Introduction
Osteosarcoma is the most common primary malignant
bone neoplasm with poor prognosis in children and young
adolescents. In the past 27 years, the rate of disease-free
survival appeared to be stable (62 and 66% for patients treated in
two respective periods 1986–1989 and 1997–1999 in our studies),
despite the use of intensive neoadjuvant chemotherapy and radiation
therapy (1). Thus, novel agents
need to be developed to establish an effective therapeutic strategy
against osteosarcoma.
Bisphosphonates are analogs of endogenous
pyrophosphate that strongly bind to hydroxyapatite on the bone
surface. Nitrogen-containing bisphosphonates (N-BPs) inhibit the
activity of farnesyl diphosphate (FPP) synthase, a key enzyme in
the mevalonate pathway, and thereby reduce the prenylation of
proteins that are essential for normal cell function and survival
(2,3). N-BPs may have direct cytostatic and
anti-proliferative effects against a variety of tumor cells,
including myeloma, breast, prostate and pancreatic cancers, in a
concentration- and time-dependent manner (4–7).
Previously, it was reported that N-BPs may also
exhibit direct effects on osteoblast cells by altering the
expression of receptor activator of nuclear factor-κB ligand
(RANKL) and osteoprotegerin (OPG) (10). OPG is a soluble decoy receptor for
RANKL that blocks osteoclast formation by inhibiting the binding of
RANKL to the receptor activator of nuclear factor-κB (RANK)
(8,9). The OPG/RANK/RANKL system was
demonstrated to be abnormally regulated in several malignant
osteolytic pathologies such as osteosarcoma (10). By contrast, production of its
endogenous counteracting decoy receptor OPG is either inhibited or
too low to compensate for the increase in RANKL production. Thus,
the OPG/RANK/RANKL system is essential for understanding of the
pathophysiology of the bone microenvironment and offers
pharmacological targets for new antitumor drugs (11,12).
Preclinical data have indicated that N-BPs
accumulated in bone are able to either directly or indirectly
inhibit osteoclastic resorption and alter the bone microenvironment
(13,14). It was confirmed that N-BPs inhibit
proliferation, decrease viability and induce apoptosis in various
tumor cells. However, few studies have examined OPG and RANKL
levels in the MG-63 osteosarcoma cell line as well as the effects
of N-BPs on these cytokines. Zoledronic acid (ZOL) is a
nitrogen-containing third generation bisphosphonate (N-BP). In the
present study, we examined the effects of ZOL on MG-63 cells. We
also assessed the expression of RANKL and OPG in MG-63 cells
treated with or without ZOL.
Materials and methods
Drug preparation
The neutralized sodium salt of ZOL (Novartis, Basel,
Switzerland) was dissolved in sterile double distilled
H2O at a final concentration of 10 mM. The stock
solution of the ZOL salt was aliquoted and maintained at −20°C for
long-term storage.
Cell lines and culture maintenance
MG-63 human osteosarcoma cells were cultured in
Eagle’s minimum essential medium (EMEM) with 2 mM L-glutamine and
Earle’s BSS adjusted to contain 1.5 g/l sodium bicarbonate, 0.1 mM
non-essential amino acids, 1.0 mM sodium pyruvate and 10%
heat-inactivated fetal bovine serum in a humidified atmosphere of
5% CO2 at 37°C.
Cell growth and viability assays
Cells (100 μl, 2×104 cells/ml) were
plated in each well in 96-well plates and allowed to attach for 24
h. Cells were then treated with 0, 0.3, 1, 3, 10, 30, 100, 300 and
1000 μM of ZOL for 24, 48 and 72 h. The proportion of viable cells
was determined by MTT assay following the manufacturer’s
instructions. In brief, the cells were incubated with MTT (0.5 g/l)
for 4 h. The formazan precipitate was dissolved in 200 μl DMSO and
the absorbance at 550 nm was measured with a Benchmark microplate
reader. The tumor inhibitory rate was calculated as (1 - OD
experimental group/OD control group) × 100%. Experiments were
performed in triplicate.
Analysis of apoptosis and cell cycle
arrest
Apoptosis induced by ZOL was quantified using the
Annexin V-FITC Apoptosis Detection kit I (BD Biosciences, Franklin
Lakes, NJ, USA). MG-63 cells were treated with 50 μM of ZOL for 24,
48 and 72 h and then washed twice with cold PBS and re-suspended in
1× binding buffer at a concentration of 1×106 cells/ml.
The solution (100 μl, 1×105 cells) was transfered to a
5-ml culture tube and 5 μl Annexin V-FITC and 5 μl propidium iodide
(PI) were added. The cells were gently vortexed and incubated for
15 min at room temperature (25°C) in the dark. 1× binding buffer
(400 μl) was added to each tube. Samples were analyzed by flow
cytometry within 1 h. In the dual parameter fluorescent dot plots,
the cells in early and late apoptosis were counted. Total cell
apoptosis was defined as the sum of cells in early and late
apoptosis. Experiments were performed in triplicate.
Confluent MG-63 cells (treated with increasing
concentrations of ZOL for 24, 48 and 72 h) were removed from
culture dishes by trypsinization and fixed with ice-cold 70%
ethanol at −20°C overnight. Cells were washed with PBS, treated
with DNase A (200 g/l, Sigma, St. Louis, MO, USA) and stained with
PI (50 g/l, Sigma) at room temperature for 30 min in the dark. Cell
cycle distribution was determined by the FACSCalibur flow cytometer
(BD Biosciences).
Hoechst 33258 staining and electron
microscopy
Cells were treated with 50 μM of ZOL for 72 h,
harvested, cytospun onto glass slides and fixed with a 50% solution
of fixative (3:1 methanol/acetic acid). The preparations were
stained with Hoechst 33258 (10 mg/l Sigma) for 30 min, rinsed,
dried and visualized under a fluorescence microscope (Nikon,
Japan). Confluent MG-63 cells were treated with 50 μM of ZOL for 72
h. Cells were trypsinized, centrifuged, fixed in 2.5%
glutaraldehyde, postfixed in 2% osmium tetroxide and embedded in
Luveak-812 (Nacalai Tesque, Japan). Ultrathin sections were stained
with lead citrate and uranyl acetate and examined with a JEM-1230
electron microscope (Jeol Tokyo, Japan).
Reverse-transcription polymerase chain
reaction (RT-PCR)
Total cellular RNA was isolated using the RNAgents
Total RNA Isolation system (Promega, Madison, WI, USA). The cDNA
was synthesized using a reverse-transcription system (Promega).
RT-PCR was performed using Takara Ex Taq Hot Start Version (Takara
Bio, Shiga, Japan) and amplified by PCR using the specific primers
(Sangon, Shanghai, China): 5′-TGCT GTTCCTACAAAGTTTACG-3′ (sense)
and 5′-CTTTGATGC TTTAGTGCGTG-3′ (antisense) for OPG; 5′-GCCAGTGGG
AGATGTTAG-3′ (sense) and 5′-TTAGCTGCAAGTTTT CCC-3′ (antisence) for
RANKL; 5′-ACCCACACGGTGCCC ATCTACGAGG-3′ (sense) and
5′-AGCTCGTAGCTCTTCT CCAGGGAGG-3′ (antisense) for β-actin. Following
denaturation at 95°C for 3 min, samples were amplified for 30
cycles for OPG (95°C, 1 min; 57°C, 1 min; 72°C, 1 min) and RANKL
(95°C, 1 min; 55°C, 1 min; 72°C, 1 min) and 30 cycles for β-actin
(95°C, 1 min; 54°C, 1 min; 72°C, 1 min). The PCR products were
subjected to electrophoresis on 1.5% agarose gel with ethidium
bromide and observed under UV light.
Western blot analysis
MG-63 cells (5×105) were plated and
treated with 0, 1, 10, 100 μM of ZOL for 72 h. Cells were then
lysed with a lysis buffer composed of 50 mmol/l Tris-HCl (pH 8.0),
150 mmol/l NaCl, 0.1% Triton X-100, 0.01 g/l aprotinin and 0.05 g/l
phenylmethylsulfonyl fluoride. Protein was quantified by the
Bradford method and equal amounts of protein were loaded and
electrophoresed on a 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis mini-gel. Proteins were transferred to a PVDF
membrane and preblocked with casein PBS and 0.05% Tween-20 for 1 h
at room temperature. Membranes were incubated with mouse monoclonal
antibody against OPG (Santa Cruz Biotechnology, Santa Cruz, CA,
USA). Horseradish peroxidase-conjugated secondary antibody against
anti-mouse monoclonal antibody was used and protein bands were
visualized with enhanced chemiluminescence reagent (ECL kit,
Amersham, UK).
Statistical analysis
Statistical significance was determined using the
Student’s t-test, using SPSS 13.0. P<0.05 was considered to
indicate a statistically significant result.
Results
Effect of ZOL on cell proliferation in
MG-63 cells
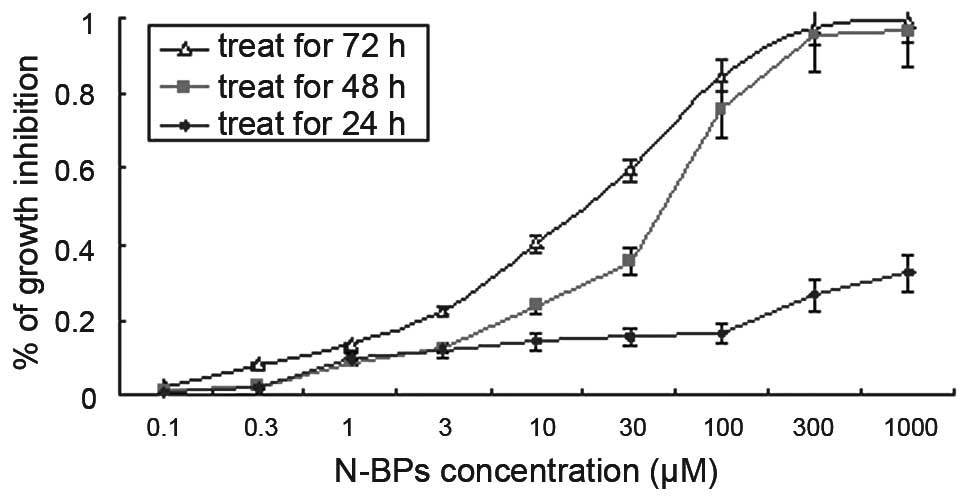
To evaluate the growth inhibitory effect of N-BPs on
the MG-63 cell line, we used the MTT assay and calculated the
IC50 values. There was no significant (P>0.001)
growth inhibition in a dose-dependent manner following treatment
with N-BPs for 24 h. However, following treatment for 48 h, ZOL
significantly inhibited (P<0.001) MG-63 cell growth in a
dose-dependent manner (Fig. 1). The
calculated IC50 value for ZOL was 52.37±1.0 μM,
following treatment with N-BPs for 72 h. This concentration was
used for subsequent experiments.
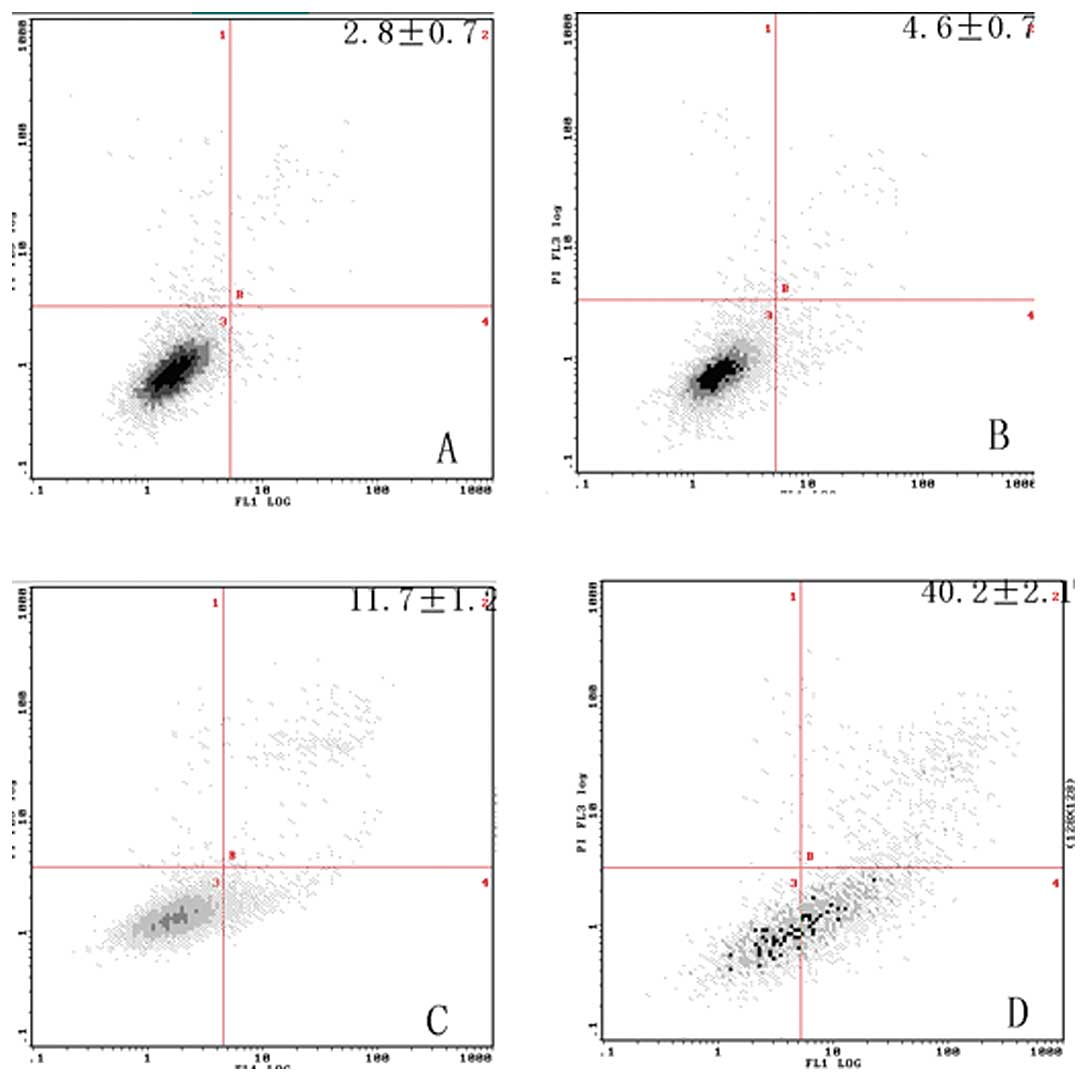
Induction of apoptosis
To determine the effects of ZOL on the induction of
apoptosis in MG-63 cells, the Annexin V assay was performed. ZOL
(50 μM) treated for 72 h induced 42.2±2.1% apoptotic cells
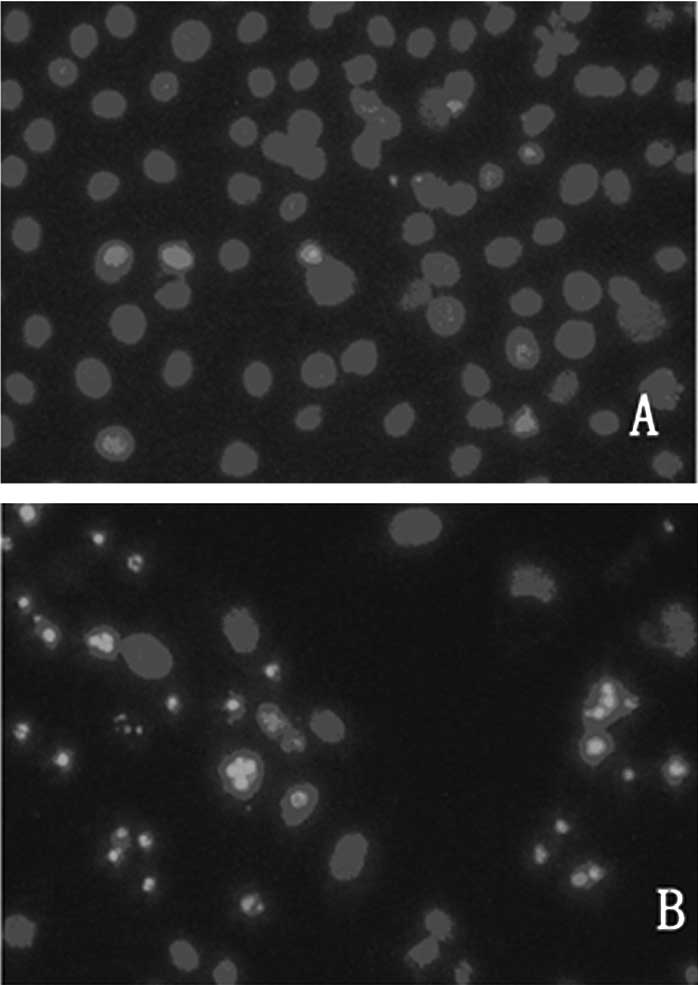
(Fig. 2). The preparations were
stained with Hoechst 33258 and examined under a fluorescence
microscope (Fig. 3). The blue
emission light in apoptotic cells was much brighter than that
observed in the control cells. Condensed chromatin was also
observed in numerous treated cells and several of these cells
formed apoptotic bodies, which is one of the classic
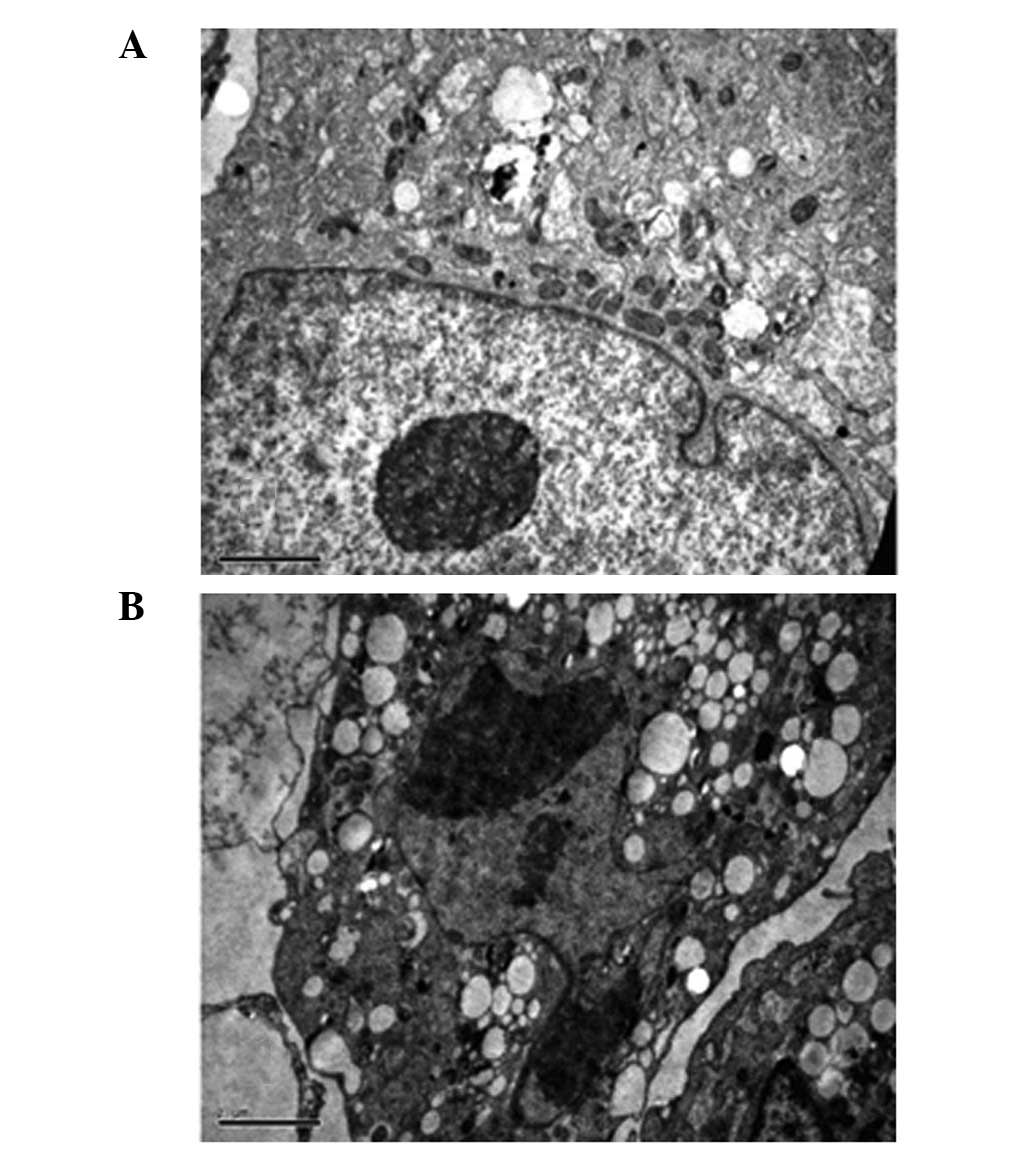
characteristics of apoptotic cells. As shown by electron
micrographs (Fig. 4), control cells
demonstrated an integrated nuclear membrane, relatively homogeneous
chromatin and extensive membrane interdigitations and microvilli.
Following treatment with 50 μM of ZOL for 72 h, MG-63 cells were
characterized by condensation into dense granules or blocks,
migration of nuclear chromatin, formation of apoptotic bodies and
numerous vacuoles in the cytoplasm. Concurrently, membrane
microvilli also disappeared. These changes indicated the apoptosis
of MG-63 cells.
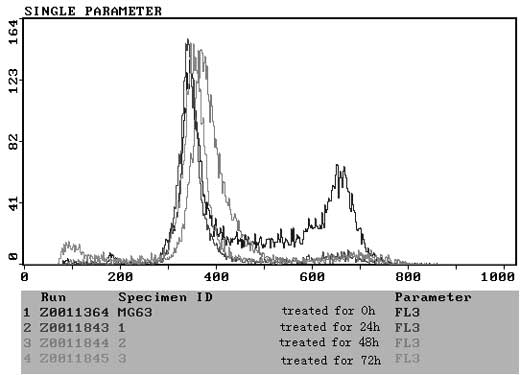
ZOL induces S-phase arrest in MG-63
cells
To elucidate the molecular mechanism underlying the
antitumor activity of N-BP, we examined the cell cycle distribution
in MG-63 cells by flow cytometric analysis following PI staining.
Significant changes were observed in the sub-G1, S, and
G2/M phase. Untreated control cells were found to have
few or no cells in the apoptotic peak and a distribution of cells
in the G0–G1, S and G2/M phase
typical of a proliferating cancer cell line. ZOL treatment induced
an increase in the sub-G1 peak in all cell lines, and an
increase in the percentage of cells in the G1 phase was
observed following treatment with 50 μM ZOL in a time-dependent
manner (Fig. 5). In view of the
above-mentioned growth-inhibitory effects, experiments were
performed to determine whether N-BP also induced apoptosis in the
cell lines. A time-dependent increase in apoptosis was observed in
cells treated with ZOL.
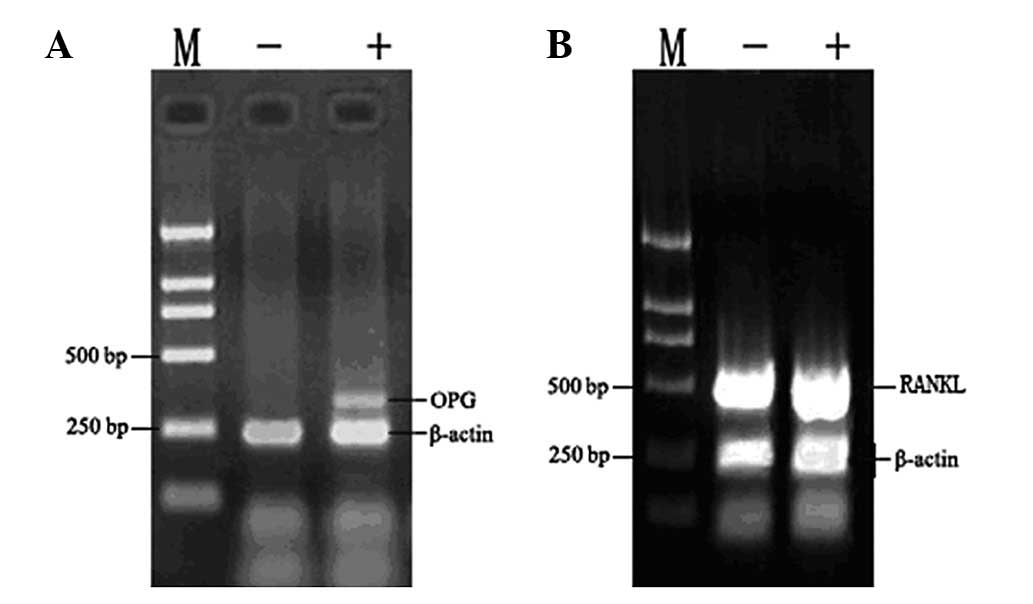
Effect of ZOL on osteoprotegerin
expression in vitro
The RANK/RANKL/OPG pathway is important in the
regulation of bone formation. Therefore, we aimed to determine
whether ZOL has any impact on the expression of these genes. ZOL
had no effect on the expression of RANKL in MG-63 cells (Fig. 6). By contrast, ZOL-treated cells
demonstrated a significantly higher expression of OPG. β-actin was
used as an internal loading control. Densitometric analysis
revealed that ZOL induced 2.2-fold increases in OPG expression in
MG-63 cells. There was no significant difference in the expression
of either RANKL or RANK in the MG-63 cells.
Western blot analysis was performed to detect the
OPG protein levels in cell culture media following treatment with
ZOL in MG-63 cells. The OPG protein level increased following
incubation with 0, 1, 10, 100 μM ZOL for 72 h (Fig. 7). The protein expression reached
peak levels at 10 μM and then decreased at 100 μM. ZOL stimulated
osteoprotegerin protein production in the MG-63 cells.
Discussion
A number of studies have demonstrated the inhibitory
potential of bisphosphonates on bone metastases from different
solid tumors, including breast, prostate and pancreatic cancer and
murine osteosarcoma (5,6,7,15). In
this study, we demonstrated that ZOL significantly inhibited human
osteosarcoma growth and induced apoptosis in in vitro
assays. Our data suggest that ZOL had direct effects on
osteosarcoma cells, including the inhibition of tumor cell
proliferation (Fig. 1) and
induction of apoptosis (Fig. 2).
These results are consistent with those of previous reports showing
that N-BPs are able to inhibit cell proliferation and induce
apoptosis in osteoclasts, myeloma, neuroblastoma and lung cancer
cells (4,13,16,17).
It has been suggested that N-BPs, including ZOL, are
capable of inhibiting the activities of FPP synthase and
geranylgeranyl diphosphate (GGPP) synthase, which are essential for
the activation of FPP and GGPP. It should be noted that the
activation of FPP and GGPP may result in the prenylation of small
GTP-binding proteins including Ras and corresponding anti-apoptotic
effects. Thus, it can be postulated that N-BPs induce cell
apoptosis (18,19). The results of the flow cytometric
analysis confirmed that treatment of MG-63 with ZOL increased the
G1 cell population (Fig.
5). From these findings, it is suggested that N-BPs exerted
their anti-proliferative effect against MG-63 by the induction of
cell apoptosis through the small GTP-binding proteins associated
signal transduction pathway (20).
Dunford et al demonstrated that YM529 showed direct
antitumor effects on NSCLC cells, induced apoptosis and caused
G1 arrest of the cell cycle through downregulation of
the phosphorylation of ERK1/2 (18). Ory et al also confirmed that
ZOL inhibited proliferation and increased atypical apoptosis in
several osteosarcoma cell lines. However, these authors found that
ZOL caused cell cycle arrest in the S and G2/M phases,
through the control of the intra-S DNA checkpoint at high doses of
ZOL, or through the control of the G1/S DNA checkpoint
at low doses (21).
In addition to apoptosis, N-BPs may regulate growth
factors and cytokines that affect cancer cell progression. OPG and
RANKL are able to affect the cancer phenotype. Bisphosphonates have
been shown to increase OPG expression in osteoblast cells in in
vitro assays. The increased OPG reduces osteolytic activity.
Other studies have shown that bisphosphonates are capable of
inhibiting FPP synthase in the mevalonate pathway, as well as the
OPG pathway (22,23). We observed that ZOL caused a
downregulated expression of two pro-osteolytic molecules,
consistent with previous reports. Our data indicate that ZOL
upregulated the OPG expression (Figs.
6 and 7). This would in turn
lead to a decrease in the RANKL/OPG ratio and decreased osteoclast
activity (24). Ishii et al
found that the sRANKL/OPG ratio decreased significantly following
therapy in several myeloma patients. The ratio of serum RANKL/OPG
correlated with the presence of osteolytic lesions and was a strong
predictor of five-year survival (25). Mintz et al (26) defined a set of 104 genes that
characterize poor histological response to chemotherapy in
osteosarcoma. These authors found that a marked decrease of −5.44
of OPG suggested the involvement of osteoclast promotion in poorly
responsive osteosarcoma tumors. They believed that the use of
bisphosphonate analogs should be considered as a potential
therapeutic intervention to suppress bone remodeling and tumor
osteolysis involved in osteosarcoma chemotherapy resistance
(26). In their study, Grimaud
et al observed an increase in the RANKL/OPG ratio in the
serum of patients with high-grade osteosarcoma (27). All of these findings suggest the
potential involvement of the RANK/RANKL/OPG axis in osteosarcoma
(28). OPG would decrease tumor
burden and select plasma parameters of tumor burden and bone
resorption by inhibiting osteoclastic bone resorption and tumor
growth in an intratibial injection model of bone metastasis of a
human pulmonary squamous cell carcinoma (29).
Previous studies have shown that N-BPs exert
antitumor properties and interact synergistically with other
antineoplastic agents (30). As
bisphosphonates accumulate in bone, they exert cytostatic effects
on tumor cells in bone metastases, either via osteoclast inhibition
or alterations in the bone microenvironment. Further in vivo
studies are required to optimize the dosing regimen of N-BPs to
fully exploit their antitumor potential. However, the present
experiments do not completely rule out that OPG inhibits tumor
growth specifically in the bone microenvironment. Further
experiments are needed to determine the true effects of OPG on
osteosarcoma.
In conclusion, our results demonstrate that ZOL is
able to directedly inhibit cell growth and induce apoptosis of
MG-63 cells. Increased expression of osteoprotegerin was also
observed, which may be an indirect effect via an alteration in the
local microenvironment.
References
|
1
|
Bacci G, Longhi A, Fagioli F, Briccoli A,
Versari M and Picci P: Adjuvant and neoadjuvant chemotherapy for
osteosarcoma of the extremities: 27 year experience at Rizzoli
Institute, Italy. Eur J Cancer. 41:2836–2845. 2005.PubMed/NCBI
|
|
2
|
Perry CM and Figgitt DP: Zoledronic acid:
a review of its use in patients with advanced cancer. Drugs.
64:1197–1211. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Green JR: Skeletal complications of
prostate cancer: pathophysiology and therapeutic potential of
bisphosphonates. Acta Oncol. 44:282–292. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Baulch-Brown C, Molloy TJ, Yeh SL, Ma D
and Spencer A: Inhibitors of the mevalonate pathway as potential
therapeutic agents in multiple myeloma. Leuk Res. 31:341–352. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Iguchi K, Nakano T, Usui S and Hirano K:
Incadronate inhibits aminopeptidase N expression in prostatic PC-3
cells. Cancer Lett. 237:223–233. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nakajima H, Magae J, Tsuruga M, Sakaguchi
K, Fujiwara I, Mizuta M, Sawai K, Yamagishi H and Mizuta N:
Induction of mitochondria-dependent apoptosis through the
inhibition of mevalonate pathway in human breast cancer cells by
YM529, a new third generation bisphosphonate. Cancer Lett.
253:89–96. 2007. View Article : Google Scholar
|
|
7
|
Tassone P, Tagliaferri P, Viscomi C,
Palmieri C, Caraglia M, D’Alessandro A, Galea E, Goel A, Abbruzzese
A, Boland CR and Venuta S: Zoledronic acid induces
antiproliferative and apoptotic effects in human pancreatic cancer
cells in vitro. Br J Cancer. 88:1971–1978. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Im GI, Qureshi SA, Kenney J, Rubash HE and
Shanbhag AS: Osteoblast proliferation and maturation by
bisphosphonates. Biomaterials. 25:4105–4115. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Viereck V, Emons G, Lauck V, Frosch KH,
Blaschke S, Gründker C and Hofbauer LC: Bisphosphonates pamidronate
and zoledronic acid stimulate osteoprotegerin production by primary
human osteoblasts. Biochem Biophys Res Commun. 291:680–686. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wittrant Y, Théoleyre S, Chipoy C,
Padrines M, Blanchard F, Heymann D and Rédini F: RANKL/RANK/OPG:
new therapeutic targets in bone tumours and associated osteolysis.
Biochim Biophys Acta. 1704:49–57. 2004.PubMed/NCBI
|
|
11
|
Dougall WC and Chaisson M: The
RANK/RANKL/OPG triad in cancer-induced bone diseases. Cancer
Metastasis Rev. 25:541–549. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Baud’huin M, Duplomb L, Ruiz Velasco C,
Fortun Y, Heymann D and Padrines M: Key roles of the OPG-RANK-RANKL
system in bone oncology. Expert Rev Anticancer Ther. 7:221–232.
2007.PubMed/NCBI
|
|
13
|
Plotkin LI, Manolagas SC and Bellido T:
Dissociation of the pro-apoptotic effects of bisphosphonates on
osteoclasts from their anti-apoptotic effects on
osteoblasts/osteocytes with novel analogs. Bone. 39:443–452. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Green JR: Antitumor effects of
bisphosphonates. Cancer. 97:840–847. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Horie N, Murata H, Nishigaki Y, Matsui T,
Segawa H, Nogawa M, Yuasa T, Kimura S, Maekawa T, Fushiki S and
Kubo T: The third-generation bisphosphonates inhibit proliferation
of murine osteosarcoma cells with induction of apoptosis. Cancer
Lett. 238:111–118. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Koshimune R, Aoe M, Toyooka S, Hara F,
Ouchida M, Tokumo M, Sano Y, Date H and Shimizu N: Anti-tumor
effect of bisphosphonate (YM529) on non-small cell lung cancer cell
lines. BMC Cancer. 7:82007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dickson PV, Hamner JB, Cauthen LA, Ng CY,
McCarville MB and Davidoff AM: Efficacy of zoledronate against
neuroblastoma. Surgery. 140:227–235. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dunford JE, Thompson K, Coxon FP, Luckman
SP, Hahn FM, Poulter CD, Ebetino FH and Rogers MJ:
Structure-activity relationships for inhibition of farnesyl
diphosphate synthase in vitro and inhibition of bone resorption in
vivo by nitrogen-containing bisphosphonates. J Pharmacol Exp Ther.
296:235–242. 2001.
|
|
19
|
Ishikawa C, Matsuda T, Okudaira T, Tomita
M, Kawakami H, Tanaka Y, Masuda M, Ohshiro K, Ohta T and Mori N:
Bisphosphonate incadronate inhibits growth of human T-cell
leukaemia virus type I-infected T-cell lines and primary adult
T-cell leukaemia cells by interfering with the mevalonate pathway.
Br J Haematol. 136:424–432. 2007. View Article : Google Scholar
|
|
20
|
Caraglia M, Santini D, Marra M, Vincenzi
B, Tonini G and Budillon A: Emerging anti-cancer molecular
mechanisms of aminobisphosphonates. Endocr Relat Cancer. 13:7–26.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ory B, Blanchard F, Battaglia S, Gouin F,
Rédini F and Heymann D: Zoledronic acid activates the DNA S-phase
checkpoint and induces osteosarcoma cell death characterized by
apoptosis-inducing factor and endonuclease-G translocation
independently of p53 and retinoblastoma status. Mol Pharmacol.
71:333–343. 2007. View Article : Google Scholar
|
|
22
|
Mori K, Le Goff B, Berreur M, Riet A,
Moreau A, Blanchard F, Chevalier C, Guisle-Marsollier I, Léger J,
Guicheux J, et al: Human osteosarcoma cells express functional
receptor activator of nuclear factor-kappa B. J Pathol.
211:555–562. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Benassi MS, Chiechi A, Ponticelli F,
Pazzaglia L, Gamberi G, Zanella L, Manara MC, Perego P, Ferrari S
and Picci P: Growth inhibition and sensitization to cisplatin by
zoledronic acid in osteosarcoma cells. Cancer Lett. 250:194–205.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang J, Dai J, Qi Y, Lin DL, Smith P,
Strayhorn C, Mizokami A, Fu Z, Westman J and Keller ET:
Osteoprotegerin inhibits prostate cancer-induced osteoclastogenesis
and prevents prostate tumor growth in the bone. J Clin Invest.
107:1235–1244. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ishii R, Morimoto A, Ikushima S, Sugimoto
T, Asami K, Bessho F, Kudo K, Tsunematu Y, Fujimoto J and Imashuku
S: High serum values of soluble CD154, IL-2 receptor, RANKL and
osteoprotegerin in Langerhans cell histiocytosis. Pediatr Blood
Cancer. 47:194–199. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mintz MB, Sowers R, Brown KM, Hilmer SC,
Mazza B, Huvos AG, Meyers PA, Lafleur B, McDonough WS, Henry MM, et
al: An expression signature classifies chemotherapy-resistant
pediatric osteosarcoma. Cancer Res. 65:1748–1754. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Grimaud E, Soubigou L, Couillaud S,
Coipeau P, Moreau A, Passuti N, Gouin F, Redini F and Heymann D:
Receptor activator of nuclear factor kappaB ligand
(RANKL)/osteoprotegerin (OPG) ratio is increased in severe
osteolysis. Am J Pathol. 163:2021–2031. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Quinn JE, Brown LG, Zhang J, Keller ET,
Vessella RL and Corey E: Comparison of Fc-osteoprotegerin and
zoledronic acid activities suggests that zoledronic acid inhibits
prostate cancer in bone by indirect mechanisms. Prostate Cancer
Prostatic Dis. 8:253–259. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tannehill-Gregg SH, Levine AL, Nadella MV,
Iguchi H and Rosol TJ: The effect of zoledronic acid and
osteoprotegerin on growth of human lung cancer in the tibias of
nude mice. Clin Exp Metastasis. 23:19–31. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zheng Y, Zhou H, Brennan K, Blair JM,
Modzelewski JR, Seibel MJ and Dunstan CR: Inhibition of bone
resorption, rather than direct cytotoxicity, mediates the
anti-tumour actions of ibandronate and osteoprotegerin in a murine
model of breast cancer bone metastasis. Bone. 40:471–478. 2007.
View Article : Google Scholar : PubMed/NCBI
|