Introduction
Radical surgery or radiotherapy may be curative
therapies for patients with localized prostate cancer, but there
are no effective treatment modalities for the management of
advanced prostate cancer. Androgen deprivation treatment is the
most effective systemic approach for patients with metastatic
disease. Although 80–90% of patients initially respond favorably to
this treatment, they eventually become unresponsive to androgen
deprivation and develop castration-resistant prostate cancer (CRPC)
(1,2). Currently, the combination of docetaxel
and prednisone is considered the standard first-line therapy for
CRPC, but the survival gain from docetaxel chemotherapy is limited
and unsatisfactory (3).
Stromal derived factor-1 (SDF-1) is a member of the
CXC subfamily of chemokines that interacts with the 7 transmembrane
G-protein-coupled receptor CXCR4 (4–6). CXCR4
expression has been reported in at least 23 epithelial, mesenchymal
and hematopoietic cancers, indicating the importance of this
ligand/receptor axis in tumor aggressiveness and metastasis
(7–9). The role of the SDF-1/CXCR4 axis in
prostate cancer has been experimentally demonstrated. It is known
that CXCR4 mRNA and protein are expressed in prostate cancer cell
lines, including LNCaP, PC-3 and DU145, and in human prostate
samples (10). Akashi et
al(11) demonstrated that
patients with a high level of CXCR4 expression in the tumor had
worse cancer-specific survival rates than patients with a low level
of expression. Darash-Yahana et al(12) reported that subcutaneous xenografts
of prostate tumors that overexpressed CXCR4 in nude mice were 2- to
3-fold larger in volume and weight compared to the controls.
Moreover, the blood vessel density, invasiveness of the tumors and
metastasis to the lymph nodes and lungs were significantly
increased in these tumors. In the present study, the potential
therapeutic effects of the CXCR4 antagonist were explored in a
prostate cancer xenograft model.
Materials and methods
Cell culture and reagents
Human prostate cancer PC-3 cells, which are
representative of CRPC cells, were obtained from the Korea Cell
Line Bank (Seoul, Korea). These cells were maintained in RPMI-1640
supplemented with 10% fetal bovine serum (FBS), 1%
penicillin-streptomycin and 1% L-glutamine. All cells were grown in
a humidified incubator at 37°C and 5% CO2. SDF-1
(R&D Systems, Minneapolis, MN, USA) was used as a specific
ligand for CXCR4, and the bicyclam derivative, AMD3100
(Sigma-Aldrich, St. Louis, MO, USA), was used as a CXCR4-specific
antagonist. In certain experiments, the cells were pretreated with
filipin (Sigma-Aldrich) to deplete membrane cholesterol (13).
In vitro proliferation assay
The PC-3 cells were seeded at a density of
2×103 cells/well into 96-well plates in culture medium
containing 10% FBS. After 24 h, the cells were washed and cultured
with serum-free medium alone (control) or with medium containing
SDF-1 or AMD3100 at various concentrations. After 72 h, the number
of viable cells was counted using a CellTiter 96®
AQueous One Solution Cell Proliferation Assay (Promega, Madison,
WI, USA) according to the manufacturer’s instructions. This assay
is based on the ability of viable cells to bioreduce
3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfonyl)-2H-tetrazolium
to formazan in the presence of phenazine methosulfate, an
electron-coupling reagent. Formazan production was quantified by
measuring absorbance at 490 nm, which is directly proportional to
the number of living cells.
Western blot analysis
Prostate cancer cells were cultured to subconfluence
(80–90%), washed and incubated in serum-free media for 12 h. SDF-1
stimulation was performed with 0–200 ng/ml SDF-1 for various
lengths of time. In certain experiments, 1 μg/ml AMD3100 or 1 μg/ml
filipin was pre-incubated with the cells. The cells were lysed with
radio-immunoprecipitation assay (RIPA) lysis buffer consisting of
50 mM HEPES (pH 7.6; USB, Cleveland, OH, USA), 150 mM NaCl
(Sigma-Aldrich), 1% NP-40, 10 ml/ml phenylmethylsulfonyl fluoride
(both Amresco, Solon, OH, USA) and 10 ml/ml aprotinin
(Sigma-Aldrich), and cleared by centrifugation at 12,000 × g. The
protein concentration was determined using a detergent compatible
protein assay kit (Bio-Rad Laboratories, Hercules, CA, USA). Equal
amounts of cell lysates were separated by 10–12% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and transferred to
nitrocellulose membranes. The membranes were blocked for 1 h at
room temperature in 5% skimmed dried milk and incubated for 2 h at
room temperature with anti-protein kinase B (Akt)1 (1:1,000),
anti-p-Akt1/2/3 (1:1,000) or β-actin (1:2,000) antibodies (all from
Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA). The membranes
were incubated with horseradish peroxidase (HRP)-conjugated goat
anti-rabbit immunoglobulin G (IgG) or goat anti-mouse IgG for 1 h
at room temperature. Protein signals were detected by
chemiluminescence with ECL detection reagents (Amersham
Biosciences, Piscataway, NJ, USA).
Xenografts in nude mice
All the protocols for the animal studies were
reviewed and approved by the Institutional Animal Care and Use
Committee at Yonsei University College of Medicine. Six-week-old
male nude mice (BALB/c) were obtained from Japan SLC (Hamamatsu,
Japan). The PC-3 cells (5×106 cells/mouse) were injected
subcutaneously into the right dorsal region. When the tumors
measured 40 mm3, the mice were randomized to receive
AMD3100 (n=6; 3 mg/kg) or vehicle alone (n=6; phosphate-buffered
saline) by intraperitoneal injection for 5 consecutive days/week
for 4 weeks. Tumor diameters were measured at regular intervals
using calipers. The tumor volume (V) was calculated using the
following formula: V = A × B2/2 (A, axial diameter; B,
rotational diameter). Tumors were excised and fixed overnight in
neutral-buffered formalin and processed by routine methods.
Immunohistochemistry
Immunohistochemical staining was performed using the
mouse anti-human Ki-67 monoclonal antibody (1:50), the mouse
anti-human Bcl-2 antibody (prediluted form; both from Dako,
Carpinteria, CA, USA) and the rabbit anti-mouse CD34 monoclonal
antibody (1:100; BD Pharmingen, San Jose, CA, USA) in a xenograft
tissue experiment. In brief, formalin-fixed, paraffin-embedded 4-μm
specimens were deparaffinized and rehydrated. The sections were
treated with 2% hydrogen peroxide to inactivate endogenous
peroxidase, and non-specific binding was blocked by treatment with
the blocking reagent. The primary antibody was applied to each
section for 1 h at 37°C, and the appropriate HRP-conjugated
secondary antibody was applied at a dilution of 1:100 for 1 h at
room temperature. Immunoreactivity was subsequently detected using
a 3,3′-diaminobenzidine system (Vector Laboratories, Burlingame,
CA, USA). Nuclei were counterstained using Meyer’s hematoxylin.
Data analysis
Statistical comparisons were performed among groups
using Student’s t-test or the Mann-Whitney U test. P<0.05 was
considered to indicate a statistically significant difference. The
Statistical Package for Social Sciences (SPSS), version 12.0 for
Windows, was used for the statistical analysis. Data are expressed
as the mean ± standard deviation. All in vitro experiments
were repeated with triplicate or quadruplicate samples and similar
results were obtained across all trials.
Results
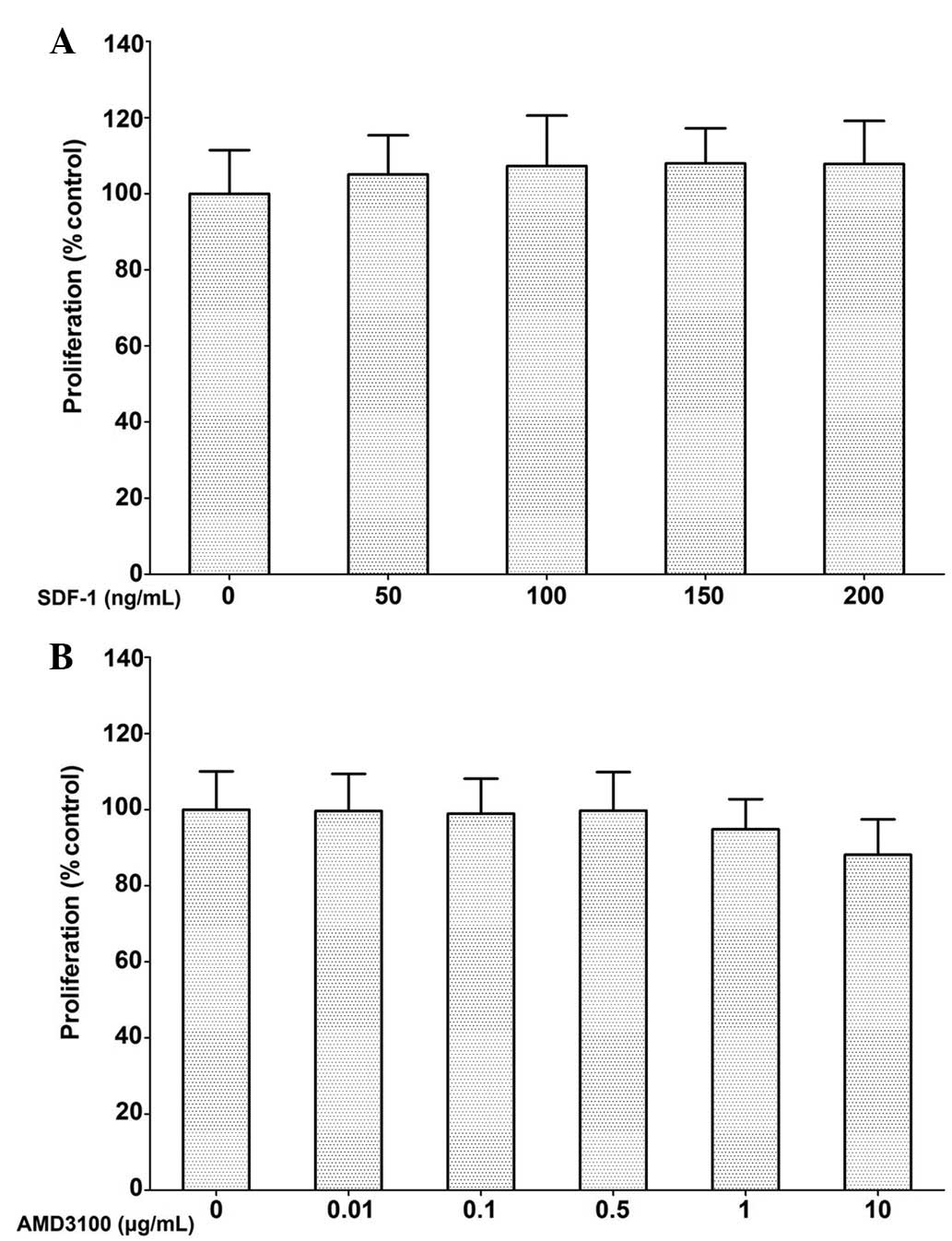
Effect of SDF-1 and a CXCR4 antagonist on
proliferation of PC-3 cells
Subsequent to incubation for 72 h, PC-3 cell
proliferation was not significantly affected by SDF-1 at
concentrations ranging between 50 and 200 ng/ml. AMD3100 also
showed no inhibitory effect on PC-3 cell proliferation at
concentrations ≤0.5 μg/ml. Although cell proliferation was
marginally suppressed at 1 and 10 μg/ml, the effect was not
statistically significant (Fig.
1).
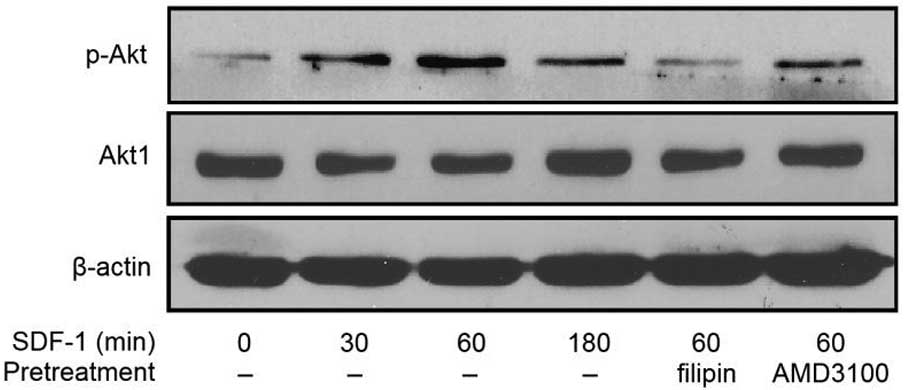
Effect of a CXCR4 antagonist on
SDF-1/CXCR4-mediated Akt signaling in PC-3 cells
Following SDF-1 stimulation, the expression of
phosphorylated Akt was enhanced in the PC-3 cells at 30, 60 and 180
min compared with the control. The highest levels of Akt
phosphorylation following SDF-1 stimulation were observed between
30 and 60 min. The expression of pAkt induced by SDF-1 was
marginally increased in the AMD3100-treated PC-3 cells compared to
the controls, but the levels did not reach those expressed in the
PC-3 cells stimulated by SDF-1 alone. Examination of the total Akt
levels demonstrated that almost all of the conditions resulted in
similar levels of Akt (Fig. 2).
These results indicate that the CXCR4-specific antagonist AMD3100
abrogates SDF-1-induced pAkt activation.
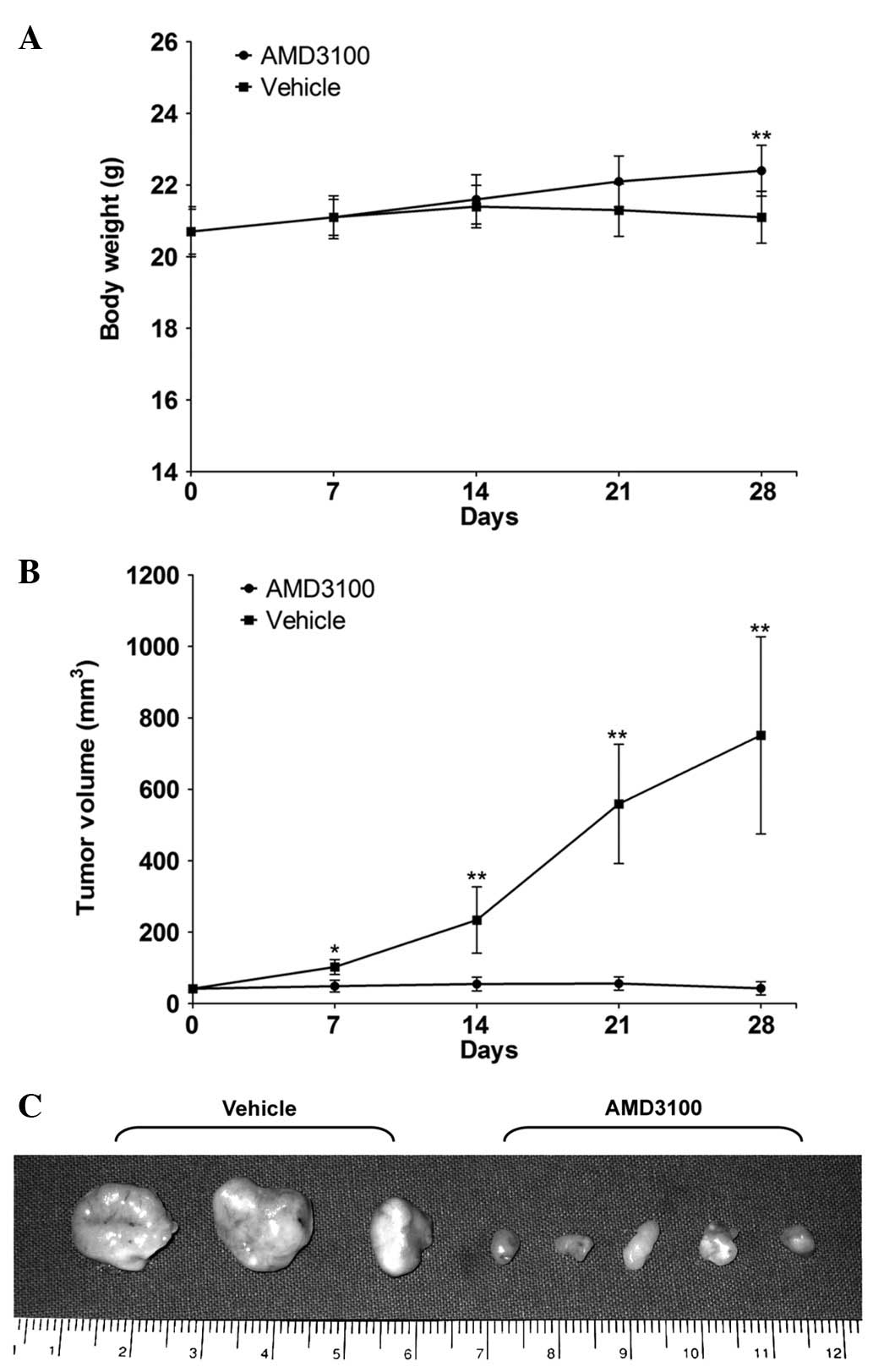
Blocking CXCR4 suppresses the tumor
growth of prostate cancer in a xenograft mouse model
The tumor volumes of the AMD3100-treated and control
groups were 49.0±16.7 and 102.5±20.9 mm3 on day 7
post-treatment, 54.9±19.4 and 234.0±92.9 mm3 on day 14,
56.3±18.5 and 559.9±167.8 mm3 on day 21 and 42.9±18.4
and 751.9±276.4 mm3 on day 28, respectively (Fig. 3). These data indicate that the CXCR4
antagonist used here delayed tumor growth at an early stage of
tumor development. There was no difference in the body weight
between the 2 groups until day 21, however, the body weight on day
28 was 22.4±0.7 g in the AMD3100-treated group and 21.1±0.7 g in
the control group, and this difference was significant (Fig. 3).
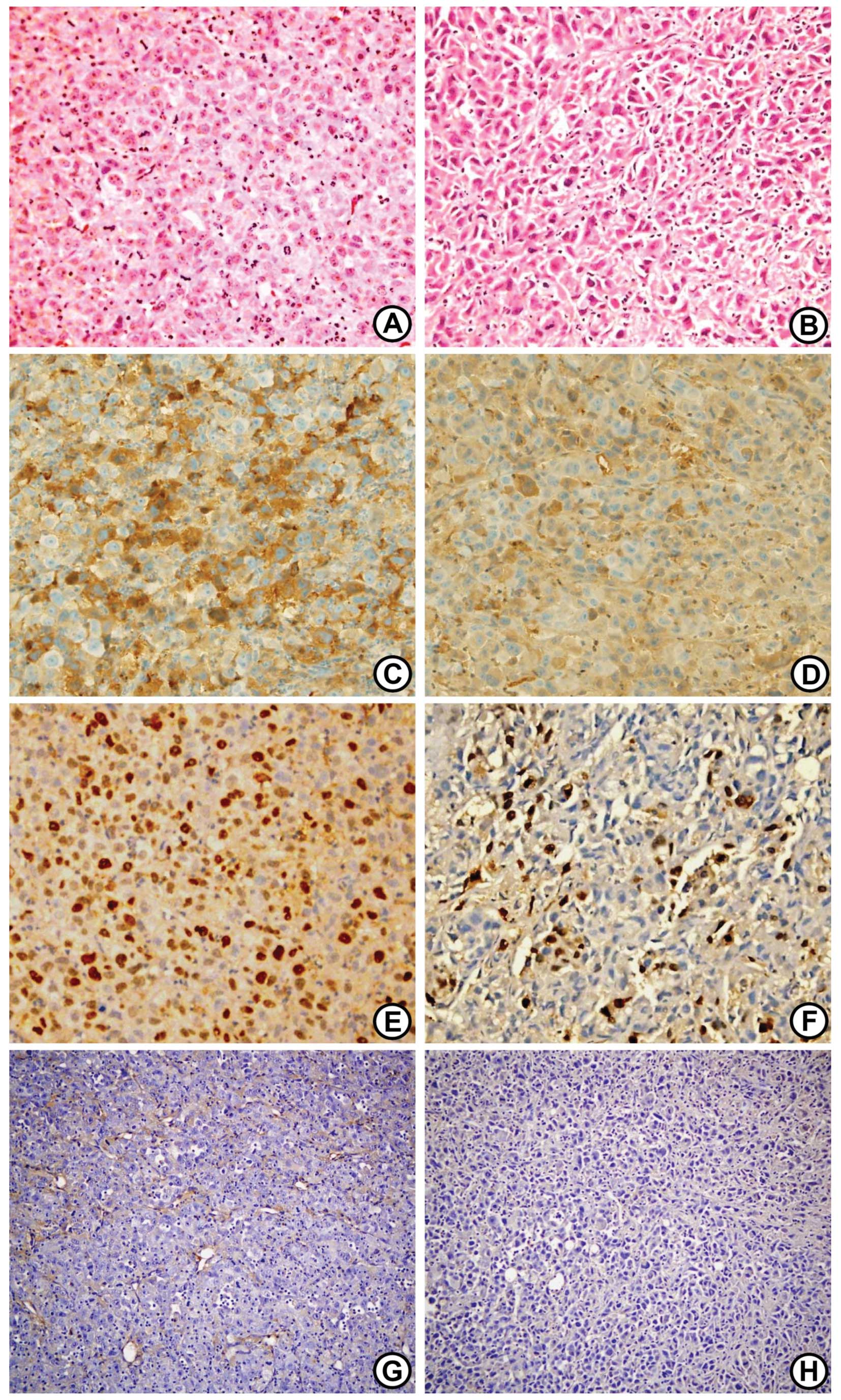
Histopathological examination of PC-3
xenograft tumors following CXCR4 antagonist treatment
Hematoxylin and eosin (HE) staining revealed a
definite histological change in the PC-3 xenograft tumors following
treatment with the CXCR4 antagonist (Fig. 4). CXCR4 antagonist-treated tumors
were characterized by their spindle cell shapes compared with the
control tumors, as well as their enlarged, pleomorphic and
hyperchromatic nuclei. Immunohistochemistry for Bcl-2 expression
showed brownish cytoplasmic staining in the two groups, but Bcl-2
immunostaining was more predominant and more frequently observed in
the control tumors (Fig. 4). The
immunohistochemistry for CD34 was examined on the primary tumor
tissue sections to determine whether the suppression of primary
tumor growth was a result of an antiangiogenic effect (i.e., the
inhibition of microvessel formation) of the CXCR4 antagonist. A
marked reduction in microvessel formation was observed in the
tumors of the CXCR4 antagonist-treated mice (Fig. 4).
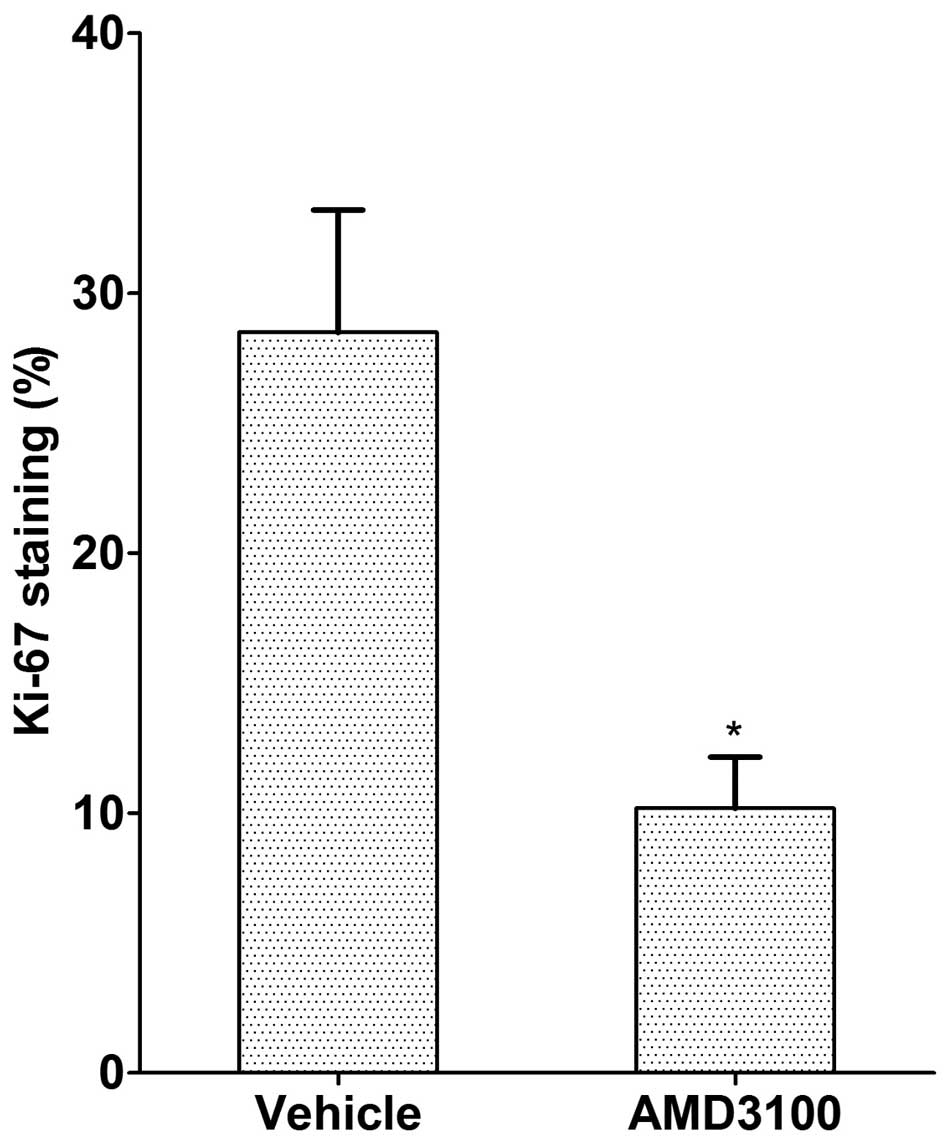
Ki-67 was used as an estimator of tumor
aggressiveness, with dark red-brown nuclear staining regarded as
positivity for Ki-67. The Ki-67 staining index was significantly
higher in the control tumors compared to the CXCR4
antagonist-treated tumors (10.2±1.9 vs. 28.5±4.7%, respectively;
P<0.05; Figs. 4 and 5).
Discussion
The present study observed that a CXCR4-specific
antagonist effectively inhibited CXCR4/Akt signal transduction in
PC-3 cells, as well as tumor growth in nude mice challenged with
PC-3 cells. This indicates that CXCR4 targeting may represent a
novel, effective strategy for the treatment of human prostate
cancer.
It is well known that the binding of chemokines to
their G protein-linked receptors on target cells leads to a series
of signal transduction events involving the generation of inositol
1,4,5-trisphosphate and cyclic adenosine monophosphate-dependent
protein kinase, the activation of phosphatidylinositol 3-kinase
(PI3K), the phosphorylation of protein kinase B (Akt) and
extracellular signal-regulated kinase (ERK), the elevation of
components of focal adhesion complexes and the activation of
protein kinase C (14). SDF-1
binding to CXCR4 generates various signaling mechanisms affecting
the regulation of angiogenesis, the activation of cell invasion,
the promotion of cell growth, the inhibition of apoptosis, and
notably, is important in metastasis (15–20).
In a previous study of prostate cancer, differential activation of
the ERK and PI3K/Akt pathways resulted in differential secretion of
interleukin (IL)-6, IL-8, tissue inhibitors of matrix
metalloproteinase-2 (MMP-2) and vascular endothelial cell growth
factor, which affected the ability of the cancer cells to induce
angiogenesis (15). Exogenous SDF-1
induces Akt phosphorylation in PC-3 cells, which is independent of
PI3K and indispensable for MMP-9 secretion, migration and invasion
(16). SDF-1 induction enhances
various MMPs (MMP-1, -2, -3, -9, -11, -13 and -14) in PC-3 cells
(17). It has also been reported
that the SDF-1-induced expression of CXCR4 in PC-3 cells is
dependent on the mitogen-activated protein kinase (MEK)/ERK
signaling cascade and on nuclear factor-κB (NF-κB) activation,
which enhances endothelial adhesion and transendothelial migration
(18). An immunohistochemical study
of human samples demonstrated that high NF-κB expression was
associated with CXCR4 expression and that they are co-expressed in
approximately one-third of patients with clinically localized
prostate cancer (19). Wang et
al(20) also showed that CXCR4
plays a significant role in prostate cancer metastasis through the
upregulation of vascular endothelial growth factor (VEGF).
The present study showed that SDF-1 has no direct
effects on PC-3 cell proliferation. These findings are in
accordance with results from previous studies of other types of
cells, showing that SDF-1 has no proliferative effect on glioma
(U251n), cholangiocarcinoma (RMCCAI and KKU100), testicular germ
cell tumor (TCAM2), rhabdomyosarcoma and pancreatic cancer cells
(21–25). Notably, Sun et al(10) also showed that recombinant SDF-1
does not alter the growth rate of PC-3 cells under various
conditions. However, an antibody to SDF-1 significantly decreased
the number of PC-3 cells, which indicated that SDF-1 derived from
the PC-3 cells themselves acts in an autocrine fashion to stimulate
growth. Previous studies have demonstrated that SDF-1 stimulates
the proliferation of small cell lung cancer cells (NCI-H69) in the
presence of serum and colorectal cancer (SW480) and epithelial
ovarian cancer (ES-2) cells in the absence of serum (26–28).
In addition, antisense CXCR4 overexpression in glioblastoma cells
has been shown to cause the inhibition of cell proliferation,
indicating that the SDF-1/CXCR4 system is also involved in cell
proliferation in glioblastoma cell lines (29,30).
These differences may be due to different culture systems or target
cells.
The effects of AMD3100 on the viability of tumor
cells are controversial. In the present study, AMD3100 did not
significantly affect the viability of the PC-3 cells. Glioma
(U251n), testicular germ cell tumor (TCAM2), epithelial ovarian
cancer (ES-2) and oral squamous carcinoma (B88-SDF-1) cells have
also been shown to be insensitive to AMD3100 (24,25,28,31).
The enhancing effect of SDF-1 on cell proliferation was markedly
inhibited by AMD3100 treatment in colorectal cancer cells (SW480),
but AMD3100 alone did not significantly affect cell proliferation
when compared with the results observed in the SDF-1 unstimulated
group, which indicated that there was no autocrine growth
stimulatory loop in the cell line (27).
Although AMD3100 did not affect the viability of the
PC-3 cells in the present study, it inhibited SDF-1-induced Akt
activation. The Akt pathway is an important signaling pathway in
prostate cancer (32,33). Akt is a serine-threonine kinase and
its phosphorylation is linked to mitogenic signals. In addition to
its role in survival, Akt participates in a number of intracellular
pathways, including the integration of proliferation and
differentiation signals that mediate migration and angiogenesis
(16,32). To determine whether a CXCR4-specific
antagonist inhibits SDF-1/CXCR4-mediated Akt phosphorylation, in
the present study, the PC-3 cells were pretreated with AMD3100. The
phosphorylation of Akt in the AMD3100 pretreated cells was
significantly lower than in the untreated cells and was similar to
that observed in studies of cholangiocarcinoma cells (21). It has also been reported that in
SDF-1-stimulated activation of ERK1/2 and Akt, rapid responses to
SDF-1 are attenuated by AMD3100 in medulloblastoma and glioblastoma
cells (Daoy and U87) (34).
The present results clearly demonstrate the
inhibitory effects of a CXCR4-specific antagonist on PC-3 tumor
growth in nude mice, although AMD3100 treatment had no direct
effect on the proliferation of PC-3 cells in vitro. Previous
studies have investigated the effects of CXCR4 antagonism on tumor
growth and metastasis in other xenograft models. Such studies have
observed that AMD3100 effectively inhibits anaplastic thyroid
carcinoma tumor growth (35).
Intraperitoneal treatment with AMD3100 has been shown to result in
reduced dissemination in nude mice inoculated with epithelial
ovarian cancer cells (ES-2) (28).
In an oral squamous cell carcinoma xenograft model, AMD3100
significantly inhibited lung metastasis of the SDF-1 transfectant,
ameliorated body weight loss and improved the survival rates of
tumor-bearing nude mice (31).
Another anti-CXCR4 treatment, TN14003, has been shown to suppress
primary tumor growth by inhibiting tumor angiogenesis and
preventing lung metastasis of squamous cell carcinoma of the head
and neck in animal models (36).
The present study also showed that there was a marked reduction in
microvessel formation in CXCR4 antagonist-treated tumors compared
with tumors in the control group.
These marked differences in the biological effects
of CXCR4 inhibition observed in animals and in cell culture may be
explained by the fact that SDF-1 acts at multiple levels in the
tumor microenvironment. Tumor stromal cells, including fibroblasts
and bone marrow-derived cells, express high levels of SDF-1, which
may directly enhance the growth of epithelial tumor cells and
recruit endothelial progenitors, thus favoring angiogenesis
(37). It is believed that chronic
treatment with AMD3100 efficiently blocks SDF-1-mediated
vasculogenesis. Accordingly, the suppression of tumor growth in
treated mice may be explained by the inhibition of
CXCR4+ tumor cell proliferation and the diminishing
recruitment of CXCR4+ angiogenic cells. The present
study observed a strong inhibition of PC-3 tumor growth following
AMD3100 treatment, but AMD3100 did not induce a complete regression
of the tumors. Therefore, we hypothesize that combined treatment
with AMD3100 and antineoplastic agents, such as platinum or
taxanes, is a promising strategy.
No toxic effects of AMD3100 were detected in the
animal model of the present study, but the appropriate therapeutic
approach for antagonizing CXCR4 remains unclear, as long-term
sustained dosing of AMD3100 results in some toxicity (38). A previous study showed that the
sustained dosing of AMD3100 over a 10-day period was associated
with mild toxicities. Reflecting the effects of AMD3100 on bone
marrow function, an elevation in white blood cell count was evident
throughout an 18-day follow-up period following cessation of
AMD3100 treatment (38). For these
reasons, further studies aimed at understanding the effects of the
long-term administration of CXCR4 inhibitors must be pursued.
Despite these considerations, the present data, together with data
from several other reports, markedly indicate that the inhibition
of this pathway should be actively evaluated as a novel anticancer
therapy. AMD3100 or other CXCR4-specific inhibitors should be
developed and tested as therapies for human prostate cancer.
In conclusion, the present study showed that the
CXCR4-specific antagonist, AMD3100, effectively inhibited
SDF-1-induced CXCR4/Akt signal transduction in the PC-3 cells.
Moreover, AMD3100 clearly suppressed tumor growth in the nude mice
inoculated with the PC-3 cells, and AMD3100-treated PC-3 tumors
showed lower levels of microvessel formation and a lower
immunoreactivity for the proliferation marker Ki-67 and the
anti-apoptotic marker Bcl-2 compared with control tumors in
vivo. Thus, the study indicates that CXCR4 targeting may
represent an effective strategy for the treatment of CRPC.
References
|
1
|
No authors listed. Leuprolide versus
diethylstilbestrol for metastatic prostate cancer. The Leuprolide
Study Group. N Engl J Med. 311:1281–1286. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gittes RF: Carcinoma of the prostate. N
Engl J Med. 324:236–245. 1991. View Article : Google Scholar
|
|
3
|
Wirth MP: Hormone-refractory prostate
cancer: what have we learned? BJU Int. 100(Suppl 2): 56–59. 2007.
View Article : Google Scholar
|
|
4
|
Tashiro K, Tada H, Heilker R, Shirozu M,
Nakano T and Honjo T: Signal sequence trap: a cloning strategy for
secreted proteins and type I membrane proteins. Science.
261:600–603. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bleul CC, Fuhlbrigge RC, Casasnovas JM,
Aiuti A and Springer TA: A highly efficacious lymphocyte
chemoattractant, stromal cell-derived factor 1 (SDF-1). J Exp Med.
184:1101–1109. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wells TN, Power CA, Lusti-Narasimhan M, et
al: Selectivity and antagonism of chemokine receptors. J Leukoc
Biol. 59:53–60. 1996.PubMed/NCBI
|
|
7
|
Zlotnik A: Chemokines and cancer. Int J
Cancer. 119:2026–2029. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Balkwill F: The significance of cancer
cell expression of the chemokine receptor CXCR4. Semin Cancer Biol.
14:171–179. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Furusato B, Mohamed A, Uhlen M and Rhim
JS: CXCR4 and cancer. Pathol Int. 60:497–505. 2010. View Article : Google Scholar
|
|
10
|
Sun YX, Wang J, Shelburne CE, et al:
Expression of CXCR4 and CXCL12 (SDF-1) in human prostate cancers
(PCa) in vivo. J Cell Biochem. 89:462–473. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Akashi T, Koizumi K, Tsuneyama K, Saiki I,
Takano Y and Fuse H: Chemokine receptor CXCR4 expression and
prognosis in patients with metastatic prostate cancer. Cancer Sci.
99:539–542. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Darash-Yahana M, Pikarsky E, Abramovitch
R, et al: Role of high expression levels of CXCR4 in tumor growth,
vascularization, and metastasis. FASEB J. 18:1240–1242.
2004.PubMed/NCBI
|
|
13
|
Smart EJ and Anderson RG: Alterations in
membrane cholesterol that affect structure and function of
caveolae. Methods Enzymol. 353:131–139. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kucia M, Reca R, Miekus K, et al:
Trafficking of normal stem cells and metastasis of cancer stem
cells involve similar mechanisms: pivotal role of the SDF-1-CXCR4
axis. Stem Cells. 23:879–894. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang J, Wang J, Sun Y, et al: Diverse
signaling pathways through the SDF-1/CXCR4 chemokine axis in
prostate cancer cell lines leads to altered patterns of cytokine
secretion and angiogenesis. Cell Signal. 17:1578–1592. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chinni SR, Sivalogan S, Dong Z, et al:
CXCL12/CXCR4 signaling activates Akt-1 and MMP-9 expression in
prostate cancer cells: the role of bone microenvironment-associated
CXCL12. Prostate. 66:32–48. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Singh S, Singh UP, Grizzle WE and Lillard
JW Jr: CXCL12-CXCR4 interactions modulate prostate cancer cell
migration, metalloproteinase expression and invasion. Lab Invest.
84:1666–1676. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kukreja P, Abdel-Mageed AB, Mondal D, Liu
K and Agrawal KC: Up-regulation of CXCR4 expression in PC-3 cells
by stromal-derived factor-1alpha (CXCL12) increases endothelial
adhesion and transendothelial migration: role of MEK/ERK signaling
pathway-dependent NF-kappaB activation. Cancer Res. 65:9891–9898.
2005. View Article : Google Scholar
|
|
19
|
Okera M, Bae K, Bernstein E, et al:
Evaluation of nuclear factor kappaB and chemokine receptor CXCR4
co-expression in patients with prostate cancer in the Radiation
Therapy Oncology Group (RTOG) 8610. BJU Int. 108:E51–E58. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang Q, Diao X, Sun J and Chen Z:
Regulation of VEGF, MMP-9 and metastasis by CXCR4 in a prostate
cancer cell line. Cell Biol Int. 35:897–904. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Leelawat K, Leelawat S, Narong S and
Hongeng S: Roles of the MEK1/2 and AKT pathways in CXCL12/CXCR4
induced cholangiocarcinoma cell invasion. World J Gastroenterol.
13:1561–1568. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Libura J, Drukala J, Majka M, et al:
CXCR4-SDF-1 signaling is active in rhabdomyosarcoma cells and
regulates locomotion, chemotaxis, and adhesion. Blood.
100:2597–2606. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mori T, Doi R, Koizumi M, et al: CXCR4
antagonist inhibits stromal cell-derived factor 1-induced migration
and invasion of human pancreatic cancer. Mol Cancer Ther. 3:29–37.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hong X, Jiang F, Kalkanis SN, et al: SDF-1
and CXCR4 are up-regulated by VEGF and contribute to glioma cell
invasion. Cancer Lett. 236:39–45. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gilbert DC, Chandler I, McIntyre A, et al:
Clinical and biological significance of CXCL12 and CXCR4 expression
in adult testes and germ cell tumours of adults and adolescents. J
Pathol. 217:94–102. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kijima T, Maulik G, Ma PC, et al:
Regulation of cellular proliferation, cytoskeletal function, and
signal transduction through CXCR4 and c-Kit in small cell lung
cancer cells. Cancer Res. 62:6304–6311. 2002.PubMed/NCBI
|
|
27
|
Li JK, Yu L, Shen Y, Zhou LS, Wang YC and
Zhang JH: Inhibition of CXCR4 activity with AMD3100 decreases
invasion of human colorectal cancer cells in vitro. World J
Gastroenterol. 14:2308–2313. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kajiyama H, Shibata K, Terauchi M, Ino K,
Nawa A and Kikkawa F: Involvement of SDF-1alpha/CXCR4 axis in the
enhanced peritoneal metastasis of epithelial ovarian carcinoma. Int
J Cancer. 122:91–99. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sehgal A, Keener C, Boynton AL, Warrick J
and Murphy GP: CXCR-4, a chemokine receptor, is overexpressed in
and required for proliferation of glioblastoma tumor cells. J Surg
Oncol. 69:99–104. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sehgal A, Ricks S, Boynton AL, Warrick J
and Murphy GP: Molecular characterization of CXCR-4: a potential
brain tumor-associated gene. J Surg Oncol. 69:239–248. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Uchida D, Onoue T, Tomizuka Y, et al:
Involvement of an autocrine stromal cell derived factor-1/CXCR4
system on the distant metastasis of human oral squamous cell
carcinoma. Mol Cancer Res. 5:685–694. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kreisberg JI, Malik SN, Prihoda TJ, et al:
Phosphorylation of Akt (Ser473) is an excellent predictor of poor
clinical outcome in prostate cancer. Cancer Res. 64:5232–5236.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ayala G, Thompson T, Yang G, et al: High
levels of phosphorylated form of Akt-1 in prostate cancer and
non-neoplastic prostate tissues are strong predictors of
biochemical recurrence. Clin Cancer Res. 10:6572–6578. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Rubin JB, Kung AL, Klein RS, et al: A
small-molecule antagonist of CXCR4 inhibits intracranial growth of
primary brain tumors. Proc Natl Acad Sci USA. 100:13513–13518.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
De Falco V, Guarino V, Avilla E, et al:
Biological role and potential therapeutic targeting of the
chemokine receptor CXCR4 in undifferentiated thyroid cancer. Cancer
Res. 67:11821–11829. 2007.PubMed/NCBI
|
|
36
|
Yoon Y, Liang Z, Zhang X, et al: CXC
chemokine receptor-4 antagonist blocks both growth of primary tumor
and metastasis of head and neck cancer in xenograft mouse models.
Cancer Res. 67:7518–7524. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Orimo A, Gupta PB, Sgroi DC, et al:
Stromal fibroblasts present in invasive human breast carcinomas
promote tumor growth and angiogenesis through elevated SDF-1/CXCL12
secretion. Cell. 121:335–348. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hendrix CW, Collier AC, Lederman MM, et
al: Safety, pharmacokinetics, and antiviral activity of AMD3100, a
selective CXCR4 receptor inhibitor, in HIV-1 infection. J Acquir
Immune Defic Syndr. 37:1253–1262. 2004. View Article : Google Scholar : PubMed/NCBI
|