1. Introduction
Epithelial ovarian cancers are a heterogeneous group
with varying pathologies, and are broadly categorized into serous,
mucinous, endometrioid and clear cell histotypes. These
histological subtypes demonstrate morphological features that
resemble the Müllerian duct-derived epithelial cells, which are
embryologically derived from the fallopian tubes, endocervix,
endometrium and gestational endometrium, respectively. Epithelial
ovarian cancer is generally managed as a single entity, and treated
with a combination of maximal cytoreductive surgery and
taxane/platinum-based chemotherapy (1). Randomized clinical trials have allowed
us to develop a number of successful therapeutic strategies using
paclitaxel and carboplatin for serous cancer, which is the most
prevalent histotype (2). However,
the current standard treatments applied to high-grade serous cancer
are ineffective for clear cell carcinoma (CCC) (1). CCC is more likely to be chemoresistant
than high-grade serous cancer. The differential expression of
chemoresistance-related genes has been observed in various
histological types of ovarian cancer (1).
The intrinsic chemoresistance of CCC appears to be
related to an increase in the detoxification of the drug within the
cell or an increase in cell cycle arrest during the DNA damage
response (3). Previous studies have
demonstrated that the majority of the CCC-specific genes display a
significant change in expression upon exposure to an oxidative
stress microenvironment (3–5). Several studies have summarized the
clinicopathological features, phenotypic and genomic changes in CCC
and provided mechanistic interpretations of the gene expression
changes, including chemoresistance, cell cycle regulation, hormone
independency, detoxification, glycogen synthesis and chromosomal
instability (3,6,7).
Endometriosis-associated neoplasms include CCC,
endometrioid adenocarcinoma, serous borderline tumors, Müllerian
mucinous borderline tumors, adenosarcoma and endometrial stromal
sarcoma (8). Substantial
histopathological data provide evidence that CCC and endometrioid
adenocarcinoma are the most common histologies in ovarian cancer
patients who have associated endometriosis, also known as
endometriosis-associated ovarian cancer. Abundant iron-mediated
oxidative stress occurs due to repeated hemorrhage in
endometriosis, prior to this compound oxidatively modifying host
genomic DNA stability, which is a significant factor that
accelerates carcinogenesis (9).
Thus, the pathogenesis of endometriosis-associated ovarian
carcinogenesis may be closely associated with iron overload. Iron
overload-related diseases, including hemochromatosis, chronic viral
hepatitis, asbestosis and endometriosis, may lead to the
development of hepatocellular carcinoma, mesothelioma or ovarian
cancer (10). Such
microenvironments may play a role in carcinogenesis. Nevertheless,
CCC and endometrioid adenocarcinoma have distinct
clinicopathological characteristics and molecular phenotypes
(11,12). The present study reviews a
considerably expanded functional set of CCC-specific genes and
discusses recent insights into the pathophysiology of CCC.
2. Review of the literature
A comprehensive review of the literature was
conducted in order to investigate the molecular basis of the
CCC-specific genes. A MEDLINE search of the literature was
performed using the key words ‘endometriosis’, ‘ovarian cancer’,
‘clear cell carcinoma’, ‘estrogen receptor’, ‘Arias-Stella
reaction’ and ‘HNF-1β’. English-language publications in PubMed and
references from relevant articles published between 2005 and 2012
were analyzed. References in the studies identified were also
searched.
3. CCC-specific genetic signaling
circuitries
Several genome-wide gene expression analyses have
led to the identification of sets of CCC-specific genes and their
regulated targets, and highlighted a complex circuitry responsible
for CCC development, maintenance and progression (5). Two characteristics define CCC cells,
hormone independency and HNF-1β overexpression. A previous study on
the immunophenotype of CCC cells showed that the tumor cells are
positive for HNF-1β and insulin-like growth factor binding
protein-1 (IGFBP-1), but negative for estrogen receptor (ER),
progesterone receptor (PR), Wilms tumor 1 (WT1) and p53 (13).
The expression of the hormone receptors, ER and PR,
is more elevated in cells of endometrioid-type than in those of
non-endometrioid origin (14). ER
and PR expression may be a significant event in the progression of
endometrioid carcinoma, which develops from endometrial hyperplasia
with the risks of unopposed estrogen signaling in the endometriotic
epithelium. Notably, CCC has shown negative or weak staining for
ER-α and PR (13). The frequent
loss of estrogen function may be a turning point in CCC progression
and aggressiveness. DNA methylation of the promoter region, histone
deacetylation, chromatin remodeling and ubiquitination may
negatively regulate ER expression in CCC (11).
Among epithelial ovarian cancers, the majority of
CCC cases showed the expression of the HNF-1β gene, whereas non-CCC
tumors hardly expressed this gene (15). The CCC-specific genes are in
extremely close linkage with HNF-1β and its target genes. HNF-1β
may be a central player in the differentiation into CCC-specific
lineages from endometriosis (4).
There is a significant overlap between the HNF-1β-related genes and
those associated with chemoresistance, detoxification and glycogen
synthesis, with the conservation of the biological features of CCC
(3,7,11,12).
HNF-1β plays a role in the cellular stress response in CCC through
its regulation of cytoprotective genes (3). Despite recent studies, our
understanding of the HNF-1β-associated molecular mechanisms and
functional players in CCC remains limited.
To further elucidate the biological pathway network,
the present study reviewed extensive functional studies of
CCC-specific genes and proteins at the transcriptome and proteome
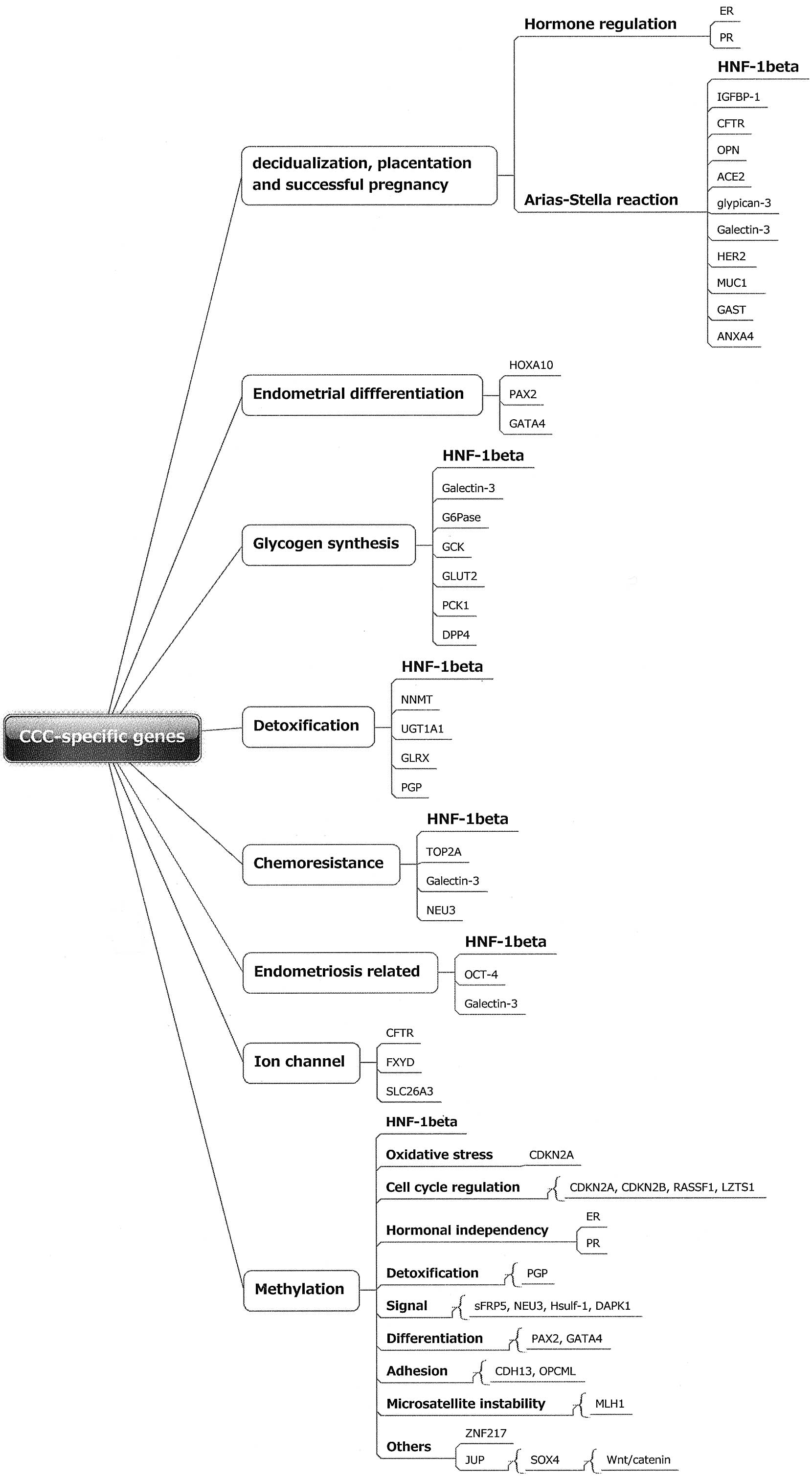
levels. The CCC-specific genes were classified into several groups
based on assorted physiological functions (Fig. 1). The differentially expressed genes
are mainly involved in several functional groups, including those
of decidualization, placentation and successful pregnancy, the
Arial-Stella reaction, differentiation, glycogen synthesis,
detoxification, chemoresistance, endometriosis development and ion
channels. Progesterone is required to develop and maintain the
decidualized phenotype of the endometrium and plays a role in
epithelial-mesenchymal communication (16). The process of the differentiation of
endometrial stromal cells into secretory cells, known as
decidualization, is a prerequisite for the successful implantation,
progression and maintenance of a pregnancy. The placenta is
critical for nourishing the fetus in order to meet the requirements
of the growing fetus throughout pregnancy, and also produces
metabolizing enzymes that detoxify foreign chemicals. Ion channels
play important roles in the feto-placental vasculature and are
essential for electrolyte absorption and replacement of essential
amino acids (17). These data allow
us to speculate that the CCC-specific genes stimulate the secretory
functions of the endometrium that are required for differentiation
and implantation, placentation, and pregnancy recognition
signaling, and are essential for fetal development.
Arias-Stella reaction
Epigenetic, genetic, morphological and functional
changes occurring within the endometrium during the menstrual cycle
are orchestrated under the influence of sequential exposure to the
cyclic expression of the ovarian steroids, estradiol and
progesterone. These changes are required for the development of
uterine receptivity, the disintegration of the decidualized
endometrium and the subsequent repair and fine remodeling for
tissue regeneration, differentiation and placentation. Two
independent ERs, ER-α and ER-β, have distinctive cellular
distribution patterns in the reproductive organs. ER-β
predominantly inhibits the activity of ER-α. The expression of ER-α
has been identified as lower during the secretory phase compared
with the proliferative phase, and significantly lower in the
first-trimester decidua (18). In
contrast to ER-α, ER-β was observed to be expressed in all
compartments of the decidual tissues. The PR gene produces two
isoforms, PRA and PRB, which are mainly localized to the nuclei of
stromal cells. PR was expressed at a lower level in the secretory
endometrium than in the proliferative endometrium (18).
Although ER expression in endometriosis is variable,
several studies have reported higher levels of ER-β and lower
levels of ER-α (19). The
expression of PR, particularly the PR-B isoform, is decreased,
demonstrating that the resistance of endometriotic tissue to
progesterone, so-called progesterone resistance, is commonly
observed in females with endometriosis (19). The endometriotic ER and PR
expression pattern resembles that of the eutopic secretory
endometrium. Furthermore, in a study on endometrioid
adenocarcinoma, ER-α expression increased with malignant
transformation. By contrast, a gradual reduction in ER-α and PR
expression was observed with malignant transformation from
endometriosis to atypical endometriosis to CCC (20). There is a marked tendency for the
disappearance of these receptors in association with the malignant
transformation of endometriosis into CCC.
The Arias-Stella reaction is a hormone-induced
hyperplastic change of the endometrial epithelium that is
associated with cellular atypia, which may be the result of the
regeneration and proliferation activities of the endometrial or
endometriotic glands (21). The
changes in this reaction, a common and universally recognized
feature of the gestational endometrium, have been frequently
observed in abortion curettage specimens (21). The reaction was also observed in the
cytoplasm of stromal cells undergoing decidual change in certain
patients treated with exogenous hormones, such as progestin
(21). Endometriosis infrequently
shows decidual changes and a Arias-Stella reaction, suggesting the
possibility of a metaplastic origin from the secondary Müllerian
system (22,23). Morphological and phenotypic features
of the CCC cells resemble those of cells undergoing the
Arias-Stella reaction (24). The
Arias-Stella reaction and CCC often show prominent hobnail features
and clear cell changes. The etiology for the cytoplasmic clearing
observed in these cells includes the accumulation of intracellular
glycogen. Phenotypical homology may emphasize common ancestry
through the genetic continuity. However, ER and PR expression has
shown a marked difference between endometrial epithelial cells,
corresponding to the Arias-Stella reaction (positive staining), and
CCC (often appears negative), indicating that clear cells in the
two entities are phenotypically similar but hormonally distinct.
CCC cells are able to exploit a progression mechanism via
hormone-independent signaling pathways (11,21).
Decidualization, placentation and a
successful pregnancy
The unique gene families discussed in this section
may be closely related to decidualization, placentation and a
successful pregnancy. These proteins are expressed at a higher
level in the secretory endometrium than in the proliferative
endometrium. The insight provided by this review may illuminate how
the hormone-independent state is established and regulated.
Osteopontin (OPN), also known as
secreted phosphoprotein 1
OPN is a glycoprotein of the extracellular matrix
that also acts as a chemokine. OPN is most likely a direct target
gene of HNF-1β, as it contains functional HNF1 binding sites in its
promoter region (25). OPN is
expressed in the cyclic endometrium, decidual stromal cells and
natural killer cells, and higher expression has been detected in
the later gestational phase compared with the early gestational
phase. OPN expression was shown to be regulated by progesterone
(25). The involvement of OPN has
been proposed for maintaining successful pregnancy. Furthermore,
OPN was shown to be overexpressed in the late secretory endometrium
of females with endometriosis (25). This glycoprotein has been shown to
mediate cellular invasion and to contribute to tumorigenesis in
several types of cancer. OPN may be involved in the pathogenesis of
endometriosis and endometriosis-associated CCC, and its regulation
may have a crucial role in CCC therapy (26).
Angiotensin converting enzyme 2
(ACE2)
The ACE2 gene contains functional HNF1 binding sites
and its overexpression has been observed in CCC. ACE2 metabolizes
Angiotensin II, a vasoconstrictor, to Angiotensin (1–7), a
vasodilator. This enzyme stimulates endothelial cell migration,
capillary formation and angiogenesis, and attenuates angiotensin
II-induced reactive oxygen species (ROS) production (27). ACE2 expression has been shown to be
elevated in the secretory phase of endometrial epithelial cells and
early gestational trophoblasts. One of the first signs of
implantation is an increase in endometrial vascular permeability
and angiogenesis (27). Thus, ACE2
may act as a local autocrine/paracrine regulator of angiogenesis,
vascular permeability and growth during decidualization and
implantation (?).
Galectin-3 and lectin,
galactoside-binding, soluble 3 (LGALS3)
Galectin-3 protein has been immunohistochemically
observed in 59.7% of CCC cases (28). It has been implicated in cell cycle
control, adhesion, migration, invasion, angiogenesis and tumor
development through the activation of the Wnt signaling pathway.
The overexpression of galectin-3 in CCC may contribute to cisplatin
resistance (29). Galectin-3 has
been shown to be expressed in endometrial epithelial cells during
the secretory phase of the menstrual cycle, and to regulate
cell-cell adhesion by interacting with integrin-β3 during embryo
implantation (30). 17β-estradiol,
progesterone and human chorionic gonadotropin induced the
expression of galectin-3 (30).
Galectin-3 also stimulates the expression of phosphorylated
glycogen synthase kinase-3β (GSK3β) and regulates glycogen
synthesis. Furthermore, galectin-3 expression has been observed to
be higher in endometriosis than in the eutopic endometrium,
indicating that galectin-3 has a potential role in the development
of endometriosis and subsequently, in CCC (31).
Glypican-3 (GPC3)
The expression of GPC3, a heparan sulfate
proteoglycan, has been observed in 44% of CCC cases, while rarely
being observed in other histological subtypes (32). GPC3 regulates cellular growth by
interacting with a variety of morphogenic or growth factors, such
as Wnt, fibroblast growth factor (FGF)2 and bone morphogenic
protein (BMP). The expression of GPC3 has been identified in the
endometrial epithelium in the gestational period (32). The differentiated
syncytiotrophoblast cells derived from the placenta expressed GPC3
mRNA and protein.
Mucin 1, cell surface associated
(MUC1)
The cancer-associated aberrantly sialylated MUC1
glycoprotein specifically expressed on CCC plays a role in
tumorigenicity, survival under anoikis conditions and malignant
behavior (33). Progesterone
stimulates the expression of MUC1. MUC1 has been observed to be
expressed in endometriosis (34).
The endometrium of fertile females also expresses MUC1, which
carries selectin ligands recognized by the human blastocyst. MUC1
expression may be required at implantation sites to permit embryo
attachment and implantation (34,35).
Annexin A4 (ANXA4)
ANXA4 expression is elevated in CCC and enhances
cancer cell chemoresistance (36).
This protein belongs to a family of calcium-dependent
phospholipid-binding proteins and is involved in ion channel
regulation, exocytosis and signal transduction. Studies have shown
that ANXA4 mRNA is significantly upregulated during
mid-to-late-secretory phases compared with proliferative phases,
and that progesterone, but not estrogen, increased the expression
of ANXA4 mRNA and protein (37).
The lower expression of ANXA4 during implantation in infertile
patients with endometriosis may be associated with the decrease of
endometrial receptivity (38).
Insulin-like growth factor binding
protein-1 (IGFBP-1)
IGFBP-1 is a specific marker for CCC (39). The expression of IGFBP-1 may be more
specific than that of HNF-1β. The specific expression of IGFBP-1
has also been confirmed in the secretory endometrium, the decidua
of the placenta and the Arias-Stella glands (39). Progesterone induces the expression
of IGFBP-1. IGFBP-1 is downregulated in endometriosis via reduced
levels of forkhead box O1 (FOXO1) and homeobox A10 (HOXA10),
upstream target genes of IGFBP-1.
Human epidermal growth factor receptor
2 (HER2), also known as v-erb-b2 erythroblastic leukemia viral
oncogene homolog 2
HER2/neu-positivity in CCC has been observed to be
significantly higher than in other ovarian cancers (40). This oncogene showed the highest
level of expression during the early secretory phase and in the
placenta. HER2 plays fundamental roles in embryogenesis,
development, proliferation and differentiation.
Octamer-binding protein 4 (OCT-4),
also known as POU class 5 homeobox 1
CCC shows positive staining (28%) for the stem cell
marker OCT-4, which is key in stem cell identity and reprogramming
(41). The transcription factor
OCT-4 is one of the most likely markers for endometrial stem cells.
OCT-4 plays a role in embryonic development, particularly during
early embryogenesis. Endometriotic cells express OCT-4, indicating
that the ectopic endometrium in endometriosis has a stem cell
origin and may explain the possible progression to ovarian cancer
(42).
Endometrial differentiation
The proteins that increase between the early and
mid-luteal phases may play direct roles in embryo-uterine
interactions during the implantation process (43). These implantation-related genes
include HOXA10, MUC1, leukemia inhibitory factor (LIF), glycodelin,
matrix metalloproteinase (MMP), tissue inhibitors of matrix
metalloproteinase (TIMP) and E-cadherin. The majority of these
genes are overexpressed in CCC.
HOXA10
In a study by Li et al(44) HOXA10 was not expressed in the normal
ovarian epithelium, endometriosis and ovarian serous carcinomas
samples, while 68.9% of the CCC samples were positive for the
expression of HOXA10. HOXA10 is a DNA-binding transcription factor
that regulates gene expression, morphogenesis and differentiation.
This gene regulates endometrial receptivity and its expression is
decreased in females with endometriosis. HOXA10 is expressed
cyclically during the menstrual cycle in the endometrium, under the
influence of sex steroids, with the highest expression during the
implantation window (45).
Glycogen synthesis
Glycogen accumulation operates as an energy source,
enabling cell growth even under conditions of oxidative stress.
Genes associated with glycogen synthesis are related to a mechanism
of cisplatin (CDDP)-resistance in CCC (3,46).
Glucose-6-phosphatase (G6Pase)
G6Pase catalyzes the hydrolysis of D-glucose
6-phosphate to D-glucose (47). The
endometrium and placenta produce glucose via G6Pase. The key
metabolic change that occurs during the implantation period is the
rise in endometrial glycogen content. Glycogen synthesis is
regulated by progesterone (47).
Glucokinase (GCK), also known as
hexokinase 4
This protein regulates glycerol uptake and is
associated with glycogen synthesis (http://www.ncbi.nlm.nih.gov/gene/2645).
Glucose transporter type 2
(GLUT2)
GLUT isoforms, of which 13 have been identified to
date, mediate facilitated glucose transport. GLUT2 expression
increases in the endometrium throughout the secretory phase and in
the decidua (48).
Phosphoenolpyruvate carboxykinase
(PCK1)
PCK1 regulates gluconeogenesis. PCK1 forms
phosphoenolpyruvate from oxaloacetate and its expression is
regulated by insulin (http://www.ncbi.nlm.nih.gov/gene/5105).
Dipeptidyl peptidase 4 (DPP4), also
known as CD26
DPP4, also known as CD26, is identical to adenosine
deaminase complexing protein-2 (http://www.ncbi.nlm.nih.gov/gene/1803). DPP4
inactivates insulin-sensing incretin hormones, such as
glucagon-like peptide 1 (GLP-1) and glucose-dependent insulintropic
polypeptide (GIP) (49).
Detoxification
The diagnosis of endometriosis is based on the
direct or indirect evidence of cyclic bleeding in ovarian tumors.
Persistent iron-induced free radicals induced by
endometriosis-dependent hemorrhage may be associated with
carcinogenesis. The HNF-1β-related detoxification enzymes are
crucial for protection against oxidative stress. Genes associated
with detoxification are also related to a mechanism of
CDDP-resistance in CCC (46).
UDP-glycosyltransferase 1 family
polypeptide A1 (UGT1A1)
UGT1A1 is an enzyme of the glucuronidation pathway
that plays a critical role in the detoxification that transforms
small lipophilic molecules, including estrogen, bilirubin, hormones
and drugs, into water-soluble, excretable metabolites. Menstruation
and hemorrhage in endometriosis are biochemically active
environments known to undergo potent oxidizing reactions. Iron and
bilirubin mediate the generation of ROS via the Fenton reaction. If
unconjugated or uncontrolled, they may lead to oxidative DNA and
protein damage. UGT1A1 is also involved in controlling the toxicity
of the chemotherapeutic agent irinotecan. The presence of UGT1A1*28
may result in an increased risk of ovarian cancer (50).
Glutaredoxin (GLRX)
GLRX is important for the antioxidant defense and
for the regulation of the cellular redox state. This protein
protects cells from H2O2-induced apoptosis by
regulating the redox state. GLRX plays a critical role in the
detoxification of iron-induced free radicals in CCC (46).
Nicotinamide N-methyltransferase
(NNMT)
NNMT catalyzes the N-methylation of
nicotinamide, pyridines and other structural analogs by using
S-adenosylmethionine as a methyl donor, playing a pivotal
role in the biotransformation and detoxification of numerous
xenobiotics. This gene contributes to MMP-2 expression, which
induces cell invasiveness via the PI3K-AKT pathway, indicating NNMT
as a novel invasion-related gene (51). This protein is most likely involved
in embryo-implantation mechanisms. NNMT is also one of the genes
upregulated in endometrial stromal cells in response to macrophage
activation in endometriosis.
Ion channels
The absorption of uterine cavity fluid in early
pregnancy results in the closure of the lumen and allows
blastocysts to make contact with the luminal epithelium (52). Fluid absorption peaks at the time of
implantation through progesterone action. The expression patterns
of the CCC-specific gene subsets resemble those of the ion
transporting proteins involved in fluid absorption.
Cystic fibrosis transmembrane
conductance regulator (CFTR), also known as ATP-binding cassette
sub-family C, member 7 (ABCC7)
The CFTR gene encodes a member of the ATP-binding
cassette (ABC) transporter superfamily. CFTR is involved in sodium
and chloride absorption within the uterus. Progesterone stimulates
amiloride-sensitive fluid absorption. Mutations in this gene are
associated with the autosomal recessive disorder cystic fibrosis.
HNF-1 interacts with multiple elements across the CFTR locus
(53). CFTR appears to be
overexpressed in endometriosis and CCC.
FXYD domain-containing ion transport
regulator 2 (FXYD2)
FXYD2 is the sodium and potassium-transporting
ATPase subunit γ and is expressed in the uterine endometrium.
HNF-1β binding sites have been detected in the FXYD2 gene (54). Mutations in this gene have been
associated with renal hypomagnesemia.
Solute carrier family 26, member 3
(SLC26A3)
SLC26A3 functions to transport chloride ions across
the cell membrane in exchange for bicarbonate ions (55). The protein is essential for chloride
absorption and is predominantly localized to the mucosa of the
lower intestinal tract.
Cell cycle regulation
Genes mapping to amplified regions in CCC include
20q13.2 (harboring ZNF217), 17q12-q21.32 (harboring HER2, TOP2A,
GAST, JUP and BRCA1) and 17q23.2 (PPM1D) (40). The resistance of CCC to
platinum-based chemotherapy may be caused by low levels of cell
proliferation (6).
Early mitotic inhibitor-1 (Emi1)
Significant overexpression of Emi1 protein was
present in 82% of CCC (55,56). Emi1 protein is a key cell cycle
regulator that promotes S-phase and mitotic entry by inhibiting the
anaphase promoting complex. The overexpression of Emi1 leads to
tetraploidy and genomic instability.
Zinc finger protein 217 (ZNF217)
ZNF217 amplification is observed in 36% of CCC, but
rarely detected in serous cancer, regardless of grade (57). ZNF217 plays a role in the
chemoresistance and poor prognosis in breast and ovarian cancer,
possibly through the Aurora-A signaling cascade.
Topoisomerase (DNA) II α 170kDa
(TOP2A)
TOP2A is a cell cycle regulating gene that is
upregulated in chemoresistant ovarian cancer, particularly in CCC
(58). TOP2 is involved in
processes, including chromosome condensation, chromatid separation
and the relief of torsional stress that occurs during DNA
transcription and replication.
Together, all these data indicate that the
CCC-specific genes significantly overlap with the genes associated
with the conservation of the biological features of CCC.
4. CCC-specific epigenetic signaling
circuitries
The changes in the DNA hypermethylation pattern is
one of the mechanisms for the earliest molecular changes in
carcinogenesis. In epithelial ovarian cancer, the frequently
methylated genes were reported to be the cyclin-dependent kinase
inhibitor 2B (CDKN2B); estrogen receptor 1 (ESR1); secreted
frizzled-related protein 5 (sFRP5); cadherin 13 (CDH13,
H-cadherin); Ras association (RalGDS/AF-6) domain family member 1
(RASSF1); mutL homolog 1, colon cancer, nonpolyposis type 2 (MLH1);
opioid binding protein/cell adhesion molecule-like (OPCML);
sulfatase 1 (Hsulf-1); GATA binding protein 4 (GATA4) and
death-associated protein kinase 1 (DAPK1) genes (59,60).
CCC samples exhibited a higher frequency of CDKN2B, ESR1 and sFRP5
promoter hypermethylation compared with those of other histological
types (Fig. 1). In addition,
homozygous deletions were detected at the CDKN2A, CDKN2B and LZTS1
loci in CCC (57). Genetic events
in CDKN2A and ESR1 genes are rare in endometriosis.
Cell cycle regulation
CDKN2B, also known as p15INK4B, p15,
inhibits cyclin-dependent kinase 4, and CDKN2A, also known as
P16INK4A
The iron-mediated formation of free radicals (e.g.
ROS) has been shown to be associated with carcinogenesis. Akatsuka
et al identified CDKN2A, a cell cycle control gene, and
CDKN2B genes in a ferric nitrilotriacetate-induced renal
carcinogenesis animal model (61).
The two genes may be vulnerable to ROS in endometriosis. The
allelic loss of the CDKN2A gene occurs specifically at the p16
loci.
RASSF1
RASSF1 is commonly silenced by promoter
hypermethylation in a variety of types of human cancer, including
ovarian cancer (62). This protein
interacts with the DNA repair protein, xeroderma pigmentosum,
complementation group A (XPA) and inhibits the accumulation of
cyclin D1, thus inducing cell cycle arrest. No epigenetic
alterations were identified in RASSF1 in endometriosis samples
(63).
Leucine zipper, putative tumor
suppressor 1 (LZTS1)
Fasciculation and elongation protein-ζ1 (FEZ1)
expression is absent or markedly reduced in 38% of ovarian cancers.
Homozygous deletions are detected at the LZTS1 loci at 8p22 in CCC.
LZTS1 has a role in cell-cycle control by interacting with the
Cdk1-cyclin B1 complex (57).
Hormonal regulation
ESR1
The hormonal receptor profile of CCC and
endometriosis is characterized by the low expression of ER-α and
PR, and by ER-β overexpression (64). Hypomethylation at the ER-β promoter
is responsible for high ER-β levels (19). ER-β suppresses ER-α levels. An
increased ER-β to ER-α ratio is responsible for decreased PR
expression.
Detoxification
P-glycoprotein (PGP)
CCC has a lower expression of multi-drug resistance
PGP than serous cancer in females (65). The proliferative endometrium has
revealed no PGP expression, while the secretory and menstrual
endometrium has been identified with positive staining.
Progesterone increases PGP expression and function. All gestational
endometria have shown positive staining for PGP in the Arias-Stella
reaction and the decidua. PGP protects the fetus from exposure to
xenobiotics during pregnancy (66).
Signaling
sFRP5
The sFRP5 promoter has been shown to be
predominantly methylated in CCC tissues, with 64.6% in CCC compared
with 13.3% in serous cancer, and 0% in endometriosis and the normal
ovarian epithelium (60). sFRP5
modulates Wnt signals, which play a significant role in
reproductive events. sFRP5 may regulate endometrial stromal cell
proliferation, survival and differentiation, which is required to
support the developing embryo.
NEU3 (encodes sialidase 3)
Plasma membrane-associated NEU3 is expressed in the
majority of CCC cases. The overexpression of NEU3 significantly
enhances cell resistance to hypoxia (67).
Hsulf-1
Heparan sulfate 6-O-endosulfatases, such as HSulf-1,
selectively remove 6-O-sulfate from heparan sulfate, upregulate
heparin-binding growth factor signaling and confer resistance to
chemotherapy-induced apoptosis. HSulf-1 inactivation in CCC is
partly mediated by the loss of heterozygosity and epigenetic
silencing (68). HNF-1β negatively
regulates HSulf-1 expression.
DAPK1
DAPK1 is a positive mediator of TNF-α and
γ-interferon-induced apoptosis via the NF-κB signaling pathways.
Collins et al reported low levels of DAPK1 expression in CCC
compared with in normal samples (69).
Differentiation
Paired-box gene 2 (PAX2)
PAX2 is a target of transcriptional suppression by
the tumor suppressor gene, WT1, and is essential in embryonic
development of Müllerian organs. Promoter hypomethylation of the
transcription factor PAX2 has been identified in 75% of CCC cases
(70).
GATA4
The family of zinc finger-containing GATA
transcription factors is frequently lost in ovarian cancer, and
this loss accounts for the dedifferentiation of the cancer cells
(71). GATA4 has also been shown to
be frequently lost in preneoplastic lesions, including
morphologically normal inclusion cysts, epithelial hyperplasia or
atypical endometriosis adjacent to malignant cells. GATA4 plays
critical roles in cell lineage specification during early embryonic
development and organ formation.
Adhesion
CDH13
CDH13, glutathione S-transferase-π1 (GSTP1) and
RASSF1 are frequently methylated in sporadic and BRCA1-associated
ovarian cancers. CDH13 is a cell adherence protein and is often
silenced in cancer cells. Like CCC, epigenetic alterations in CDH13
and CDKN2A have been observed in a silica-induced lung cancer model
(72).
OPCML
OPCML is frequently inactivated by allelic loss and
CpG island promoter methylation in epithelial ovarian cancer
(73). OPCML may have an accessory
role in opioid receptor function and negatively regulate a specific
repertoire of receptor tyrosine kinases, including EPH receptor A2
(EPHA2), fibroblast growth factor receptor 1 (FGFR1), FGFR3, HER2
and HER4.
Microsatellite instability
MLH1
Microsatellite instability is proposed to be limited
to CCC and endometrioid cancer. The epigenetic inactivation of
hMLH1 is also an early event in the malignant transformation of
endometriosis (74). Abnormal
methylation has been detected in ~10% of endometriosis cases.
5. HNF-1β and its related genes
Members of the HNF-1 protein family, HNF-1α and
HNF-1β, are homeobox transcription factors involved in embryonic
development and tissue-specific gene expression in several organs,
including the ovaries and uterus. HNF-1β plays a crucial role in
cell differentiation and tissue morphogenesis. HNF-1β-knockout is
embryonic lethal due to the defective differentiation of the
extra-embryonic visceral endoderm (75). This transcription factor controls
endoderm development. HNF-1β may be activated during the
differentiation of embryonic endodermal stem cells.
In the kidney, HNF-1β is upregulated after acute
kidney injury in proximal tubular cells. HNF-1β mutations have been
associated with a variety of disorders of renal development with
polycystic kidney disease (76).
HNF-1β controls cellular proliferation and tubule formation by
regulating the expression of a number of kidney-specific genes,
including polycystic kidney and hepatic disease 1 (PKHD1),
uromodulin (UMOD) and suppressor of cytokine signaling-3 (SOCS3)
expression and the signal transducer and activator of transcription
3 (STAT3)/mitogen-activated protein kinase 1 (MAPK1, Erk)
activation cascade (77).
The intestinal epithelium is a complex system
characterized by continuous cell renewal, differentiation and
apoptosis. In the gut, the clusters of co-regulated genes
associated with HNF-1β are HNF-1α, HNF-4α, FABP1 and UGT2B7
(78). HNF-1β is also able to
control the expression of a number of intestinal target genes,
including FABP1, LPH, CFTR, G6Pase and DPP4. The concerted action
of HNF-1α and HNF-1β activates the expression of Notch, SLC26A3,
ATOH1 and JAG1, which act on cell fate determination, stem cell
self-renewal, epithelial cell polarity, adhesion, cell division,
differentiation and intestinal water absorption.
In the uterine endometrium, epithelial glandular
nuclei have demonstrated no HNF-1β expression in the proliferative
phase, with significant increases in the secretory and menstrual
phases (46). The pregnant
endometrial glandular cells have also been demonstrated to be
uniformly and continuously positive for HNF-1β. The majority of
HNF-1β-associated gene products co-localize to the cilium, a
crucial organelle that plays a significant role in controlling
proliferation and differentiation (79). The present review identifies direct
and indirect target genes of HNF-1β and shows that this
transcription factor plays a crucial role in defining cell fate and
controlling terminal functions in the endoderm epithelium.
Polycystic kidney and hepatic disease 1
(PKHD1)
This gene modulates calcium-dependent renal
epithelial cell proliferation and differentiation. PKHD1 mutations
cause autosomal recessive polycystic kidney disease (80).
Uromodulin (UMOD)
UMOD in urine provides a defense against urinary
tract infections (81). UMOD also
acts as a constitutive inhibitor of calcium crystallization.
Suppressor of cytokine signaling 3
(SOCS3)
SOCS3 has emerged as a critical attenuator of
cytokine-mediated processes in a negative-feedback mechanism to
hinder cytokine receptor activity, indicating a role in the
suppression of tumorigenesis. The DNA-hypermethylation of SOCS
genes in ovarian cancer has led to speculation that silencing of
the SOCS3 gene may promote the oncogenic transformation of
epithelial tissues (82). In the
human endometrium, stromal cell decidualization has been induced in
response to the expression of SOCS3, which was regulated by
hormonal stimulation (83).
Jagged 1 (JAG1)
JAG1 is the ligand for the receptor Notch1 and is
involved in the biological processes of cell adhesion, motility,
cell cycle regulation, cell communication and angiogenesis. Notch1
is expressed in endometrial epithelial and stromal cells, and
mediates stromal differentiation and decidualization. Notch1 and 2
are believed to be stem cell markers for ovarian cancer (84).
The present review shows that HNF-1β is involved in
the differentiation program of tissue structures and
tissue-specific lineage in several organs, including the ovaries,
endometrium, liver, pancreas, kidneys and intestine.
6. Summary
CCC has several significant characteristics, based
on morphological, behavioral and molecular features, which are
distinct from those of other ovarian cancer histologies (85). CCC is the most common entity in
Japan, accounting for ~70% of all endometriosis-associated ovarian
cancers. The scope of computational technologies, including DNA
microarrays, genome-wide gene expression profiling, microRNAs,
methylation arrays and subsequent methods for the visualization of
these datasets, has been extended to various transcriptomic and
proteomic features that are focused on individual platforms
(86).
The present review systematically identified and
reevaluated the CCC-associated genes (Fig. 1). Firstly, due to advancement in
computational predictions, HNF-1β has been unveiled as a major hub
in the biology of CCC (91). With
promoter hypomethylation, the expression of HNF-1β is significantly
upregulated in endometriosis and endometriosis-associated ovarian
CCC. HNF-1β is believed to be a master regulator of endodermal
organogenesis and has long been discussed with regard to its role
in clear cell carcinogenesis (3–5,12,15,46,56,85–87).
While the overexpression of HNF-1β may lead to cell regeneration,
its potential role in malignant transformation has remained
obscure. The present findings of this review highlight the
importance of the HNF-1β-induced global reproductive gene
expression changes on morphology and ultimately on tissue/organ
function. These changes included the process of the abnormal gain
or loss of several significant genes, including decidualization,
endometrial differentiation and regeneration, hormonal dependency,
glycogen synthesis, detoxification, ion exchange and cell cycle
regulation (3,4,12,46,56,85).
Collectively, the present review uncovers an unanticipated link
between HNF-1β upregulation in CCC and the acquisition of cell
cycle regulation under conditions of oxidative stress and
inflammation. Cell cycle arrest may be a failsafe mechanism against
the oxidative stress-induced DNA damage response (88).
Secondly, the present review demonstrated that
HNF-1β is a strong inducer of endometrial receptivity, a
morphogenetic program that is key to successful pregnancy. The
endometrium undergoes morphological and functional changes during
the menstrual cycle, which are essential for uterine receptivity
(12,56,88).
These changes are driven by estrogen and progesterone and involve
the fine control of numerous different genes, which have been
induced by epigenetically-regulated HNF-1β (87). The specific expression of HNF-1β and
its target genes has been confirmed in not only endometriosis and
CCC, but also the secretory endometrium, the Arias-Stella glands
and the decidua of the placenta (11). The Arias-Stella reaction may be the
result of the regeneration and proliferation activity of the
endometrial glands. A number of the HNF-1β target proteins may be
hormonally-regulated and involved in the uteroplacental transport
of substrates important in the implantation process and in early
embryo-endometrial interactions (46,56).
The ectopic overexpression of HNF-1β in the normal secretory and
gestational endometria may be sufficient to induce the
morphological and molecular changes characteristic of a successful
pregnancy. HNF-1β may be an activator to establish female
reproduction, including secretory phase organogenesis, endometrial
regeneration, differentiation, decidualization, glycogen synthesis,
detoxification, cell cycle regulation, implantation, uterine
receptivity, placentation and a successful pregnancy.
Thirdly, the overexpression of HNF-1β along with a
lack of ER/PR expression may be used as a sensitive model system to
identify the molecular characteristics of CCC. ER-α/ER-β and
PRA/PRB are frequently expressed in ovarian cancer with a certain
variability relating to histological subtype, grade and stage. In
total, 70–100% of serous, mucinous and endometrioid cancers show
positive nuclear staining for ER-α. Conversely, CCC is negative for
ER-α (89). ER-β staining for CCC
is similar to that of non-CCC. PRs have been detected in only 10%
of CCC cases compared with non-CCC (80–90%). DNA methylation and
chromatin remodeling are two epigenetic mechanisms that have been
linked with the lack of ER-α expression (90). These epigenetically modified genes
may be involved in the expression of HNF-1β and hormone
receptors.
Finally, cellular glycogen accumulation due to the
promotion of glycogen synthesis is the most conspicuous feature of
CCC (91). The regulated expression
of the CCC-specific genes offers a mechanism to control the
glycogen accumulation processes. HNF-1β regulates numerous aspects
of glycogen synthesis and its target genes also delineate their
interactions with signaling pathways in dictating glycogen
synthesis. Glycogen is synthesized from glucose presumably to act
as a source of energy for CCC cells and endometrial cells at
implantation, placentation and successful pregnancy.
The present review expanded the repertoire of HNF-1β
target genes, its downstream targets and its associated biological
consequences, thereby shedding light on the complex regulatory
circuitries of CCC. The evolutionarily conserved HNF family of
transcription factors may play fundamental roles in regulating the
hormonal microenvironment, secretory endometrial cell
specification, glycogen synthesis and detoxification during
pregnancy. The characteristics of two different entities, CCC cells
and gestational endometrial cells (e.g. the Arias-Stella reaction),
exhibit certain shared phenotypic and genetic features.
In conclusion, the results of the present review
provide support for a marked degree of genetic overlap between CCC
carcinogenesis and early gestational development.
Acknowledgements
This study was supported by a Grant-in-aid for
Scientific Research from the Ministry of Education, Science and
Culture of Japan granted to the Department of Obstetrics and
Gynecology, Nara Medical University.
References
|
1
|
Pectasides D, Pectasides E, Psyrri A and
Economopoulos T: Treatment issues in clear cell carcinoma of the
ovary: a different entity? Oncologist. 11:1089–1094. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ozols RF, Bundy BN, Greer BE, Fowler JM,
Clarke-Pearson D, Burger RA, Mannel RS, DeGeest K, Hartenbach EM
and Baergen R; Gynecologic Oncology Group. Phase III trial of
carboplatin and paclitaxel compared with cisplatin and paclitaxel
in patients with optimally resected stage III ovarian cancer: a
Gynecologic Oncology Group study. J Clin Oncol. 21:3194–3200. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kajihara H, Yamada Y, Kanayama S, Furukawa
N, Noguchi T, Haruta S, Yoshida S, Sado T, Oi H and Kobayashi H:
Clear cell carcinoma of the ovary: potential pathogenic mechanisms
(Review). Oncol Rep. 23:1193–1203. 2010.PubMed/NCBI
|
|
4
|
Yamaguchi K, Mandai M, Oura T, Matsumura
N, Hamanishi J, Baba T, Matsui S, Murphy SK and Konishi I:
Identification of an ovarian clear cell carcinoma gene signature
that reflects inherent disease biology and the carcinogenic
processes. Oncogene. 29:1741–1752. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mandai M, Matsumura N, Baba T, Yamaguchi
K, Hamanishi J and Konishi I: Ovarian clear cell carcinoma as a
stress-responsive cancer: influence of the microenvironment on the
carcinogenesis and cancer phenotype. Cancer Lett. 310:129–133.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Itamochi H, Kigawa J, Akeshima R, Sato S,
Kamazawa S, Takahashi M, Kanamori Y, Suzuki M, Ohwada M and
Terakawa N: Mechanisms of cisplatin resistance in clear cell
carcinoma of the ovary. Oncology. 62:349–353. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yoshida S, Furukawa N, Haruta S, Tanase Y,
Kanayama S, Noguchi T, Sakata M, Yamada Y, Oi H and Kobayashi H:
Theoretical model of treatment strategies for clear cell carcinoma
of the ovary: focus on perspectives. Cancer Treat Rev. 35:608–615.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Higashiura Y, Kajihara H, Shigetomi H and
Kobayashi H: Identification of multiple pathways involved in the
malignant transformation of endometriosis (Review). Oncol Lett.
4:3–9. 2012.PubMed/NCBI
|
|
9
|
Maeda D and Shih IeM: Pathogenesis and the
role of ARID1A mutation in endometriosis-related ovarian neoplasms.
Adv Anat Pathol. 20:45–52. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Toyokuni S: Mysterious link between iron
overload and CDKN2A/2B. J Clin Biochem Nutr. 48:46–49. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tanase Y, Yamada Y, Shigetomi H, Kajihara
H, Oonogi A, Yoshizawa Y, Furukawa N, Haruta S, Yoshida S, Sado T,
Oi H and Kobayashi H: Modulation of estrogenic action in clear cell
carcinoma of the ovary. Exp Ther Med. 3:18–24. 2012.PubMed/NCBI
|
|
12
|
Kajihara H, Yamada Y, Shigetomi H,
Higashiura Y and Kobayashi H: The dichotomy in the histogenesis of
endometriosis-associated ovarian cancer: clear cell-type versus
endometrioid-type adenocarcinoma. Int J Gynecol Pathol. 31:304–312.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
DeLair D, Oliva E, Köbel M, Macias A,
Gilks CB and Soslow RA: Morphologic spectrum of
immunohistochemically characterized clear cell carcinoma of the
ovary: a study of 155 cases. Am J Surg Pathol. 35:36–44. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hecht JL, Kotsopoulos J, Hankinson SE and
Tworoger SS: Relationship between epidemiologic risk factors and
hormone receptor expression in ovarian cancer: results from the
Nurses’ Health Study. Cancer Epidemiol Biomarkers Prev.
18:1624–3160. 2009.PubMed/NCBI
|
|
15
|
Tsuchiya A, Sakamoto M, Yasuda J, Chuma M,
Ohta T, Ohki M, Yasugi T, Taketani Y and Hirohashi S: Expression
profiling in ovarian clear cell carcinoma: identification of
hepatocyte nuclear factor-1 beta as a molecular marker and a
possible molecular target for therapy of ovarian clear cell
carcinoma. Am J Pathol. 163:2503–2512. 2003. View Article : Google Scholar
|
|
16
|
Uchida H, Maruyama T, Nishikawa-Uchida S,
Oda H, Miyazaki K, Yamasaki A and Yoshimura Y: Studies using an in
vitro model show evidence of involvement of epithelial-mesenchymal
transition of human endometrial epithelial cells in human embryo
implantation. J Biol Chem. 287:4441–4450. 2012. View Article : Google Scholar
|
|
17
|
Wareing M and Greenwood SL: Review:
Potassium channels in the human fetoplacental vasculature.
Placenta. 32:S203–S206. 2011. View Article : Google Scholar
|
|
18
|
Milne SA, Henderson TA, Kelly RW, Saunders
PT, Baird DT and Critchley HO: Leukocyte populations and steroid
receptor expression in human first-trimester decidua; regulation by
antiprogestin and prostaglandin E analog. J Clin Endocrinol Metab.
90:4315–4321. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bulun SE, Cheng YH, Pavone ME, Xue Q,
Attar E, Trukhacheva E, Tokunaga H, Utsunomiya H, Yin P, Luo X, Lin
Z, Imir G, Thung S, Su EJ and Kim JJ: Estrogen receptor-beta,
estrogen receptor-alpha, and progesterone resistance in
endometriosis. Semin Reprod Med. 28:36–43. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Akahane T, Sekizawa A, Okuda T, Kushima M,
Saito H and Okai T: Disappearance of steroid hormone dependency
during malignant transformation of ovarian clear cell cancer. Int J
Gynecol Pathol. 24:369–376. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dhingra N, Punia RS, Radotra A and Mohan
H: Arias-Stella reaction in upper genital tract in pregnant and
non-pregnant women: a study of 120 randomly selected cases. Arch
Gynecol Obstet. 276:47–52. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Trpkov K, Guggisberg K and Yilmaz A:
Arias-Stella reaction as a diagnostic pitfall in a bladder biopsy
with endometriosis: case report and review of the pseudoneoplastic
bladder lesions. Pathol Res Pract. 205:653–656. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sakaki M, Hirokawa M, Sano T, Takahashi H,
Tezuka K, Abe K and Sano M: Ovarian endometriosis showing decidual
change and Arias-Stella reaction with biotin-containing
intranuclear inclusions. Acta Cytol. 47:321–324. 2003.
|
|
24
|
Ohkawa K, Amasaki H, Terashima Y, Aizawa S
and Ishikawa E: Clear cell carcinoma of the ovary: light and
electron microscopic studies. Cancer. 40:3019–3029. 1977.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kato N and Motoyama T: Overexpression of
osteopontin in clear cell carcinoma of the ovary: close association
with HNF-1beta expression. Histopathology. 52:682–688. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Matsuura M, Suzuki T and Saito T:
Osteopontin is a new target molecule for ovarian clear cell
carcinoma therapy. Cancer Sci. 101:1828–1833. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fraga-Silva RA, Costa-Fraga FP, Murça TM,
et al: Angiotensin-converting enzyme 2 activation improves
endothelial function. Hypertension. 61:1233–1238. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Min KW, Park MH, Hong SR, Lee H, Kwon SY,
Hong SH, Joo HJ, Park IA, An HJ, Suh KS, Oh HK, Yoo CW, Kim MJ,
Chang HK, Jun SY, Yoon HK, Chang ED, Kim DW and Kim I; The
Gynecologic Pathology Study Group of the Korean Society of
Pathologists. Clear Cell Carcinomas of the Ovary: A
Multi-Institutional Study of 129 Cases in Korea With Prognostic
Significance of Emi1 and Galectin-3. Int J Gynecol Pathol. 32:3–14.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Oishi T, Itamochi H, Kigawa J, Kanamori Y,
Shimada M, Takahashi M, Shimogai R, Kawaguchi W, Sato S and
Terakawa N: Galectin-3 may contribute to Cisplatin resistance in
clear cell carcinoma of the ovary. Int J Gynecol Cancer.
17:1040–1046. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
von Wolff M, Wang X, Gabius HJ and
Strowitzki T: Galectin fingerprinting in human endometrium and
decidua during the menstrual cycle and in early gestation. Mol Hum
Reprod. 11:189–194. 2005.PubMed/NCBI
|
|
31
|
Noël JC, Chapron C, Borghese B, Fayt I and
Anaf V: Galectin-3 is overexpressed in various forms of
endometriosis. Appl Immunohistochem Mol Morphol. 19:253–257.
2011.PubMed/NCBI
|
|
32
|
Maeda D, Ota S, Takazawa Y, Aburatani H,
Nakagawa S, Yano T, Taketani Y, Kodama T and Fukayama M: Glypican-3
expression in clear cell adenocarcinoma of the ovary. Mod Pathol.
22:824–832. 2009.PubMed/NCBI
|
|
33
|
Tamada Y, Takeuchi H, Suzuki N, Susumu N,
Aoki D and Irimura T: Biological and therapeutic significance of
MUC1 with sialoglycans in clear cell adenocarcinoma of the ovary.
Cancer Sci. 98:1586–1591. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Vlad AM, Diaconu I and Gantt KR: MUC1 in
endometriosis and ovarian cancer. Immunol Res. 36:229–236. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Carson DD, Julian J, Lessey BA, Prakobphol
A and Fisher SJ: MUC1 is a scaffold for selectin ligands in the
human uterus. Front Biosci. 11:2903–2908. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kim A, Enomoto T, Serada S, Ueda Y,
Takahashi T, Ripley B, Miyatake T, Fujita M, Lee CM, Morimoto K,
Fujimoto M, Kimura T and Naka T: Enhanced expression of Annexin A4
in clear cell carcinoma of the ovary and its association with
chemoresistance to carboplatin. Int J Cancer. 125:2316–2322. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ponnampalam AP and Rogers PA: Cyclic
changes and hormonal regulation of annexin IV mRNA and protein in
human endometrium. Mol Hum Reprod. 12:661–669. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jiang YL, Li B, Xing FQ, Wang F and Feng
JH: Study on the relationship between altered expression of annexin
A4 and endometrial receptivity during the implantation window in
infertile patients with endometriosis. Zhonghua Fu Chan Ke Za Zhi.
47:324–327. 2012.(In Chinese).
|
|
39
|
Sugita S, Morishita Y, Kano J, Furuya S,
Shiba-Ishii A and Noguchi M: IGFBP-1 is expressed specifically in
ovarian clear cell adenocarcinoma. Histopathology. 58:729–738.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tan DS, Iravani M, McCluggage WG, Lambros
MB, Milanezi F, Mackay A, Gourley C, Geyer FC, Vatcheva R, Millar
J, Thomas K, Natrajan R, Savage K, Fenwick K, Williams A, Jameson
C, El-Bahrawy M, Gore ME, Gabra H, Kaye SB, Ashworth A and
Reis-Filho JS: Genomic analysis reveals the molecular heterogeneity
of ovarian clear cell carcinomas. Clin Cancer Res. 17:1521–1534.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Howell NR, Zheng W, Cheng L, Tornos C,
Kane P, Pearl M, Chalas E and Liang SX: Carcinomas of ovary and
lung with clear cell features: can immunohistochemistry help in
differential diagnosis? Int J Gynecol Pathol. 26:134–140. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Pacchiarotti A, Caserta D, Sbracia M and
Moscarini M: Expression of oct-4 and c-kit antigens in
endometriosis. Fertil Steril. 95:1171–1173. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ghosh D and Sengupta J: Endocrine and
paracrine correlates of endometrial receptivity to blastocyst
implantation in the human. Indian J Physiol Pharmacol. 48:6–30.
2004.PubMed/NCBI
|
|
44
|
Li B, Jin H, Yu Y, Gu C, Zhou X, Zhao N
and Feng Y: HOXA10 is overexpressed in human ovarian clear cell
adenocarcinoma and correlates with poor survival. Int J Gynecol
Cancer. 19:1347–1352. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zanatta A, Rocha AM, Carvalho FM, Pereira
RM, Taylor HS, Motta EL, Baracat EC and Serafini PC: The role of
the Hoxa10/HOXA10 gene in the etiology of endometriosis and its
related infertility: a review. J Assist Reprod Genet. 27:701–710.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kobayashi H, Yamada Y, Kanayama S,
Furukawa N, Noguchi T, Haruta S, Yoshida S, Sakata M, Sado T and Oi
H: The role of hepatocyte nuclear factor-1beta in the pathogenesis
of clear cell carcinoma of the ovary. Int J Gynecol Cancer.
19:471–479. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Salameh W, Helliwell JP, Han G, McPhaul L
and Khorram O: Expression of endometrial glycogen synthase
kinase-3beta protein throughout the menstrual cycle and its
regulation by progesterone. Mol Hum Reprod. 12:543–549. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
von Wolff M, Ursel S, Hahn U, Steldinger R
and Strowitzki T: Glucose transporter proteins (GLUT) in human
endometrium: expression, regulation, and function throughout the
menstrual cycle and in early pregnancy. J Clin Endocrinol Metab.
88:3885–3892. 2003.PubMed/NCBI
|
|
49
|
Senkel S, Lucas B, Klein-Hitpass L and
Ryffel GU: Identification of target genes of the transcription
factor HNF1beta and HNF1alpha in a human embryonic kidney cell
line. Biochim Biophys Acta. 1731:179–190. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Xu JM, Wang Y, Ge FJ, Lin L, Liu ZY and
Sharma MR: Severe irinotecan-induced toxicity in a patient with
UGT1A1*28 and UGT1A1*6 polymorphisms. World J Gastroenterol.
19:3899–3903. 2013.PubMed/NCBI
|
|
51
|
Tang SW, Yang TC, Lin WC, Chang WH, Wang
CC, Lai MK and Lin JY: Nicotinamide N-methyltransferase induces
cellular invasion through activating matrix metalloproteinase-2
expression in clear cell renal cell carcinoma cells.
Carcinogenesis. 32:138–145. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Salleh N, Baines DL, Naftalin RJ and
Milligan SR: The hormonal control of uterine luminal fluid
secretion and absorption. J Membr Biol. 206:17–28. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kerschner JL and Harris A: Transcriptional
networks driving enhancer function in the CFTR gene. Biochem J.
446:203–212. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ferrè S, Veenstra GJ, Bouwmeester R,
Hoenderop JG and Bindels RJ: HNF-1B specifically regulates the
transcription of the γa-subunit of the Na+/K+-ATPase. Biochem
Biophys Res Commun. 404:284–290. 2011.
|
|
55
|
Kobayashi H, Kajiwara H, Kanayama S,
Yamada Y, Furukawa N, Noguchi T, Haruta S, Yoshida S, Sakata M,
Sado T and Oi H: Molecular pathogenesis of endometriosis-associated
clear cell carcinoma of the ovary (Review). Oncol Rep. 22:233–240.
2009.PubMed/NCBI
|
|
56
|
Gütgemann I, Lehman NL, Jackson PK and
Longacre TA: Emi1 protein accumulation implicates misregulation of
the anaphase promoting complex/cyclosome pathway in ovarian clear
cell carcinoma. Mod Pathol. 21:445–454. 2008.PubMed/NCBI
|
|
57
|
Kuo KT, Mao TL, Chen X, Feng Y, Nakayama
K, Wang Y, Glas R, Ma MJ, Kurman RJ, Shih IeM and Wang TL: DNA copy
numbers profiles in affinity-purified ovarian clear cell carcinoma.
Clin Cancer Res. 16:1997–2008. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Ju W, Yoo BC, Kim IJ, Kim JW, Kim SC and
Lee HP: Identification of genes with differential expression in
chemoresistant epithelial ovarian cancer using high-density
oligonucleotide microarrays. Oncol Res. 18:47–56. 2009. View Article : Google Scholar
|
|
59
|
Ozdemir F, Altinisik J, Karateke A,
Coksuer H and Buyru N: Methylation of tumor suppressor genes in
ovarian cancer. Exp Ther Med. 4:1092–1096. 2012.PubMed/NCBI
|
|
60
|
Ho CM, Lai HC, Huang SH, Chien TY, Lin MC
and Chang SF: Promoter methylation of sFRP5 in patients with
ovarian clear cell adenocarcinoma. Eur J Clin Invest. 40:310–318.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Akatsuka S, Yamashita Y, Ohara H, Liu YT,
Izumiya M, Abe K, Ochiai M, Jiang L, Nagai H, Okazaki Y, Murakami
H, Sekido Y, Arai E, Kanai Y, Hino O, Takahashi T, Nakagama H and
Toyokuni S: Fenton reaction induced cancer in wild type rats
recapitulates genomic alterations observed in human cancer. PLoS
One. 7:e434032012. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Choi YL, Kang SY, Shin YK, Choi JS, Kim
SH, Lee SJ, Bae DS and Ahn G: Aberrant hypermethylation of RASSF1A
promoter in ovarian borderline tumors and carcinomas. Virchows
Arch. 448:331–336. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Vestergaard AL, Thorup K, Knudsen UB, Munk
T, Rosbach H, Poulsen JB, Guldberg P and Martensen PM: Oncogenic
events associated with endometrial and ovarian cancers are rare in
endometriosis. Mol Hum Reprod. 17:758–761. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Zannoni GF, Morassi F, Prisco MG, De
Stefano I, Vellone VG, Arena V, Scambia G and Gallo D:
Clinicopathologic and immunohistochemical features of ovarian clear
cell carcinomas in comparison with type I and type II tumors. Int J
Gynecol Pathol. 31:507–516. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Eltabbakh GH, Mount SL, Beatty B,
Simmons-Arnold L and Cooper K: Clinical and molecular differences
between clear cell and papillary serous ovarian carcinoma. J Surg
Oncol. 93:379–386. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Axiotis CA, Guarch R, Merino MJ, Laporte N
and Neumann RD: P-glycoprotein expression is increased in human
secretory and gestational endometrium. Lab Invest. 65:577–581.
1991.PubMed/NCBI
|
|
67
|
Nomura H, Tamada Y, Miyagi T, Suzuki A,
Taira M, Suzuki N, Susumu N, Irimura T and Aoki D: Expression of
NEU3 (plasma membrane-associated sialidase) in clear cell
adenocarcinoma of the ovary: its relationship with T factor of pTNM
classification. Oncol Res. 16:289–297. 2006.PubMed/NCBI
|
|
68
|
Liu P, Khurana A, Rattan R, He X, Kalloger
S, Dowdy S, Gilks B and Shridhar V: Regulation of HSulf-1
expression by variant hepatic nuclear factor 1 in ovarian cancer.
Cancer Res. 69:4843–4850. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Collins Y, Dicioccio R, Keitz B, Lele S
and Odunsi K: Methylation of death-associated protein kinase in
ovarian carcinomas. Int J Gynecol Cancer. 16:195–199. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Song H, Kwan SY, Izaguirre DI, Zu Z, Tsang
YT, Tung CS, King ER, Mok SC, Gershenson DM and Wong KK: PAX2
Expression in Ovarian Cancer. Int J Mol Sci. 14:6090–6105. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Cai KQ, Caslini C, Capo-chichi CD, Slater
C, Smith ER, Wu H, Klein-Szanto AJ, Godwin AK and Xu XX: Loss of
GATA4 and GATA6 expression specifies ovarian cancer histological
subtypes and precedes neoplastic transformation of ovarian surface
epithelia. PLoS One. 4:e64542009. View Article : Google Scholar
|
|
72
|
Blanco D, Vicent S, Fraga MF,
Fernandez-Garcia I, Freire J, Lujambio A, Esteller M,
Ortiz-de-Solorzano C, Pio R, Lecanda F and Montuenga LM: Molecular
analysis of a multistep lung cancer model induced by chronic
inflammation reveals epigenetic regulation of p16 and activation of
the DNA damage response pathway. Neoplasia. 9:840–852. 2007.
View Article : Google Scholar
|
|
73
|
McKie AB, Vaughan S, Zanini E, Okon IS,
Louis L, de Sousa C, Greene MI, Wang Q, Agarwal R, Shaposhnikov D,
Wong JL, Gungor H, Janczar S, El-Bahrawy M, Lam EW, Chayen NE and
Gabra H: The OPCML tumor suppressor functions as a cell surface
repressor-adaptor, negatively regulating receptor tyrosine kinases
in epithelial ovarian cancer. Cancer Discov. 2:156–171. 2012.
View Article : Google Scholar
|
|
74
|
Ren F, Wang D, Jiang Y and Ren F:
Epigenetic inactivation of hMLH1 in the malignant transformation of
ovarian endometriosis. Arch Gynecol Obstet. 285:215–221. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Coffinier C, Thepot D, Babinet C, Yaniv M
and Barra J: Essential role for the homeoprotein vHNF1/HNF1beta in
visceral endoderm differentiation. Development. 126:4785–4794.
1999.PubMed/NCBI
|
|
76
|
Horikawa Y, Iwasaki N, Hara M, Furuta H,
Hinokio Y, Cockburn BN, Lindner T, Yamagata K, Ogata M, Tomonaga O,
Kuroki H, Kasahara T, Iwamoto Y and Bell GI: Mutation in hepatocyte
nuclear factor-1β gene (TCF2) associated with MODY. Nat Genet.
17:384–385. 1997.
|
|
77
|
Ogata K, Shimamura Y, Hamada K, Hisa M,
Bun M, Okada N, Inoue K, Taniguchi Y, Ishihara M, Kagawa T, Horino
T, Fujimoto S and Terada Y: Upregulation of HNF-1β during
experimental acute kidney injury plays a crucial role in renal
tubule regeneration. Am J Physiol Renal Physiol. 303:F689–F699.
2012.
|
|
78
|
D’Angelo A, Bluteau O, Garcia-Gonzalez MA,
Gresh L, Doyen A, Garbay S, Robine S and Pontoglio M: Hepatocyte
nuclear factor 1alpha and beta control terminal differentiation and
cell fate commitment in the gut epithelium. Development.
137:1573–1582. 2010.PubMed/NCBI
|
|
79
|
Avni FE and Hall M: Renal cystic diseases
in children: new concepts. Pediatr Radiol. 40:939–946. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Zhou XH, Hui ZY and Li Y: Clinical and
pathological features of a neonate with autosomal recessive
polycystic kidney disease caused by a nonsense PKHD1 mutation.
World J Pediatr. 9:76–79. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Serafini-Cessi F, Monti A and Cavallone D:
N-Glycans carried by Tamm-Horsfall glycoprotein have a crucial role
in the defense against urinary tract diseases. Glycoconj J.
22:383–394. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Sutherland KD, Lindeman GJ, Choong DY,
Wittlin S, Brentzell L, Phillips W, Campbell IG and Visvader JE:
Differential hypermethylation of SOCS genes in ovarian and breast
carcinomas. Oncogene. 23:7726–7733. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Dimitriadis E, Stoikos C, Tan YL and
Salamonsen LA: Interleukin 11 signaling components signal
transducer and activator of transcription 3 (STAT3) and suppressor
of cytokine signaling 3 (SOCS3) regulate human endometrial stromal
cell differentiation. Endocrinol. 147:3809–3817. 2006. View Article : Google Scholar
|
|
84
|
Oktem G, Sanci M, Bilir A, Yildirim Y,
Kececi SD, Ayla S and Inan S: Cancer stem cell and embryonic
development-associated molecules contribute to prognostic
significance in ovarian cancer. Int J Gynecol Cancer. 22:23–29.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Yamada Y, Shigetomi H, Onogi A, Haruta S,
Kawaguchi R, Yoshida S, Furukawa N, Nagai A, Tanase Y, Tsunemi T,
Oi H and Kobayashi H: Redox-active iron-induced oxidative stress in
the pathogenesis of clear cell carcinoma of the ovary. Int J
Gynecol Cancer. 21:1200–1207. 2011.PubMed/NCBI
|
|
86
|
Haruta S, Furukawa N, Yoshizawa Y, Tsunemi
T, Nagai A, Kawaguchi R, Tanase Y, Yoshida S and Kobayashi H:
Molecular genetics and epidemiology of epithelial ovarian cancer.
Oncol Rep. 26:1347–1356. 2011.PubMed/NCBI
|
|
87
|
Kato N, Tamura G and Motoyama T:
Hypomethylation of hepatocyte nuclear factor-1beta (HNF-1beta) CpG
island in clear cell carcinoma of the ovary. Virchows Arch.
452:175–180. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Shigetomi H, Higashiura Y, Kajihara H and
Kobayashi H: A potential link of oxidative stress and cell cycle
regulation for development of endometriosis. Gynecol Endocrinol.
28:897–902. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Cameron RI, Ashe P, O’Rourke DM, Foster H
and McCluggage WG: A panel of immunohistochemical stains assists in
the distinction between ovarian and renal clear cell carcinoma. Int
J Gynecol Pathol. 22:272–276. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Brinkman JA and El-Ashry D: ER
re-expression and re-sensitization to endocrine therapies in
ER-negative breast cancers. J Mammary Gland Biol Neoplasia.
14:67–78. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Iida Y, Aoki K, Asakura T, Ueda K,
Yanaihara N, Takakura S, Yamada K, Okamoto A, Tanaka T and Ohkawa
K: Hypoxia promotes glycogen synthesis and accumulation in human
ovarian clear cell carcinoma. Int J Oncol. 40:2122–2130.
2012.PubMed/NCBI
|