Introduction
The search for effective anticancer agents has been
one of the most important areas in cancer control research. There
are numerous natural plants used in clinical therapy for cancer in
traditional Chinese medicine. The root of Cynanchum
auriculatum Royle ex Wight, also known as Baishouwu, has been
widely used in clinics since ancient times as an agent for
anti-aging and prolonging life. Its major components,
C21-steroidal glycosides (CG), are of considerable
interest due to their bioactivities. Previous studies have revealed
that the CGs isolated from Baishouwu were able to protect
hepatocytes and neurons, as well as cells in the digestive system
(1–3). Studies have also revealed the
anticancer activity of these compounds (4–8). A
number of CGs have been isolated from the roots of Cynanchum
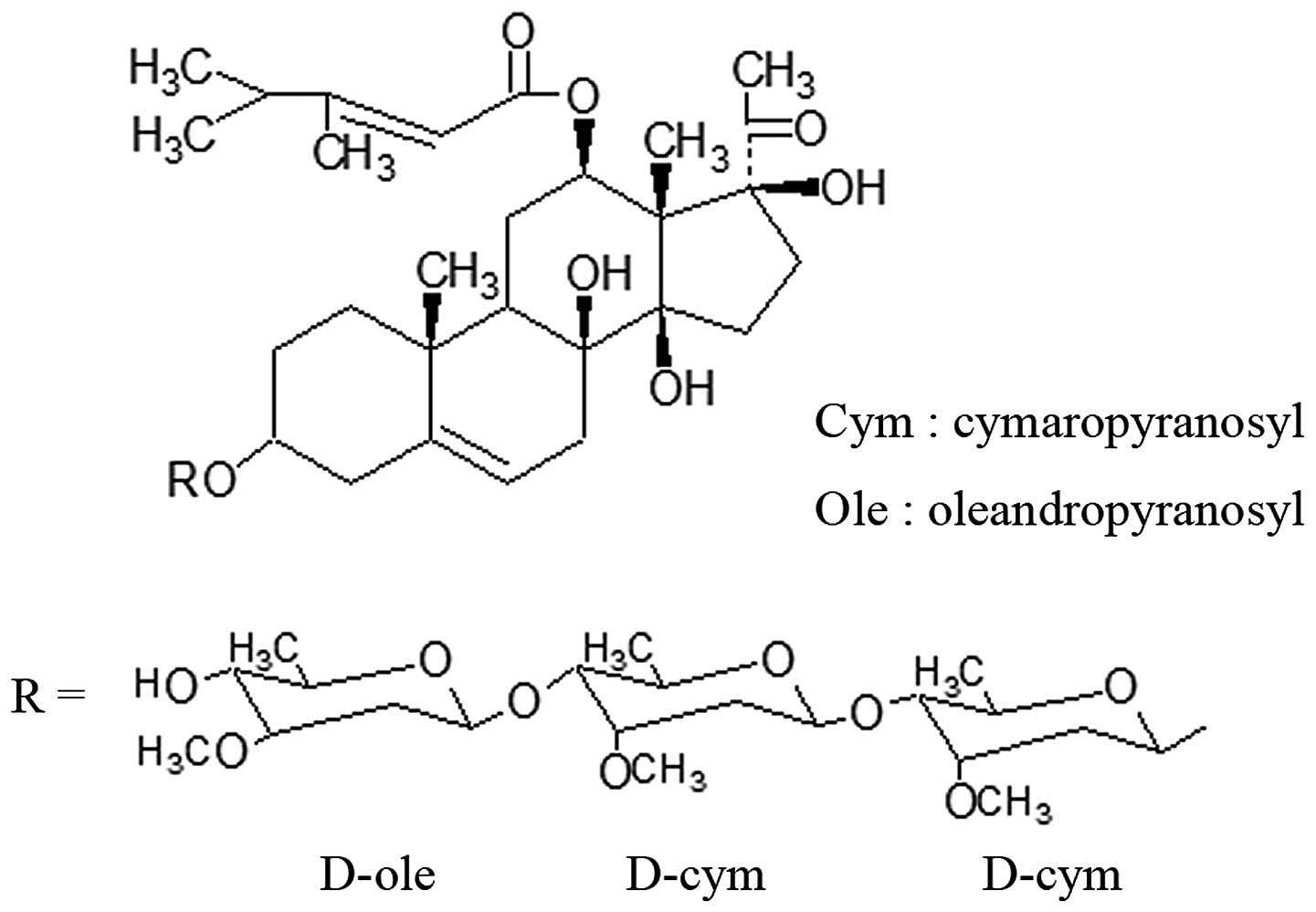
auriculatum Royle ex Wight. CG (Fig. 1) was one of the
C21-steroidal glycosides obtained from Baishouwu and was
first isolated by Warashina et al in 1995 (9). Few of the pharmacological functions of
this compound have been reported. This compound attracted our
attention due to its anticancer activity, which was more marked
than that of the C21-steroidal glycosides previously
reported (8). To further understand
the potential anticancer properties of this compound, CG’s
cytotoxic and apoptosis-inducing activities in several human cancer
cells lines were studied.
Materials and methods
Cell lines
The human gastric cancer (SGC-7901), human colon
cancer (HT-29) and human hepatoma (HEPG-2) cell lines were obtained
from the Institute of Biochemistry and Cell Biology, Shanghai
Institutes for Biological Sciences, Chinese Academy of Sciences
(Shanghai, China). The cells were incubated at 37°C in humidified
air containing 5% CO2.
Concentration-dependent inhibitory effect
assay
The SGC-7901, HT-29 and HEPG-2 cell viability was
determined with an MTT assay. In brief, cells (3×103)
were seeded in 96-well microtiter plates with each well containing
culture medium (100 μl) supplemented with 10% FBS and
incubated at 37°C overnight. The cells were then treated with CG
(2.7, 5.4, 10.8, 21.6 and 43.2 μM) or DMSO (0.1%, negative
control). Following incubation for 48 h, 20 μl of MTT
solution (5 g/l) was added to each well. After 4 h of incubation at
37°C, the medium was discarded, DMSO (150 μl) was added to
each well and the optical density (OD) was measured at 570 nm using
an ELISA Elx8000 plate reader (BioTek, Winooski, VT, USA). The cell
viability was measured using the OD values. The rate of inhibition
was calculated by the following equation: Viability inhibition =
(ODc − ODt) / ODc × 100, where ODc is the optical density of the
control group and ODt is the optical density of the drug-treated
group. Based on the viability inhibition rate, the IC50
(concentration required to inhibit the cell viability by 50%)
values were then calculated with NDST software (BioGuider Medicinal
Technology, China).
Time-dependent inhibitory effect
assay
SGC-7901 cells were plated as described in the
previous section. The cells were left to adhere overnight, then
treated with CG (5.4–21.6 μM) or 0.1% DMSO for 0, 24, 48 and
72 h. At the end of each treatment, the cell viability was
determined using an MTT assay as described previously.
Flow cytometry analysis
Propidium iodide (PI) staining was used to analyze
cell apoptosis and the cell cycle. SGC-7901 cells
(1×106) were seeded in culture flasks prior to drug
treatment. The cells were left to adhere overnight, then treated
with CG (10.8 and 21.6 μM) or DMSO (0.1%) for 24 h. At the
end of the treatment, the cells were trypsinized and washed twice
with ice-cold PBS, then fixed with ice-cold 70% ethanol in PBS at
4°C. The fixed cells were then centrifuged and washed with staining
buffer. After washing, the pellets were treated with 100 μl
RNase A (1 g/l) for 30 min at 37°C. After the incubation, 900
μl staining buffer and 20 μl PI (1 g/l) were added to
each sample and incubated in the dark for 30 min. The samples were
then analyzed with a FACSCalibur flow cytometer (BD Biosciences,
Franklin Lakes, NJ, USA).
Morphological analysis
SGC-7901 cells (1×105) were seeded in
6-well culture plates with each well containing medium (2 ml). The
cells were left to adhere overnight, then treated with CG (10.8 and
21.6 μM) or 0.1% DMSO for 24 h. Following drug treatment,
the medium was discarded and the cells were washed twice with PBS.
The cells were then observed with a light microscope (Olympus,
Tokyo, Japan) or stained with acridine orange (5 μl) for 10
min at room temperature in the dark and observed under a
fluorescence microscope (Olympus).
Determination of cleaved caspase-3
production
Cells (5×106) in medium (20 ml) were
transferred to each flask (75 cm2) prior to drug
treatment. The cells were left to adhere overnight, then treated
with CG (10.8 and 21.6 μM) or DMSO (0.1%) for 24 h. At the
end of the treatment, the cells were harvested and washed twice
with ice-cold PBS, then lysed with lysis buffer (100 μl).
Proteins for the assay were obtained by collecting the supernatant
and were used for the determination of cleaved caspase-3 by sodium
dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and
western blot analysis. The procedure was as follows: The protein
concentration in the supernatant was determined using a BCA protein
assay kit (KeyGen, Nanjing, China) with albumin as the standard.
Equal amounts of proteins (60 μg) from each group were
loaded onto SDS-PAGE gels (12.5%) and electrophoresed to separate
the proteins. The proteins from the gels were transferred onto
nitrocellulose membranes. The membranes were blocked with non-fat
milk (5%) in TBS (10 mM Tris and 100 mM NaCl) for 1 h and probed
with primary antibodies (Santa Cruz Biotechnology, Santa Cruz, CA,
USA) against cleaved caspase-3 and β-actin, followed by an
appropriate horseradish peroxidase-conjugated secondary antibody
(Zhongshan Bio-Tech Co., Ltd., Zhongshan, China) and ECL
detection.
Colorimetric caspase-3 activity
assays
Caspase-3 activity was measured using a colorimetric
assay kit (KeyGen) according to the the manufacturer’s
instructions. In brief, the cells were plated and treated as
described for the determination of cleaved caspase-3 production. At
the end of the treatment, the cells were harvested, washed twice
with ice-cold PBS and lysed for 60 min on ice in the lysis buffer
provided in the kit. The proteins were collected by centrifuging at
10,000 × g for 1 min. The protein concentration in the supernatant
was determined using a BCA protein assay kit (KeyGen) and samples
were diluted to a concentration of 2 g/l using lysis buffer.
Samples containing 50 μg of proteins in lysis buffer (100
μl) were added to the reaction buffer and caspase-3
substrates to measure the caspase-3 activity. The samples were
incubated at 37°C for 4 h. The absorbance density was measured
using a spectrophotometer (Amersham, Piscataway, NJ, USA) at 400
nm.
Statistical analysis
The data are expressed as the mean ± SD. Values were
analyzed with SPSS 16.0 software for Windows and the statistical
significance of differences among the values was evaluated by
one-way ANOVA. P<0.05 was considered to indicate a statistically
significant difference.
Results
Concentration-dependent inhibitory effect
of CG on three human cancer cell lines
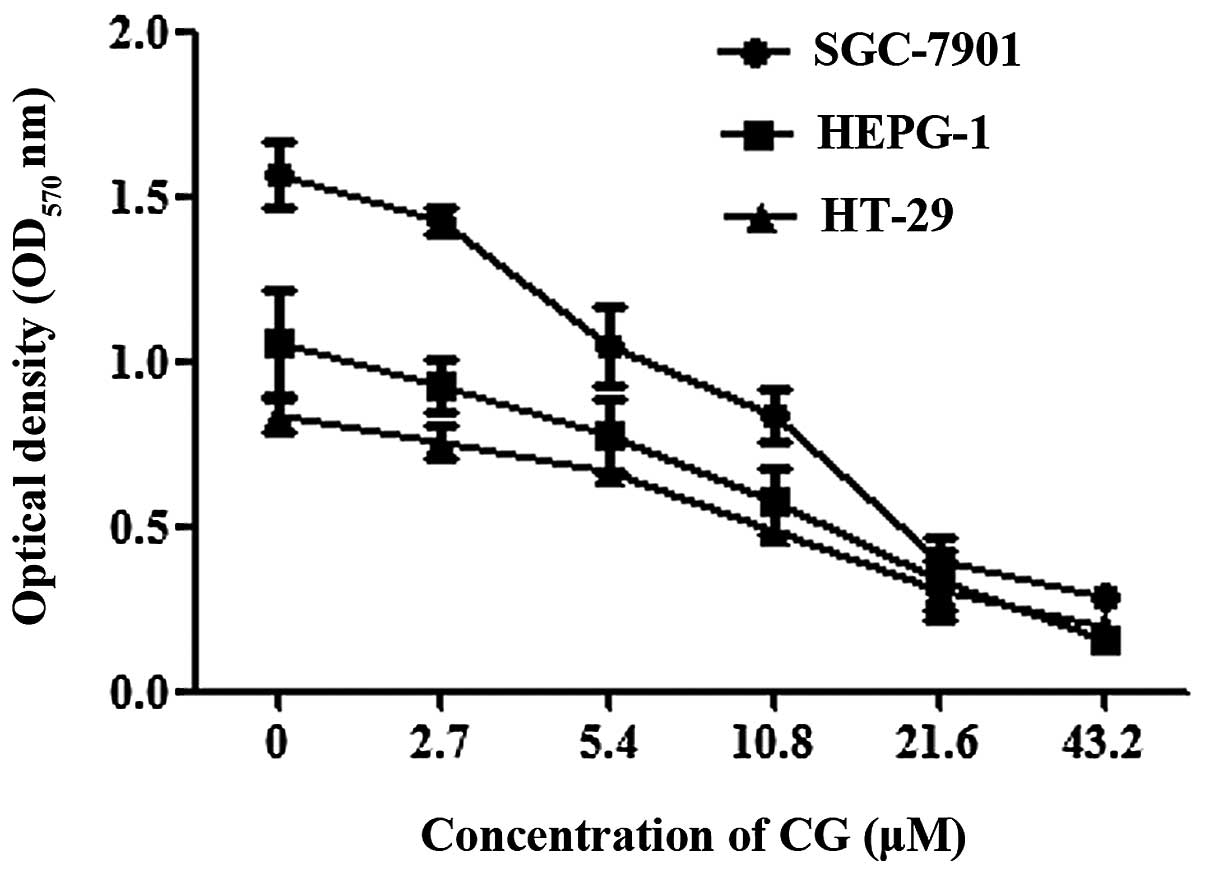
The cytotoxicity of CG against three human
cancer-cell lines; gastric cancer cell line (SGC-7901), colon
cancer cell line (HT-29) and hepatoma cell line (HEPG-2), was
determined. The cell viability was evaluated by measuring the
mitochondrial metabolic activity of the cells using MTT assays. A
concentration-dependent decrease in optical density at 570 nm
(OD570nm) was observed following CG treatment in all
three cell lines (Fig. 2). The cell
viability inhibition rates were also calculated. Treatment with CG
(2.7 43.2 μM) for 48 h resulted in 11.9 to 85.3% decrease in
cell viability in HEPG 2 cells, 10 to 75.9% decrease in HT29 cells
and 9 to 81.4% decrease in SGC 7901 cells. The IC50
values of CG in the HEPG 2, HT 29 and SGC 7901 cell lines are 12.2,
16.4 and 12.6 μM, respectively (Table I).
 | Table IIC50 values of CG against
three human cancer cell lines (mean ± SD, n=3). |
Table I
IC50 values of CG against
three human cancer cell lines (mean ± SD, n=3).
| Cancer cell line | Cell type | IC50
(μM) |
|---|
| HEPG-2 | Liver | 12.2±0.6 |
| HT-29 | Colon | 16.4±2.1 |
| SGC-7901 | Stomach | 12.6±0.5 |
Time-dependent inhibitory effect of CG on
SGC-7901 cells
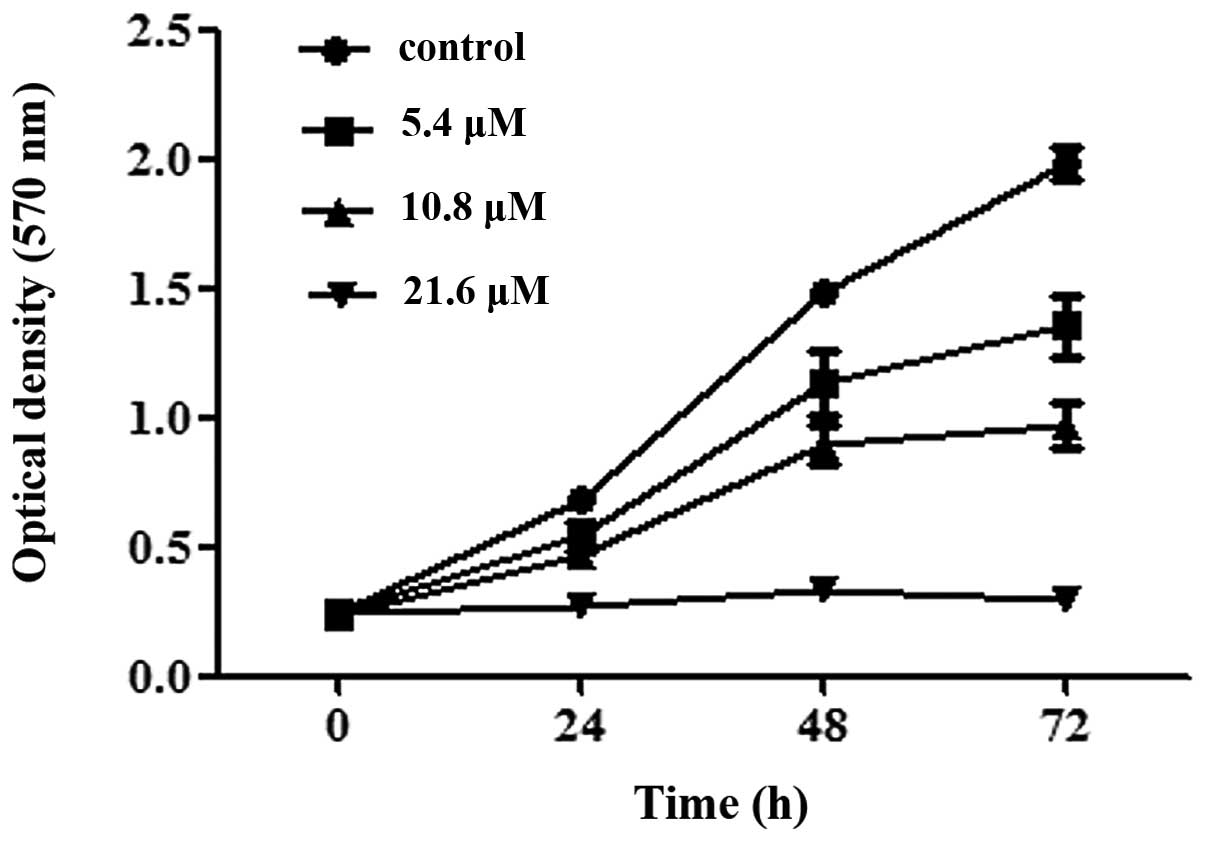
To further understand the time-dependent inhibitory
effect of CG, SGC-7901 cells were treated with CG (5.4–21.6
μM) for 24, 48 and 72 h and an MTT assay was used to
evaluate the cell viability. As shown in Fig. 3, the untreated SGC-7901 cells grew
in an unrestrained manner and the OD570 nm increased
noticeably with the culture time. However, in the SGC-7901 cells
treated with CG, the OD570 nm increased slowly. At 21.6
μM, the OD570 nm did not increase with the
culture time and the cell viability was inhibited by >50%
compared with the control, even when the SGC-7901 cells were only
treated for 24 h.
Cell cycle distribution and apoptosis
rate in SGC-7901 cells
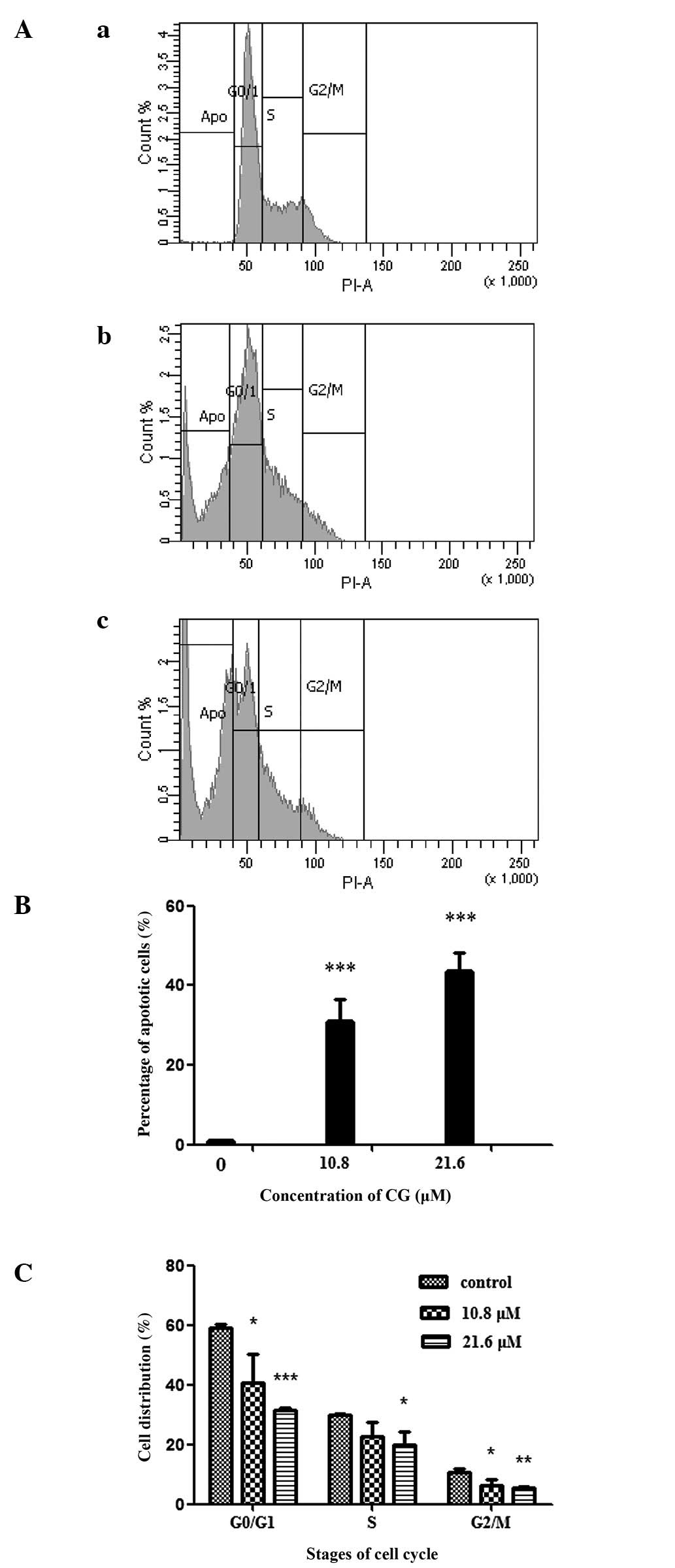
In order to gain an improved understanding of the
cell growth inhibition mechanism, the effects of CG on the cell
cycle distribution and apoptosis rate of SGC-7901 cells were
investigated. PI, a fluorescent dye, was used to stain the nuclear
chromatin and flow cytometry was used to analyze cell apoptosis and
the cell cycle according to the content of the cell chromatin. The
total analyzed cells were separated into four groups; apoptosis,
G0/G1, S and G2/M with regard to
the distribution of DNA content. The gray area in Fig. 4A denotes the number of the cells in
each group. As shown in Fig. 4,
after the cells were treated with CG at 10.8 and 21.6 μM for
24 h, the percentage of apoptotic cells increased to 30.4 and 43.2%
respectively (P<0.001) while the cells in the
G0/G1, S and G2/M phases decreased
(P<0.05).
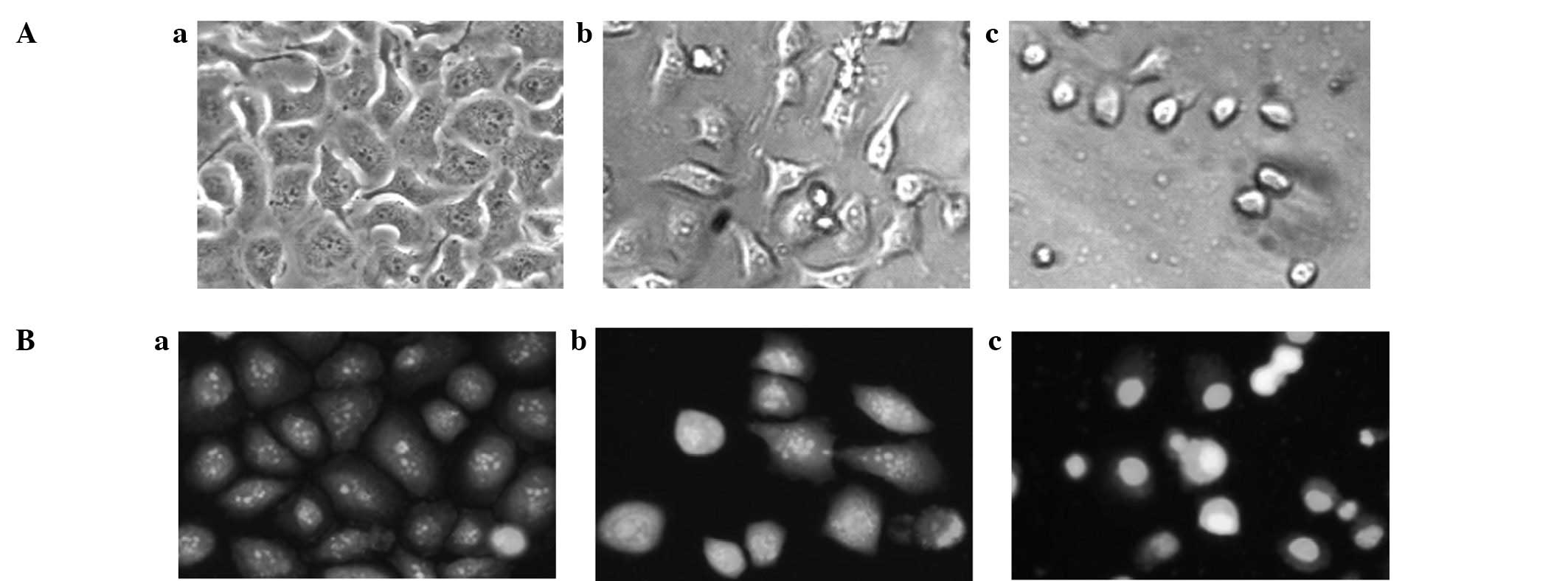
Morphological changes in SGC-7901
cells
To confirm the apoptosis-inducing properties of CG,
the morphological changes in SGC-7901 cells were studied. Under the
light microscope, it was observed that the untreated SGC-7901 cells
grew in an unrestricted manner with tight contact between
neighboring cells. After being treated with CG (10.8 μM) for
24 h, the gaps between the SGC-7901 cells became larger and a
number of the cells shrank and became round in shape. When treated
with CG at 21.6 μM, the cell morphology changed
significantly and the majority of the cells became round in shape
(Fig. 5A). To observe the nuclear
morphological changes, acridine orange, a fluorescent dye which
binds to nuclear chromatin, was used. Under the fluorescence
microscope, it was observed that the fluorescent dye was
distributed evenly in the nuclei of SGC-7901 cells in the control
group (Fig. 5Ba). When the cells
were exposed to CG (10.8 and 21.6 μM) for 24 h, clear
apoptotic characteristics, such as chromatin condensation, nuclear
fragmentation and apoptotic bodies, were observed in numerous
cells. The morphological changes became more distinct when the
concentration of CG was increased (Fig.
5Bb and c).
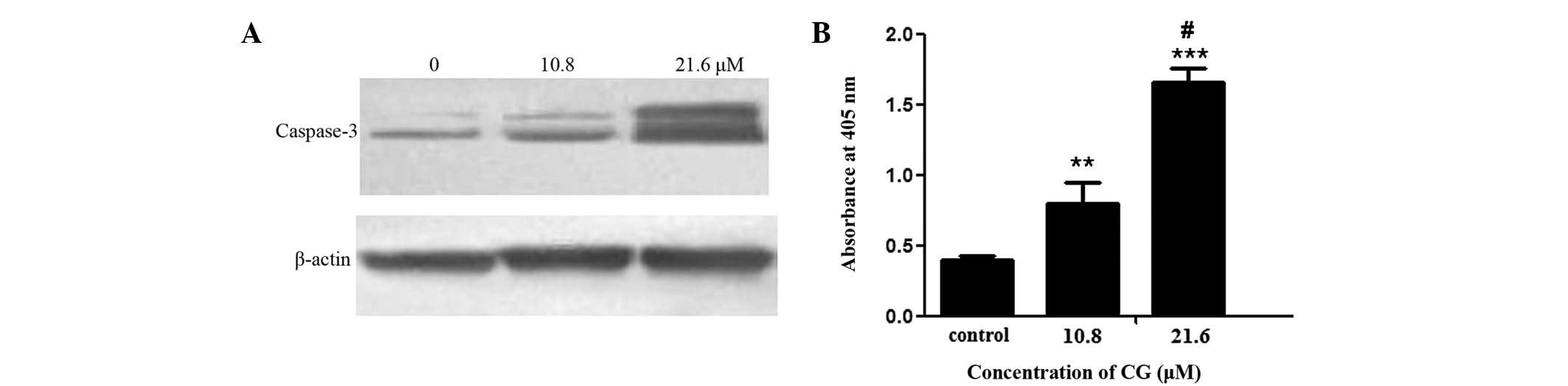
Caspase-3 acivation in SGC-7901
cells
To further demonstrate the apoptosis-inducing effect
of CG, the activation of protease caspase-3, which is activated in
the classic apoptosis pathway and used as a marker of apoptosis
induction (10), was studied. The
results showed that CG (10.8 and 21.6 μM) treatment for 24 h
increased the production of cleaved caspase-3 in a
concentration-dependent manner. As shown in Fig. 6A, the western blot analysis results
revealed two bands at 19 and 17 kDa, which were the bands for
cleaved caspase-3. The cleaved caspase-3 bands of the control were
extremely faint. CG (10.8 and 21.6 μM) treatment for 24 h
increased the intensity of the two bands, particularly at 21.6
μM. To further confirm the activation of caspase-3, a
colorimetric assay kit (KeyGen) was used to examine the activity of
caspase-3. The enzymatic activity of caspase-3 was quantified by
measuring chromophores obtained from the cleaved substrates. As
shown in Fig. 6B, after treatment
with CG (10.8 and 21.6 μM) for 24 h, the absorbance at 405
nm, denoting the caspase-3 activity, increased significantly
(P<0.01). At 21.6 μM, the activity of caspase-3 was
increased ∼3-fold.
Discussion
In our previous study, we isolated four new
C21-steroidal glycosides from the roots of Cynanchum
auriculatum and reported the anticancer activity of
auriculoside A (AA) which was the most active of the four (8,11,12).
In the present study, CG, a C21-steroidal glycoside
first isolated by Warashina et al in 1995, demonstrated
greater cytotoxic effects than AA. The IC50 values of
CG, which ranged between 12.2 and 16.4 μM, were lower than
those of AA, which ranged between 23.2 and 36.7 μM (8). Furthermore, the IC50 values
of CG were lower than the C21-steroidal glycosides
reported by other studies. For example, the IC50 value
of caudatin, a C21-steroidal aglycone from the roots of
Cynanchum auriculatum was reported to be 84.51 μM at
48 h in HEPG-2 (13). The great
difference in the inhibitory effects of these
C21-steroidal glycosides in similar cancer cells
suggests that there may be certain rules governing the association
between the structure and activity of this type of compound.
Apoptosis induction is a key event that is the
target of numerous chemopreventive agents (13,14).
C21-steroidal glycosides also show a clear
apoptosis-inducing effect (6,8,13).
Accordingly, the apoptosis-inducing properties of CG were
investigated in the present study. The results showed that CG
induced cancer cell apoptosis at a lower concentration than a
number of other C21-steroidal glycosides (6,8,13). As
analyzed by flow cytometry, 21.6 μM CG treatment for only 24
h induces apoptosis in >40% cells and this is consistent with
its strong cytotoxicity in cancer cells in vitro.
Morphological changes are clear evidence of apoptosis. Apoptotic
cells are characterized by distinct morphological features,
including cell shrinkage and loss of contact with neighboring
cells, chromatin condensation, nuclear fragmentation and apoptotic
body formation. To demonstrate the apoptosis-inducing effect of CG,
the cell outline and nucleus were observed under light and
fluorescence microscopes. It was revealed that SGC-7901 cells
treated with CG exhibited typical apoptotic morphological features.
Usually in apoptosis, caspase-3 is activated and this is often used
as a marker of apoptosis induction (15). Activated caspase-3 is cleaved into
two segments (cleaved caspase-3) with molecular weights of 17 and
19 kDa. Cleaved caspase-3 is able to activate deoxyribonuclease,
leading to DNA fragmentation and apoptotic cell death (16). In the present study, the production
of cleaved caspase-3 and the activity of caspase-3 were also
investigated to assess caspase-3 activation. The results showed
that the production of the two segments of cleaved caspase-3 and
the activity of caspase-3 increased following CG treatment for 24 h
particularly at 21.6 μM. Thus the apoptosis-inducing effect
of CG was further demonstrated.
In conclusion, the present study demonstrated the
cytotoxic and apoptosis-inducing properties of CG and showed that
CG is a potent anticancer C21-steroidal glycoside from
the roots of Cynanchum auriculatum. This study also
emphasized the importance of comparing the pharmacological
properties between the various C21-steroidal glycosides.
In future research, emphasis should be placed on identifying the
rules governing the association between the structure and activity
of this type of compound.
Acknowledgements
This study was supported by the
Zhejiang Provincial Health Department of China (grant No.
2011KYB055).
References
|
1
|
Lee MK, Yeo H, Kim J and Kim YC:
Protection of rat hepatocytes exposed to CCl4 in-vitro
by cynandione A, a biacetophenone from Cynanchum wilfordii.
J Pharm Pharmacol. 52:341–345. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lee MK, Yeo H, Kim J, Markelonis GJ, Oh TH
and Kim YC: Cynandione A from Cynanchum wilfordii protects
cultured cortical neurons from toxicity induced by
H2O2, L-glutamate, and kainate. Neurosci Res.
59:259–264. 2000.
|
|
3
|
Shan L, Liu RH, Shen YH, et al:
Gastroprotective effect of a traditional Chinese herbal drug
‘Baishouwu’ on experimental gastric lesions in rats. J
Ethnopharmacol. 107:389–394. 2006.
|
|
4
|
Shan L, Zhang WD, Zhang C, Liu RH, Su J
and Zhou Y: Antitumor activity of crude extract and fractions from
root tuber of Cynanchum auriculatum Royle ex Wight.
Phytother Res. 19:259–261. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu K, Chen F and Zhang H: Antitumor
effects by Wilfoside C3N treatment in ECA109 cells. Anticancer
Drugs. 21:625–631. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Peng YR, Ding YF, Wei YJ, Shu B, Li YB and
Liu XD: Caudatin-2, 6-dideoxy-3-O-methy-β-D-cymaropyranoside 1
induced apoptosis through caspase 3-dependent pathway in human
hepatoma cell line SMMC7721. Phytother Res. 25:631–637. 2011.
|
|
7
|
Li Y, Zhang J, Gu X, Peng Y, Huang W and
Qian S: Two new cytotoxic pregnane glycosides from Cynanchum
auriculatum. Planta Med. 74:551–554. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang R, Liu Y, Wang Y, Ye Y and Li X:
Cytotoxic and apoptosis-inducing properties of auriculoside A in
tumor cells. Chem Biodivers. 4:887–892. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Warashina T and Noro T: Steroidal
glycosides from roots of Cynanchum caudatum M. Chem Pharm
Bull (Tokyo). 43:977–982. 1995. View Article : Google Scholar
|
|
10
|
Zhang RS, Ye YP, Shen YM and Liang HL:
Studies on the cytotoxic constituents of Cynanchum
auriculatum Royle ex Wight. Yao Xue Xue Bao. 35:431–437.
2000.
|
|
11
|
Zhang RS, Ye YP, Shen YM and Liang HL: Two
new cytotoxic C-21 steroidal glucosides from the root of
Cynanchum auriculatum. Tetrahedron. 56:3875–3879. 2000.
View Article : Google Scholar
|
|
12
|
Fei HR, Chen HL, Xiao T, Chen G and Wang
FZ: Caudatin induces cell cycle arrest and caspase-dependent
apoptosis in HepG2 cell. Mol Biol Rep. 39:131–138. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sun SY, Hail NJ and Lotan R: Apoptosis as
a novel target for cancer chemoprevention. J Natl Cancer Inst.
96:662–672. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kasibhatla S and Tseng B: Why target
apoptosis in cancer treatment. Mol Cancer Ther. 2:573–580.
2003.PubMed/NCBI
|
|
15
|
Nicholson DW and Thornberry NA: Caspases:
killer proteases. Trends Biochem Sci. 22:299–306. 1997. View Article : Google Scholar
|
|
16
|
Ghobrial IM, Witzig TE and Adjei AA:
Targeting apoptosis pathways in cancer therapy. CA Cancer J Clin.
55:178–194. 2005. View Article : Google Scholar : PubMed/NCBI
|