Introduction
Shikonin, a naphthoquinone pigment, is the primary
component of root extracts from Lithospermum erythrorhizon.
Shikonin and its analogs have been used for the treatment of burns,
measles, sore throat, macular eruption and carbuncles (1). Shikonin and its analogs have also been
shown to possess in vitro and in vivo anticancer
activity against various types of cancer (2–4).
Shikonins can inhibit tumor growth and prolong the lifespan of
tumor-bearing mice (5) and patients
with lung cancer (6). Shikonins
mediate apoptosis through multiple mechanisms, including induction
of the generation of reactive oxygen species (ROS) (7,8) and
cell cycle arrest via a caspase-dependent mechanism (9). In addition, shikonin exhibits
antiangiogenic activity (10) and
can also regulate the activity of topoisomerase I and II, leading
to DNA cleavage (11,12).
Along with surgery and chemotherapy, radiotherapy is
one of the most significant modalities for cancer treatment. The
use of radiotherapy is primarily limited by intrinsic or acquired
resistance to ionizing radiation (IR). In an effort to overcome the
radioresistance of cancer cells to improve radiotherapy, a variety
of chemical compounds have been tested for their radiosensitizing
effects. Curcumin (13–15), resveratrol (16), genistein (17–19)
and flavopiridol (20) have been
shown to exhibit radiosensitizing effects on a variety of cancer
cells. IR kills cancer cells by inducing DNA damage and generating
ROS, which in turn induces further damage of biomolecules,
including DNA. The accumulation of ROS also induces the
deregulation of the apoptotic signaling pathway, ultimately leading
to apoptosis. The radiosensitizing effect of compounds is often
associated with ROS upregulation, indicating that the ROS-mediated
mechanism may be a significant target for achieving biological
enhancement of the effects of radiotherapy (21).
Although shikonin and its derivatives have been
reported to have potential anticancer activity, they have not been
examined for their effects on radiotherapy. The present study
examined whether shikonin and its analog,
β,β-dimethylacrylshikonin, exhibit radiosensitizing effects, and
investigated the possible utilization of these compounds as
radiotherapy-enhancing agents.
Materials and methods
Cell culture
The HCT-116, H460 and A549 cells obtained from the
American Type Culture Collection (Manassas, VA, USA) were grown in
RPMI-1640 medium (Mediatech, Manassas, VA, USA) and LN428 was grown
in MEM (Mediatech) supplemented with 10% fetal bovine serum (Tissue
Culture Biologicals, Los Alamitos, CA, USA),
penicillin/streptomycin (1X; PAA Laboratories GmbH, Morningside,
QLD, Australia) and mycokill (5 mg/ml, PAA laboratories). Cells
were maintained at 37°C in a humidified incubator containing 5%
CO2. Subconfluent cells were treated with shikonin
(Biomol Research Laboratories, Inc., Plymouth Meeting, PA, USA) or
its analogue, β,β-dimethylacrylshikonin (Tokyo Chemical Industry
Co., Ltd., Tokyo, Japan) for 4 h followed by IR treatment at 5 Gy
for the indicated time.
Growth inhibition assay
Cells were seeded in 96-well plates and pre-treated
with shikonins for 4 h, and subsequently exposed to IR at the
indicated doses. The number of viable cells was determined using
the
3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium,
inner salt (MTS) reduction assay (CellTiter 96 AQueous
Non-Radioactive Cell Proliferation assay; Promega, Madison, WI,
USA) according to the manufacturer’s instructions. The experiments
were carried out in triplicate.
Colony formation assay
Cells were seeded in 60-mm dishes at a density of
500 cells per dish. Shikonins were added to each dish 4 h prior to
IR treatment. After 14 days, media were removed, then cells were
stained with 1% crystal violet (Sigma-Aldrich, St. Louis, MO, USA)
in 10% ethanol and counted. The experiments were carried out in
triplicate.
Apoptosis assay
Cells were pre-treated with shikonins (0.5 μM) for 4
h and irradiated. The media were then exchanged with fresh media
and the cells were incubated for 72 h, followed by Annexin
V/propidium iodide (PI)-double staining using Annexin V-FITC
Apoptosis Detection kit I (BD Biosciences, Franklin Lakes, NJ,
USA). For the ROS scavenging experiment, N-acetylcysteine
(NAC; Sigma-Aldrich) was pre-treated at 1 mM for 2 h prior to the
treatment with shikonins. Cell death was analyzed using a
fluorescence-activated cell sorting (FACS)Calibur apparatus (BD
Biosciences).
Determination of intracellular ROS
level
Intracellular ROS production was measured by
staining cells with the fluorescent probe,
2′-7′-dichlorofluoresceindiacetate (DCF-DA; Invitrogen Life
Technologies, Carlsbad, CA, USA). The cells that were treated with
a combination of shikonins and IR were incubated with DCF-DA at 1
μM for 30 min. The changes in fluorescence intensity were monitored
by flow cytometry using a FACSCalibur apparatus (BD
Biosciences).
Western blot analysis
Following drug treatment, cell lysates were prepared
for western blot analysis. Proteins were resolved by
SDS-polyacrylamide gel electrophoresis (Bio-Rad, Hercules, CA, USA)
and transferred to nitrocellulose membrane (Whatman, Pittsburgh,
PA, USA). Subsequent to the transfer, the membranes were blocked in
5% skimmed milk in 10 mM Tris-HCl (pH 8.0), 150 mM NaCl and 0.05%
Tween-20 (TBST; Amresco, Solon, OH, USA) for 30 min, and then
incubated with a primary antibody in 5% skimmed milk in TBST for 2
h at room temperature. The membranes were washed three times with
TBST and incubated for 1 h in TBST containing horseradish
peroxidase-linked anti-immunoglobulin G (IgG). Following three
washes in TBST, immunoreactive products were detected by
chemiluminescence (ECL Plus; Amersham Pharmacia Biotech,
Piscataway, NJ, USA). Mouse monoclonal anti-γH2AX and anti-β-actin
antibodies were purchased from Millipore (Billerica, MA, USA) and
Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA),
respectively.
Ionizing irradiation of cells
Cells were exposed to γ-rays with a 137Cs
γ-ray source (Atomic Energy of Canada, Ltd., Ontario, ON, Canada)
and a dose rate of 2.6 Gy/min.
Immunofluorescence microscopy
The cells were seeded on a cover glass in 24-well
plates. The media were removed and carefully rinsed with
phosphate-buffered saline (PBS) 30 min following the treatments
with shikonins and IR. The cells were fixed with 3.7%
paraformaldehyde in PBS for 10 min and washed twice with PBS. Cells
were permeabilized for 10 min with 0.1% Triton X-100 followed by
blocking with CAS-block (Invitrogen Life Technologies) for 30 min.
Cells were then stained by incubating with mouse monoclonal
anti-γH2AX antibody (1:500 dilution) followed by goat anti-mouse
IgG-Alexa Fluor555 (Invitrogen Life Technologies) (1:1000
dilution). 4,6-Diamidino-2-phenylindole (DAPI) (50 μg/ml) was added
to the secondary antibody mixture to visualize the nuclei.
Fluorescence images were obtained using a LSM710 confocal
microscope (Carl Zeiss Group, Jena, Germany).
Tumor xenograft growth in athymic
mice
Athymic nude mice (4-week-old males) were obtained
from Orientbio, Inc., (Seoul, South Korea) and were maintained in a
laminar air-flow cabinet under specific pathogen-free conditions.
The human colon cancer HCT-116 xenograft mouse model was
established by subcutaneous inoculation of 2×106 cells
into the right hind leg. When the tumor size reached 150
mm3, the mice were randomly divided into six groups
(seven mice per group) and treated with either the vehicle (10%
dimethylsulfoxide in PBS) or shikonins (2.0 mg/kg) in the presence
or absence of IR. Two days after treatment, the second injection
was prepared. Locoregional irradiation was applied in single 8-Gy
doses using a Co-60 irradiator (Theratron 780; Atomic Energy of
Canada). Two perpendicular diameters of tumors were measured twice
a week with a caliper square by the same investigator, and the
tumor volume was calculated using the following equation: Tumor
volume (V) mm3 = (smaller diameter)2 ×
(larger diameter) × (π/6). The experiment was terminated when the
tumor volume in the control group reached 3000 mm3. All
animal protocols were reviewed using the Good Laboratory Practice
guidelines of the Radiotherapy Research Center, Korea Institute of
Radiological and Medical Sciences (Seoul, Korea). The use of these
animals and the experimental procedures were approved by the
Institutional Animal Care and Use Committee of the Korea Institute
of Radiological and Medical Sciences.
Statistical analysis
All data were plotted in terms of mean ± standard
error of the mean values. Statistical analysis was assessed using a
parametric repeated-measures one-way analysis of variance followed
by Tukey’s multiple comparison test (Graph Pad version 3; San
Diego, CA, USA). A value of P<0.05 was considered to indicate a
statistically significant difference.
Results
Shikonins sensitize cancer cell lines to
IR
To investigate the effect of shikonin analogs on the
cancer cell response to IR, shikonin and its analog
β,β-dimethylacrylshikonin were selected for the present study and
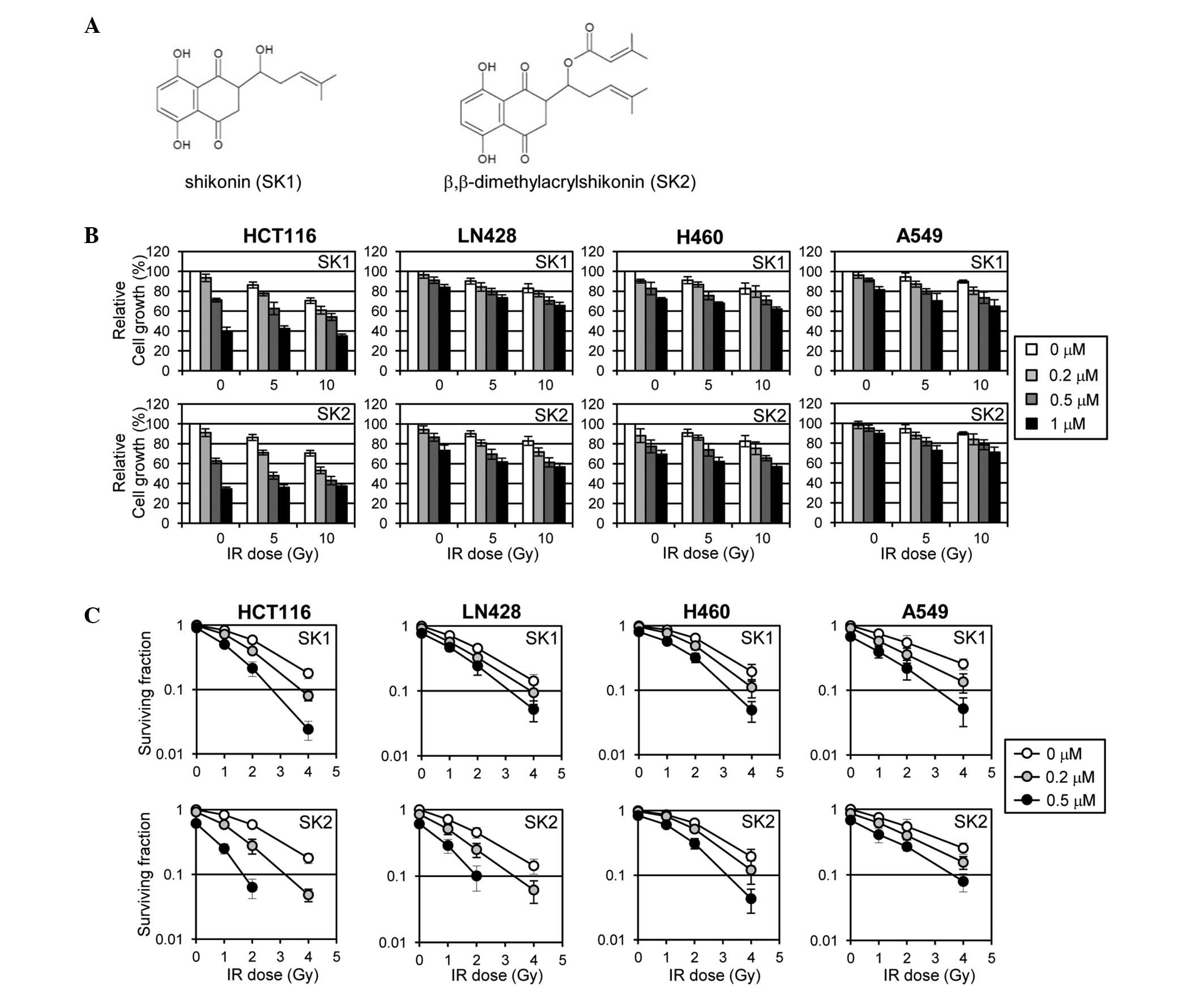
their chemical structures are shown in Fig. 1A. First, the effect of shikonins on
the proliferation of cancer cells when used alone or in combination
with IR was determined. For this experiment, various cancer cell
lines, including HCT-116 (colon cancer), LN428 (glioma), H460 (lung
cancer) and A549 (lung cancer) cells, were used. The cells were
pre-treated with shikonins for 4 h and irradiated at the indicated
doses in Fig. 1B. The viability of
cells was determined using the MTS assay. Shikonins inhibited the
overall proliferation of the cell lines in a dose-dependent manner
and exhibited additional effects when combined with IR (Fig. 1B). Among the cell lines examined,
HCT-116 was the most sensitive to shikonin treatment with respect
to inhibition of proliferation, as determined by the MTS assay.
Subsequently, the effect of shikonins on the cellular response to
IR was evaluated by determining clonogenic cell survival following
IR treatment through a colony formation assay. Shikonin exhibited a
moderate radiosensitizing effect for HCT-116, but minor effects for
the other cell lines, while the radiosensitizing effect of
β,β-dimethylacrylshikonin was considerable for HCT-116 and LN428,
but minor for H460 and A549 (Fig.
1C). Overall, these data show that shikonins sensitize HCT-116
cells more efficiently to IR treatment and, therefore, the HCT-116
cells were used for further in-depth study of the radiosensitizing
effects of shikonins.
Shikonins enhance IR-induced
apoptosis
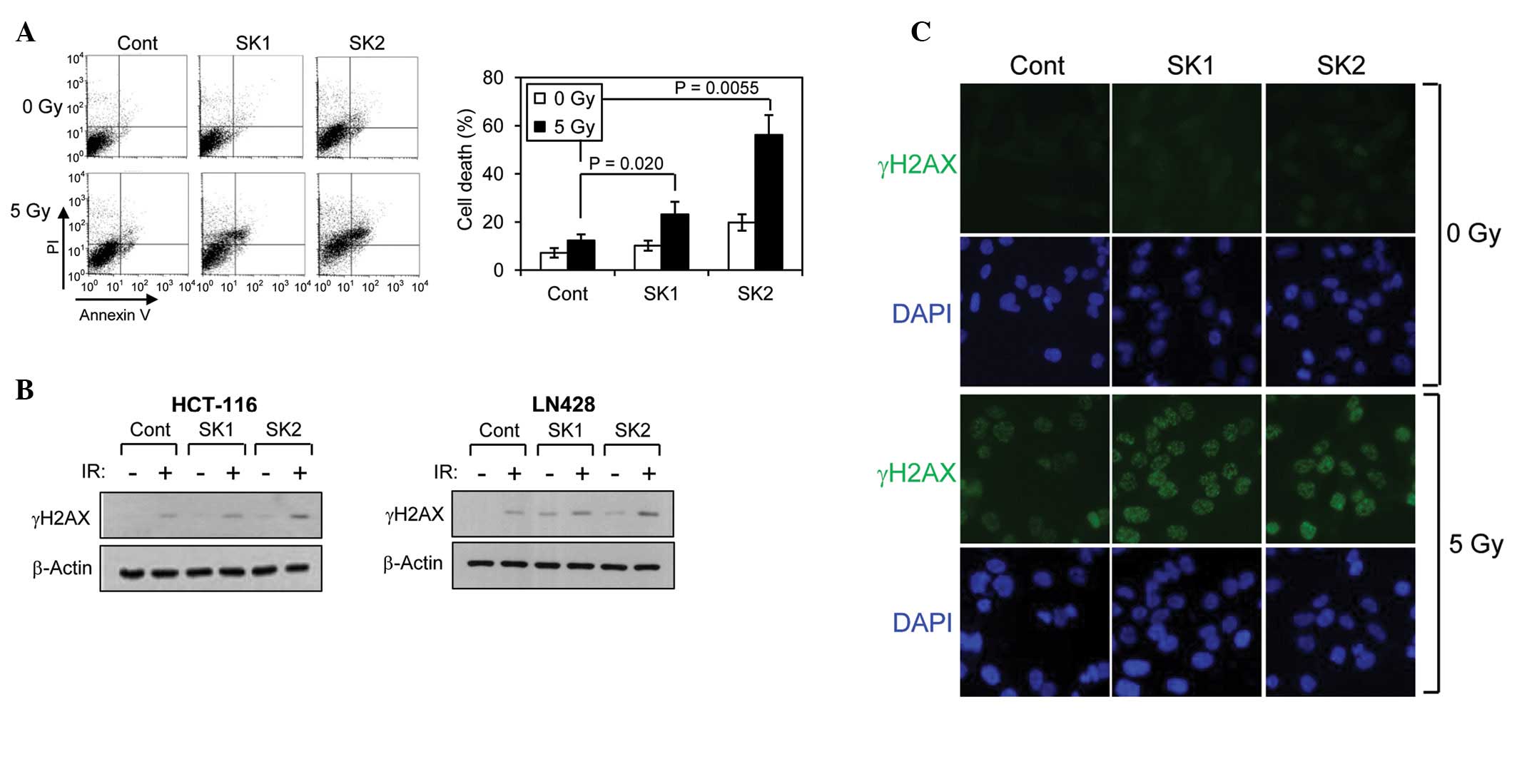
The induction of apoptosis in HCT-116 cells was
analyzed following combined treatment of shikonins with IR. The
cells were irradiated following pre-treatment or no pre-treatment
with shikonins and were analyzed for apoptosis by Annexin V/PI
staining at 72 h following irradiation. The cells that stained
negative for Annexin V and PI were assigned as undamaged live
cells. Shikonin induced marginal cell death, and the extent of
further enhancement of cell death by combination with IR was not
significant (Fig. 2A). By contrast,
treatment with β,β-dimethylacrylshikonin alone induced significant
cell death, and further enhancement of cell death was observed when
combined with IR. These results indicate that
β,β-dimethylacrylshikonin is extremely effective and more effective
compared with shikonin in rendering HCT-116 cells more susceptible
to IR-induced cell death. It has great potential as a
radiosensitizing agent.
Shikonins enhance IR-induced DNA
damage
The effect of shikonins on the extent of IR-induced
DNA damage was examined by determining the level of the
phosphorylated histone H2AX (γH2AX), a well-known marker for DNA
double-strand breaks. Single treatment with either shikonin or
β,β-dimethylacrylshikonin caused weak accumulation of γH2AX in
HCT-116 and LN428 cells, indicating that shikonin and
β,β-dimethylacrylshikonin can individually induce DNA damage to a
certain extent. However, when the cells were treated with a
combination of shikonins and IR, only β,β-dimethylacrylshikonin
strongly enhanced further IR-induced γH2AX increases (Fig. 2B). The effect of shikonins on the
induction of DNA damage was also assessed by visualizing γH2AX foci
with immunofluorescence microscopy. Treatment with either of the
shikonins increased the formation of γH2AX foci, but
β,β-dimethylacrylshikonin-treated cells showed a stronger γH2AX
signal intensity compared with shikonin-treated cells following
exposure to IR (Fig. 2C). These
results indicate that β,β-dimethylacrylshikonin strongly
potentiates the induction of DNA damage by IR treatment and that
this potentiation is greater compared with that observed with
shikonin.
Combined treatment of shikonins and IR
causes ROS accumulation
ROS generation is one of the primary mechanisms by
which IR kills cells, and it has been reported that shikonin causes
apoptosis through an ROS/c-Jun N-terminal kinase-mediated signaling
pathway in the breakpoint cluster region/Abelson-positive chronic
myelogenous leukemia cells (8).
Therefore, it was initially postulated that shikonins modulate the
cellular response to IR through the regulation of ROS levels. To
investigate this possibility, the effect of a combined treatment of
shikonins and IR at the intracellular ROS level was examined. A
single IR treatment (5 Gy) with either of the shikonins caused a
minor increase (~15–25%) in ROS levels in the HCT-116 cells.
However, IR treatment following pre-treatment with either of the
shikonins resulted in a significant increase (~80%) in ROS levels
(Fig. 3A). This synergistic effect
of combined treatment of shikonins and IR on ROS accumulation
indicates that shikonins may predispose cancer cells to accumulate
more ROS in response to IR treatment.
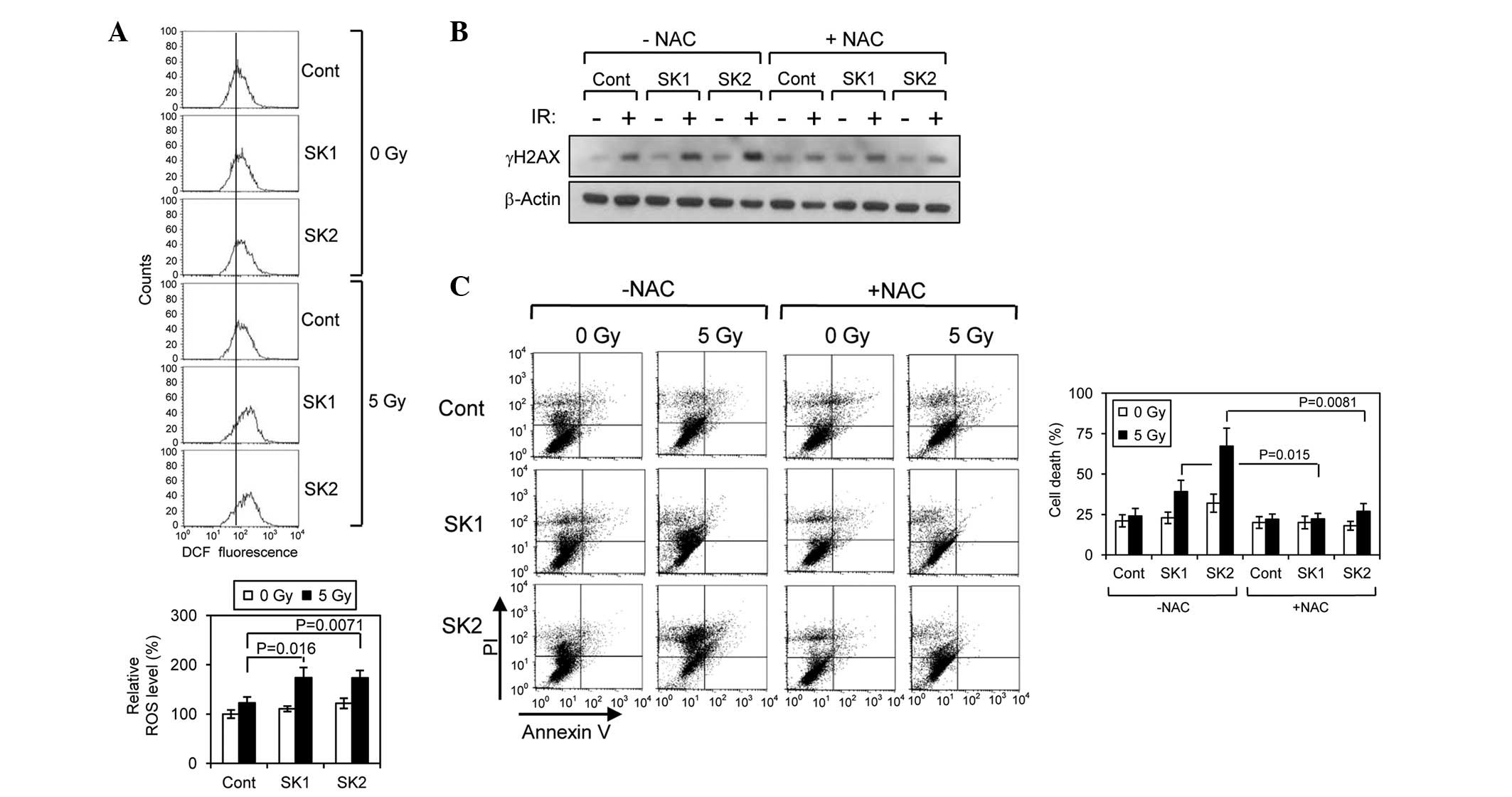 | Figure 3Shikonins enhance IR-induced apoptosis
through ROS upregulation. (A) Determination of the cellular ROS
level by staining with DCF-DA followed by FACS analysis. The cells
were treated with shikonins (0.5 μM) in the presence or absence of
IR treatment (5 Gy), and the ROS level was measured by FACS
analysis. (B) Suppression of the effect of shikonins on H2AX
phosphorylation by NAC pretreatment. (C) Suppression of the effect
of shikonins on IR-induced apoptosis by NAC pre-treatment. IR,
ionizing radiation; NAC, N-acetylcysteine; ROS, reactive
oxygen species; PI, propidium iodide; DCF-DA,
2′-7′-dichlorofluoresceindiacetate; FACS, fluorescence-activated
cell sorting; SK1, shikonin; SK2, β,β-dimethylacrylshikonin. |
ROS is involved in the synergistic effect
of shikonins on IR-induced DNA damage and cell death
The preceding observations that shikonins enhance
ROS accumulation, DNA damage and apoptosis indicate that ROS
accumulation may be responsible for the synergistic effect of
shikonins on DNA damage and subsequent cell death. To test this
hypothesis, the effect of pretreatment of cells with NAC on
ROS-induced DNA damage was examined, which can be assessed by
determining the level of γH2AX (Fig.
3B). NAC significantly attenuated the increase in the γH2AX
level induced by either of the shikonins. NAC also significantly
suppressed the synergistic effect of β,β-dimethylacrylshikonin on
IR-induced apoptosis (Fig. 3C).
These observations indicate that ROS accumulation plays a critical
role in the enhancement of IR-induced DNA damage and subsequent
apoptosis by β,β-dimethylacrylshikonin treatment.
β,β-Dimethylacrylshikonin potentiates the
antitumor effect of IR on tumor growth in the HCT-116 xenograft
mouse model
To validate the radiosensitizing effect of shikonins
in vivo, the HCT-116 xenografts in athymic nude mice were
established. Using the experimental procedure described in Fig. 4A, the change in tumor volume was
monitored twice a week following the combined treatment with the
shikonins and IR. While tumor growth was moderately suppressed by
IR alone, β,β-dimethylacrylshikonin completely retarded tumor
growth when coupled with IR treatment (Fig. 4B). The tumor size endpoint, which
was measured 30 days subsequent to IR treatment when the tumor
volume of the vehicle group reached 3000 mm3, also
manifested the strong effect of β,β-dimethylacrylshikonin acting
synergistically with IR to suppress tumor growth (Fig. 4C).
Discussion
Radiotherapy is one of the primary modalities in
cancer treatment and is generally used in combination with surgery
or chemotherapy (22). The use of
high-dose IR also inevitably causes damage to surrounding normal
tissues, necessitating the use of agents to sensitize cancer cells
to IR treatment, thereby allowing the use of lower doses of
radiation. In spite of numerous reports that have demonstrated the
antitumor effect of shikonins (2–4), the
potential applicability of shikonins as radiosensitizers has not
been fully examined. In an effort to identify novel
radiosensitizers, the effect of shikonin and its analog
β,β-dimethylacrylshikonin on the sensitivity of cancer cells to IR
treatment was examined.
IR-induced cell death was promoted by pre-treatment
with shikonin or more strongly with β,β-dimethylacrylshikonin.
Synergistic increases in intracellular ROS levels and DNA damage
accompanied the IR-sensitizing action of shikonins. It was also
found that the enhancement of IR-induced DNA damage and cell death
mediated by shikonins was abolished in the presence of the
antioxidant NAC. Since the generation of ROS is one of the primary
mechanisms by which IR induces DNA damage and kills cells, these
results indicate that further upregulation of ROS to intolerable
levels accounts for the radiosensitizing effects of shikonins. A
recent study using leukemia cells indicated that the cytotoxicity
of shikonin involves the disruption of mitochondrial function,
including ROS production and the inhibition of cytoskeleton
formation (23). Shikonin
immediately accumulates in the mitochondria and disrupts the
mitochondrial membrane potential, followed by the induction of
oxidative damage due to the generation of ROS. Several studies have
demonstrated the antitumor activity of β,β-dimethylacrylshikonin
via various signaling pathways, including the extracellular
signal-regulated kinase and Notch-1 pathways (24–26).
Notably, β,β-dimethylacrylshikonin has been reported to inhibit the
cellular growth of HCT-116 cells in vitro and of xenografts
in vivo (26). A previous
study showed that the induction of apoptosis by
β,β-dimethylacrylshikonin is associated with the upregulation of
the proapoptotic proteins, Bax and Bid, and a reduction in the
expression of the antiapoptotic proteins, B-cell lymphoma 2 (Bcl-2)
and Bcl-XL (27). This change in
the ratio of the proapoptotic/antiapoptotic Bcl-2 family of
proteins may have led to ROS generation. These observations are in
accordance with the overall results in the present study that
demonstrated the ROS-mediated radiosensitizing effect of
shikonins.
In summary, the present study has demonstrated
significant radiosensitizing activity of β,β-dimethylacrylshikonin
in vitro and in vivo. These findings indicate that
β,β-dimethylacrylshikonin is a promising candidate for a
radiosensitizing agent and may be exploited for the development of
a novel strategy for improving radiotherapy against cancerous
tumors.
Acknowledgements
This study was supported by a grant from the Nuclear
Research and Development Program (NRF-2010-0018713) and the Basic
Science Research Program (NRF-2011-0007381) through the National
Research Foundation of Korea funded by the Ministry of Education,
Science and Technology.
References
|
1
|
Chen X, Yang L, Oppenheim JJ and Howard
MZ: Cellular pharmacology studies of shikonin derivatives.
Phytother Res. 16:199–209. 2002. View
Article : Google Scholar
|
|
2
|
Gao D, Hiromura M, Yasui H and Sakurai H:
Direct reaction between shikonin and thiols induces apoptosis in
HL60 cells. Biol Pharm Bull. 25:827–832. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yao Y, Brodie AM, Davidson NE, Kensler TW
and Zhou Q: Inhibition of estrogen signaling activates the NRF2
pathway in breast cancer. Breast Cancer Res Treat. 124:585–591.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yingkun N, Lvsong Z and Huimin Y: Shikonin
inhibits the proliferation and induces the apoptosis of human HepG2
cells. Can J Physiol Pharmacol. 88:1138–1146. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sankawa U, Ebizuka Y, Miyazaki T, Isomura
Y and Otsuka H: Antitumor activity of shikonin and its derivatives.
Chem Pharm Bull (Tokyo). 25:2392–2395. 1977. View Article : Google Scholar
|
|
6
|
Guo XP, Zhang XY and Zhang SD: Clinical
trial on the effects of shikonin mixture on later stage lung
cancer. Zhong Xi Yi Jie He Za Zhi. 11:598–599. 1991.(In
Chinese).
|
|
7
|
Chen CH, Chern CL, Lin CC, Lu FJ, Shih MK,
Hsieh PY and Liu TZ: Involvement of reactive oxygen species, but
not mitochondrial permeability transition in the apoptotic
induction of human SK-hep-1 hepatoma cells by shikonin. Planta Med.
69:1119–1124. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mao X, Yu CR, Li WH and Li WX: Induction
of apoptosis by shikonin through a ROS/JNK-mediated process in
Bcr/Abl-positive chronic myelogenous leukemia (CML) cells. Cell
Res. 18:879–888. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu Z, Wu L, Li L, Tashiro S, Onodera S and
Ikejima T: p53-mediated cell cycle arrest and apoptosis induced by
shikonin via a caspase-9-dependent mechanism in human malignant
melanoma A375-S2 cells. J Pharmacol Sci. 94:166–176. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hisa T, Kimura Y, Takada K, Suzuki F and
Takigawa M: Shikonin, an ingredient of Lithospermum erythrorhizon,
inhibits angiogenesis in vivo and in vitro. Anticancer Res.
18:783–790. 1998.PubMed/NCBI
|
|
11
|
Fujii N, Yamashita Y, Arima Y, Nagashima M
and Nakano H: Induction of topoisomerase II-mediated DNA cleavage
by the plant naphthoquinones plumbagin and shikonin. Antimicrob
Agents Chemother. 36:2589–2594. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ahn BZ, Baik KU, Kweon GR, Lim K and Hwang
BD: Acylshikonin analogues: Synthesis and inhibition of DNA
topoisomerase-I. J Med Chem. 38:1044–1047. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chendil D, Ranga RS, Meigooni D,
Sathishkumar S and Ahmed MM: Curcumin confers radiosensitizing
effect in prostate cancer cell line PC-3. Oncogene. 23:1599–1607.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shi HS, Gao X, Li D, et al: A systemic
administration of liposomal curcumin inhibits radiation pneumonitis
and sensitizes lung carcinoma to radiation. Int J Nanomedicine.
7:2601–2611. 2012.PubMed/NCBI
|
|
15
|
Yang Y, Duan W, Liang Z, et al: Curcumin
attenuates endothelial cell oxidative stress injury through Notch
signaling inhibition. Cell Signal. 25:615–629. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zoberi I, Bradbury CM, Curry HA, Bisht KS,
Goswami PC, Roti Roti JL and Gius D: Radiosensitizing and
anti-proliferative effects of resveratrol in two human cervical
tumor cell lines. Cancer Lett. 175:165–173. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Papazisis KT, Zambouli D, Kimoundri OT, et
al: Protein tyrosine kinase inhibitor, genistein, enhances
apoptosis and cell cycle arrest in K562 cells treated with
gamma-irradiation. Cancer Lett. 160:107–113. 2000. View Article : Google Scholar
|
|
18
|
Akimoto T, Nonaka T, Ishikawa H, Sakurai
H, Saitoh JI, Takahashi T and Mitsuhashi N: Genistein, a tyrosine
kinase inhibitor, enhanced radiosensitivity in human esophageal
cancer cell lines in vitro: possible involvement of inhibition of
survival signal transduction pathways. Int J Radiat Oncol Biol
Phys. 50:195–201. 2001. View Article : Google Scholar
|
|
19
|
Hillman GG, Forman JD, Kucuk O, et al:
Genistein potentiates the radiation effect on prostate carcinoma
cells. Clin Cancer Res. 7:382–390. 2001.PubMed/NCBI
|
|
20
|
Jung C, Motwani M, Kortmansky J, et al:
The cyclin-dependent kinase inhibitor flavopiridol potentiates
gamma-irradiation-induced apoptosis in colon and gastric cancer
cells. Clin Cancer Res. 9:6052–6601. 2003.
|
|
21
|
Pelicano H, Carney D and Huang P: ROS
stress in cancer cells and therapeutic implications. Drug Resist
Updat. 7:97–110. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pavlov A, Pirogov A, Trachtenberg A,
Volkova M, Maximov T and Matveeva T: Results of combination
treatment of lung cancer patients: surgery plus radiotherapy and
surgery plus chemotherapy. Cancer Chemother Rep. 4:133–135.
1973.PubMed/NCBI
|
|
23
|
Wiench B, Eichhorn T, Paulsen M and
Efferth T: Shikonin directly targets mitochondria and causes
mitochondrial dysfunction in cancer cells. Evid Based Complement
Alternat Med. 2012:7260252012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shen XJ, Wang HB, Ma XQ and Chen JH:
β,β-Dimethylacrylshikonin induces mitochondria dependent apoptosis
through ERK pathway in human gastric cancer SGC-7901 cells. PLoS
One. 7:e417732012.
|
|
25
|
Zhen-Jun S, Yuan-Yuan Z, Ying-Ying F, et
al: β,β-Dimethylacrylshikonin exerts antitumor activity via Notch-1
signaling pathway in vitro and in vivo. Biochem Pharmacol.
84:507–512. 2012.
|
|
26
|
Fan Y, Jin S, He J, Shao Z, Yan J, Feng T
and Li H: Effect of β,β-dimethylacrylshikonin on inhibition of
human colorectal cancer cell growth in vitro and in vivo. Int J Mol
Sci. 13:9184–9193. 2012.
|
|
27
|
Fan Y, Jin S, He J, Shao Z, Yan J, Feng T
and Li H: Effect of β,β-dimethylacrylshikonin on inhibition of
human colorectal cancer cell growth in vitro and in vivo. Int J Mol
Sci. 13:9184–9193. 2012.
|