Introduction
As epithelial ovarian cancer is difficult to
diagnose during the early stages, treatment is often administered
later than would be ideal. Epithelial ovarian cancer is the leading
cause of gynecological cancer mortalities and poses a serious
threat to female health. DNA methylation is a covalent modification
of the cytosine residue in a CpG dinucleotide. It is a natural
means of controlling gene transcription and does not require any
changes to the DNA sequence. DNA methylation is important in the
occurrence and development of epithelial ovarian cancer (1,2). The
WWOX gene is a newly discovered tumor suppressor gene. It was first
isolated and identified using shotgun sequencing, by Bednarek et
al in 2000 (3). Studies have
shown that the primary site of WWOX gene transcription is rich in
CpG islands (4). Therefore,
methylation may be the key mechanism behind defects in expression.
Abnormal methylation of CpG islands in the promoter region of the
WWOX gene have been shown to be closely associated with the
occurrence and development of breast cancer (5,6). The
present study aimed to further explore the correlation between the
abnormal methylation of the WWOX gene and epithelial ovarian
cancer.
Materials and methods
Materials
The HO-8910 human ovarian cancer cell line was
obtained from the Department of Obstetrics and Gynecology
Laboratory of the Affiliated Hospital of Xuzhou Medical College
(Xuzhou, China) and the RPMI-1640 medium was purchased from Hyclone
(South Logan, UT, USA). Fetal bovine serum was purchased from
Hangzhou Sijiqing Biology Engineering Materials Co., Ltd.
(Hangzhou, China). 5-Aza-2′-deoxycytidine (5-Aza-CdR) and MTT were
produced by Sigma (St Louis, MO, USA) and the Wizard DNA Clean-up
System kits were obtained from Promega (Madison, WI, USA). The Taq
DNA polymerases systems were obtained from Qiagen (Hilden,
Germany). The oligonucleotide primer was synthesized by Shanghai
Shenggong Biological Engineering Co., Ltd. (Shanghai, China). The
Transwell chamber and WWOX primary antibody were provided by
Chemicon (Temecula, CA, USA). BALB/c female nude mice, aged 4–6
weeks, were supplied by the Shanghai Laboratory Animal Center,
Chinese Academy of Sciences (Shanghai, China). This study was
approved by the ethics committee of Xuzhou Medical College
(Jiangsu, China).
Cell culture
The HO-8910 cell line was maintained in RPMI-1640
medium (including 70 U/ml penicillin and 70 μg/ml streptomycin)
supplemented with 10% fetal bovine serum. The cells were
subcultured in a humidified atmosphere of 5% CO2 at 37°C
in an airtight incubator. Logarithm vegetal period cells were added
to the fluid, which included 5.0 μmol/l 5-Aza-CdR, to culture for
24 h. The solution was then replaced by fresh culture fluid
containing the same concentration of 5-Aza-CdR. Subsequent to being
cultured for 3 days, the solution was replaced by a fresh culture
medium that did not contain the drug. The cells were allowed to
incubate and the experiment was performed 5 days later.
WWOX gene methylation state detected by
methylation-specific PCR (MSP)
The cells were divided into two groups. Protease
K-phenol extraction was used to extract the total DNA, and an
ultraviolet spectrophotometer was used to determine the quantity
and purity of the DNA. Agarose gel electrophoresis was performed to
determine the DNA integrity. The ultraviolet spectrophotometer
quantitatively adjusted the final concentration of DNA to 0.1 g/l,
and the DNA was stored at −20°C. Subsequently, DNA modification,
purification and PCR were conducted. The purification steps of the
experiment were performed in accordance with the Wizard DNA
Clean-up System kit instructions (Promega). PCR was performed using
a Qiagen reaction system. The 20-μl reaction system consisted of 2
μl 10X PCR buffer (Shanghai Shenggong Biological Engineering Co.,
Ltd.), 0.4 μl dNTP, 0.4 μl upstream and downstream primers, 0.1 μl
DNA Taq enzyme, 15.7 μl de-ionized water and 1 μl template. The PCR
and MSP amplification conditions were 15 min at 95°C; 40 sec at
95°C, 40 sec at 60°C and 40 sec at 70°C, for 35 cycles; and 10 min
at 72°C. An agarose gel electrophoresis was performed and a gel
imaging system was used for the scanning analysis. The primer
sequences for the methylated WWOX gene were forward,
5′-TATGGGCGTCGTTTTTTTAGTT-3′ and reverse,
5′-CAATCTCCGCAATATCGCGACA-3′. The sequences of the unmethylated
primers were forward, 5′-TATGGGTGTTGTTTTTTTAHTT-3′ and reverse,
5′-CAATCTCCACAATATCACAACA-3′. The product size was 347 bp.
WWOX protein expression detected by
western blot analysis
The two groups of cells that were in the growth
period were placed in 200 μl cell lysis solution (Shanghai
Shenggong Biological Engineering Co., Ltd.) for cracking on ice,
and the bicinchoninic acid (BCA) method was used to detect the
protein concentration. In order to transfer the proteins to a
nitrocellulose membrane, sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) was performed. The membranes were
incubated with with 5% skimmed milk powder for 60 min, prior to the
addition of 1:1,000 anti-WWOX antibody (rabbit anti-human) and
incubation overnight at 4°C. The cells were reacted with
horseradish peroxidase-labeled sheep anti-rabbit antibody as a
secondary antibody (1:10,000; Shanghai Shenggong Biological
Engineering Co., Ltd.). This reaction took place at room
temperature and was allowed to continue for 2 h. Subsequent to
adding enhanced chemiluminescence (ECL) agent (Hangzhou Sijiqing
Biology Engineering Materials Co., Ltd.), the mixtures were placed
in an anechoic chamber for exposure imaging with highly sensitive
X-ray film. The gray value of the target and β-actin protein bands
were measured and the ratio of the target band to β-actin indicated
the relative expression level of the target protein.
Proliferation of the HO-8910 cell line
detected by MTT
Two groups of HO-8910 cells were obtained in the
growth period, and then plated into 96-well plates at a density of
1.5×104/well and cultured for 1–6 days. Following this,
20 μl MTT working fluid was added to each well and incubated in
CO2 at 37°C for 4 h. Dimethyl sulfoxide was added to
terminate the reaction and the absorbance value was detected at 490
nm. Cellular growth curves were constructed based on these
results.
Invasion ability of HO-8910 cells
detected by an invasion assay in vitro
Using a Transwell chamber cell invasion assay in
vitro, an invasion membrane synthesized from matrix adhesive
(Matrigel, Promega) was placed between the upper and lower areas of
the Transwell chamber. A single HO-8910 cell suspension was
vaccinated on the invasion membrane at 200 μl per hole, containing
~105 cells, and cultured at 37°C in 5% CO2
for 12 h. The cells and matrix adhesive were wiped on the membrane,
which was fixed and stained with H&E. The number of
transmembrane cells was calculated using a microscope. Each group
of cells had three duplicate wells and the experiment was repeated
three times.
Flow cytometry to detect the HO-8910 cell
cycle
The cells that were in the growth period were
divided into two groups and digested using 0.25% trypsin.
Physiological saline was used for conventional washing. The cells
were centrifuged and fixed in 70% ethanol overnight prior to the
addition of 100 μl phosphate buffer liquid to disperse them into
single cell suspensions. The cells were analyzed using flow
cytometry.
In vivo experiment
Animal groups
A total of 20 BALB/c 4- to 6-week-old female nude
mice were randomly divided into two groups, a control group and a
5-Aza-CdR-treated group, each containing 10 mice. They were
maintained at a constant temperature (25–27°C) and humidity
(45–50%), with fresh air and high dust-removing sterilizing and
pathogen-free conditions. The mice were allowed free access to food
and sterilized water.
Animal vaccination and treatment
The two groups of cells in the growth period were
digested to form single cell suspensions with a density of
4×107/ml. The cells were inoculated into the abdominal
cavities of the nude mice at a volume of 0.5 ml each. The nude mice
were observed and the date of death was recorded. A necropsy was
performed on the abdominal cavity immediately following death. The
volume of abdominal fluid was calculated, and the number, weight
and maximum diameter of the tumors and the abdominal viscera
metastasis were observed.
Western blot analysis to detect WWOX
protein expression in the tumor tissue
The tumor tissues from the nude mice in the
5-Aza-CdR-treated and control groups were obtained and weighed. The
tumors were then ground with lysis buffer to extract the total
protein. These were quantified using the BCA method. SDS-PAGE was
performed using a 30-μg protein sample from each group to transfer
the proteins to a nitrocellulose membrane. The membranes were
incubated with 5% skimmed milk powder for 60 min at room
temperature, and then 1:1,000 anti-WWOX antibody (rabbit
anti-human) was added and the membranes were incubated overnight at
4°C. The mixture was then allowed to react with horseradish
peroxidase-labeled sheep anti-rabbit antibody as a secondary
antibody (1:10,000). This reaction took place at room temperature
for 2 h. Subsequent to adding ECL agent, the mixture was placed in
the anechoic chamber for exposure imaging with a highly sensitive
X-ray film.
Statistical analysis
All data are expressed as the group mean ± standard
deviation, and were processed using SPSS 13.0 statistical software
(SPSS, Inc., Chicago, IL, USA). The statistical differences between
the groups of data were assessed using the Student’s t-test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
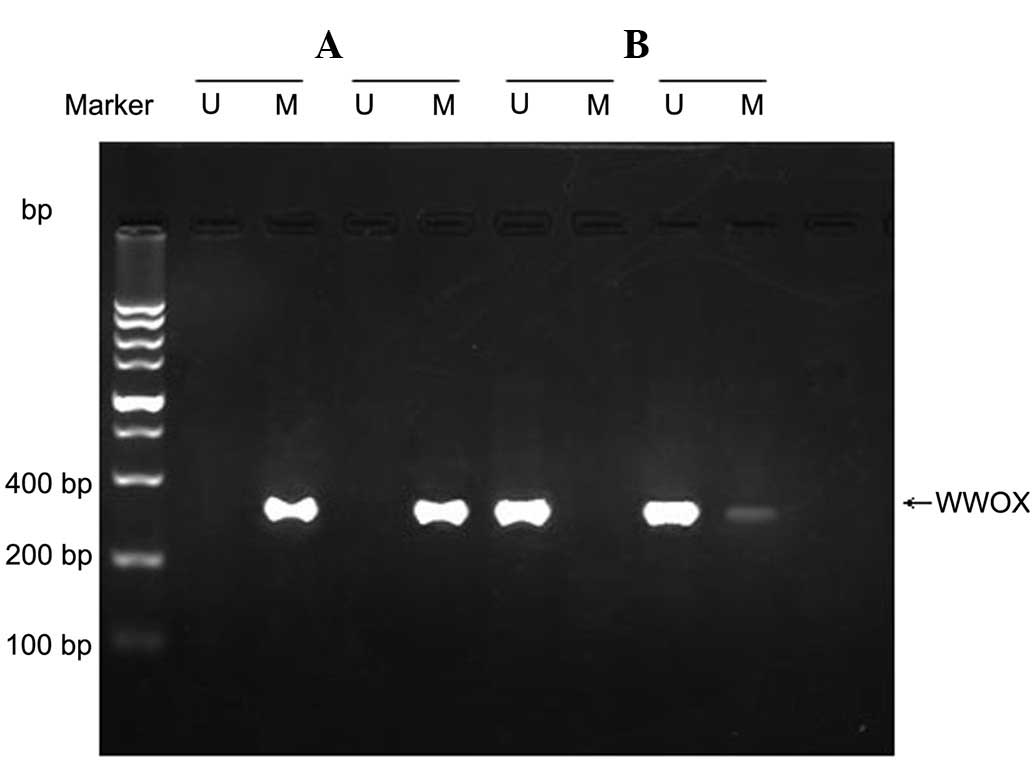
WWOX gene methylation in HO-8910
cells
The WWOX gene was in a state of methylation in the
HO-8910 ovarian cancer cell line. Unmethylated and partially
methylated samples were identified following treatment with
5-Aza-CdR (Fig. 1).
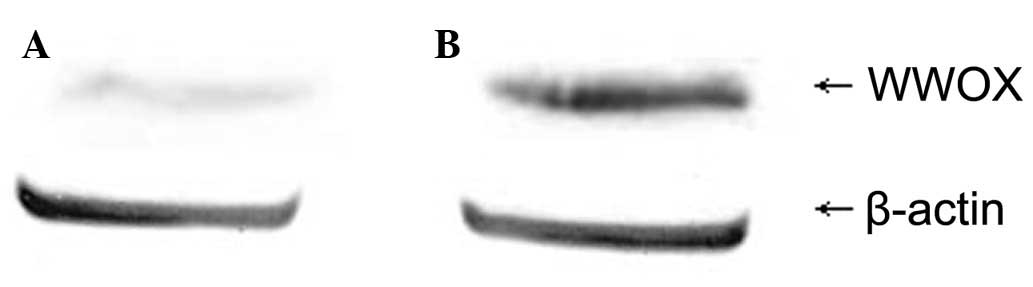
Changes in WWOX expression in
5-Aza-CdR-treated HO-8910 cells
Western blot analysis revealed that the expression
level of the WWOX protein in the 5-Aza-CdR-treated group was
0.71±0.023, whereas that of the control group was 0.13±0.012. The
difference was identified to be statistically significant
(P<0.05; Fig. 2).
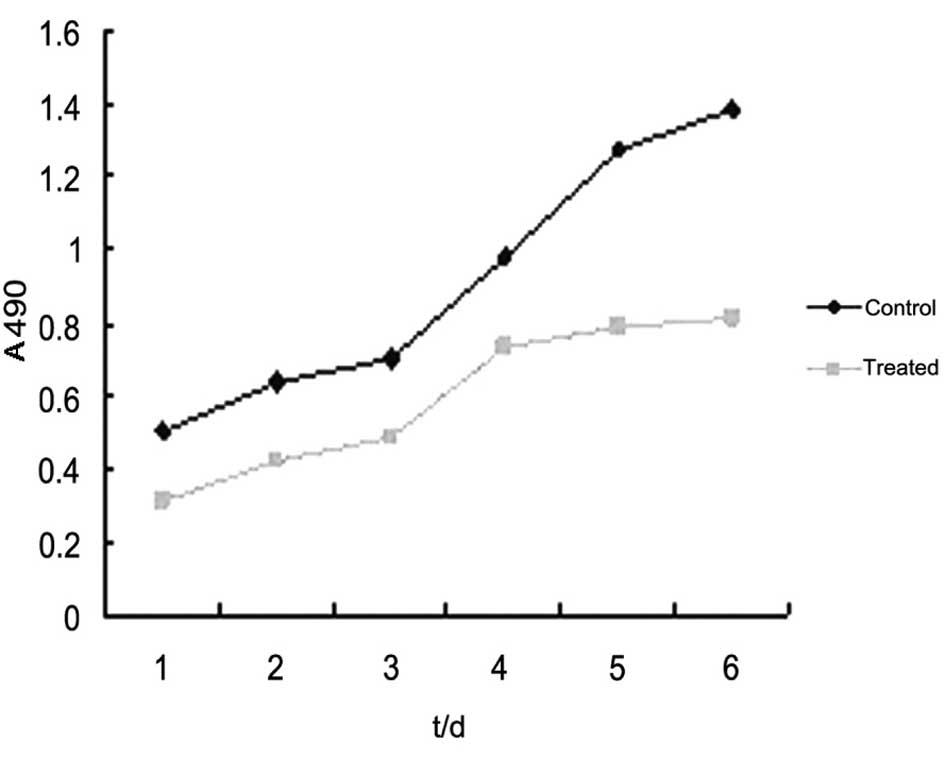
Changes in the proliferation ability of
5-Aza-CdR-treated HO-8910 cells
The MTT assay showed that subsequent to being
cultured for 1–6 days, the absorbance values of cells in the
5-Aza-CdR-treated group were 0.312±0.013, 0.423±0.015, 0.488±0.022,
0.734±0.031, 0.791±0.019 and 0.814±0.018. The growth rate at each
time point was significantly lower than that of the control group
at the corresponding time (P<0.05; Fig. 3).
Changes in the invasion ability of
5-Aza-CdR-treated HO-8910 cells
The in vitro invasion assay showed that the
number of transmembrane cells in the 5-Aza-CdR-treated group was
92.2±4.7, while 172.1±5.2 transmembrane cells were observed in the
control group. This difference was identified to be statistically
significant (P<0.05, data not shown).
Changes in the cell cycle of
5-Aza-CdR-treated HO-8910 cells
Flow cytometry revealed that the cells in the
5-Aza-CdR-treated group apparently arrested in the
G0/G1 phase, and that the number of cells in
the S phase was significantly lower than that in the control group
(P<0.05; Table I).
 | Table IChanges to the 5-Aza-CdR-treated
HO-8910 cellular cycle. |
Table I
Changes to the 5-Aza-CdR-treated
HO-8910 cellular cycle.
| Group |
G0/G1 | S | G2/M |
|---|
| 5Aza-CdR-treated | 67.13±0.26 | 19.56±1.36 | 13.45±0.38 |
| Control | 21.52±0.37 | 47.09±1.03 | 31.06±0.61 |
Changes in the tumorigenic ability of
5-Aza-CdR-treated HO-8910 cells in nude mice
The necropsy revealed that transplanted tumors were
present in the abdominal cavities of all the nude mice. The tumor
formation rate was 100% in the treated and control groups
(P>0.05). Each of the nude mice in the control group contained
bloody ascites in the abdominal cavity, while two nude mice in the
5-Aza-CdR-treated group showed no significant bloody ascites. The
two groups of nude mice had tumor nodules of various sizes
scattered in numerous parts of the peritoneum, epiploon, diaphragm,
bowel and the surface of the mesentery. Metastasis was observed in
the liver, but not in the heart, kidneys, lungs or uterine
appendages in the two groups. A comparison between the control and
5-Aza-CdR-treated group revealed significant differences in the
survival time, number of metastases, metastatic foci weight,
maximum tumor diameter and abdominal volume (P<0.05; Table II).
| Table IIComparison of survival time, number of
metastases, tumor weight, maximum diameter and abdominal volume
between the two groups of nude mice. |
Table II
Comparison of survival time, number of
metastases, tumor weight, maximum diameter and abdominal volume
between the two groups of nude mice.
| Group | No. | Survival time
(days) | No. of
metastases | Tumor weight (g) | Maximum diameter
(mm) | Abdominal volume
(ml) |
|---|
|
5-Aza-CdR-treated | 10 | 58.3±3.6 | 33±5 | 4.73±0.52 | 4.9±0.7 | 3.75±0.62 |
| Control | 10 | 37.4±3.1 | 58±6 | 9.02±0.17 | 9.2±0.4 | 7.53±0.22 |
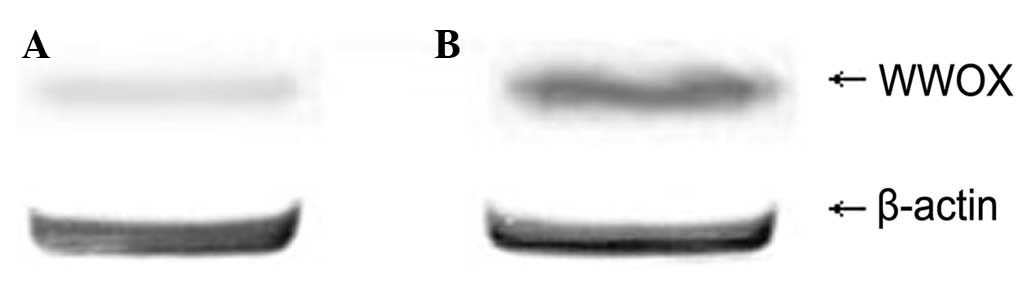
Western blot analysis showed the expression level of
WWOX protein in the tumor issue of the nude mice in the
experimental group to be 0.65±0.031, while that of the control
group was 0.25±0.047. The difference was statistically significant
(P<0.05; Fig. 4).
Discussion
DNA methylation is the only known natural chemical
modification of DNA in mammals. It is one of the main and most
studied factors in mammalian epigenetics. In DNA methylation,
S-adenosyl-L-methionine supplies the methyl group, which is
transferred to the fifth carbon atom of cytosine to generate
5-methylcytosine under the action of the methyltransferase. This
may affect gene expression without causing any changes to the gene
sequence. The site of DNA methylation is usually located on the CpG
island in the promoter region of a gene (7). DNA hypomethylation may promote the
expression of tumor suppressor genes, while DNA hypermethylation
may decrease or stop the expression of tumor suppressor genes and
cause the tumor suppressor genes to lose function. This may result
in unrestricted cell growth and ultimately lead to tumorigenesis
(8–10).
Epigenetic changes are different from genetic
alterations in several ways. One of these variations is that
epigenetic changes are able to be reversed. Therefore, the normal
regulation of cells is restored by recovering the expression of
unmutated genes to achieve the purpose of the treatment. Normal
human cell genes are not controlled by CpG island methylation.
Therefore, the inhibition of methylation does not affect the
expression of genes in normal cells. Genes in static states due to
methylation are sensitive to DNA methylation inhibitors and to the
abnormal expression of corresponding HLA molecules, allowing gene
therapy to recover the activity (11–16).
At present, the primary drug for reversing DNA methylation is
5-Aza-CdR. The methyltransferase-specific inhibitor 5-Aza-CdR is a
pyrimidine analog whose mechanism is to combine with DNA during DNA
replication, form covalent complexes with DNMT1, inhibit the
enzyme’s methyl transfer activity, produce low-methyl annihilator
chains and reduce the hypermethylation of the gene promoter region
to recover gene activity.
The present study revealed that the WWOX gene was in
a state of hypermethylation in the HO-8910 ovarian cancer cell
line. The cells were in a demethylated state following treatment
with 5-Aza-CdR. Due to demethylation, WWOX gene expression
increased. A cell growth curve experiment showed a decrease in the
growth rate of the HO-8910 cells that were treated with 5-Aza-CdR,
suggesting that 5-Aza-CdR may inactivate the WWOX gene through
aberrant recovery of methylation. The WWOX gene tumor suppressor
function was recovered, which inhibited further growth of the
HO-8910 cells. In vitro invasion ability was a significant
characteristic that was used to measure tumor metastatic potential.
A chamber invasion in vitro experiment showed that the
ability of the HO-8910 cells that were treated with 5-Aza-CdR to
invade other tissues decreased markedly, suggesting that the
invasion and cancer cell movement abilities were reduced. Flow
cytometry showed that the HO-8910 cells were arrested in the S
phase following treatment with 5-Aza-CdR. The number of cells in
the G2/M phases decreased, indicating that the antitumor
effect of the WWOX gene may work by affecting the cell cycle
following demethylation. In the present study, the HO-8910 cells
that were treated with 5-Aza-CdR were also inoculated into the
abdominal cavity of nude mice, and the effects of WWOX
demethylation on the behavior of the HO-8910 cells were studied
further. It was identified that the growth rate of the demethylated
HO-8910 cells in the nude mice was significantly slower than that
of the control group. Tumorigenicity decreased significantly and
the expression levels of the WWOX gene increased in the
experimental group. The WWOX gene was observed to be associated
with the occurrence and development of ovarian cancer; normal
expression of the WWOX gene suppresses the occurrence of ovarian
cancer.
In the present study, the in vivo and in
vitro experiments revealed that aberrant methylation of the
WWOX gene was the main reason for the reduction in gene
transcription and expression, which may be closely associated with
the occurrence and development of ovarian cancer. These results
also indicated that the tumor suppressor activity was recovered by
demethylation of the WWOX gene, which may provide new experimental
evidence for the diagnosis and treatment of ovarian cancer.
References
|
1
|
Barton CA, Hacker NF, Clark SJ and O’Brien
PM: DNA methylation changes in ovarian cancer: implications for
early diagnosis, prognosis and treatment. Gynecol Oncol.
109:129–139. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Houshdaran S, Hawley S, Palmer C, et al:
DNA methylation profiles of ovarian epithelial carcinoma tumors and
cell lines. PLoS One. 5:e93592010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bednarek AK, Laflin KJ, Daniel RL, et al:
WWOX, a novel WW domain-containing protein mapping to human
chromosome 16q23.3–24.1, a region frequently affected in breast
cancer. Cancer Res. 60:2140–2145. 2000.PubMed/NCBI
|
|
4
|
Del Mare S, Salah Z and Aqeilan RI: WWOX:
its genomics, partners, and functions. J Cell Biochem. 108:737–745.
2009.PubMed/NCBI
|
|
5
|
Lewandowska U, Zelazowski M, Seta K, et
al: WWOX, the tumour suppressor gene affected in multiple cancers.
J Physiol Pharmacol. 60:47–56. 2009.PubMed/NCBI
|
|
6
|
Wang X, Chao L, Jin G, et al: Association
between CpG island methylation of the WWOX gene and its expression
in breast cancers. Tumour Biol. 30:8–14. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dehan P, Kustermans G, Guenin S, et al:
DNA methylation and cancer diagnosis: new methods and applications.
Expert Rev Mol Diagn. 9:651–657. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Worthley DL, Whitehall VL, Buttenshaw RL,
et al: DNA methylation within the normal colorectal mucosa is
associated with pathway-specific predisposition to cancer.
Oncogene. 29:1653–1662. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kiran M, Chawla YK and Kaur J: Methylation
profiling of tumor suppressor genes and oncogenes in hepatitis
virus-related hepatocellular carcinoma in northern India. Cancer
Genet Cytogenet. 195:112–119. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sharma G, Mirza S, Parshad R, et al: CpG
hypomethylation of MDR1 gene in tumor and serum of invasive ductal
breast carcinoma patients. Clin Biochem. 43:373–379. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kuroda A, Rauch TA, Todorov I, et al:
Insulin gene expression is regulated by DNA methylation. PLoS One.
4:e69532009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lewandowska J and Bartoszek A: DNA
methylation in cancer development, diagnosis and therapy - multiple
opportunities for genotoxic agents to act as methylome disruptors
or remediators. Mutagenesis. 26:475–487. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gaudet MM, Campan M, Figueroa JD, et al:
DNA hypermethylation of ESR1 and PGR in breast cancer: pathologic
and epidemiologic associations. Cancer Epidemiol Biomarkers Prev.
18:3036–3043. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Weng YI, Huang TH and Yan PS: Methylated
DNA immunoprecipitation and microarray-based analysis: detection of
DNA methylation in breast cancer cell lines. Methods Mol Biol.
590:165–176. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sceusi EL, Loose DS and Wray CJ: Clinical
implications of DNA methylation in hepatocellular carcinoma. HPB
(Oxford). 13:369–376. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Espada J and Esteller M: DNA methylation
and the functional organization of the nuclear compartment. Semin
Cell Dev Biol. 21:238–246. 2010. View Article : Google Scholar : PubMed/NCBI
|