Introduction
Phosphatidylcholine (PC), an essential phospholipid
component of cell membranes, is synthesized in all nucleated cells
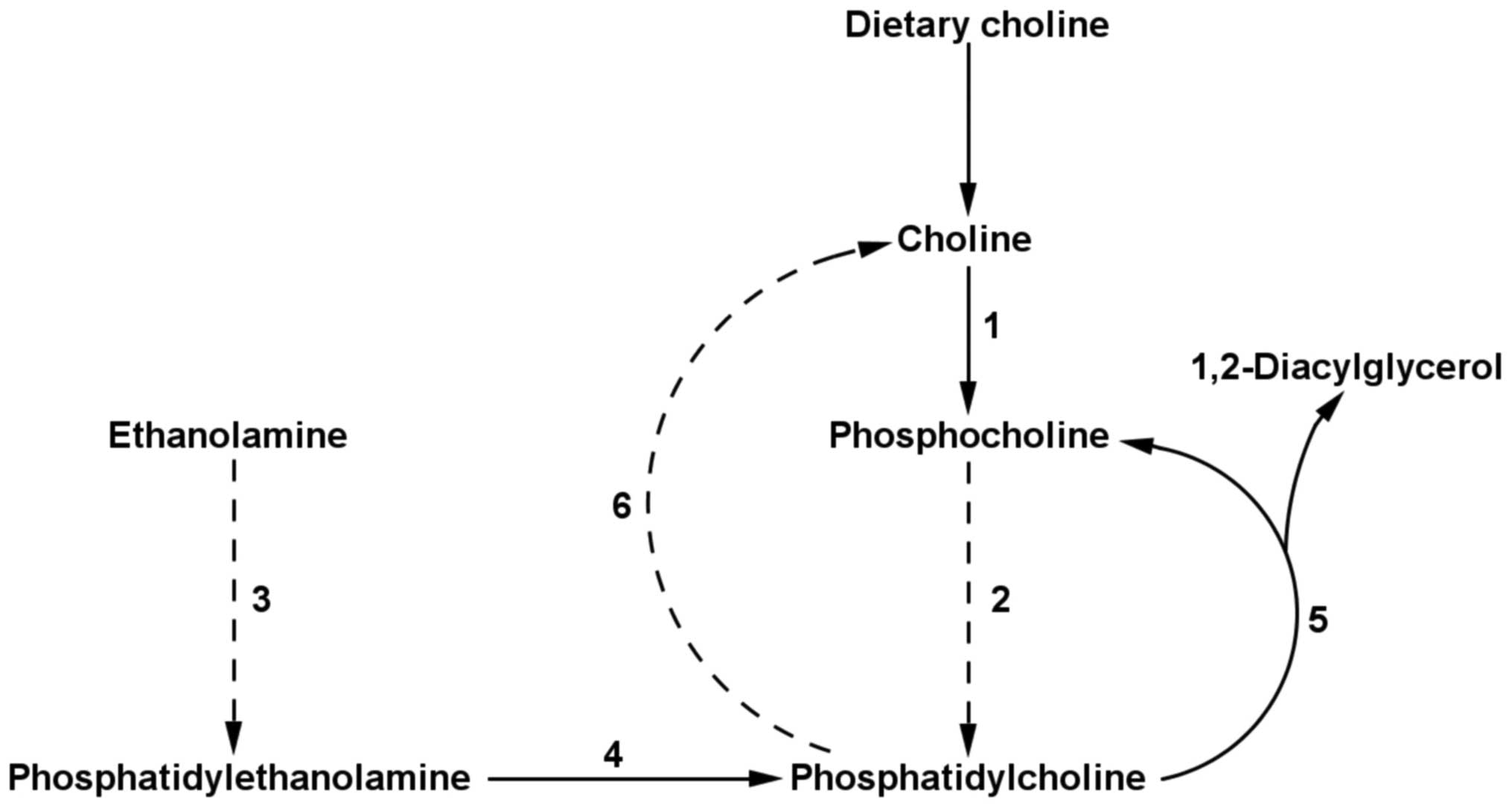
via the cytidine diphosphate (CDP)-choline pathway (1), which requires the uptake of dietary
choline. Alternatively, PC is synthesized by
phosphatidylethanolamine N-methyltransferase (PEMT; EC 2.1.1.17),
an enzyme responsible for the catalytic conversion of
phosphatidylethanolamine (PE) to PC (Fig. 1) (2). PC synthesis via the PEMT pathway is
dietary choline independent. PEMT expression and its
activity are highest in the liver, where the PEMT pathway accounts
for 30% of PC synthesis (1). PEMT
plays an important role in hepatocytes. For example,
PEMT−/− mice had a decreased PC/PE
ratio, which resulted in the loss of cell membrane integrity and
liver failure (3). All other
tissues exhibit low PEMT activity of, at most, 2% of the liver PEMT
activity (4).
Highly proliferating cancer cells must continuously
provide lipids for membrane assembly, signal transduction and
protein modification (5). The
biosynthetic and bioenergetic requirements for cell proliferation
and survival are supported by vigorous fatty acid metabolism
(6). Importantly, endogenously
synthesized fatty acids are not stored as triglycerides, but are
predominantly converted to phospholipids for membrane assembly
(6,7). This may require active PC-synthesis
pathways. PC concentration was found to be elevated in colorectal
cancer (8,9) and a higher expression of the
CDP-choline pathway enzymes was found in various types of human
cancers (8,10). With regards to the PEMT pathway,
data are inconsistent. Malignant neoplasm cells with metastases
were characterized by a higher PC/PE ratio compared with malignant
neoplasm cells without metastases (9), which suggests that higher PEMT
activity may be associated with tumor aggressiveness. On the other
hand, in rats with chemically induced hepatocarcinogenesis,
PEMT2 expression (an isoform exclusively located in
mitochondrial membranes of hepatocytes) was found to be diminished
in liver cancer lesions and, consistently, total PEMT activity was
decreased during different stages of tumor progression (11). Thus, PEMT involvement in cancer
requires further evaluation.
Fatty acid synthase (FASN; EC:2.3.1.85), the key
enzyme involved in neoplastic lipogenesis (12), is the sole protein capable of de
novo synthesis of long-chain fatty acids (7). As such, FASN is highly
expressed in various types of human cancers and in several cancer
types elevated FASN expression is linked to poor prognosis
(12–17). Consistently, in patients with
resectable non-small-cell lung cancer (NSCLC), higher FASN activity
in the cancer tissues (relative to the adjacent non-cancer lung
tissue) was associated with adverse outcomes and predicts shorter
patient survival (18).
Lipoprotein lipase (LPL; EC:3.1.1.34) is another
lipid-related enzyme associated with tumor growth. Following
parenchymal synthesis, LPL is translocated to the luminal
endothelial surface where it is responsible for intravascular
catabolism of triglyceride-rich lipoproteins and extracellular
supply of long-chain fatty acids, lipids and lipoproteins to
adjacent tissues. Thus, patients with resectable NSCLC had a higher
LPL activity in the cancer tissue than in the adjacent, healthy
non-cancer lung tissue (19), and
elevated LPL activity in resectable NSCLC tissue (relative to
adjacent non-cancer lung tissue) predicts shorter patient survival
independently of the standard prognostic factors (20).
To the best of our knowledge, PEMT has not yet been
studied in NSCLC tissue in the literature to date and has not been
studied concurrently with other lipid-related enzymes known to be
associated with tumor growth, such as FASN or LPL. In order to
study PEMT involvement in tumor growth, we hypothesized that
PEMT expression is increased in NSCLC tissue and that
increased gene expression acts as a predictor of shorter patient
survival. PEMT expression was determined in the same NSCLC
tissue samples as used for FASN and LPL analyses (18–20).
Materials and methods
Patient selection and tissue
sampling
Forty-two patients (median age of 62.5 years)
undergoing surgical removal of resectable, stages I, II and III
NSCLC tissues at University Medical Centre Ljubljana (Ljubljana,
Slovenia) were enrolled consecutively in this study. Samples of
lung cancer tissues and of adjacent, visually unaffected tissues
were cut from the resected lung within 15 min of surgery. Tumor
tissues were taken from the periphery, where the tumor was most
vigorous. Presumed normal tissues, taken from each subject as
control, were cut from the periphery of the resected lung, far away
from the tumor. Tissue samples were stored in liquid nitrogen until
analyzed. Staging was performed according to the
tumor-node-metastasis classification (21). Histological analysis of tumor tissue
samples was performed in accordance with the World Health
Organization histological classification (22). The study was approved by the
National Ethics Committee and informed consent was obtained from
all the patients participating in the study.
Patients were followed-up for four years. For the
first two years, patients were examined every three months and in
the second year, every six months. Clinical status and X-ray
imaging results were assessed routinely at the time of examination.
In progressively suspicious cases, bronchoscopy, computed
tomography scans and scintigraphy were performed as required. None
of the patients had undergone neoadjuvant therapy prior to surgery.
Patients with stage III NSCLC underwent postoperative
chemotherapy.
Quantification of PEMT expression in
tissue samples
RNA was isolated from tissue with RNeasy Mini kit
(Qiagen, Hilden, Germany) and reverse transcribed to cDNA using the
High-Capacity cDNA Archive kit (Applied Biosystems, Foster, CA,
USA) as previously described (23).
SYBR Green-based real-time polymerase chain reaction
(PCR) was used for the quantification of PEMT mRNA. A total of 2 μl
cDNA, previously diluted with purified water (dilution ratio 1:5)
(PCR Grade Water; Roche Diagnostics, Basel, Switzerland), was mixed
with 0.5 μl of primer [PEMT or β-glucuronidase (GUSB) QuantiTect
Primer Assays; Qiagen] and 2.5 μl FastStart Essential Green Master
(Roche Diagnostics). Two separate analyses of all the samples were
performed. In each analysis, all the reactions were run in
duplicate. Reaction mixtures were incubated in a
LightCycler® 480 Instrument (Roche Diagnostics) for 10
min at 95°C, followed by 40 cycles of 15 sec denaturation at 95°C
and 30 sec annealing/extension at 60°C.
GUSB was selected as the most appropriate internal
control among the 11 candidates available on TaqMan Human
Endogenous Control Plate (Applied Biosystems), as previously
described (23). The average PEMT
and GUSB PCR amplification efficiency (E) was considered in
calculating relative PEMT mRNA quantities (RQ) using the following
equation: RQ=(1+E) exp (−ΔΔCt).
Statistical analysis
Data are presented as the medians with 25th and 75th
percentiles and range values. Comparisons of parameters between
cancer and control tissues were made with the Wilcoxon matched-pair
test. Mann-Whitney or Pearson’s χ2 tests were used for
comparisons of the same parameter between groups. Associations
between variables were evaluated with Spearman rho analysis. Uni-
and multivariate Cox regression analyses were used to assess the
prognostic factors. Statistical analysis was performed with SPSS
software, v.17.0 (SPSS Inc., Chicago, IL, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
During the follow-up, one patient did not survive
due to non-cancer reasons and, therefore, was excluded from the
statistical analysis. Baseline clinical characteristics of the
remaining 41 patients are summarized in the first part of Table I. PEMT expression in the
cancer tissues was equal to its expression in the adjacent
non-cancer tissues and did not correlate with the disease stage
(results not shown). Furthermore, in patients with squamous cell
carcinoma, PEMT expression in the cancer tissue was equal to
that in patients with adenocarcinoma (results not shown). During
the four-year follow-up, 21 of the 41 patients succumbed to tumor
progression with a mean survival time of 24.6±16.4 months.
 | Table IClinical characteristics and
laboratory findings of patients with resectable non-small-cell lung
cancer. |
Table I
Clinical characteristics and
laboratory findings of patients with resectable non-small-cell lung
cancer.
| Parameters | Subjects |
|---|
| No. | 41 |
| Gender,
female/male | 11/30 |
| Age, years | 63.0 {53.0/67.0}
(44–77)a |
| Body mass index,
kg/m2 | 24.8 {22.2/28.3}
(16.4–46.5)a |
| Weight loss in the
last three months, no/yes | 24/17 |
| Smoking status,
never/current or former | 5/36 |
| Tumor stageb, I/II/III | 17/14/10 |
| Histological type,
squamous cell/adenocarcinoma/macrocellular/other | 19/13/5/4 |
| PEMT
expression ratio (expression in lung cancer tissue vs. non-cancer
lung tissue) | 0.93 {0.59/1.22}
(0.25–3.27)a |
| FASN activity
ratioc (activity in lung cancer
tissue vs. non-cancer lung tissue) | 1.48 {0.77/2.79}
(0.04–54.07)a |
| LPL activity
ratioc (activity in lung cancer
tissue vs. non-cancer lung tissue) | 1.91 {1.08/2.42}
(0.10–19.27)a |
For the survival analysis, PEMT expression
was specified as the ratio of gene expression in cancer tissues to
that in non-cancer tissues. Descriptive statistics of PEMT
expression ratios are presented in the second part of Table I. Deceased patients had a 32% higher
PEMT expression ratio in comparison with the patients that
were alive four years after tumor excision; however, the difference
was not significant (median, 1.135 vs. 0.0860; P=0.0556).
PEMT expression ratio was negatively correlated with
survival time following surgery, but this correlation was not
statistically significant (Spearman’s ρ=−0.2745; P=0.0786).
Cox-regression analysis revealed that increased PEMT
expression ratio predicted shorter patient survival, independently
of increased disease stage and weight loss, the two other
predictors of shorter patient survival in univariate statistical
analysis (Table II, Model 1).
| Table IIPredictors of shorter survival in
patients with resectable non-small-cell lung cancer. |
Table II
Predictors of shorter survival in
patients with resectable non-small-cell lung cancer.
| Model No. | Variables in the
model (Cox regression analysis) | Relative risk (95%
CI) | P-value |
|---|
| 1 | Disease stage (I
vs. II or III) | 5.87
(1.64–20.96) | 0.006 |
| Weight loss in the
last three months (no vs. yes) | 3.83
(1.45–10.15) | 0.007 |
| PEMT
expression ratio (low vs. high)a | 3.55
(1.35–9.33) | 0.010 |
| 2 | Disease stage (I
vs. II or III) | 4.42
(1.23–15.95) | 0.023 |
| Weight loss in the
last three months (no vs. yes) | 3.50
(1.27–9.70) | 0.016 |
| PEMT
expression ratio (low vs. high)a | 3.08
(1.16–8.15) | 0.023 |
| FASN activity ratio
(low vs. high)a,b | 2.36
(0.85–6.57) | 0.101 |
| 3 | Disease stage (I
vs. II or III) | 3.39
(0.94–12.24) | 0.063 |
| Weight loss in the
last three months (no vs. yes) | 2.51
(0.88–7.15) | 0.084 |
| PEMT
expression ratio (low vs. high)a | 3.00
(1.09–8.25) | 0.033 |
| LPL activity ratio
(low vs. high)a,c | 4.18
(1.26–13.87) | 0.019 |
In order to study PEMT involvement in tumor
growth and progression concurrently with other lipid-related
enzymes associated with tumor progression, such as FASN and LPL,
FASN activity ratios (ratio of activity in cancer tissue to that in
adjacent non-cancer tissue for each individual patient) and LPL
activity ratios (ratio of activity in cancer tissue to that in
adjacent non-cancer tissue for each individual patient) were used,
which were estimated previously in the same tissues and shown to
predict shorter patient survival (18–20).
Descriptive statistics of FASN and LPL activity ratios are
presented in the second part of Table
I. PEMT expression ratio did not correlate with FASN and
LPL activity ratios (results not shown) and increased PEMT
expression ratio predicted shorter patient survival, independently
of the increased FASN (Table II,
Model 2) and LPL activity ratios (Table II, Model 3).
Discussion
The present study shows that in patients with
resectable NSCLC, elevated PEMT expression in the cancer
tissue (relative to the adjacent non-cancer lung tissue) predicts
shorter survival, independently of standard prognostic factors and
also independently of increased FASN or LPL activities in the
cancer tissue.
Notably, increased PEMT expression in lung
cancer tissue is associated with adverse patient outcomes and is
explicable by several potential causes. Current data regarding PEMT
in cancer is scarce and controversial. On the one hand, malignant
colorectal tumors have an increased PC/PE ratio (9), which suggests that increased PEMT
activity is an essential mechanism for maintaining membrane
integrity and to prevent cell death. Furthermore,
PEMT−/− mice fed with a
choline-deficient diet did not develop hepatic cancer, but hepatic
steatosis (24). On the other hand,
in hepatocarcinomas, PEMT2 expression and hepatic PEMT
activity are decreased (11), and
hepatocytes with overexpressed PEMT2 are poorly tumorigenic
(25). Furthermore, PEMT
promoter polymorphism, −774G>C, which results in lower
PEMT expression and choline deficiency (26), was associated with increased breast
cancer risk among women receiving hormone replacement therapy
(27). PEMT predictive capacity
seems reasonable due to several causes. First, lipid mobilization
is required to sustain tumor growth (28). Highly proliferating cancer cells use
fatty acids (either synthesized endogenously by FASN or supplied
exogenously from the microvascular system) predominantly for the
synthesis of phospholipids, which are then incorporated in cell
membranes (7). PEMT may play an
important role in membrane assembly as PC, the end-product of the
PEMT reaction, is the most abundant phospholipid in cell membranes.
The CDP-choline pathway, which is the sole pathway for de
novo synthesis of PC in non-hepatic cells, may not be
sufficient for PC supply in cancer cells; therefore, cancer cells
activate the PEMT pathway to cover the increased demand for PC to
maintain an adequate PC/PE ratio, membrane integrity and cell
survival. Furthermore, PC synthesis via the PEMT pathway is dietary
choline independent. White adipose tissue covers its increased
demand for new phospholipids, which are constituents of lipid
droplets, by increasing PEMT expression (29). The second potential cause of PEMT
predictive capacity may be its role in the only known endogenous
pathway of choline synthesis, via the degradation of PC (Fig. 1) (30). The increased total level of
choline-containing compounds has been found in several types of
cancer (10) and the endogenous
synthesis of choline appears to be upregulated in various cancer
cells (31–35). Endogenous choline synthesis via the
PEMT pathway may be essential particularly in the case of
inadequate dietary choline uptake during increased cell
requirements. Furthermore, choline is utilized as a methyl group
donor and increased choline affects DNA methylation and results in
disruption of DNA repair (36). The
third potential cause of PEMT predictive capacity is that products
of PC hydrolysis (phosphocholine and diacylglycerol) may function
as secondary messengers (37) and
certain enzymes involved in PC degradation (such as phospholipase
D) may be implicated in cell proliferation, signal transduction,
malignant transformation and tumor progression (36). Thus, PEMT may play an important role
in tumor growth and progression in addition to lipid mobilization;
however, these speculations require further investigation.
The finding that elevated PEMT expression in
NSCLC tissue predicts shorter patient survival independently of
increased FASN or LPL activities further strengthens the evidence
that lipid mobilization is required to sustain tumor growth. Our
previous studies showed that increased LPL and FASN activities are
predictors of shorter survival in patients with NSCLC (18–20).
PEMT predictive capacity in addition to LPL and FASN is explicable
as FASN and LPL only supply the cancer tissue with long-chain fatty
acids, which are then further used as constituents in phospholipid
synthesis. PEMT is then involved in the synthesis of PC from PE, as
cancer cells are characterized by increased growth and synthesis of
new cell membranes has high requirements for PC. Moreover, other
metabolic pathways associated with PEMT (such as endogenous choline
synthesis, PC hydrolysis to secondary messengers and disruption of
DNA repair) are also independent of long-chain fatty acid
metabolism and, therefore independent of LPL and FASN activity
levels. As both enzymes are molecular targets for cancer therapy,
with specific inhibitors of FASN activity (7,38) or
by inducing LPL activity in non-cancer tissues by peroxisome
proliferator-activated receptor γ agonists (39,40),
PEMT may also be a promising diagnostic and therapeutic target.
Choline-containing compounds were found to be significantly
elevated in prostate cancer versus benign prostate tissues, hence
choline phospholipid metabolites can potentially be used for
diagnosing and managing prostate cancer patients (36). In addition, it was reported that
hexadecylphosphocholine, the alkylphospholipid analog, reduces cell
proliferation in hepatoma cells and simultaneously inhibits PC
synthesis via the CDP-choline and PEMT pathways (41).
In conclusion, increased PEMT expression in
NSCLC tissues (relative to the adjacent non-cancer lung tissue)
predicts shorter patient survival, independently of increased FASN
or LPL activities in the same cancer tissues.
References
|
1
|
Shields DJ, Agellon LB and Vance DE:
Structure, expression profile and alternative processing of the
human phosphatidylethanolamine N-methyltransferase (PEMT)
gene. Biochim Biophys Acta. 1532:105–114. 2001.PubMed/NCBI
|
|
2
|
Vance DE, Li Z and Jacobs RL: Hepatic
phosphatidylethanolamine N-methyltransferase, unexpected roles in
animal biochemistry and physiology. J Biol Chem. 282:33237–33241.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li Z, Agellon LB, Allen TM, et al: The
ratio of phosphatidylcholine to phosphatidylethanolamine influences
membrane integrity and steatohepatitis. Cell Metab. 3:321–331.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vance DE and Vance JE: Phospholipid
biosynthesis in eukaryotes. Biochemistry of lipids, lipoproteins
and membranes. 5th edition. Vance DE and Vance JE: Elsevier;
Amsterdam: pp. 213–244. 2008, View Article : Google Scholar
|
|
5
|
Bauer DE, Hatzivassiliou G, Zhao F,
Andreadis C and Thompson CB: ATP citrate lyase is an important
component of cell growth and transformation. Oncogene.
24:6314–6322. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Barger JF and Plas DR: Balancing
biosynthesis and bioenergetics: metabolic programs in oncogenesis.
Endocr Relat Cancer. 17:R287–R304. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kuhajda FP: Fatty acid synthase and
cancer: new application of an old pathway. Cancer Res.
66:5977–5980. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dueck DA, Chan M, Tran K, et al: The
modulation of choline phosphoglyceride metabolism in human colon
cancer. Mol Cell Biochem. 162:97–103. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dobrzyńska I, Szachowicz-Petelska B,
Sulkowski S and Figaszewski Z: Changes in electric charge and
phospholipids composition in human colorectal cancer cells. Mol
Cell Biochem. 276:113–119. 2005.PubMed/NCBI
|
|
10
|
Glunde K, Bhujwalla ZM and Ronen SM:
Choline metabolism in malignant transformation. Nat Rev Cancer.
11:835–848. 2011.PubMed/NCBI
|
|
11
|
Tessitore L, Dianzani I, Cui Z and Vance
DE: Diminished expression of phosphatidylethanolamine
N-methyltransferase 2 during hepatocarcinogenesis. Biochem J.
337:23–27. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Flavin R, Peluso S, Nguyen PL and Loda M:
Fatty acid synthase as a potential therapeutic target in cancer.
Future Oncol. 6:551–562. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ogino S, Nosho K, Meyerhardt JA, et al:
Cohort study of fatty acid synthase expression and patient survival
in colon cancer. J Clin Oncol. 26:5713–5720. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rossi S, Graner E, Febbo P, et al: Fatty
acid synthase expression defines distinct molecular signatures in
prostate cancer. Mol Cancer Res. 1:707–715. 2003.PubMed/NCBI
|
|
15
|
Shurbaji MS, Kalbfleisch JH and Thurmond
TS: Immunohistochemical detection of a fatty acid synthase (OA-519)
as a predictor of progression of prostate cancer. Hum Pathol.
27:917–921. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Takahiro T, Shinichi K and Toshimitsu S:
Expression of fatty acid synthase as a prognostic indicator in soft
tissue sarcomas. Clin Cancer Res. 9:2204–2212. 2003.PubMed/NCBI
|
|
17
|
Visca P, Sebastiani V, Botti C, et al:
Fatty acid synthase (FAS) is a marker of increased risk of
recurrence in lung carcinoma. Anticancer Res. 24:4169–4173.
2004.PubMed/NCBI
|
|
18
|
Cerne D, Zitnik IP and Sok M: Increased
fatty acid synthase activity in non-small cell lung cancer tissue
is a weaker predictor of shorter patient survival than increased
lipoprotein lipase activity. Arch Med Res. 41:405–409. 2010.
View Article : Google Scholar
|
|
19
|
Cerne D, Melkic E, Trost Z, Sok M and Marc
J: Lipoprotein lipase activity and gene expression in lung cancer
and in adjacent noncancer lung tissue. Exp Lung Res. 33:217–225.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Trost Z, Sok M, Marc J and Cerne D:
Increased lipoprotein lipase activity in non-small cell lung cancer
tissue predicts shorter patient survival. Arch Med Res. 40:364–368.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sobin LH and Wittekind C: Lung and pleural
tumours. TNM Classification of Malignant Tumours. 6th edition. John
Wiley & Sons; Hoboken, NJ, USA: pp. 97–107. 2002
|
|
22
|
Travis WD, Brambilla E, Muller-Hermelink
HK and Harris CC: World Health Organization Classification of
Tumours. Pathology and Genetics of Tumours of the Lung, Pleura,
Thymus and Heart. IARC Press; Lyon, France: 2004
|
|
23
|
Trost Z, Marc J, Sok M and Cerne D:
Increased apolipoprotein E gene expression and protein
concentration in lung cancer tissue do not contribute to the
clinical assessment of non-small cell lung cancer patients. Arch
Med Res. 39:663–667. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Vance JE and Vance DE: Metabolic insights
into phospholipid function using gene-targeted mice. J Biol Chem.
280:10877–10880. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tessitore L, Sesca E and Vance DE:
Inactivation of phosphatidylethanolamine N-methyltransferase-2 in
aflatoxin-induced liver cancer and partial reversion of the
neoplastic phenotype by PEMT transfection of hepatoma cells. Int J
Cancer. 86:362–367. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
da Costa KA, Kozyreva OG, Song J, et al:
Common genetic polymorphisms affect the human requirement for the
nutrient choline. FASEB J. 20:1336–1344. 2006.PubMed/NCBI
|
|
27
|
Xu X, Gammon MD, Ziesel SH, et al: Choline
metabolism and risk of breast cancer in a population-based study.
FASEB J. 22:2045–2052. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mulligan HD and Tisdale MJ: Effect of the
lipid-lowering agent bezafibrate on tumour growth rate in
vivo. Br J Cancer. 64:1035–1038. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hörl G, Wagner A, Cole LK, et al:
Sequential synthesis and methylation of phosphatidylethanolamine
promote lipid droplet biosynthesis and stability in tissue culture
in vivo. J Biol Chem. 286:17338–17350. 2011.PubMed/NCBI
|
|
30
|
Li Z and Vance DE: Phosphatidylcholine and
choline homeostasis. J Lip Res. 49:1187–1194. 2008. View Article : Google Scholar
|
|
31
|
Qi C, Park JH, Gibbs TC, et al:
Lysophosphatidic acid stimulates phospholipase D activity and cell
proliferation in PC-3 human prostate cancer cells. J Cell Physiol.
174:261–272. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Foster DA and Xu L: Phospholipase D in
cell proliferation and cancer. Mol Cancer Res. 1:789–800.
2003.PubMed/NCBI
|
|
33
|
Stewart JD, Marchan R, Lesjak MS, et al:
Choline-releasing glycerophosphodiesterase EDI3 drives tumor cell
migration and metastasis. Proc Natl Acad Sci USA. 109:8155–8160.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cao MD, Döpkens M, Krishnamachary B, et
al: Glycerophosphodiester phosphodiesterase domain containing 5
(GDPD5) expression correlates with malignant choline phospholipid
metabolite profiles in human breast cancer. NMR Biomed.
25:1033–1042. 2012. View
Article : Google Scholar
|
|
35
|
Kang DW, Choi KY and Min do S:
Phospholipase D meets Wnt signaling: a new target for cancer
therapy. Cancer Res. 71:293–297. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Awwad HM, Geisel J and Obeid R: The role
of choline in prostate cancer. Clin Biochem. 45:1548–1553. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ackerstaff E, Glunde K and Bhujwalla ZM:
Choline phospholipid metabolism: a target in cancer cells? J Cell
Biochem. 90:525–533. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mashima T, Seimiya H and Tsuruo T: De novo
fatty-acid synthesis and related pathways as molecular targets for
cancer therapy. Br J Cancer. 100:1369–1372. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Nomura K, Noguchi Y and Matsumoto A:
Stimulation of decreased lipoprotein lipase activity in the
tumour-bearing state by the antihyperlipidemic drug bezafibrate.
Surg Today. 26:89–94. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Mutoh M, Niho N and Wakabayashi K:
Concomitant suppression of hyperlipidemia and intestinal polyp
formation by increasing lipoprotein lipase activity in
Apc-deficient mice. Biol Chem. 387:381–385. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Jiménez-López JM, Carrasco MP, Segovia JL
and Marco C: Hexadecylphosphocholine inhibits phosphatidylcholine
synthesis via both the methylation of phosphatidylethanolamine and
CDP-choline pathways in HepG2 cells. Int J Biochem Cell Biol.
36:153–161. 2004.PubMed/NCBI
|