Introduction
Pancreatic cancer is a highly aggressive malignancy
with a poor survival rate and invasion and metastasis are the most
common cause of mortality in patients (1). Studies have demonstrated that tumor
invasion and metastasis are complex processes that are regulated by
various molecules. The majority of these molecules are ligands and
receptors, which are involved in mediating the cell-to-cell or
cell-to-extracellular matrix (ECM) interactions (2,3).
Ephrin (EPH) receptors belong to the family of
receptor tyrosine kinases and have been demonstrated to be elevated
in the majority of human cancers (4,5). EPH
receptors can be classified into EPHA and EPHB receptors to which
the EPHs are the ligands; divided into type A and B EPH ligands
(6). Generally, type A EPH ligands
are glycosylphosphatidylinisotol-anchored peripheral membrane
molecules that bind EPHA receptors, while type B EPH ligands are
transmembrane molecules that bind EPHB receptors. However, EPHA4
has the ability to bind to type A or B EPH ligands (7). Studies have shown that the expression
level of EPHA4 is upregulated in gastric cancer (8). Furthermore, the overexpression of
EPHA4 has been proven to correlate with liver metastasis in
colorectal cancer (9). However, the
role of EPHA4 in pancreatic cancer remains unclear. Therefore, the
aim of the present study was to determine the effect of EPHA4 on
the motility and invasion of pancreatic cancer cells.
Materials and methods
Antibodies
The antibodies against EPHA4 (mouse monoclonal
antibody), epithelial (E)-cadherin (rabbit monoclonal antibody) and
β-actin (mouse monoclonal antibody) were purchased from Santa Cruz
Biotechnology, Inc. (Santa Cruz, CA, USA) and the antibody against
Snail (rabbit monoclonal antibody) was obtained from Cell Signaling
Technology, Inc. (Beverly, MA, USA).
Cell lines and culture conditions
The pancreatic cancer cell lines, MIA PaCa-2, HAPC,
SW1990, BxPC-3 and Panc-1, were purchased from the American Type
Culture Collection (Manassas, VA, USA). All cells were incubated in
Dulbecco’s modified Eagle’s medium (DMEM; Gibco-BRL, Gaithersburg,
MD, USA) containing 10% fetal bovine serum (FBS; Hyclone, Logan,
UT, USA) and cultured at 37°C in a humidified atmosphere of 5%
CO2.
Small interfering RNA (siRNA) and
transfection
The following EPHA4 siRNA sequence was obtained from
GenePharma (Shanghai, China): 5′-UCAUGAAGCUGAACACCGA-3′. As a
control, the following scramble siRNA sequence was also used:
5′-UUCUCCGAACGUGUCACGU-3′. The cells were transfected with siRNAs
using Lipofectamine reagent (Invitrogen Life Technologies,
Carlsbad, CA, USA) according to the manufacturer’s instructions and
further incubated for 48 h prior to being used in the subsequent
experiments.
Western blotting
The total protein of Panc-1 and BxPC-3 cells was
extracted by radioimmunoprecipitation assay buffer with protease
inhibitor and the concentration of the protein was examined using
the Bicinchoninic acid (BCA) Assay kit (Applygen Technologies Inc.,
Beijing, China). The protein of each sample was then separated by
SDS-PAGE (Invitrogen Life Technologies, Carlsbad, CA, USA) and
transferred onto the polyvinylidene difluoride membranes
(Invitrogen Life Technologies). The membranes were blocked with 5%
non-fat milk for 1 h and incubated with the primary antibodies at
4°C overnight. Next, the membranes were probed with the secondary
antibodies for 1 h at room temperature. Immunopositive bands were
then detected and exposed to film following incubation with an
enhanced chemiluminescence system (Applygen Technologies, Inc.,
Beijing, China). The expression of protein was normalized to the
levels of β-actin.
Wound healing assay
The cells were cultured in six-well plates at a
density of 2×105 cells/well and when the cells achieved
80% confluence, the cell monolayers were scratched with a sterile
plastic pipette tip. The cells were then washed twice with
phosphate-buffered saline and incubated with DMEM for 20 h. Images
were captured at 0 and 20 h under an Olympus X71 inverted
microscope (Olympus Corporation, Tokyo, Japan).
Invasion assay
The invasion assay was performed using the 24-well
Transwell inserts (Costar, Cambridge, MA, USA) and each filter of
the Transwell was coated with Matrigel (BD Biosciences, Franklin
Lakes, NJ, USA). The cells (1×105 cells/well) were
seeded onto the top chamber and 600 μl DMEM with 30% FBS was placed
in the lower chamber. Following incubation for 20 h in a
CO2 incubator, the invaded cells were fixed and stained
with crystal violet (Amresco, Solon, OH, USA). Next, the invaded
cells were observed under a microscope (YS100; Nikon, Tokyo, Japan)
at a magnification of ×200. The mean number of cells in five random
fields was calculated and the data are presented as a percentage of
the invaded cells compared with the control.
Gelatin zymography
The cells were transfected with siRNAs and incubated
in a CO2 incubator for 48 h. The cell supernatant was
then collected and concentrated at 8,000 × g for 30 min in a
concentrator (Amicon Ultra concentrator, 30,000 Da MWCO; Millipore,
Billerica, MA, USA). The concentration of the protein was
determined by BCA assay and equal amounts of protein were separated
on SDS-PAGE containing 1 mg/ml gelatin. The gel was washed with
renaturing buffer (2.5% Triton X-100; Amresco) for 30 min at room
temperature and then incubated with developing buffer [50 mM
Tris-HCl buffer (pH 7.6), 5 mM CaCl2, 200 mM NaCl and
0.02% Brij-35; Invitrogen Life Technologies] overnight at 37°C. The
gel was further stained with 0.5% Coomassie brilliant blue R-250
solution (Amresco) for 30 min, followed by destaining with 7.5%
acetic acid solution containing 10% methanol. The areas of protease
activity appeared as clear bands against the dark blue
background.
Statistical analysis
All experiments were repeated at least three times
and the data are presented as the mean ± standard deviation. The
results were analyzed using the SPSS 17.0 software (SPSS, Inc.,
Chicago, IL, USA) and the differences between the two groups were
determined by Student’s t-test. P<0.05 was considered to
indicate a statistically significant difference.
Results
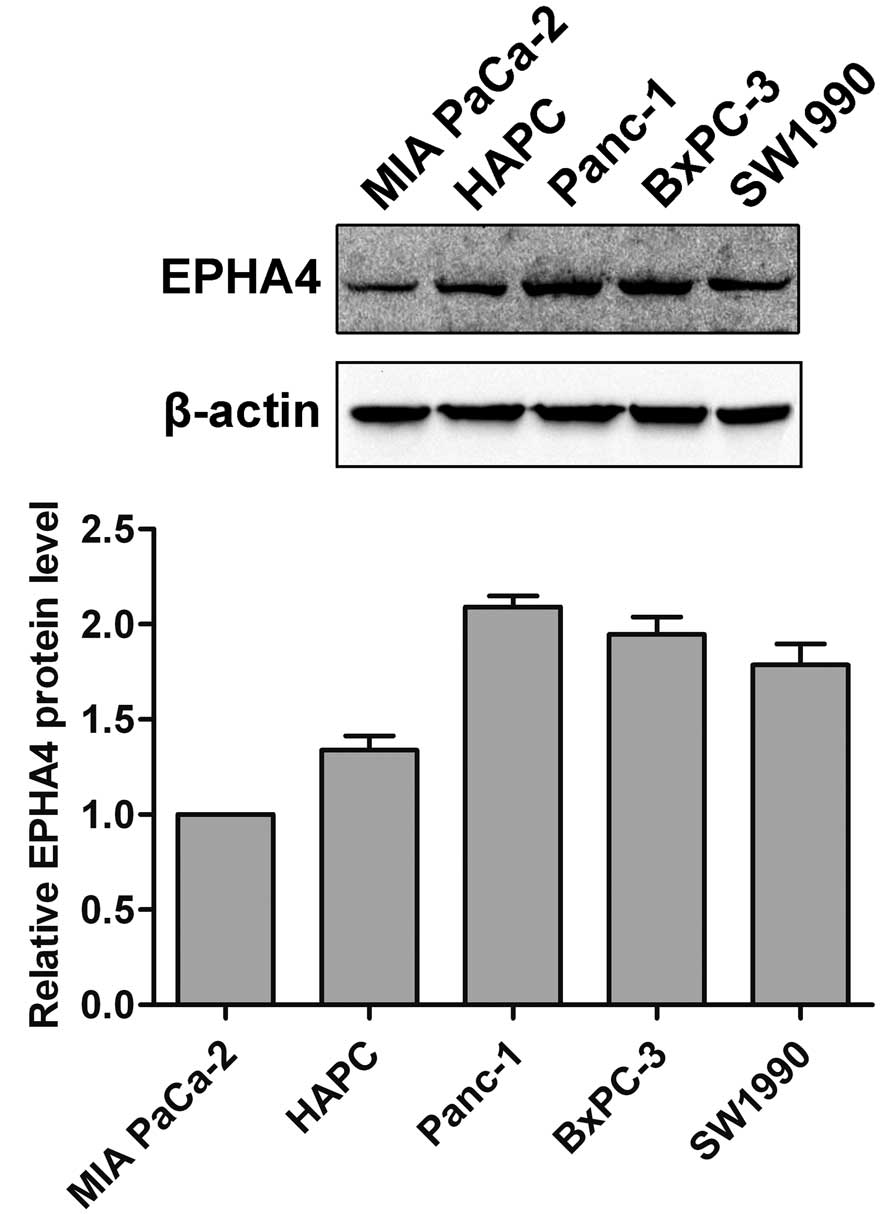
Expression of EPHA4 in pancreatic cancer
cells
The expression of EPHA4 in pancreatic cancer cell
lines (MIA PaCa-2, HAPC, SW1990, BxPC-3 and Panc-1) was examined by
western blotting. As shown in Fig.
1, EPHA4 was significantly expressed in all the examined
pancreatic cancer cells.
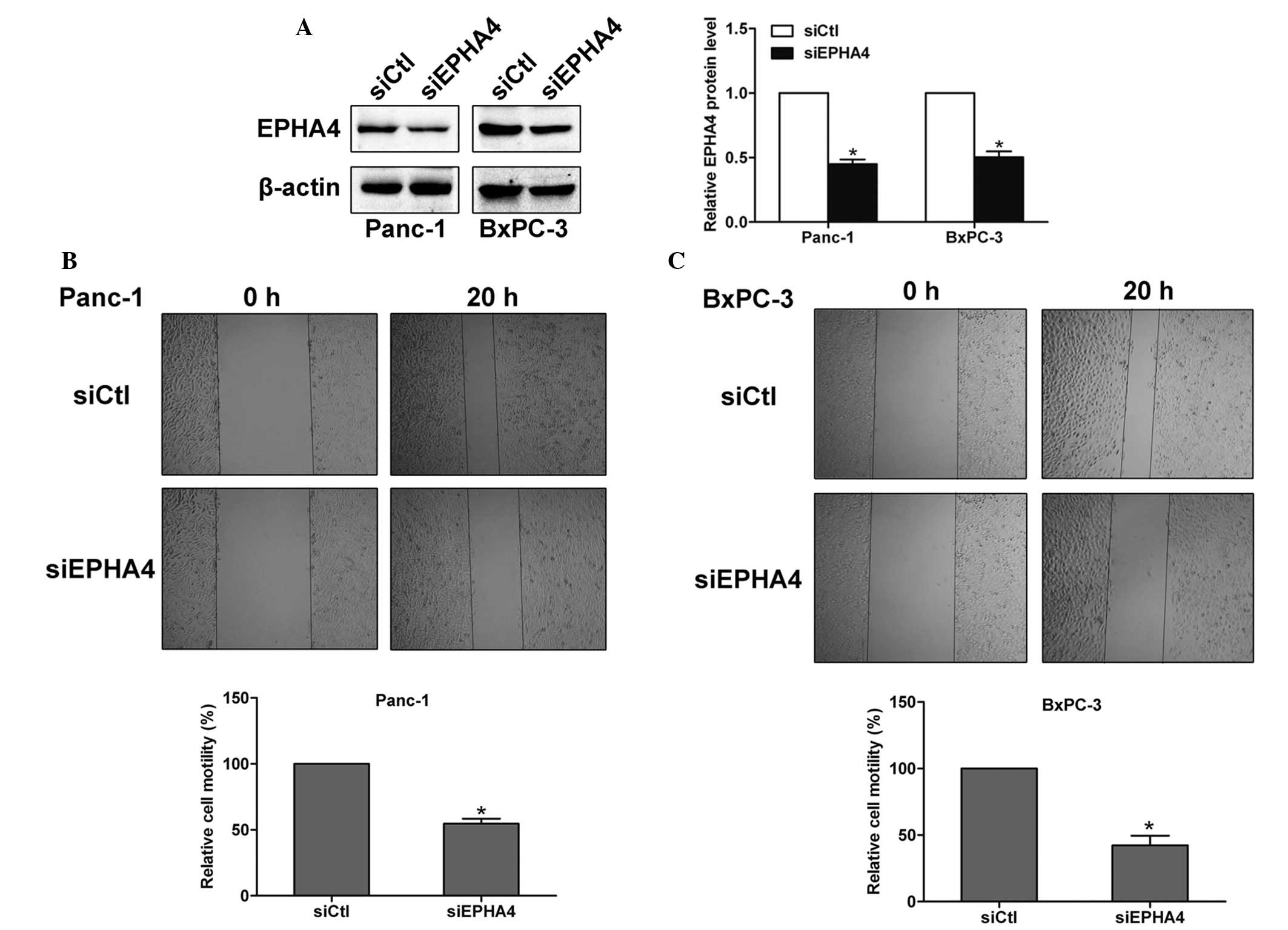
EPHA4 enhances the motility of pancreatic
cancer cells
To explore the function of EPHA4 in the pancreatic
cancer cells, siRNA was introduced to silence the expression of
EPHA4 in Panc-1 and BxPC-3 cells. The knockdown efficiency was
examined by western blotting and the EPHA4 siRNA-transfected cells
were shown to express lower levels of the EPHA4 protein (Fig. 2A). Next, a wound healing assay was
performed in Panc-1 and BxPC-3 cells and the results showed that
the motility of EPHA4 siRNA-transfected cells was significantly
reduced compared with the control siRNA cells, which indicated that
EPHA4 enhances the motility of pancreatic cancer cells (Fig. 2B and C).
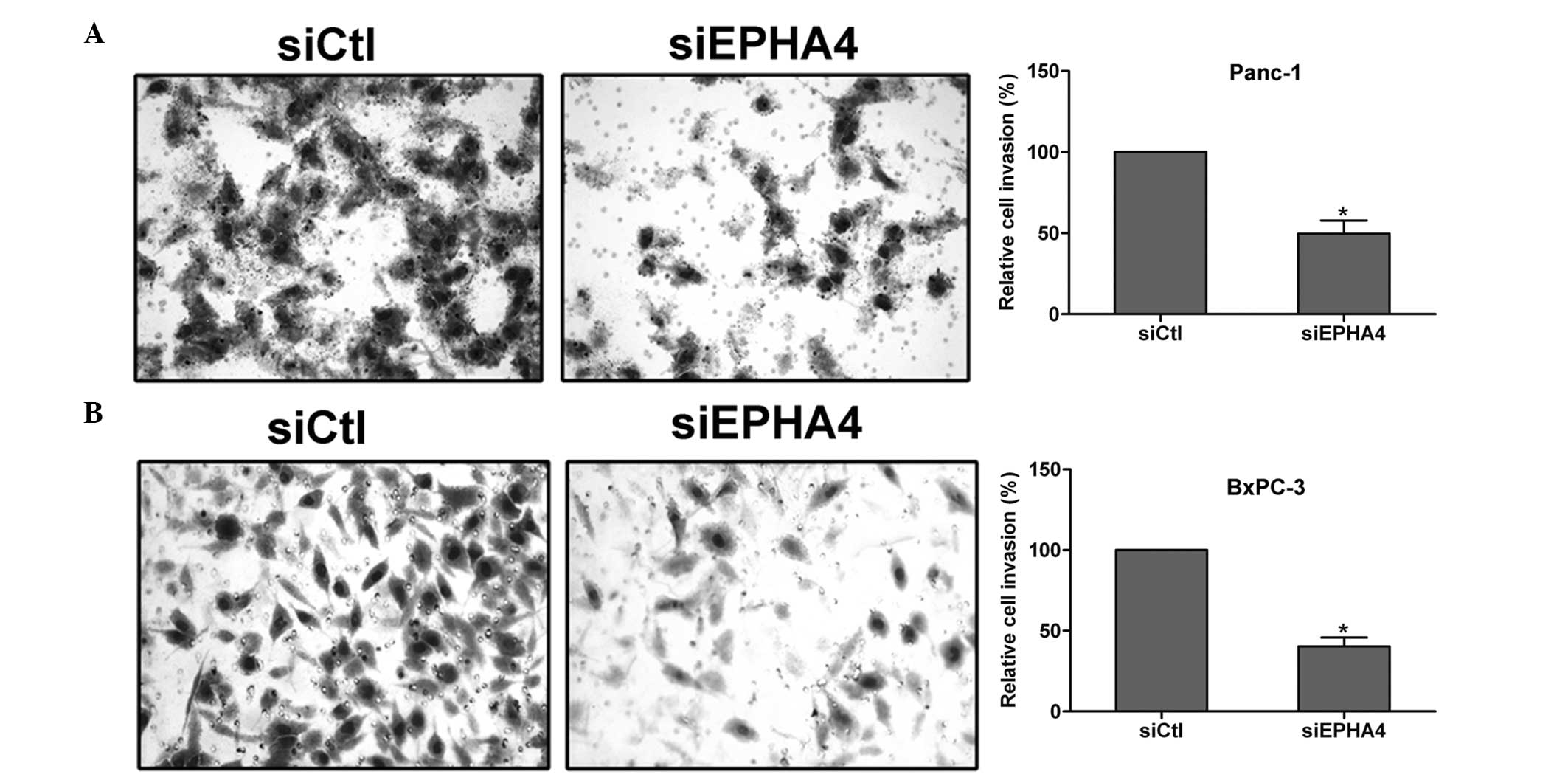
EPHA4 promotes the invasion of pancreatic
cancer cells
Next, an invasion assay was performed to determine
the function of EPHA4 in pancreatic cancer cell invasion. Notably,
the knockdown of EPHA4 was found to suppress the invasion of Panc-1
and BxPC-3 cells, which suggested that EPHA4 is involved in the
invasion of pancreatic cancer cells (Fig. 3A and B).
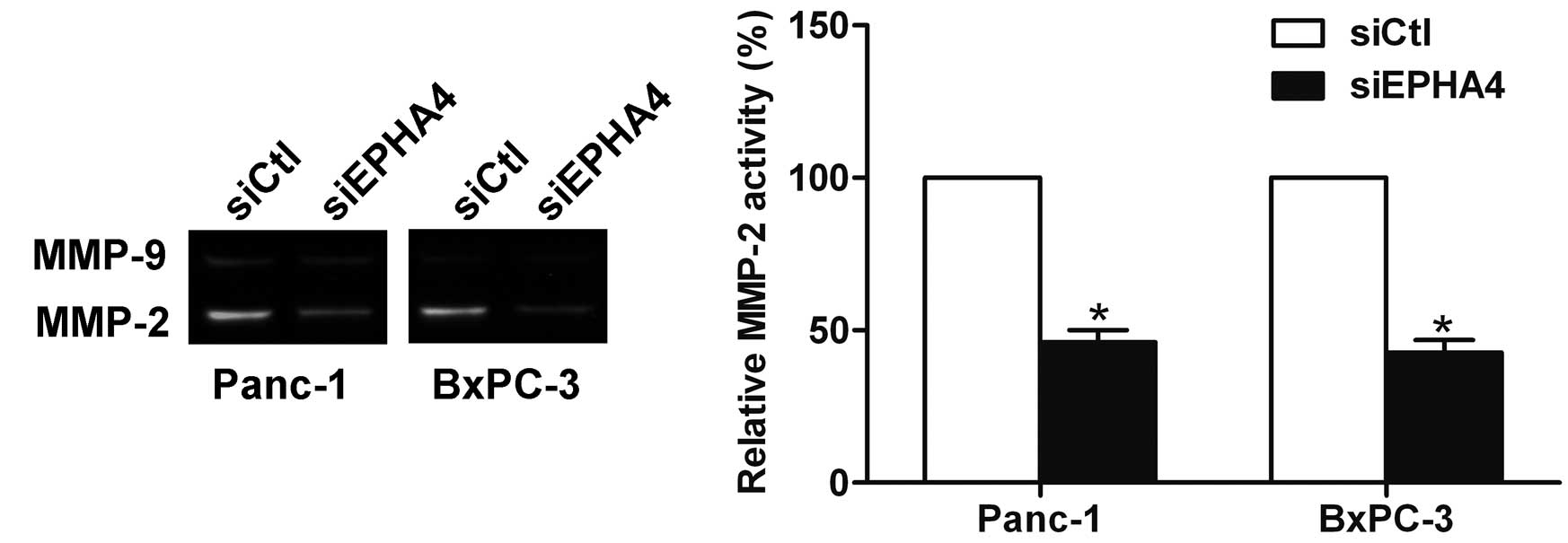
EPHA4 is involved in the regulation of
matrix metalloproteinase (MMP)-2 activity
MMP-2 and MMP-9 are crucial in ECM degradation and
are essential for the invasion and metastasis of pancreatic cancer
(10). The knockdown of EPHA4 was
found to inhibit the activity of MMP-2 by gelatin zymography, which
suggested that EPHA4 is involved in the regulation of MMP-2
activity (Fig. 4).
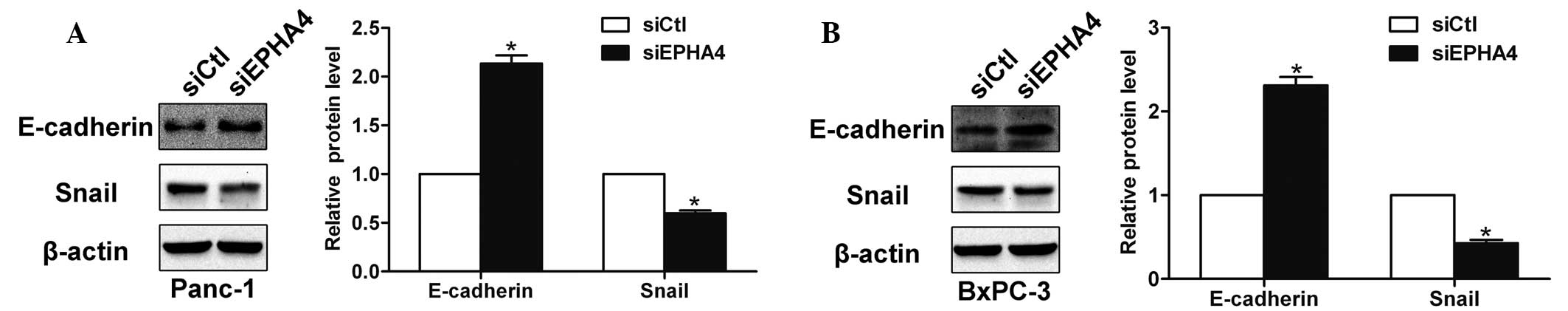
EPHA4 affects the expression of
E-cadherin and Snail
E-cadherin is an important cell adhesion molecule
that is regulated by zinc-finger transcription factors, such as
Snail, and is involved in epithelial-mesenchymal transition (EMT)
and metastatic processes (11).
Following the knockdown of EPHA4, the expression of E-cadherin was
found to increase by western blotting, whereas the level of Snail
was found to decrease (Fig. 5A and
B). Thus, the results indicated that EPHA4 affects the
expression of E-cadherin and Snail in pancreatic cancer cells.
Discussion
As the largest family of receptor tyrosine kinases,
the EPH receptors have been found to be overexpressed in a number
of human cancers (12,13) and studies have shown that EPH
receptors and their ligands are crucial in tumor progression
(14,15). In addition, it has been reported
that EPH receptors affect the cell-ECM attachment, thereby
contributing to the invasion and metastasis of cancer (16). The current study demonstrated that
EPHA4, a member of the EPH receptors, may promote the motility and
invasion of pancreatic cancer cells. The knockdown of EPHA4 was
found to suppress the activity of MMP-2, as well as increase the
expression of E-cadherin and decrease the expression of Snail.
Upregulation of EPHA4 has been found in various
types of tumors, such as gastric, colorectal and pancreatic cancer
(8,9,17), and
high expression of EPHA4 has been found to correlate with tumor
progression, including the invasion, pathological stage and distant
metastasis (18). Furthermore,
overexpression of EPHA4 promotes the growth of pancreatic cancer
cells (19) and enhances the
proliferation and migration of glioma cells (20). The present study revealed that EPHA4
was greatly expressed in the pancreatic cancer cells. In addition,
the knockdown of EPHA4 by siRNA inhibited the motility and invasion
of pancreatic cancer cells, which indicated the involvement of
EPHA4 in the motility and invasion of pancreatic cancer cells.
The MMPs are crucial enzymes for the degradation of
the ECM and are involved in cancer cell invasion and metastasis
(21). Furthermore, it has been
reported that the overexpression of EPHA2 upregulates the
expression of MMP-2 in Capan2 pancreatic ductal adenocarcinoma
cells (22). The results of the
present study demonstrated that EPHA4 has the ability to regulate
the activity of MMP-2 in Panc-1 and BxPC-3 cells. As a cell
adhesion molecule, E-cadherin can be negatively regulated by Snail
and, therefore, acts as a crucial marker in EMT and the invasion of
pancreatic cancer (23). Studies
have found that EPHA4 mediates the EMT process of human
hepatocellular carcinoma by downregulating the expression of
E-cadherin and vimentin (24). The
results of the current study also showed that the knockdown of
EPHA4 may increase the expression of E-cadherin and decrease the
expression of Snail in Panc-1 and BxPC-3 cells, which indicated
that EPHA4 may be involved in the EMT process of pancreatic cancer
cells.
In conclusion, the current study demonstrated that
EPHA4 may promote the motility and invasion of pancreatic cancer
cells. Furthermore, these processes may involve the upregulation of
MMP-2 and Snail, as well as the downregulation of E-cadherin.
However, further investigation is required to determine the
signaling pathways by which EPHA4 enhances the motility and
invasion of pancreatic cells.
References
|
1
|
Bardeesy N and DePinho RA: Pancreatic
cancer biology and genetics. Nat Rev Cancer. 2:897–909. 2002.
View Article : Google Scholar
|
|
2
|
Curran S and Murray GI: Matrix
metalloproteinases: molecular aspects of their roles in tumour
invasion and metastasis. Eur J Cancer. 36:1621–1630. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Takeichi M: Cadherins in cancer:
implications for invasion and metastasis. Curr Opin Cell Biol.
5:806–811. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lugli A, Spichtin H, Maurer R, et al:
EphB2 expression across 138 human tumor types in a tissue
microarray: high levels of expression in gastrointestinal cancers.
Clin Cancer Res. 11:6450–6458. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Walker-Daniels J, Hess AR, Hendrix MJ and
Kinch MS: Differential regulation of EphA2 in normal and malignant
cells. Am J Pathol. 162:1037–1042. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Héroult M, Schaffner F and Augustin HG:
Eph receptor and ephrin ligand-mediated interactions during
angiogenesis and tumor progression. Exp Cell Res. 312:642–650.
2006.PubMed/NCBI
|
|
7
|
Gale NW, Holland SJ, Valenzuela DM, et al:
Eph receptors and ligands comprise two major specificity subclasses
and are reciprocally compartmentalized during embryogenesis.
Neuron. 17:9–19. 1996. View Article : Google Scholar
|
|
8
|
Oki M, Yamamoto H, Taniguchi H, Adachi Y,
Imai K and Shinomura Y: Overexpression of the receptor tyrosine
kinase EphA4 in human gastric cancers. World J Gastroenterol.
14:5650–5656. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Oshima T, Akaike M, Yoshihara K, et al:
Overexpression of EphA4 gene and reduced expression of EphB2 gene
correlates with liver metastasis in colorectal cancer. Int J Oncol.
33:573–577. 2008.PubMed/NCBI
|
|
10
|
Bloomston M, Zervos EE and Rosemurgy AS
II: Matrix metalloproteinases and their role in pancreatic cancer:
A review of preclinical studies and clinical trials. Ann Surg
Oncol. 9:668–674. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guarino M, Rubino B and Ballabio G: The
role of epithelial-mesenchymal transition in cancer pathology.
Pathology. 39:305–318. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dodelet VC and Pasquale EB: Eph receptors
and ephrin ligands: embryogenesis to tumorigenesis. Oncogene.
19:5614–5619. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lu Z, Zhang Y, Li Z, et al: Overexpression
of the B-type Eph and ephrin genes correlates with progression and
pain in human pancreatic cancer. Oncol Lett. 3:1207–1212.
2012.PubMed/NCBI
|
|
14
|
Pasquale EB: Eph receptors and ephrins in
cancer: bidirectional signalling and beyond. Nat Rev Cancer.
10:165–180. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tan P, Liu Y, Yu C, et al: EphA2 silencing
in nasopharyngeal carcinoma leads to decreased proliferation,
invasion and increased sensitization to paclitaxel. Oncol Lett.
4:429–434. 2012.PubMed/NCBI
|
|
16
|
Singh A, Winterbottom E and Daar IO:
Eph/ephrin signaling in cell-cell and cell-substrate adhesion.
Front Biosci (Landmark Ed). 17:473–497. 2012. View Article : Google Scholar
|
|
17
|
Giaginis C, Tsourouflis G,
Zizi-Serbetzoglou A, et al: Clinical significance of ephrin
(eph)-A1, -A2, -a4, -a5 and -a7 receptors in pancreatic ductal
adenocarcinoma. Pathol Oncol Res. 16:267–276. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Miyazaki K, Inokuchi M, Takagi Y, Kato K,
Kojima K and Sugihara K: EphA4 is a prognostic factor in gastric
cancer. BMC Clin Pathol. 13:192013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Iiizumi M, Hosokawa M, Takehara A, et al:
EphA4 receptor, overexpressed in pancreatic ductal adenocarcinoma,
promotes cancer cell growth. Cancer Sci. 97:1211–1216. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fukai J, Yokote H, Yamanaka R, Arao T,
Nishio K and Itakura T: EphA4 promotes cell proliferation and
migration through a novel EphA4-FGFR1 signaling pathway in the
human glioma U251 cell line. Mol Cancer Ther. 7:2768–2778. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Deryugina EI and Quigley JP: Matrix
metalloproteinases and tumor metastasis. Cancer Metastasis Rev.
25:9–34. 2006. View Article : Google Scholar
|
|
22
|
Duxbury MS, Ito H, Zinner MJ, Ashley SW
and Whang EE: Ligation of EphA2 by Ephrin A1-Fc inhibits pancreatic
adenocarcinoma cellular invasiveness. Biochem Biophys Res Commun.
320:1096–1102. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
von Burstin J, Eser S, Paul MC, et al:
E-Cadherin regulates metastasis of pancreatic cancer in vivo and is
suppressed by a SNAIL/HDAC1/HDAC2 repressor complex.
Gastroenterology. 137:361–371. 2009.PubMed/NCBI
|
|
24
|
Yan Y, Luo YC, Wan HY, et al: MicroRNA-10a
is involved in the metastatic process by regulating Eph tyrosine
kinase receptor A4-mediated epithelial-mesenchymal transition and
adhesion in hepatoma cells. Hepatology. 57:667–677. 2013.
View Article : Google Scholar : PubMed/NCBI
|