Contents
Introduction
IRF-1 in human cancer
Tumor suppressive activity of IRF-1
Regulation of the cell cycle
Regulation of apoptosis
Regulation of immune response
Conclusion
Introduction
The original identification of the first member of
the interferon (IFN) regulatory factor (IRF) family, IRF-1, was
followed by additional studies on type I IFN and IFN-inducible
genes in the IFN system, which is referred to as ‘the IRF kingdom’.
The mammalian IRF family of transcription factors is comprised of 9
members: IRF-1, -2, -3, -4/PIP/LSIRF/ICSAT, IRF-5, -6, -7, -8/ICSBP
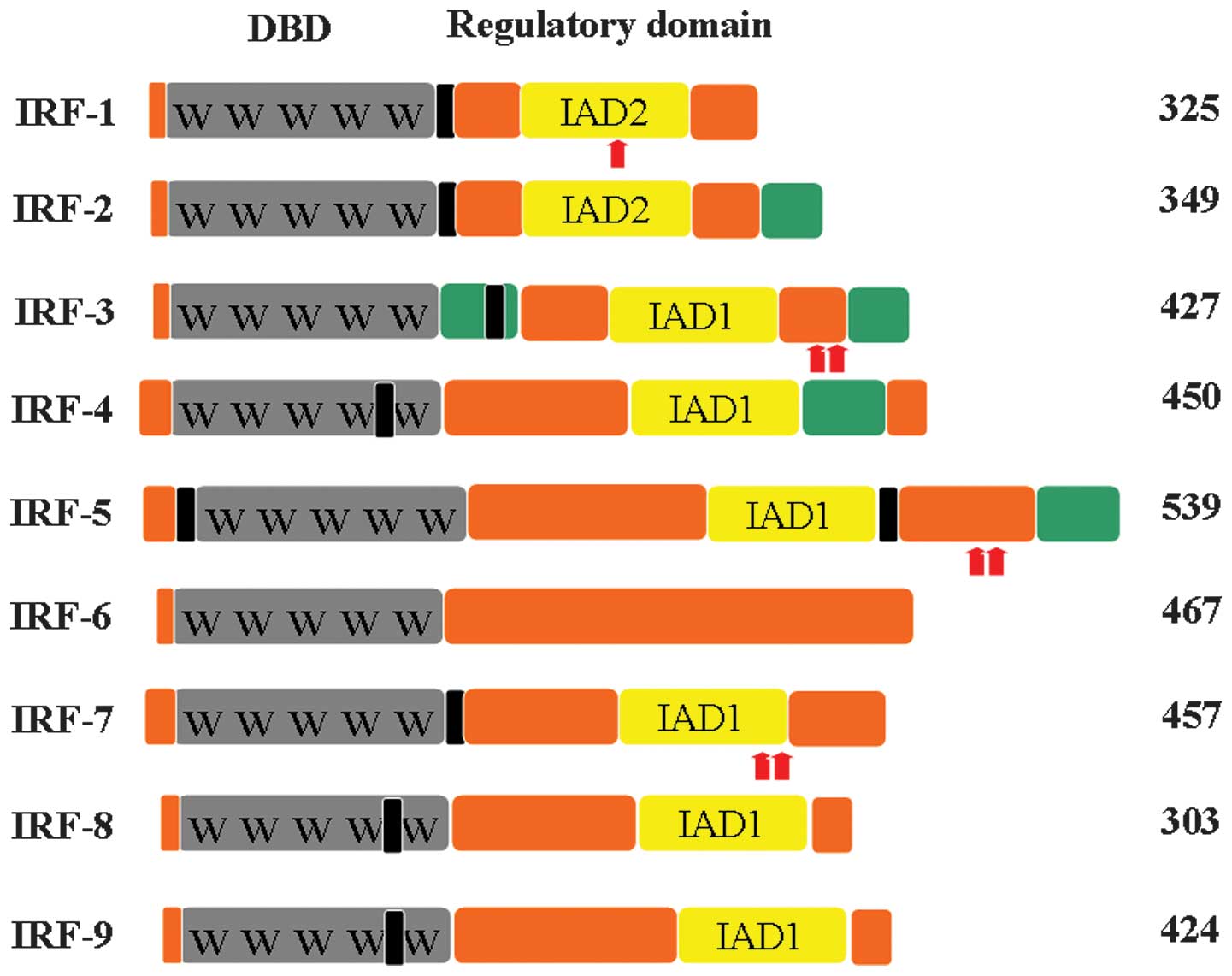
and -9 (1,2). IRFs contain an amino-terminal DNA
binding domain (DBD) which is characterized by a series of five
well-conserved tryptophan-rich repeats (3,4). The
DBD forms a helix-turn-helix domain and recognizes a DNA sequence
similar to the IFN-stimulated response element (A/GNGAAANNGAAACT)
(5). The carboxyl-terminal regions
of IRFs are less well-conserved and mediate interactions with
additional IRF members, transcription factors and cofactors,
conferring specific activities upon each IRF (Fig. 1) (5,6).
Although IRFs were first characterized as transcriptional
regulators of type I IFNs and IFN-inducible genes, more recent
studies indicate that this family is important for the regulation
of oncogenesis beyond the IFN system (6).
IRF-1 was the first member of the IRF family to be
isolated by virtue of its affinity to specific DNA sequences in the
IFN-β promoter (7). The molecule is
markedly induced by IFN, double-stranded RNA (dsRNA), cytokines and
specific hormones. Induced IRF-1 activates target genes leading to
inhibition of cell proliferation and stimulation of cell apoptosis
(5). IRF-2 binds to the same DNA
sequences as IRF-1 but downregulates or blocks the activation of
IRF-1 target genes. Accumulating evidence indicates that IRF-1 and
IRF-2 have antioncogenic and oncogenic potentials, respectively
(6). A correlation between IRF-4
and oncogenesis has also been reported in Epstein-Barr virus
(EBV)-transformed lymphocytes and HTLV-1-induced leukemogenesis
(8,9). Notably, IRF-4 alone is not sufficient
for oncognesis in transgenic mice overexpressing IRF-4 in
lymphocytes. IRF-4 may regulate cellular growth by targeting
pro-apoptotic IRF-5 during EBV transformation (10,11).
In addition, IRF-8 has been revealed to exhibit antitumor activity
through direct control of cell growth, differentiation and
apoptosis and modulation of antitumor immunity (12). IRF-6 may also act as a tumor
suppressor via its interaction with maspin, a tumor suppressor gene
(13). An additional member of the
IRF family, IRF-7, mediates the EBV transformation process in the
pathogenesis of EBV-associated lymphomas indicative of oncogenic
properties (14). Together, these
observations indicate that the IRF family is important for
regulation of oncogenesis.
Accumulating evidence indicates that deletion or
inactivation of the IRF-1 gene may be a critical step in the
development of specific types of human cancer. A previous study on
IRF-1 identified a tumor suppressive function demonstrating the
importance of additional investigation of IRF-1 in oncogenesis
regulation. In the present review, we summarize the contribution of
IRF-1 to regulation of oncogenesis. In addition, current hypotheses
with regard to the molecular mechanisms by which IRF-1 regulates
oncogenesis are discussed. IRF-1 may suppress tumor proliferation
by inhibiting the cell cycle, reducing susceptibility to
transformation by oncogenes or inducing apoptosis, development of
natural killer (NK) cells and differentiation of Th1 and
CD8+ T cells. Review of the current literature on IRF-1
is likely to improve understanding of the role of this molecule in
the pathological processes that lead to human malignant diseases
and may provide novel therapeutic strategies.
IRF-1 in human cancer
Previous clinical studies have indicated that loss
of IRF-1 may affect development of specific forms of human cancer.
This hypothesis was developed following the observation that the
IRF-1 gene maps to the chromosomal region 5q31.1 (15). Deletion of this region is one of the
most frequently observed cytogenetic abnormalities in leukemia or
preleukemic myelodysplastic syndrome (MDS) (16). Loss of one IRF-1 allele has also
been reported in esophageal and gastric cancers (17–19).
Loss of heterozygosity analysis at the IRF-1 locus of sporadic
breast cancer demonstrated frequent loss of heterozygosity at the
IRF-1 gene, which may induce low IRF-1 mRNA expression. These
observations imply a correlation between the IRF-1 gene and poor
clinical outcome in breast cancer (20). Notably, a polymorphism in the IRF-1
gene was detected at a higher frequency in human breast cancer cell
lines than in the general population and is more frequent in
African-American than Caucasian individuals of European-ancestry
(21). A significant difference in
genotype distribution between these populations was identified
(21). Collectively, these studies
indicate that genetic alterations of IRF-1 are important for the
development of specific types of human cancer.
In addition to genetic alterations of IRF-1
genotypes, several additional mechanisms by which IRF-1 may be
inactivated in cancer have been reported. For instance,
nucleophosmin, a putative ribosome assembly factor commonly
overexpressed in myeloid leukaemia cells, binds to and inhibits the
function of IRF-1 (22,23). Specific splicing aberrations of the
IRF-1 gene leads to loss of functional IRF-1 in MDS and leukemias
(24,25). A mechanism associated with the human
papilloma virus (HPV) 16-encoded E7 oncoprotein has also been
reported (26). In addition,
previous studies indicate that SUMOylated IRF-1 inhibits apoptosis
by repression of transcriptional activity (27). Finally, numerous reports reveal low
expression levels of IRF-1 mRNA in specific forms of cancer,
including breast cancer and hepatocellular carinoma (28,29).
To develop an improved understanding of the role of IRF-1 in human
cancer, additional studies must be performed to clarify the
mechanism by which IRF-1 regulates oncogenesis.
Tumor suppressive activity of IRF-1
The tumor suppressor-like activity of IRF-1 was
previously demonstrated by oncogenic transformation assays in which
the activated c-Ha-Ras, a single oncogene, was introduced to and
transformed IRF−/− MEF cells (30). Wild-type MEF cells are known to
require at least two oncogenes to undergo transformation. In
addition, conditions under which activated Ras paradoxically
inhibits cell growth of myeloid cells have been demonstrated to be
involved with IRF-1 and the induction of
p21WAF1/CIP1(31). A
previous study revealed that ectopic expression of IRF-1 suppresses
the malignant properties of cancer cell lines and
oncogene-transformed cell lines in vitro and in
vivo(29).
In contrast to other tumor suppressor genes, loss of
IRF-1 alleles alone rarely induces tumor development, however,
IRF-1 deficiency was previously demonstrated to markedly enhance
tumor predispositions caused by the expression of a c-Ha-Ras
transgene or nullizygosity of the transformation-related protein 53
(Trp53) gene (32). This
accelerated tumor development may not be due to the discussed
immunological disorders caused by the absence of IRF-1 (33). Therefore, IRF-1 belongs to a class
of tumor susceptibility genes whose loss in combination with other
genetic alterations significantly increases the incidence of
developing tumors.
The mechanisms by which IRF-1 mediates tumor
suppression are not well understood. Several IRF-1 induced genes
that exert growth-inhibitory and promote apoptosis effects,
including 2′,5′-oligo(A) synthetase (34), indoleamine 2,3-dioxigenase (35), RNA-dependent protein kinase (PKR)
(36),
p21WAF/CIP1(37), Lox
(38), angiotensin type II receptor
(39), TNF-related
apoptosis-inducing ligand (TRAIL) (40), caspase-1 (41), -7 (42) and -8 (43) may be associated with inhibition of
proliferation, induction of apoptosis, stimulation of the immune
response and reversion of the transformed phenotype (Table I). Among them, Lox is important for
biogenesis of connective tissue matrices and is identical to the
Ras recision gene, which is associated with in the reversion of
Ras-transformed NIH3T3 cells by preventing the activation of
nuclear factor κ-light-chain-enhancer of activated B (44). Aberrant expression of Lox leads to
tumorigenesis and tumor progression. However, this molecule is also
a potential downstream mediator for the tumor-suppressive activity
of IRF-1. By contrast, an additional study demonstrated that growth
of the Ras+ myc-induced transforming phenotype in soft
agar is not altered by overexpression of Lox, but is suppressed by
IRF-1 expression (45).
 | Table I.IRF-1 target genes. |
Table I.
IRF-1 target genes.
| Gene | Role | Reference |
|---|
| IFN-α/β | Antiviral
response | 5,7 |
|
p21WAF/CIP1 | Cell cycle | 38 |
| p53 | Apoptosis | 21 |
| PKR | Cell cycle | 37 |
| 2′,5′-OAS | Antiviral
response | 53 |
| Lox | Inhibition of cell
transformation | 39 |
| Angiotensin type II
receptor | Apoptosis | 40 |
| TRAIL | Apoptosis | 41 |
| Caspase-1 | Apoptosis | 42 |
| Caspase-7 | Apoptosis | 43 |
| Caspase-8 | Apoptosis | 44, 69 |
| PUMA | Apoptosis | 64 |
| BRIP1 | N/A | 49 |
| BARD1 | N/A | 49 |
| HPGD | Metabolic
process | 49 |
| PLAGL1 |
Anti-proliferation | 49 |
| RASSF5 | Cell adhesion | 49 |
| AKAP12 | Signal
transduction | 49 |
| IL-15 | NK cell
development | 76 |
| iNOS | Th1
differentiation | 73 |
| LMP-2 | CD8+ T
cell development | 80 |
| TAP-1 | CD8+ T
cell development | 79 |
| MHC-I | CD8+ T
cell development | 78 |
| IL-12p40 | Th1
differentiation | 74 |
More recent studies have identified additional IRF-1
target genes. The tumor suppressor activity of IRF-1 has been
associated with downregulation of cyclin D1 (46) and survivin (47). A ChIP-chip approach performed by
Frontini et al(48) revealed
a novel role for IRF-1 in the regulation of the DNA interstrand
crosslink damage response. IRF-1 regulates ATP-dependent RNA
helicase (BRIP1), a component of the Fanconi anemia/BRCA DNA repair
pathway and a newly identified breast cancer susceptibility gene
(49,50). The association between IRF-1 and
BRIP1 further validates the importance of IRF-1 as a tumor
susceptibility gene (48). In
addition, hydroxyprostaglandin, pleiomorphic adenoma gene-like 1,
Ras association domain family 5, a kinase (PRKA) anchor protein 12
and deleted in colorectal cancer have been identified as IRF-1
target genes with tumor suppressor activities (48). Further studies to identify the role
of these individual genes in the antitumor activity of IRF-1 must
be performed.
Regulation of the cell cycle
The involvement of IRF-1 in cell cycle regulation
has been studied extensively and its tumor suppressive activity may
be explained, at least in part, by its cell cycle checkpoint
function. In fact, the expression of IRF-1 appears to be regulated
throughout the cell cycle. IRF-1 mRNA expression is markedly
elevated in NIH3T3 cells subjected to serum-induced cell cycle
progression; however, it rapidly decreases as the serum-induced
cell cycle continues, suggesting that IRF-1 is involved in cell
cycle regulation (51). Several
studies have provided insight into the mechanism by which IRF-1
inhibits the cell cycle.
The growth inhibitory effects of IRF-1 may be
mediated by stimulation of antiproliferative gene transcription. As
with p53, transcriptional induction of the gene encoding
p21WAF1/CIP1 by γ irradiation has also been identified
to be regulated by IRF-1 (37). In
DNA-damaged cells, IRF-1 protein levels were elevated via an Ataxia
telangiectasia mutated-dependent increase in mRNA expression and
protein half-life, acting on the p21WAF1/CIP1 promoter
region containing the IRF-1 and p53-binding sites to induce a
G1 cell cycle-specific arrest (37).
Moreover, additional genes or secreted factors
induced by IRF-1 may also cause cell growth inhibition, including
2′,5′-OAS, whose products activate the mRNA-degrading enzyme RNase
L (34). Xie et al
established a casual series of events that functionally connect the
antiproliferative effects of IFNs with the IRF-1-dependent
suppression of the CDK2 gene, which encodes a key regulator of the
G1/S phase transition. Although IRF-1, -2, -3 and -7
have all been demonstrated to activate IRF-1-responsive reporter
genes, only IRF-1 inhibits CDK2 gene transcription (52).
The IRF-1-induced enzymes, including lysyl oxidase
and indoleamine 2,3-dioxygenase, may lower the biosynthetic
capacity of the cell by enhanced degradation of rate-limiting
precursors (35,38). PKR is important for the regulation
of cell proliferation and exerts antigrowth activities by
IFN-inducible genes, including IRF-1 (53).
Specific signal pathways are also vital for the
regulation of growth activity. For instance, the Janus kinase and
signal transducer and activator of transcription (JAK-STAT) pathway
may be an IRF-1 target for growth regulation at the transcriptional
level (54). However, STAT1 is
known to function upstream of IRF-1 and regulate IRF-1 promoter
expression. This mechanism is currently hypothesized to involve
IRF-1 upregulation in response to IFN induction through STAT1.
Newly synthesized IRF-1 may in turn activate expression of STAT1,
resulting in positive feedback regulation of IRF-1 expression
(55).
Regulation of apoptosis
Apoptosis is an additional mechanism used to control
cell number in tissues and eliminate individual cells that threaten
the host’s survival. IRF-1 is associated with apoptosis induced by
DNA damage or other stimuli (56).
Wild-type MEFs, when introduced with an activated oncogene, i.e.,
c-Ha-Ras, undergo apoptosis instead of cell cycle arrest when
treated with anti-cancer drugs or ionizing radiation. Apoptosis is
a hallmark of tumor suppression and is dependent, in this case, on
IRF-1 and p53 (30).
However, DNA damage-induced apoptosis in
mitogenically activated mature T lymphocytes is dependent on IRF-1
but independent of p53 (57,58).
Bowie et al demonstrated that IRF-1 is critical for the
promotion of p53-independent apoptosis in acutely damaged
basal-type human mammary epithelial cells, providing evidence that
loss of IRF-1 is a short-term marker of early basal-type breast
cancer risk (59). Pizzoferrato
et al identified that ectopic expression of IRF-1 protein
results in downregulation of survivin protein expression that is
independent of p53 and promotes breast cancer cell death (47). In addition, IRF-1 binds to distinct
sites in the promoter and upregulates expression of PUMA, a
p53-upregulated modulator of apoptosis that activates apoptosis by
the intrinsic pathway. PUMA has also been identified to function in
a p53-independent manner (60).
Therefore, IRF-1 induces apoptosis by the intrinsic pathway,
independent of the extrinsic pathway, by upregulation of PUMA.
However, in thymocytes, apoptosis is dependent on p53 but not on
IRF-1. Thus, IRF-1 and p53 regulate DNA damage-induced apoptosis
cooperatively and independently, depending on the type and
differentiation stage of the cell. Notably, gatekeeper of apoptosis
activating proteins-1, a transcriptional activator of IRF-1 and
p53, has a proapoptotic activity (61).
Caspases are unique proteases that comprise an
activation cascade downstream in the apoptosis mechanism. IRF-1 has
been demonstrated to directly mediate IFN-γ-induced apoptosis via
activation of caspase-1 gene expression in IFN-γ-sensitive ovarian
cancer cells and other cancer cells (62). Furthermore, IRF-1 is known to
activate caspase-8 expression in response to IFN-γ/STAT1 signaling,
a component of the events that sensitize cells for apoptosis
(63). Caspase activity assays are
used to determine the overexpression of wild-type IRF-1 or dominant
negative IRF-1 in breast cancer cells. Thus, IRF-1 controls
apoptosis through caspase-8 in breast cancer cells. These
observations are consistent with the hypothesis that IRF-1
regulates apoptosis through caspase-8 in breast cancer cells
(64). Moreover, RNA interference
experiments also indicated that IRF-1 and -2 are associated with
constitutive caspase-8 expression in neuroblastoma cells (65). In addition, Tomita et al
demonstrated that IRF-1 is important for IFN-γ mediated-enhancement
of Fas/CD95-mediated apoptosis through the regulation of
DEVD-CHO-sensitive caspases, most likely caspase-7 (66).
Moreover, specific genes or signaling molecules may
be involved in IRF-1-regulated apoptosis. TRAIL signaling is
critically involved in immune surveillance against tumor
development. IFN-γ enhances the anticancer activities of TRAIL
through IRF-1 (67). Promoter
mapping, chromatin immunoprecipitation and RNA interference reveal
that retinoid-induced IRF-1 is required for TRAIL induction by
retinoic acid (RA) and IFN-γ (40).
Regulation of immune response
IRF-1 regulates the expression of a number of genes
whose products are central to innate and adaptive immunity,
indicating that IRF-1 may provide a link between the two systems.
IRF-1 induces transcription of various genes involved in the first
reaction to viral invasion, including PKR and 2′,5′-OAS (36,34).
IRF-1 also binds MyD88 and is modified by currently unidentified
signaling molecule(s) to migrate into the nucleus and induce genes
encoding IFN-β, iNOS and IL-12p35 (68).
In addition to the functions assigned to IRF-1 in
differentiated immune cells, previous studies have revealed roles
for IRF-1 in the development of various immune cells. IRF-1 affects
the development and function of NK cells. Analysis of the spleen
and liver of Irf-1−/− mice demonstrates a reduction in
NK cell counts and function (69).
Reduced IRF-1 selectively affects bone marrow stromal cells that
constitute the microenvironment for NK cell development. IRF-1 does
not affect NK progenitors (70).
However, IRF-1 in stromal cells is required for transcription of
the gene encoding IL-15, which is essential for NK cell development
(71).
IRF-1 also regulates dendritic cell subset
development. Mice lacking IRF-1 have reduced numbers of mature
CD8+ T cells, despite normal maturation of
CD4+ T cells in the thymus and peripheral lymphoid
organs (72,73). Low molecular weight protein-2,
antigen processing-1 and major histocompatibility complex I are
decreased in Irf-1−/− thymic stromal cells (73–75),
however, the defect in CD8+ T cell development does not
reside in the thymic environment but is instead due to a
thymocyte-intrinsic defect during differentiation from immature to
mature CD8+ T cells (76). TCR stimulation induces IRF-1
expression in immature thymocytes, while Irf-1−/−
thymocytes are defective in TCR-mediated signal transduction.
Therefore, IRF-1 may regulate genes in developing T cells that are
crucial for signal transduction in the thymus and in
lineage-specific differentiation of CD8+ T cells
(76).
Moreover, IRF-1 promotes the differentiation of Th1.
T-cells from Irf-1−/− mice fail to mount Th1 responses
and instead exclusively undergo Th2 differentiation in
vitro(77). IRF-1 has also been
identified to be involved in the differentiation of Th1 combined
with HPV E7 (78). However,
compromised Th1 differentiation is associated with defects in
multiple cell types, including impaired production of the p40
subunit of interleukin-12 (IL-12p40) by macrophages and
hyporesponsiveness of CD4+ T cells to IL-12, which is essential for
Th1 differentiation (69,77). IRF-1 regulates the expression of
genes encoding iNOS induced by IFN-γ. iNOS catalyzes the production
of nitric oxide, a short-lived volatile gas important in the
effector phase of the Th1 response (79). In essence, IRF-1 is indispensable
for the differentiation of Th1.
The discussed observations demonstrate that tumor
surveillance by the immune system is impaired by loss of IRF-1.
Therefore, IRF-1 may function as a ‘systemic gatekeeper’ involved
in the protection of the host against exogenous mutagens that may
lead to caricinogenesis.
Conclusion
The previous studies discussed reveal that the IRF-1
transcription factor is involved in the regulation of tumor
suppression and oncogenesis. However, the mechanisms underlying
IRF-1-mediated tumor suppression and oncogenesis remain
undefined.
The high frequency of mutated or rearranged IRF-1 in
several cancer types indicates its importance in oncogenesis.
Additional studies demonstrate that IRF-1 exhibits tumor suppressor
activities in a number of human tumors. IRF-1 domains and
post-translational modifications may affect the function of the
transcription factor. IRF-1 activates a set of target genes
responsible for inhibition of the cell cycle and induction of
apoptosis and may regulate DNA repair. Therefore, in order to gain
an improved understanding of the role of IRF-1 in oncogenesis, new
targets of IRF-1 must be identified which are critical in the
cascade of events involved in oncogenesis.
IRF-1 is a unique member of the IRF transcription
factor family as it functions in the regulation of the innate and
adaptive immune systems, linking the immune response and
oncogenesis together. Elucidation of the intricate gene network
operation associated with regulation of host defense and the
function of IRF-1 in this network is not currently understood
fully. However, the multiple functions of IRF-1 imply that IRF-1
may be suitable as a selective gene with an engineered delivery
system for biotherapies of various types of cancer and autoimmune
diseases.
Acknowledgements
The present review was supported by
the Natural Science Fund for Colleges and Universities in Jiangsu
Province (no. 10KJB320023).
References
|
1.
|
Mamane Y, Heylbroeck C, Genin P, Algarte
M, Servant MJ, LePage C, DeLuca C, Kwon H, Lin R and Hiscott J:
Interferon regulatory factors: the next generation. Gene. 237:1–14.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Taniguchi T, Ogasawara K, Takaoka A and
Tanaka N: IRF family of transcription factors as regulators of host
defense. Annu Rev Immunol. 19:623–655. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Wathelet MG, Lin CH, Parekh BS, Ronco LV,
Howley PM and Maniatis T: Virus infection induces the assembly of
coordinately activated transcription factors on the IFN-beta
enhancer in vivo. Mol Cell. 1:507–518. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Meraro D, Hashmueli S, Koren B, Azriel A,
Oumard A, Kirchhoff S, Hauser H, Nagulapalli S, Atchison ML and
Levi BZ: Protein-protein and DNA-protein interactions affect the
activity of lymphoid-specific IFN regulatory factors. J Immunol.
163:6468–6478. 1999.PubMed/NCBI
|
|
5.
|
Lohoff M and Mak TW: Roles of
interferon-regulatory factors in T-helper-cell differentiation. Nat
Rev Immunol. 5:125–135. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Nobuyuki T and Tadatsugu T: The interferon
regulatory factors and oncogenesis. Semin Cancer Biol. 10:73–81.
2000. View Article : Google Scholar
|
|
7.
|
Fujita T, Sakakibara J, Sudo Y, Miyamoto
M, Kimura Y and Taniguchi T: Evidence for a nuclear factor(s),
IRF-1, mediating induction and silencing properties to human
IFN-beta gene regulatory elements. EMBO J. 7:3397–3405. 1988.
|
|
8.
|
Mamane Y, Grandvaux N, Hernandez E, Sharma
S and Innocente SA: Repression of IRF-4 target genes in human T
cell leukemia virus-1 infection. Oncogene. 21:6751–6765. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Xu DS, Zhao LJ, Valle LD, Miklossy J and
Zhang LW: Interferon regulatory factor 4 is involved in
Epstein-Barr Virus-mediated transformation of human B lymphocytes.
J Virol. 82:6251–6258. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Xu D, Meyer F, Ehlers E, Blasnitz L and
Zhang L: Interferon regulatory factor 4 (IRF-4) targets IRF-5 to
regulate Epstein-Barr virus transformation. J Biol Chem.
286:18261–18267. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Barnes BJ, Kellum MJ, Pinder KE, Frisancho
JA and Pitha PM: Interferon regulatory factor 5, a novel mediator
of cell cycle arrest and cell death. Cancer Res. 63:6424–6431.
2003.PubMed/NCBI
|
|
12.
|
Deng M and Daley GQ: Expression of
interferon consensus sequence binding protein induces potent
immunity against BCR/ABL-induced leukemia. Blood. 97:3491–3497.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Bailey CM, Khalkhali-Ellis Z, Kondo S,
Margaryan NV, Seftor RE, Wheaton WW, Amir S, Pins MR, Schutte BC
and Hendrix MJ: Mammary serine protease inhibitor (Maspin) binds
directly to interferon regulatory factor 6: identification of a
novel serpin partnership. J Biol Chem. 280:34210–34217. 2005.
View Article : Google Scholar
|
|
14.
|
Zhao Y, Xu DS, Jiang YJ and Zhang LW: Dual
functions of interferon regulatory factors 7C in Epstein-Barr
virus-mediated transformation of human B lymphocytes. PLoS One.
5:e94592010. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Itoh S, Harada H, Nakamura Y, White R and
Taniguchi T: Assignment of the human interferon regulatory factor-1
(IRF1) gene to chromosome 5q23-q31. Genomics. 10:1097–1099. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Willman CL, Sever CE, Pallavicini MG,
Harada H, Tanaka N, Slovak ML, Yamamoto H, Harada K, Meeker TC and
List AF: Deletion of IRF-1, mapping to chromosome 5q31.1, in human
leukemia and preleukemic myelodysplasia. Science. 259:968–971.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Ogasawara S, Tamura G, Maesawa C, Suzuki
Y, Ishida K, Satoh N, Uesugi N, Saito K and Satodate R: Common
deleted region on the long arm of chromosome 5 in esophageal
carcinoma. Gastroenterology. 110:52–57. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Tamura G, Ogasawara S, Nishizuka S, Sakata
K, Maesawa C, Suzuki Y, Terashima M, Saito K and Satodate R: Two
distinct regions of deletion on the long arm of chromosome 5 in
differentiated adenocarcinomas of the stomach. Cancer Res.
56:612–615. 1996.PubMed/NCBI
|
|
19.
|
Nozawa H, Oda E, Ueda S, Tamura G, Maesawa
C, Muto T, Taniguchi T and Tanaka N: Functionally inactivating
point mutation in the tumor-suppressor IRF-1 gene identified in
human gastric cancer. Int J Cancer. 77:522–527. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Cavalli LR, Riggins RB, Wang A, Clarke R
and Haddad BR: Frequent loss of heterozygosity at the interferon
regulatory factor-1 gene locus in breast cancer. Breast Cancer Res
Treat. 121:227–231. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Bouker KB, Skaar TC, Harburger DS, Riggins
RB, Fernandez DR, Zwart A and Clarke R: The A4396G polymorphism in
interferon regulatory factor 1 is frequently expressed in breast
cancer cell lines. Cancer Genet Cytogenet. 175:61–64. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Falini B, Mecucci C, Tiacci E, Alcalay M,
Rosati R, Pasqualucci L, Starza R, Diverio D, Colombo E, Santucci
A, Bigerna B, Pacini R, Pucciarini A, Liso A, Vignetti M, Fazi P,
Meani N, Pettirossi V, Saglio G, Mandelli F, Lo-Coco F, Pelicci PG
and Martelli MF: Cytoplasmic nucleophosmin in acute myelogenous
leukemia with a normal karyotype. N Engl J Med. 352:254–266. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Kondo T, Minamino N, Nagamura-Inoue T,
Matsumoto M, Taniguchi T and Tanaka N: Identification and
characterization of nucleophosmin/B23/numatrin which binds the
anti-oncogenic transcription factor IRF-1 and manifests oncogenic
activity. Oncogene. 15:1275–1281. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Harada H, Kondo T, Ogawa S, Tamura T,
Kitagawa M, Tanaka N, Lamphier MS, Hirai H and Taniguchi T:
Accelerated exon skipping of IRF-1 mRNA in human
myelodysplasia/leukemia; a possible mechanism of tumor suppressor
inactivation. Oncogene. 9:3313–3320. 1994.PubMed/NCBI
|
|
25.
|
Tzoanopoulos D, Speletas M, Arvanitidis K,
Veiopoulou C, Kyriaki S, Thyphronitis G, Sideras P, Kartalis G and
Ritis K: Low expression of interferon regulatory factor-1 and
identification of novel exons skipping in patients with chronic
myeloid leukaemia. Br J Haematol. 119:46–53. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Park JS, Kim EJ, Kwon HJ, Hwang ES,
Namkoong SE and Um SJ: Inactivation of interferon regulatory
factor-1 tumor suppressor protein by HPV E7 oncoprotein.
Implication for the E7-mediated immune evasion mechanism in
cervical carcinogenesis. J Biol Chem. 275:6764–6769. 2000.
View Article : Google Scholar
|
|
27.
|
Junsoo P, Kim K, Lee EJ, Seo YJ, Lim SN,
Park K, Rho SB, Lee SH and Lee JH: Elevated level of SUMOylated
IRF-1 in tumor cells interferes with IRF-1-mediated apoptosis. Proc
Natl Acad Sci USA. 104:17028–17033. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Tamura T, Yanai H, Savitsky D and
Taniguchi T: The IRF family transcription factors in immunity and
oncogenesis. Annu Rev Immunol. 26:535–584. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Takaoka A, Tamura T and Taniguchi T:
Interferon regulatory factor family of transcription factors and
regulation of oncogenesis. Cancer Sci. 99:467–478. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Tanaka N, Ishihara M, Kitagawa M, Harada
H, Kimura T, Matsuyama T, Lamphier MS, Aizawa S, Mak TW and
Taniguchi T: Cellular commitment to oncogene-induced transformation
or apoptosis is dependent on the transcription factor IRF-1. Cell.
77:829–839. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Passioura T, Dolnikov A, Shen S and
Symonds G: N-ras induced growth suppression of myeloid cells is
mediated by IRF-1. Cancer Res. 65:797–804. 2005.PubMed/NCBI
|
|
32.
|
Nozawa H, Oda E, Nakao K, Ishihara M, Ueda
S, Yokochi T, Ogasawara K, Nakatsuru Y, Shimizu S, Ohira Y, Hioki
K, Aizawa S, Ishikawa T, Katsuki M, Muto T, Taniguchi T and Tanaka
N: Loss of transcription factor IRF-1 affects tumor susceptibility
in mice carrying the Ha-ras transgene or nullizygosity for p53.
Genes Dev. 13:1240–1245. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Romeo G, Fiorucci G, Chiantore MV,
Percario ZA, Vannucchi S and Affabris E: IRF-1 as a negative
regulator of cell proliferation. J Interferon Cytokine Res.
22:39–47. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Benech P, Vigneron M, Peretz D, Revel M
and Chebath J: Interferon-responsive regulatory elements in the
promoter of the human 2′,5′-oligo(A)synthetase gene. Mol Cell Biol.
7:4498–4504. 1987.
|
|
35.
|
Takikawa O, Kuroiwa T, Yamazaki F and Kido
R: Mechanism of interferon-γ action. Characterization of
indoleamine 2,3-dioxygenase in cultured human cells induced by
interferon-γ and evaluation of the enzyme-mediated tryptophan
degradation in its anticellular activity. J Biol Chem.
263:2041–2048. 1988.
|
|
36.
|
Kirchhoff S, Koromilas AE, Schaper F,
Grashoff M, Sonenberg N and Hauser H: IRF-1 induced cell growth
inhibition and interferon induction requires the activity of the
protein kinase PKR. Oncogene. 11:439–445. 1995.PubMed/NCBI
|
|
37.
|
Pamment J, Ramsay E, Kelleher M, Dornan D
and Ball KL: Regulation of the IRF-1 tumour modifier during the
response to genotoxic stress involves an ATM-dependent signalling
pathway. Oncogene. 21:7776–7785. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38.
|
Tan RS, Taniguchi T and Harada H:
Identification of the lysyl oxidase gene as target of the
antioncogenic transcription factor, IRF-1 and its possible role in
tumor suppression. Cancer Res. 56:2417–2421. 1996.PubMed/NCBI
|
|
39.
|
Horiuchi M, Yamada T, Hayashida W and Dzau
VJ: Interferon regulatory factor-1 up-regulates angiotensin II type
2 receptor and induces apoptosis. J Biol Chem. 272:1952–1958. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
40.
|
Clarke N, Jimenez AM, Voltz E and
Gronemeyer H: Tumor suppressor IRF-1 mediates retinoid and
interferon anticancer signaling to death ligand TRAIL. EMBO J.
23:3051–3060. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41.
|
Chin YE, Kitagawa M, Kuida K, Flavell RA
and Fu XY: Activation of the STAT signaling pathway can cause
expression of caspase 1 and apoptosis. Mol Cell Biol. 17:5328–5337.
1997.PubMed/NCBI
|
|
42.
|
Sancéau J, Hiscott J, Delattre O and
Wietzerbin J: IFN-beta induces serine phosphorylation of Stat-1 in
Ewing’s sarcoma cells and mediates apoptosis via induction of IRF-1
and activation of caspase-7. Oncogene. 19:3372–3383.
2000.PubMed/NCBI
|
|
43.
|
Ruiz C, Ruiz AC, Rodriguez A, Ortiz G,
Redondo JM and López A: The up-regulation of human caspase-8 by
interferon-gamma in breast tumor cells requires the induction and
action of the transcription factor interferon regulatory factor-1.
J Biol Chem. 279:19712–19720. 2004. View Article : Google Scholar
|
|
44.
|
Jeay S, Pianetti S, Kagan HM and
Sonenshein GE: Lysyl oxidase inhibits ras-mediated transformation
by preventing activation of NF-κB. Mol Cell Biol. 23:2251–2263.
2003.PubMed/NCBI
|
|
45.
|
Kroger A, Dallugge A, Kirchhoff S and
Hauser H: IRF-1 reverts the transformed phenotype of oncogenically
transformed cells in vitro and in vivo. Oncogene. 22:1045–1056.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
46.
|
Kroger A, Stirnweiss A, Pulverer JE,
Klages, Grashoff M, Reimann J and Hauser H: Tumor suppression by
IFN regulatory factor-1 is mediated by transcriptional
down-regulation of cyclin D1. Cancer Res. 67:2972–2981. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
47.
|
Pizzoferrato E, Liu Y, Gambotto A,
Armstrong MJ, Stang MT, Gooding WE, Alber SM, Shand SH, Watkins SC
and Storkus WJ: Ectopic expression of interferon regulatory
factor-1 promotes human breast cancer cell death and results in
reduced expression of survivin. Cancer Res. 64:8381–8388. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
48.
|
Frontini M, Vijayakumar M, Garvin A and
Clarke N: A ChIP-chip approach reveals a novel role for
transcription factor IRF1 in the DNA damage response. Nucleic Acids
Res. 37:1073–1085. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
49.
|
Walsh T and King MC: Ten genes for
inherited breast cancer. Cancer Cell. 11:103–105. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
50.
|
Seal S, Thompson D, Renwick A, Elliott A,
Kelly P, Barfoot R, Chagtai T, Jayatilake H, Ahmed M, Spanova K,
North B, McGuffog L, Evans DG, Eccles D; Breast Cancer
Susceptibility Collaboration (UK); Easton DF, Stratton MR and
Rahman N: Truncating mutations in the Fanconi anemia J gene BRIP1
are low penetrance breast cancer susceptibility alleles. Nat Genet.
38:1239–1241. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
51.
|
Harada H, Kitagawa M, Tanaka N, Yamamoto
H, Harada K, Ishihara M and Taniguchi T: Anti-oncogenic and
oncogenic potentials of interferon regulatory factors-1 and -2.
Science. 259:971–974. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
52.
|
Xie RL, Gupta S, Miele A, Shiffman D,
Stein JL, Stein GS and van Wijnen AJ: The tumor suppressor
interferon regulatory factor 1 interferes with SP1 activation to
repress the human CDK2 promoter. J Biol Chem. 278:26589–26596.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
53.
|
Mori K, Yoshida K, Tani J, Nakagawa Y,
Hoshikawa S and Ito S: Double-stranded RNA-induced interferon
regulatory factor-1 gene expression in FRTL-5 rat thyroid cells.
Mol Cell Endocrinol. 84:77–86. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
54.
|
Wang J, Zhang WJ, Zhang YS, Chen Y, Zou B,
Jiang B, Pang R, Gu Q, Qiao L, Lan HY, Kung HF and Wong BC: c-Jun
N-terminal kinase (JNK1) upregulates XIAP-associated factor 1
(XAF1) through interferon regulatory factor 1 (IRF-1) in
gastrointestinal cancer. Carcinogenesis. 30:222–229. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
55.
|
Nguyen H, Lin R and Hiscott J: Activation
of multiple growth regulatory genes following inducible expression
of IRF-1 or IRF/RelA fusion proteins. Oncogene. 15:1425–1435. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
56.
|
Savitsky D, Tamura T, Yanai H and
Taniguchi T: Regulation of immunity and oncogenesis by the IRF
transcription factor family. Cancer Immunol Immunother. 59:489–510.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
57.
|
Strasser A, Harris AW, Jacks T and Cory S:
DNA damage can induce apoptosis in proliferating lymphoid cells via
p53-independent mechanisms inhibitable by Bcl-2. Cell. 79:329–339.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
58.
|
Tamura T, Ishihara M, Lamphier MS, Tanaka
N, Oishi I, Aizawa S, Matsuyama T, Mak TW, Taki S and Taniguchi T:
An IRF-1-dependent pathway of DNA damage-induced apoptosis in
mitogen-activated T lymphocytes. Nature. 376:596–599. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
59.
|
Bowie ML, Ibarra C and Seewalt VL: IRF-1
promotes apoptosis in p53-damaged basal-type human mammary
epithelial cells: a model for early basal-type mammary
carcinogenesis. Adv Exp Med Biol. 617:367–374. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
60.
|
Gao J, Senthil M, Ren B, Yan J, Xing Q, Yu
J, Zhang L and Yim JH: IRF-1 transcriptionally upregulates PUMA,
which mediates the mitochondrial apoptotic pathway in IRF-1-induced
apoptosis in cancer cells. Cell Death Differ. 17:699–709. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
61.
|
Lallemand C, Palmieri M, Blanchard B,
Meritet JF and Tovey MG: GAAP-1: a transcriptional activator of p53
and IRF-1 possesses proapoptotic activity. EMBO Rep. 3:153–158.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
62.
|
Kim EJ, Lee JM, Namkoong SE, Um SJ and
Park JS: Interferon regulatory factor-1 mediates
interferon-gamma-induced apoptosis in ovarian carcinoma cells. J
Cell Biochem. 85:369–380. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
63.
|
Fulda S and Debatin KM: IFN gamma
sensitizes for apoptosis by upregulating caspase-8 expression
through the Stat1 pathway. Oncogene. 21:2295–2308. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
64.
|
Bouker KB, Skaar TC, Riggins R, Harburger
DS, Fernandez DR, Zwart A, Wang A and Clarke R: Interferon
regulatory factor-1 (IRF-1) exhibits tumor suppressor activities in
breast cancer associated with caspase activation and induction of
apoptosis. Carcinogenesis. 26:1527–1535. 2005. View Article : Google Scholar
|
|
65.
|
De Ambrosis A, Casciano I, Croce M, Pagnan
G, Radic L, Banelli B, Di Vinci A, Allemanni G, Tonini GP, Ponzoni
M, Romani M and Ferrini S: An interferon-sensitive response element
is involved in constitutive caspase-8 gene expression in
neuroblastoma cells. Int J Cancer. 120:39–47. 2007.PubMed/NCBI
|
|
66.
|
Tomita Y, Bilim V, Hara N, Kasahara T and
Takahashi K: Role of IRF-1 and caspase-7 in IFN-gamma enhancement
of Fas-mediated apoptosis in ACHN renal cell carcinoma cells. Int J
Cancer. 104:400–408. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
67.
|
Park SY, Seol JW, Lee YJ, Cho JH, Kang HS
and Kim IS: IFN-gamma enhances TRAIL-induced apoptosis through
IRF-1. Eur J Biochem. 271:4222–4228. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
68.
|
Negishi H, Fujita Y, Yanai H, Sakaguchi S,
Ouyang X, Shinohara M, Takayanagi H, Ohba Y, Taniguchi T and Honda
K: Evidence for licensing of IFN-gamma-induced IFN regulatory
factor 1 transcription factor by MyD88 in toll-like receptor
dependent gene induction program. Proc Natl Acad Sci USA.
103:15136–15141. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
69.
|
Taki S, Sato T, Ogasawara K, Fukuda T,
Sato M, Hida S, Suzuki G, Mitsuyama M, Shin EH, Kojima S, Taniguchi
T and Asano Y: Multistage regulation of Th1-type immune responses
by the transcription factor IRF-1. Immunity. 6:673–679. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
70.
|
Trinchieri G: Biology of natural killer
cells. Adv Immunol. 47:187–376. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
71.
|
Ogasawara K, Hida S, Azimi N, Tagaya Y,
Sato T, Yokochi T, Waldmann TA, Taniguchi T and Taki S: Requirement
for IRF-1 in the microenvironment supporting development of natural
killer cells. Nature. 391:700–703. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
72.
|
Gabriele L, Fragale A, Borghi P, Sestili
P, Stellacci E, Venditti M, Schiavoni G, Sanchez M, Belardelli F
and Battistini A: IRF-1 deficiency skews the differentiation of
dendritic cells toward plasmacytoid and tolerogenic features. J
Leukoc Biol. 80:1500–1511. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
73.
|
Matsuyama T, Kimura T, Kitagawa M, Pfeffer
K, Kawakami T, Watanabe N, Kundig TM, Amakawa R, Kishihara K and
Wakeham A: Targeted disruption of IRF-1 or IRF-2 results in
abnormal type I IFN gene induction and aberrant lymphocyte
development. Cell. 75:83–97. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
74.
|
Reis LF, Ruffner H, Stark G, Aguet M and
Weissmann C: Mice devoid of interferon regulatory factor 1 (IRF-1)
show normal expression of type I interferon genes. EMBO J.
13:4798–4806. 1994.PubMed/NCBI
|
|
75.
|
White LC, Wright KL, Felix NJ, Ruffner H,
Reis LF, Pine R and Ting JP: Regulation of LMP2 and TAP1 genes by
IRF-1 explains the paucity of CD8 + T cells in IRF-1−/− mice.
Immunity. 5:365–376. 1996.PubMed/NCBI
|
|
76.
|
Penninger JM, Sirard C, Mittrucker HW,
Chidgey A, Kozieradzki I, Nghiem M, Hakem A, Kimura T, Timms E,
Boyd R, Taniguchi T, Matsuyama T and Mak TW: The interferon
regulatory transcription factor IRF-1 controls positive and
negative selection of CD8+ thymocytes. Immunity. 7:243–254.
1997.PubMed/NCBI
|
|
77.
|
Lohoff M, Ferrick D, Mittrucker HW, Duncan
GS, Bischof S, Rollinghoff M and Mak TW: Interferon regulatory
factor-1 is required for a T helper 1 immune response in vivo.
Immunity. 6:681–689. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
78.
|
Wiwanitkit V: Interaction between
interferon regulatory factor-1 and human papillomavirus E7 oncogene
in cervical cancer: an ontology study. Taiwan J Obstet Gynecol.
48:138–141. 2009. View Article : Google Scholar
|
|
79.
|
MacMicking J, Xie QW and Nathan C: Nitric
oxide and macrophage function. Annu Rev Immunol. 15:323–350. 1997.
View Article : Google Scholar : PubMed/NCBI
|