Introduction
Lung cancers, the leading cause of cancer-related
mortality in the world, accounted for 30% of all cancer mortalities
in 2010 (1). Clinically, there is
no curative therapy for lung cancer; all treatment modalities,
including combined chemotherapy and radiotherapy or intensive
high-dose chemotherapy have failed to prevent disease recurrence
and progression (2).
The recent availability of tyrosine kinase
inhibitors (TKI) that specifically block the phosphorylation and
function of epidermal growth factor receptor (EGFR) has provided a
novel therapeutic strategy. One such inhibitor is gefitinib
(Iressa), a selective EGFR-TKI (3).
It acts through competitive inhibition of ATP binding to the
receptor tyrosine kinase domain, resulting in growth delay in
cancer cell lines (4). Recent phase
II/III clinical studies showed that gefitinib was well tolerated
and provided antitumor activity in patients with lung cancer as
well as with other types of cancer when used as a monotherapy or in
combination with other therapies (5). However, the clinical data also showed
that only certain patients respond to gefitinib, indicating the
existence of an intrinsic or de novo resistance to this type
of TKI (6). In some clinical
studies, the absence of a direct association between EGFR
expression levels and response to the therapy with gefitinib was
noted (7). Even high EGFR
expression per se was not sufficient to determine
sensitivity to gefitinib, due to molecular defects in downstream
signaling pathways might impair transmission of the signal and the
action of the drug (8). A thorough
understanding of the downstream cellular targets of gefitinib will
allow the discovery of markers that serve to identify the subset of
patients likely to benefit from the treatment (9).
The transducer of erbB-2.1 (TOB1) gene was initially
identified as a member of the anti-proliferative TOB/BTG
(transducer of erbB-2/B-cell translocation gene) protein family,
which was discovered in the 1990s (10). Recent studies in regard to TOB1
showed that TOB1 contributed to the genesis, transformation and
progression of various malignancies, including lung cancer
(11–13). It has been reported that TOB1 was
upregulated following EGF stimulation in breast cancer cells;
however, the role of TOB1 in gefitinib treatment is poorly
understood (14).
In the present study, we investigated the function
of TOB1 in gefitinib treatment and its potential as a novel
therapeutic target in gefitinib lung cancer therapy.
Materials and methods
Cell culture
The human lung carcinoma cell lines NCI-H1975 and
A549 were purchased from American Type Culture Collection
(Manassas, VA, USA). Cells were maintained in DMEM supplemented
with 10% fetal bovine serum, L-glutamine (5 mmol/l), non-essential
amino acids (5 mmol/l), penicillin (100 U/ml), and streptomycin
(100 U/ml) (Invitrogen, Carlsbad, CA, USA), at 37°C in a humidified
5% CO2 atmosphere. Gefitinib (Iressa) was purchased from
AstraZeneca (Macclesfield, UK) and dissolved in DMSO to make 10
mmol/l stock solution. The study was approved by the Ethics
Committee of Soochow University, Jiangsu, Suzhou, China.
RNA interfering
For gene silencing, cells were transiently
transfected with 50 nmol/l of the small interfering RNA (siRNA)
using Lipofectamine™ RNAiMAX reagent (Invitrogen). Twenty-four
hours after transfection, the cells were treated with 10 μmol/l
gefitinib or DMSO vehicle. The sequences for TOB1-siRNA #1 were
GCUGUAAGCCCUACCUUCATT and UGAAGGUAG GGCUUACAGCTT (5’ to 3’).
Cell cycle analysis
Cell cycle analysis was performed using propidium
iodide staining. For this, cells were trypsinized, washed once with
1X phosphate-buffered saline (PBS), and then fixed in 75% ethanol
for at least 4 h. Fixed cells were centrifuged and washed with 1X
PBS and stained with 10 μg/ml propidium iodide in PBS containing
300 μg/ml RNase (Sigma-Aldrich, St. Louis, MO, USA) for 1 h, and
then filtered with a 53 μm nylon mesh. The cell cycle distribution
was calculated from 10,000 cells using ModFit LT software (Becton
Dickinson, CA, USA) using FACSCalibur (Becton Dickinson).
Immunofluorescence
Cells were grown on sterile coverslips and fixed in
4% formaldehyde before being permeabilized in 0.25% Triton X-100.
Coverslips were blocked in PBS containing 3% BSA, and antibody
recognizing TOB1 was added at 10 μg/ml. Specific staining was
visualized with a secondary antibody conjugated to FITC 488 and
analyzed on a Zeiss confocal microscope.
Preparation of nuclear and cytoplasmic
extracts
Following treatment, cells were collected by
scraping and were washed three times with ice-cold PBS. Cells were
vortexed for 5 sec in CER II, and then the cells were lysed on ice
for 1 min. Nuclei were sedimented by 5 min centrifugation at 16,000
x g, and the supernatant containing the cytoplasmic fraction was
snap frozen. The pellet, containing the nuclei, was resuspended in
Nuclear Extraction reagent and rotated at maximum speed on a wheel
at 4°C for 40 min. The samples were then centrifuged at 4°C for 5
min at 16,000 x g. The supernatant containing the nuclear fraction
was snap frozen and kept at −80°C.
Western blot and immunoprecipitation
(IP)/immunoblot analyses
Western blot and IP analyses were performed as
previously described (15). For
western blot assay, the following primary antibodies were used for
immunoblotting (Santa Cruz Biotech, Santa Cruz, CA, 1:500
dilution): β-actin (C-4), TOB1 (H-18) and cyclin D1 (A-12). For the
IP/western blot, the anti-TOB1 (H-18) and anti-cyclin D1 (A-12)
antibodies were applied for immunoblot. The protein bands were
visualized using an enhanced chemiluminescence system (Union
Bioscience Corporation, Hangzhou, China) with prestained markers as
molecular size standards.
Statistical analysis
The data are presented as the means and standard
deviations. Statistical comparisons of the experimental results
between the treated group and the control group were made using a
two-tailed Student’s t-test. All statistical tests were performed
using SPSS version 17.0. P≤0.05 was considered to indicate a
statistically significant result.
Results
Gefitinib causes G0/G1 arrest and TOB1
overexpression in lung cancer cell lines
To examine the effect of gefitinib on cell
proliferation, we analyzed the cell cycle phase distribution by
flow-cytometric analysis of propidium iodide-stained cells after
treatment with 10 μmol/l gefitinib for 0, 12, 24 and 48 h. As shown
in Fig. 1A, gefitinib induced G0/G1
arrest in both lung cancer cell lines A549 and NCI-H1975, of which
A549 cells are EGFR wild-type and NCI-H1975 are EGFR mutated. To
investigate the possible effect of TOB1 in mediating cell cycle
arrest response to gefitinib, we compared the expression of TOB1 as
well as cyclin D1 in the two lung cancer cell lines after gefitinib
treatment. The result showed that gefitinib caused a significant
reduction in cyclin D1 expression (P<0.05), and an obvious
increase in TOB1 protein expression (P<0.05; Fig. 1B and C). This finding suggests that
TOB1 may be a candidate indicator for gefitinib activity and
response.
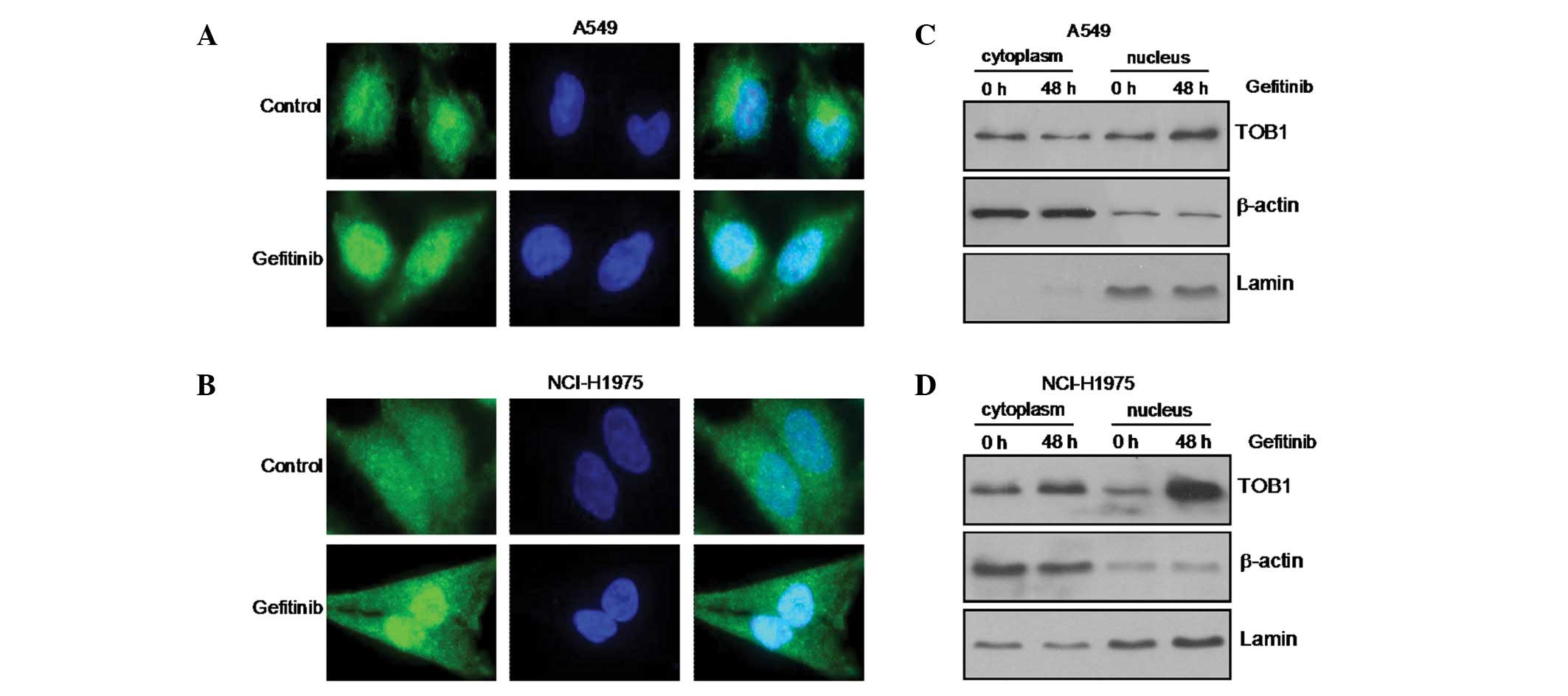
Gefitinib induces nuclear translocation
of TOB1
To investigate further the mechanism of action of
gefitinib, we next studied the subcellular localization of TOB1 in
response to gefitinib treatment by confocal microscopy following
staining with a TOB1-specific antibody. In the parental A549 and
NCI-H1975 cells, endogenous TOB1 resided both in the cytoplasm and
nucleus, whereas 48 h after gefitinib treatment, TOB1 relocated
predominantly to the nucleus (Fig. 2A
and B). The change in TOB1 localization was further confirmed
by western blot analysis of nuclear/cytoplasmic lysates. As shown
in Fig. 2C and D, treatment of A549
and NCI-H1975 cells with gefitinib resulted in a decrease in the
TOB1 level in the cytoplasm and a parallel increase in TOB1 in the
nucleus. Together, these results suggest that gefitinib induces
TOB1 overexpression and nuclear translocation in A549 and NCI-H1975
lung cancer cells to mediate G0/G1 cell cycle arrest.
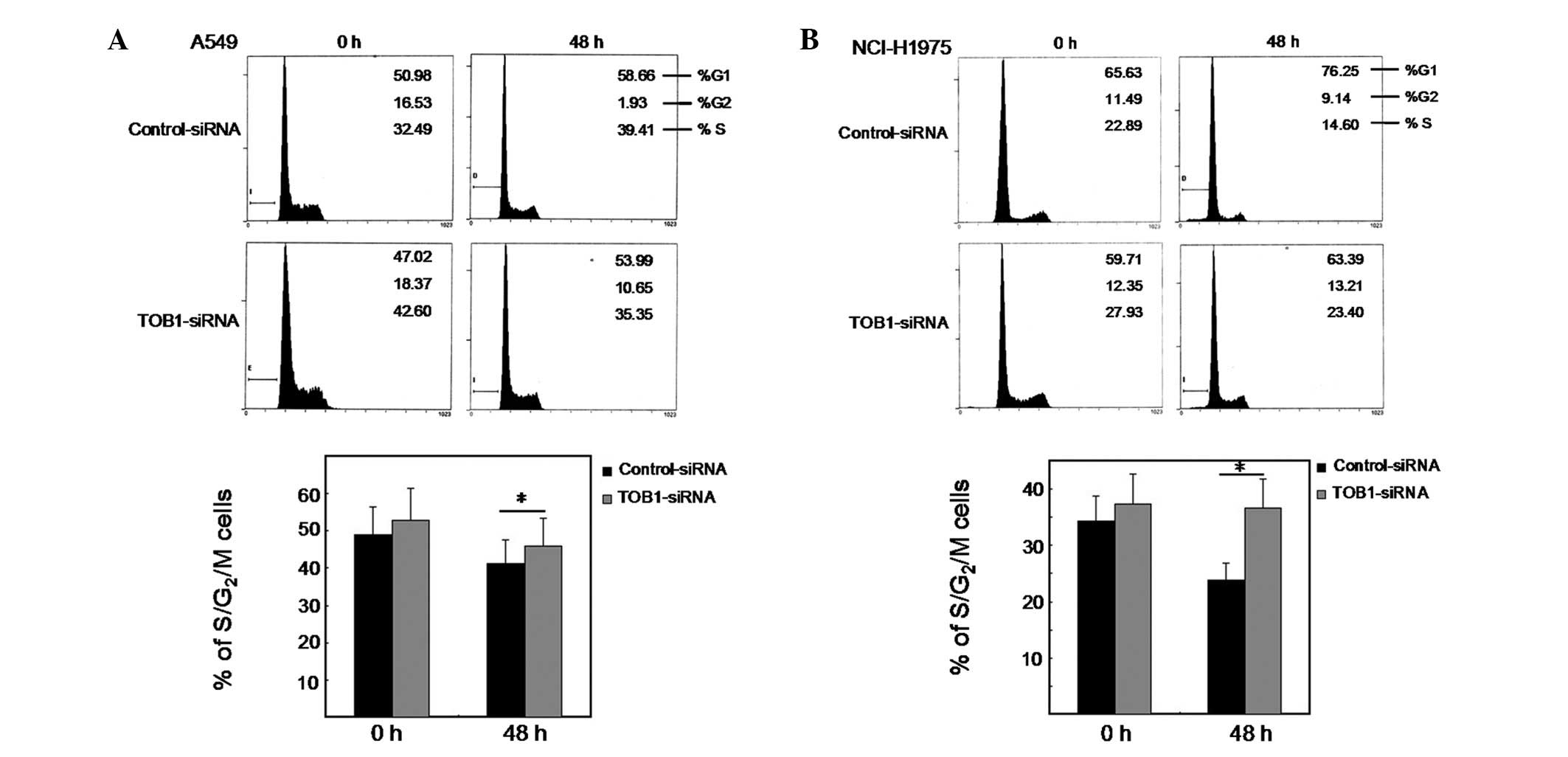
TOB1 knockdown suppresses
gefitinib-induced proliferative arrest
To certify the potential effect of TOB1 in response
to gefitinib treatment, we transfected the two lung cancer cell
lines with either a TOB1-specific siRNA or a nonspecific control
siRNA pool and studied the proliferation of these cells after
gefitinib treatment. Consistent with this, cell cycle analysis
showed that silencing of TOB1 by siRNA rescued cells from
gefitinib-induced proliferative arrest (Fig. 3A and B). This is evident from the
increase in the fraction of cells in S and G2/M phases in
TOB1-siRNA-transfected A549 and NCI-H1975 cells compared with
control siRNA transfected cells. The results suggest that gefitinib
mediates proliferative arrest in lung cancer cells, partly through
targeting TOB1 expression.
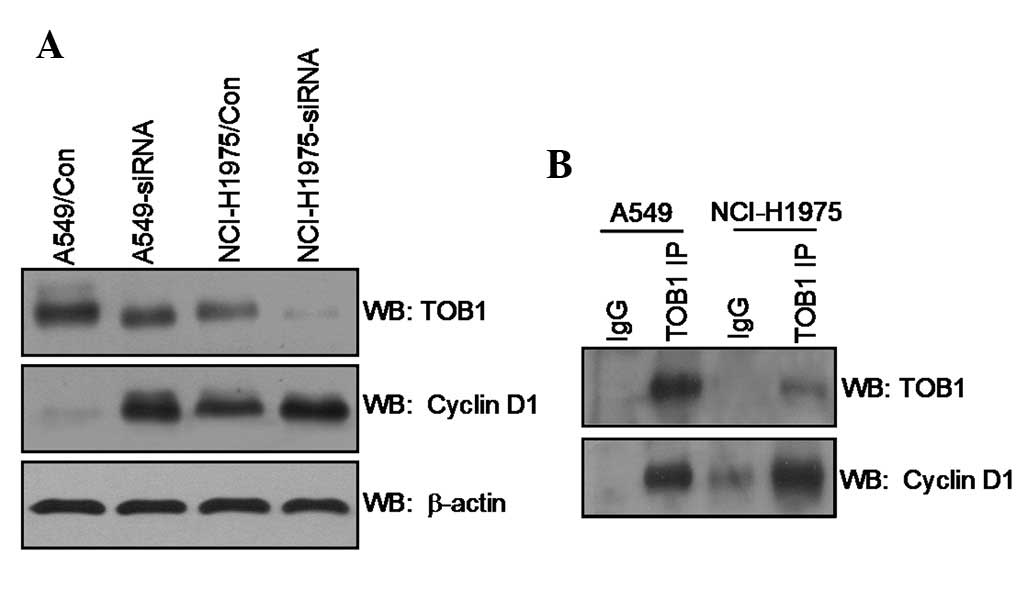
TOB1 interacts with cyclin D1
Western blot analysis was utilized to evaluate the
expression level of cyclin D1 in TOB1-siRNA transfected A549 cells
and NCI-H1975 cells compared with control siRNA transfected cells.
The result showed that TOB1 knockdown slightly increased cyclin D1
expression (Fig. 4A). The potential
interaction between cyclin D1 and TOB1 was examined through an
IP/immunoblot assay. For the first time we confirmed the presence
of cyclin D1 protein in the TOB1 IP of A549 and NCI-H1975 cells
(Fig. 4B).
Discussion
EGFR plays a central role in the development and
progression of lung cancer. As a consequence, EGFR has emerged as a
notable molecular target for lung cancer therapy, with monoclonal
antibodies and small-molecule tyrosine kinase inhibitors (TKIs),
such as gefitinib, being developed to specifically block EGFR
signaling (16). These anti-EGFR
agents have shown promising outcomes in initial preclinical and
clinical settings; however, there is a high rate of de novo
and acquired resistance, which limits the efficacy of such
treatments (17). Several
investigators have reported results from retrospective analyses of
associations between EGFR gene mutations and EGFR-TKI sensitivity
(18). These analyses indicated
that approximately 70–80% of mutation positive cases are EGFR-TKI
sensitive whereas in wild-type patients the response rate is
10–20%, but these mutations are uncommon (19).
The biological function of TOB1 in carcinogenesis
became apparent when it was observed that mice lacking TOB1
spontaneously developed tumors, primarily in lung, liver and lymph
nodes (20). In human lung cancers,
decreased TOB1 expression and increased phosphorylation-triggered
TOB1 inactivation are observed compared to that of normal
epithelial tissues (21).
Overexpression of TOB1 negatively regulates the cell cycle by
inhibiting G1 progression (22).
This evidence suggests that TOB1 is involved in the negative
regulation of cell growth. In the present study, using
immunofluorescence studies and western blot analysis of
nuclear/cytoplasmic cellular protein fractions, we observed that
endogenous TOB1 resided both in the cytoplasm and nucleus, whereas
TOB1 relocated predominantly to the nucleus following gefitinib
treatment. Gefitinib treatment causes cell cycle arrest
predominantly at the G0/G1 phase, which is associated with TOB1
overexpression and nuclear translocation. Furthermore, silencing of
the endogenous TOB1 expression by siRNA at least partially rescued
A549 and NCI-H1975 cells from undergoing proliferative arrest in
response to gefitinib, suggesting that the induction of TOB1
expression has a direct role in mediating the effect of gefitinib
on cell proliferation.
The well-known function ascribed to cyclin D1 is the
positive regulation of cell cycle distribution (23). In lung cancer cells, constitutive
cyclin D1 activation maintains retinoblastoma (RB) protein in a
phosphorylated state and promotes cell proliferation, thus likely
initiating tumorigenesis (24).
Previous studies have reported that TOB1 regulated proliferation by
affecting cyclin D1 activity (25).
Based on these findings, we investigated the mechanisms responsible
for the inhibited therapeutic efficacy of gefitinib in lung cancer
cells following TOB1 inhibition. Notably, using western blot
analysis, we found that cyclin D1 was increased after TOB1
silencing. Further analysis demonstrated that TOB1 can interact
with cyclin D1 in A549 and NCI-H1975 cells. These results suggest
that TOB1 interacting with cyclin D1 and nuclear translocation have
a direct role in mediating the gefitinib-induced proliferative
arrest. Taken together, our data showed that TOB1 may be a
potential target of anti-EGFR therapy in lung cancer. However,
since silencing of TOB1 expression in A549 and NCI-H1975 cells did
not completely rescue the cells from proliferation suppression, it
is reasonable to conclude that other mechanisms may be involved in
the response to gefitinib treatment.
In summary, the present study helps to define the
mechanism by which gefitinib mediates cell cycle arrest in lung
cancer. It also reveals that TOB1 could be a promising biomarker
for monitoring anti-EGFR therapy.
Acknowledgements
This study was supported by grants
from the Doctoral Fund of the Ministry of Education of China (Grant
No. K512602110), the College Nature Science Foundation of Jiangsu
Province (Grant No. SZ126821), the Social Development Projects of
Kunshan City (Grant No. KS1224) and the Priority Academic Program
Development of Jiangsu Higher Education Institutions.
References
|
1
|
Jemal A, Bray F, Center MM, et al: Global
cancer statistics. CA Cancer J Clin. 2:69–90. 2011. View Article : Google Scholar
|
|
2
|
Inamura K and Ishikawa Y: Lung cancer
progression and metastasis from the prognostic point of view. Clin
Exp Metastasis. 6:389–397. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wakeling AE, Guy SP, Woodburn JR, et al:
ZD1839 (Iressa): an orally active inhibitor of epidermal growth
factor signaling with potential for cancer therapy. Cancer Res.
62:5749–5754. 2002.PubMed/NCBI
|
|
4
|
Ciardiello F: Epidermal growth factor
receptor inhibitors in cancer treatment. Future Oncol. 1:221–234.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Maione P, Gridelli C, Troiani T, et al:
Combining targeted therapies and drugs with multiple targets in the
treatment of NSCLC. Oncologist. 11:274–284. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Polychronis A, Sinnett HD, Hadjiminas D,
et al: Preoperative gefitinib versus gefitinib and anastrozole in
postmenopausal patients with oestrogen-receptor positive and
epidermal-growt h-factor-receptor-positive primary breast cancer: a
double-blind placebo-controlled phase II randomised trial. Lancet
Oncol. 6:383–391. 2005. View Article : Google Scholar
|
|
7
|
Montemurro F, Valabrega G and Aglietta M:
Lapatinib: a dual inhibitor of EGFR and HER2 tyrosine kinase
activity. Expert Opin Biol Ther. 7:257–268. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ranson M, Hammond LA, Ferry D, et al:
ZD1839, a selective oral epidermal growth factor receptor-tyrosine
kinase inhibitor, is well tolerated and active in patients with
solid, malignant tumors: results of a phase I trial. J Clin Oncol.
20:2240–2250. 2002. View Article : Google Scholar
|
|
9
|
Krol J, Francis RE, Albergaria A, et al:
The transcription factor FOXO3a is a crucial cellular target of
gefitinib (Iressa) in breast cancer cells. Mol Cancer Ther.
6:3169–3179. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Matsuda S, Kawamura-Tsuzuku J, Ohsugi M,
et al: Tob, a novel protein that interacts with p185erbB2, is
associated with anti-proliferative activity. Oncogene. 4:705–713.
1996.PubMed/NCBI
|
|
11
|
Winkler GS: The mammalian
anti-proliferative BTG/Tob protein family. J Cell Physiol. 1:66–72.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang XM, Gao X, Zhang XH, et al: The
negative cell cycle regulator, Tob (transducer of ErbB-2), is
involved in motor skill learning. Biochem Biophys Res Commun.
4:1023–1027. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jia S and Meng A: Tob genes in development
and homeostasis. Dev Dyn. 4:913–921. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Helms MW, Kemming D, Contag CH, et al:
TOB1 is regulated by EGF-dependent HER2 and EGFR signaling, is
highly phosphorylated, and indicates poor prognosis in
node-negative breast cancer. Cancer Res. 12:5049–5056. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jiao Y, Sun KK, Zhao L, et al: Suppression
of human lung cancer cell proliferation and metastasis in vitro by
the transducer of ErbB-2.1 (TOB1). Acta Pharmacol Sin. 33:250–260.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Astsaturov I, Cohen RB and Harari P:
Targeting epidermal growth factor receptor signaling in the
treatment of head and neck cancer. Expert Rev Anticancer Ther.
6:1179–1193. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Scaltriti M and Baselga J: The epidermal
growth factor receptor pathway: a model for targeted therapy. Clin
Cancer Res. 12:5268–5272. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Johnston JB, Navaratnam S, Pitz MW, et al:
Targeting the EGFR pathway for cancer therapy. Curr Med Chem.
13:3483–3492. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Uchida A, Hirano S, Kitao H, et al:
Activation of downstream epidermal growth factor receptor (EGFR)
signaling provides gefitinib-resistance in cells carrying EGFR
mutation. Cancer Sci. 98:357–363. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Iwanaga K, Sueoka N, Sato A, et al:
Alteration of expression or phosphorylation status of tob, a novel
tumor suppressor gene product, is an early event in lung cancer.
Cancer Lett. 1:71–79. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ito Y, Suzuki T, Yoshida H, et al:
Phosphorylation and inactivation of Tob contributes to the
progression of papillary carcinoma of the thyroid. Cancer Lett.
2:237–242. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yanagie H, Tanabe T, Sumimoto H, et al:
Tumor growth suppression by adenovirus-mediated introduction of a
cell-growth-suppressing gene tob in a pancreatic cancer model.
Biomed Pharmacother. 4:275–286. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lapenna S and Giordano A: Cell cycle
kinases as therapeutic targets for cancer. Nat Rev Drug Discov.
8:547–566. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Felsher DW: Cancer revoked: oncogenes as
therapeutic targets. Nat Rev Cancer. 3:375–380. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Suzuki T, K-Tsuzuku J, Ajima R, et al:
Phosphorylation of three regulatory serines of Tob by Erk1 and Erk2
is required for Ras-mediated cell proliferation and transformation.
Genes Dev. 11:1356–1370. 2002. View Article : Google Scholar : PubMed/NCBI
|