CD38 protects
cells from apoptosis, thereby increasing their survival rate and
conferring them increased proliferative potential (5). Thus, CD38 expression appears to have
dual implications in the context of leukemia. Firstly, as a key
step in the activation of the immune system, it reflects the
reaction of the host on the ongoing proliferative process. On the
other hand, in the particular case of a cancer type originating
from the immune system itself, CD38 expression as a survival
mechanism for the cancer cell apparently contributes to its
proliferative proficiency and malignancy.
Acknowledgements
This study was supported by the Research Fund of the
Istanbul University (project G-147/20082003 and UDP-G/4128). The
authors thank Mr. Çağatay Korkut for his data which was useful in
the initial stage of the investigation and Mrs. Suzan Adın-Çınar
for her help in the flow cytometric analysis. The help of Mr. Halim
İşsever in statistical analysis is gratefully acknowledged.
Abbreviations:
|
BCIP
|
5-bromo-4-chloro-3′-indolyl phosphate
p-tolui-dine
|
|
BSA
|
bovine serum albumin
|
|
EATC
|
Ehrlich ascites tumor cell
|
|
EAT
|
Ehrlich ascites tumor
|
|
FITC
|
fluorescein isothiocynate
|
|
FCS
|
fetal calf serum
|
|
CEA
|
carcinoembryonic antigen
|
|
NBT
|
nitroblue tetrazolium chloride
|
|
NAD+
|
nicotinamide guanine dinucleotide
|
|
PBS
|
phosphate-buffered saline
|
|
SDS/PAGE
|
sodium dodecyl sulfate/polyacrylamide
gel electrophoresis
|
References
|
1
|
Friedman H and Rapoport SM: Enzymes of the
red cell: a critical catalogue. Cellular and Molecular Biology of
Erythrocytes. Yoshikawa H and Rapoport SM: London University Park
Press; London: pp. 181–259. 1974
|
|
2
|
Zocchi E, Franco L, Guida L, Benatti U,
Bargellesi A, Malavasi F, Lee HC and De Flora A: A single protein
immunologically identified as CD38 displays NAD+
glycohydrolase, ADP-ribosyl cyclase and cyclic ADP-ribose hydrolase
activities at the outer surface of human erythrocytes. Biochem
Biophys Res Commun. 196:1459–1465. 1993.PubMed/NCBI
|
|
3
|
Howard M, Grimaldi JC, Bazan JF, Lund FE,
Santos-Argumedo L, Parkhouse RM, Walseth TF and Lee HC: Formation
and hydrolysis of cyclic ADP-ribose catalyzed by lymphocyte antigen
CD38. Science. 262:1056–1059. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gelman L, Deterre P, Gouy H, Boumsell L,
Debre P and Bismuth G: The lymphocyte surface antigen CD38 acts as
a nicotinamide adenine dinucleotide glycohydrolase in human T
lymphocytes. Eur J Immunol. 23:3361–3364. 1993. View Article : Google Scholar
|
|
5
|
Deaglio S, Aydin S, Vaisitti T, Bergui L
and Malavasi F: CD38 at the junction between prognostic marker and
therapeutic target. Trends Mol Med. 14:210–218. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Albeniz I, Demir Ö, Nurten R and Bermek E:
NAD glycohydrolase activities and ADP-ribose uptake in erythrocytes
from normal subject and cancer patients. Biosci Rep. 24:41–53.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Albeniz I, Demir Ö, Türker-Şener L,
Yalçıntepe L, Nurten R and Bermek E: Erythrocyte CD38 as a
prognostic marker in cancer. Hematology. 12:409–414. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu Z, Cumberland WG, Hultin LE, Prince
HE, Detels R and Giorgi JV: Elevated CD38 antigen expression on
CD8+ T cells is a stronger marker for the risk of
chronic HIV disease progression to AIDS and death in the
multicenter AIDS cohort study than CD4+ cell count,
soluble immune activation markers or combinations of HLA-DR and
CD38 expression. J Acquir Immune Defic Syndr Hum Retrovirol.
16:83–92. 1997.PubMed/NCBI
|
|
9
|
Dodge JT, Mitchell C and Hanahan DJ: The
preparation and chemical characteristics of the hemoglobin free
ghosts of human erythrocytes. Arch Biochem Biophys. 100:119–130.
1963. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Böyum A: Separation of leukocytes from
blood and bone marrow. Scand J Clin Lab Invest Suppl. 97:7–106.
1968.
|
|
11
|
Liebert M, Ballou B, Taylor RJ, Reiland JM
and Hakala TR: A method of membrane preparation for immunoassay. J
Immunol Methods. 85:97–104. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Davis LG, Dibner MD and Battey JF: Basic
Methods in Molecular Biology. Elsevier Science Publishing Co Inc;
New York: 1986
|
|
13
|
Kim H, Jacobson EL and Jacobson MK:
Synthesis and degradation of cyclic ADP-ribose by NAD
glycohydrolase. Science. 261:1330–1333. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yalçıntepe L, Albeniz I, Adın-Çınar S,
Tiryaki D, Bermek E, Graeff RM and Lee HC: Nuclear CD38 in retinoic
acid-induced HL-60 cells. Exp Cell Res. 303:14–21. 2005.PubMed/NCBI
|
|
15
|
Laemmli UK: Cleavage of structural
proteins during the assembly of the head of bacteriophage T4.
Nature. 227:680–685. 1970. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gershoni JM and Palade GE: Protein
blotting: principles and applications. Anal Biochem. 131:1–15.
1983. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nurten R, Üstündağ I, Sayhan N and Bermek
E: ADP-ribosylation of human proteins promoted by endogenous NAD
glycohydrolase activity. Biochem Biophys Res Commun. 200:450–458.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Watson JM, Sensintaffar JL, Berek JS and
Martinez-Maza O: Constitutive production of interleukin 6 by
ovarian cancer cell lines and by primary ovarian tumor cultures.
Cancer Res. 50:6959–6965. 1990.PubMed/NCBI
|
|
19
|
Siegall CB, Schwab G, Nordan RP,
FitzGerald DJ and Pastan I: Expression of the interleukin 6
receptor and interleukin 6 in prostate carcinoma cells. Cancer Res.
50:7786–7788. 1990.PubMed/NCBI
|
|
20
|
Bendre MS, Margulies AG, Walser B, Akel
NS, Bhattacharrya S, Skinner RA, Swain F, Ramani V, Mohammad KS,
Wessner LL, Martinez A, Guise TA, Chirgwin JM, Gaddy D and Suva LJ:
Tumor-derived interleukin-8 stimulates osteolysis independent of
the receptor activator of nuclear factor-kappaB ligand pathway.
Cancer Res. 65:11001–11009. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Blay JY, Negrier S, Combaret V, Attali S,
Goillot E, Merrouche Y, Mercatello A, Ravault A, Tourani JM and
Moskovtchenko JF: Serum level of interleukin 6 as a prognosis
factor in metastatic renal cell carcinoma. Cancer Res.
15:3317–3322. 1992.PubMed/NCBI
|
|
22
|
Wittke F, Hoffmann R, Buer J, Dallmann I,
Oevermann K, Sel S, Wandert T, Ganser A and Atzpodien J:
Interleukin 10 (IL-10): an immunosuppressive factor and independent
predictor in patients with metastatic renal cell carcinoma. Br J
Cancer. 79:1182–1184. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Michalaki V, Syrigos K, Charles P and
Waxman J: Serum levels of IL-6 and TNF-alpha correlate with
clinicopathological features and patient survival in patients with
prostate cancer. Br J Cancer. 90:2312–2316. 2004.PubMed/NCBI
|
|
24
|
Zhang GJ and Adachi I: Serum interleukin-6
levels correlate to tumor progression and prognosis in metastatic
breast carcinoma. Anticancer Res. 19:1427–1432. 1999.PubMed/NCBI
|
|
25
|
Macrì A, Versaci A, Loddo S, Scuderi G,
Travagliante M, Trimarchi G, Teti D and Famulari C: Serum levels of
interleukin 1beta, interleukin 8 and tumour necrosis factor alpha
as markers of gastric cancer. Biomarkers. 11:184–193.
2006.PubMed/NCBI
|
|
26
|
Kang BN, Tirimurugaan KG, Deshpande DA,
Amrani Y, Panettieri RA, Walseth TF and Kannan MS: Transcriptional
regulation of CD38 expression by tumor necrosis factor-alpha in
human airway smooth cells: role of NF-κB and sensitivity to
glucocorticoids. FASEB J. 20:1000–1002. 2006.
|
|
27
|
Lee HC: Cyclic ADP-ribose: a new member of
a super family of signalling cyclic nucleotides. Cell Signal.
6:591–600. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lande R, Urbani F, Di Carlo B, Sconocchia
G, Deaglio S, Funaro A, Malavasi F and Ausiello CM: CD38 ligation
plays a direct role in the induction of IL-1beta, IL-6, and IL-10
secretion in resting human monocytes. Cell Immunol. 220:30–38.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Damle RN, Wasil T, Fais F, Ghiotto F,
Valetto A, Allen SL, Bunchbinder A, Budman D, D ittmar K, Kolitz J,
Lichtman SM, Schulman P, Vincinquerra VP, Rai KR, Ferrarini M and
Chiorazzi N: IgV gene mutation status and CD38 expression as novel
prognostic indicators in chronic lymphocytic leukemia. Blood.
94:1840–1847. 1999.PubMed/NCBI
|
|
30
|
Matrai Z: CD38 as a prognostic marker in
CLL. Hematology. 10:39–46. 2005. View Article : Google Scholar : PubMed/NCBI
|