Introduction
Colorectal cancer is the third most common type of
cancer and the second most frequent cause of cancer mortality in
numerous industrialized countries (1). The majority of tumors arise
sporadically with no clear cause or genetic predisposition. Several
risk factors have been considered as causes of colorectal cancer,
but little has been confirmed. Viruses are among the few known
causes of cancer and contribute to various malignancies worldwide
(2). Previous studies on viral
etiology in colon cancer have reported contradictory findings
(3,4).
Parvovirus B19 (B19) is a non-enveloped virus with a
linear, single-stranded DNA genome. The B19 viral genome encodes
three proteins: The non-structural protein, NS1, and two viral
capsid proteins, VP1 and VP2 (5).
In our previous study, significantly higher levels of B19 nucleic
acids and proteins were found in neoplastic colon tissues (6). This finding indicates that an
association may exist between B19 infection and the development of
colon neoplasia.
Infection with parvovirus B19 is a global concern.
The infection rate is similar in the United States, Europe and
Asia, with ~50% of 15-year-old adolescents and >60% of adults
being seropositive (5). A previous
study has shown that B19 infection may contribute to the
pathogenesis of acute lymphoblastic and myeloblastic leukemia
(7). However, few studies have
investigated B19 in solid tumors or the mechanisms or regulatory
proteins that could be involved. Therefore, it is important to
establish whether B19 contributes to the pathogenesis of colorectal
cancer and its underlying mechanism.
The present study aimed to investigate the
pathogenic mechanisms underlying B19 in colon carcinoma by
analyzing differential gene expression and biological functions,
through assessing the changes in primary human colorectal
epithelial cells (HCECs) and LoVo cells following transfection with
plasmids containing VP1, VP2 and NS1.
Materials and methods
Plasmid construction
The recombinant eukaryotic cell inducible expression
vectors, pReceiver-M03-VP1, pReceiver-M33-VP2 and
pReceiver-M16-NS1, were constructed by inserting B19 full-length
VP1, VP2 and NS1 complementary DNA into pReceiver-M03,
pReceiver-M33 or pReceiver-M16 (GeneCopoeia, Inc., Rockville, MD,
USA), respectively. The VP1, VP2 and NS1 sequences were amplified
by polymerase chain reaction (PCR) analysis using the pGEM/1-B19
plasmid provided by Professor J.P. Clewley at the Central Public
Health Laboratory (London, UK) as the template, which contained the
B19 full-length open reading frame. The presence of the recombinant
plasmid was confirmed using DNA sequencing.
Cell culture and transfection
Samples of normal human colon tissue >10 cm
distant to the tumors was obtained from patients with colorectal
adenocarcinoma. HCECs were isolated from the normal human
colorectal tissue and were washed several times using
phosphate-buffered saline (PBS) containing penicillin, streptomycin
and amphotericin B using thermolysin and collagenase type I
(Sigma-Aldrich, St. Louis, MO, USA), as described previously
(8). The cells were then cultured
in Epithelial Cell Growth Medium-2 (ScienCell Research
Laboratories, Carlsbad, CA, UDA) containing amphotericin B. LoVo
carcinoma cells were obtained from the American Type Culture
Collection (Rockville, MD, USA) and were cultured in Dulbecco’s
modified Eagle’s medium containing 10% fetal bovine serum. All
cells were cultured at 37°C in an atmosphere containing 5%
CO2. All procedures were performed in accordance with
standard guidelines for the study of humans and were approved by
the Research Ethics Committee of Urumqi Military General Hospital
(Urumqi, China). All patients provided written informed
consent.
The cells were transfected with pReceiver-M03-VP1,
pReceiver-M33-VP2 and pReceiver-M16-NS1, using pReceiver-M03,
pReceiver-M33 and pReceiver-M16 as controls. Transfection was
performed using Lipofectamine® LTX and PLUS™ Reagents
(Invitrogen Life Technologies, Carlsbad, CA, USA) according to the
manufacturer’s instructions, with untransfected cells used as blank
controls.
Fluorescence microscopy
The expression of enhanced green fluorescent protein
(eGFP), enhanced cyan fluorescent protein (eCFP) and enhanced
yellow fluorescent protein (eYFP) in the transfected HCECs and LoVo
cells was observed using a fluorescence microscope (TE2000-U, Nikon
Corporation, Tokyo, Japan) equipped with a fluorescence filter.
Digital images of the cells were captured using a spot camera
system (Nikon Corporation).
Flow cytometric analysis
The cells (1×106) were fixed in 75%
alcohol for 12–16 h at 4°C, followed by ethidium bromide (50 μg/ml)
and RNase (100 μg/ml) treatment at 25°C for 30 min. Analysis was
performed using a flow cytometer (FACScan; Becton Dickinson,
Bedford, MA, USA).
Reverse transcription PCR (RT-PCR)
analysis
RNA was extracted 24 h and 48 h subsequent to
transfection, and RT-PCR was performed. The following primer pairs
were used: Vp1 forward, ttctgcatgactgctactgga and reverse, atc
ccctagaaaacccatcct; Vp2 forward, tatttgaggaggtggctgatg and reverse,
ccaataaaggaacccagcaat; Ns1 forward, ggtggtctggga tgaaggtat and
reverse, gtgttcccgcttacaacaaaa; and glyceraldehyde-3-phosphate
dehydrogenase (GAPDH). forward, tcggagtcaacggatttggtcgta and
reverse, tggcatggactgtggtcatgagtc.
Western blot analysis
Protein extraction was performed by washing the
cells twice with ice-cold PBS, followed by homogenization in lysis
buffer [50 mM HEPES (pH 7.5), 150 mM NaCl, 10% glycerol, 1% Triton
X-100, 1.5 mM MgCl2, 1 mM EDTA, 10 mM
Na4(PO4)2, 25 μg/ml
aprotinin and 25 μg/ml leupeptin] at 24 h, 48 h and 72 h
post-transfection. The insoluble fraction was removed using
centrifugation at 1,000 × g for 15 min at 4°C. Proteins were
analyzed using electrophoresis (50 μg per lane) on 10%
polyacrylamide gels and transferred to polyvinylidene fluoride
(PVDF) membranes. Mouse monoclonal antibodies against the B19
proteins VP1 and VP2 (clone, R92F6; Chemicon, Billerica, MA, USA)
and anti-NS1 antibodies [a gift from Professor Susanne Modrow
(9) and Dr Simon Bredl, Institute
of Medical Microbiology, University of Regensburg, Germany] were
used to identify the proteins on the PVDF membranes. Horseradish
peroxidase-conjugated goat antimouse secondary antibodies
(Sigma-Aldrich) were detected using enhanced chemiluminescence
western blot analysis reagents (Pierce Biotechnology, Inc.,
Rockford, IL, USA).
Microarray hybridization and data
analysis
Microarray hybridization was performed by Shanghai
Biochip Co., Ltd., (Shanghai, China) using an Agilent SurePrint G3
Human GE 8×60k microarray (Agilent Technologies, Santa Clara, CA,
USA) that targeted 27,958 Entrez Gene RNAs and 7,419 long
non-coding RNAs (reference). In brief, total RNAs from the
transfected cells were extracted and purified using the Qiagen
RNeasy® Mini kit (Qiagen, Hilden, Germany). Total RNA
was amplified using the Low Input Quick Amp Labeling kit, One-Color
(Agilent Technologies). For hybridization, each slide was
hybridized with 1.65 μg Cy-3 labeled complementary RNA using the
Gene Expression Hybridization kit (Agilent Technologies) in a
Hybridization Oven (Agilent Technologies) according to the
manufacturer’s instructions. Subsequent to 17 h of hybridization,
the slides were washed in staining dishes (Thermo Fisher
Scientific, Waltham, MA, USA) with Gene Expression Wash Buffer
(Agilent Technologies) according to the manufacturer’s
instructions. The slides were scanned at a 3-μm resolution using
the green dye channel in an Agilent Microarray Scanner (Agilent
Technologies). The data were read using Feature Extraction Software
10.7 (Agilent Technologies), and were normalized using Quantile
Algorithm, Gene Spring 11.0 software (Agilent Technologies).
The data from three replicates were averaged. Genes
were defined as differentially expressed if the intensity ratio
(Cy5) was found to increase or decrease >2-fold and if the
intensity ratio (Cy5) showed the same direction of change
(upregulated or downregulated) in all three experimental repeats.
Gene ontology (GO) and pathway analyses were performed using
Database for Annotation, Visualization and Integrated Discovery
v6.7 software (10,11).
Quantitative (q)PCR analysis
qPCR analysis using SYBR® Green
(Invitrogen Life Technologies) was performed in order to verify the
results of the microarray analysis. Total RNA was extracted from
the transfected cells. The RNA was reverse transcribed using Murine
Leukemia Virus reverse transcriptase (Promega Corp., Madison, WI,
USA). The expression of the 12 genes that were identified as being
associated with apoptosis in the microarray analysis was determined
using qPCR analysis with SYBR-Green I (Invitrogen Life
Technologies). GAPDH was used as an internal control and distilled
water was used as a negative control. The amplification reaction
consisted of 10X PCR buffer, 1.25 units of JumpStart™ Taq
(Sigma-Aldrich), 10 pmol forward and reverse primers, 0.2 μmol
dNTP, 100 ng template and 0.2X SYBR-Green I (Amresco Inc., Solon,
OH, USA) in a final volume of 50 μl. The reactions were performed
using the StepOneTM Real-Time PCR System (Applied
Biosystems, Inc., Foster City, CA, USA). The mRNA expression of the
12 genes was normalized with GAPDH using the 2−ΔΔCt
method (12). The primer sequences
used for GAPDH and the 12 genes were retrieved from PrimerBank
(http://pga.mgh.harvard.edu/primerbank/).
Results
VP1, VP2 and NS1 expression in HCECs and
LoVo cells
Primary normal HCECs were isolated, cultured and
transiently transfected with the pReceiver-M03-VP1,
pReceiver-M33-VP2 and pReceiver-M16-NS1 constructs. The expression
of eGFP-VP1, eCFP-VP2 and eYFP-NS1 in the HCECs and LoVo cells was
analyzed and confirmed using fluorescence microscopy, RT-PCR
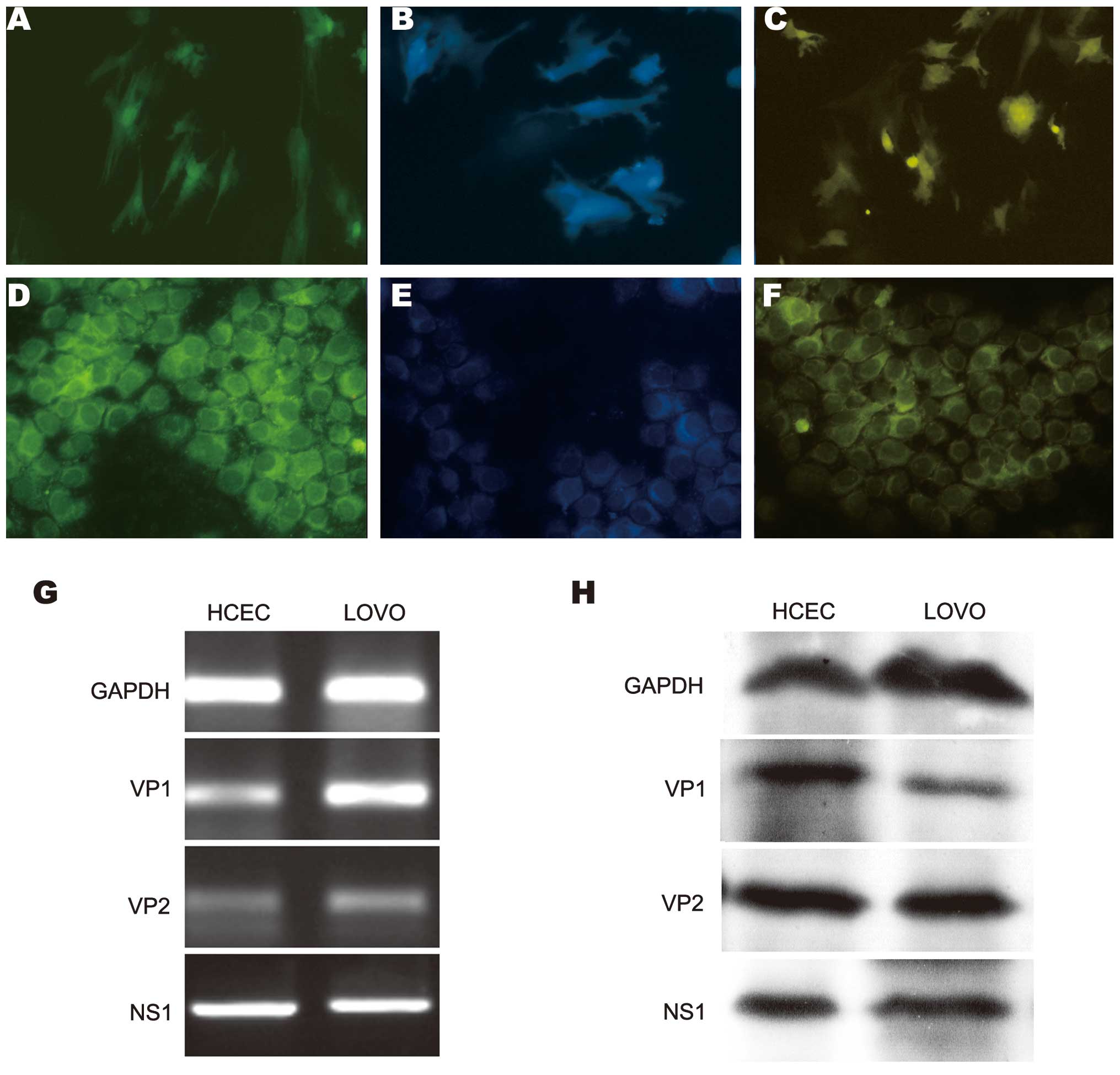
analysis and western blot analysis. Marked green, cyan and yellow
fluorescence, indicating the expression of VP1, VP2 and NS1,
respectively, was observed at 24 h post-transfection, with the
strongest expression observed after 48 h (Fig. 1A–F). The mRNA expression of VP1, VP2
and NS1 was detected using RT-PCR analysis in the transfected cells
(Fig. 1G). To further assess the
expression of eGFP-VP1, eCFP-VP2 and eYFP-NS1 in the HCECs and LoVo
cells, the protein expression of VP1, VP2 and NS1 was confirmed
using anti-VP1, -VP2 and -NS1 antibodies in western blot analysis
(Fig. 1H).
Flow cytometric analysis
No significant changes in the cell cycle or
apoptosis were identified in the HCECs transfected with
pReceiver-M03-VP1, pReceiver-M33-VP2 or pReceiver-M16-NS1 and
pReceiver-M03, pReceiver-M33 or pReceiver-M16. Similarly, no
significant changes in cell cycle or apoptosis were identified in
the LoVo cells transfected with pReceiver-M03-VP1,
pReceiver-M33-VP2 or pReceiver-M16-NS1 and pReceiver-M03,
pReceiver-M33 or pReceiver-M16.
Differential gene analysis
Using the human genome expression microarray,
differential gene expression was detected in the HCECs and LoVo
cells transfected with pReceiver-M03-VP1, pReceiver-M33-VP2 or
pReceiver-M16-NS1 compared with those transfected with
pReceiver-M03, pReceiver-M33 or pReceiver-M16, respectively. The
number of upregulated and downregulated genes (P<0.05; false
discovery rate <0.05; fold-change >2.0) are shown in Table I. The top five differential genes in
the six groups are shown in Table
II. The fold change of the differentially-expressed genes
associated with colorectal cancer are shown in Table III.
 | Table INumber of differentially-expressed
genes in HCECs and LoVo cells transfected with plasmids. |
Table I
Number of differentially-expressed
genes in HCECs and LoVo cells transfected with plasmids.
| VP1 | VP2 | NS1 |
|---|
|
|
|
|
|---|
| Cell | Upregulated, n | Downregulated, n | Upregulated, n | Downregulated, n | Upregulated, n | Downregulated, n |
|---|
| HCEC | 740 | 361 | 131 | 124 | 447 | 690 |
| LoVo | 613 | 328 | 546 | 562 | 508 | 652 |
| Table IITop five differentially-expressed
genes in HCECs and LoVo cells. |
Table II
Top five differentially-expressed
genes in HCECs and LoVo cells.
| Upregulated | Downregulated |
|---|
|
|
|
|---|
| Group | Gene symbol | Gene name | Gene ID | Log2
ratio | Gene symbol | Gene name | Gene ID | Log2
ratio |
|---|
| HCEC |
| VP1 |
| NME4 | Non-metastatic cells
4, protein expressed in | NM_005009 | 10.9 | RWDD4A | RWD domain containing
4A | NM_152682 | −7.7 |
| TMCO1 | Transmembrane and
coiled-coil domains 1 | NM_019026 | 5.9 | POU3F3 | POU class 3 homeobox
3 | NM_006236 | −4.0 |
| IL6 | Interleukin 6 | NM_000600 | 5.1 | ABCC6P1 | ATP-binding cassette,
sub-family C, member 6 pseudogene 1 | NR_003569 | −3.8 |
| CCL20 | Chemokine (C-C motif)
ligand 20 | NM_004591 | 5.1 | LOC100131599 | Hypothetical protein
LOC100131599 | AK126221 | −3.6 |
| CCL5 | Chemokine (C-C motif)
ligand 5 | NM_002985 | 5.1 | MYBPC2 | Myosin-binding
protein C, fast type | NM_004533 | −3.2 |
| VP2 |
| TSPAN32 | Tetraspanin 32 | NM_139022 | 4.9 | NME4 | Non-metastatic cells
4, protein expressed in | NM_005009 | −11.2 |
| LOC100294501 | Hypothetical protein
LOC100294501 | XM_002344054 | 4.1 | TMCO1 | Transmembrane and
coiled-coil domains 1 | NM_019026 | −6.1 |
| BTBD19 | BTB (POZ) domain
containing 19 | NM_001136537 | 3.7 | BTN1A1 | Butyrophilin,
subfamily 1, member A1 | NM_001732 | −5.0 |
| TTC29 | Tetratricopeptide
repeat domain 29 | NM_031956 | 3.7 | LOC221122 | Hypothetical
LOC221122 | NR_026681 | −4.5 |
| CCDC114 | Coiled-coil domain
containing 114 | NM_144577 | 3.6 | HIBCH |
3-hydroxyisobutyryl-Coenzyme A
hydrolase | NM_014362 | −4.2 |
| NS1 |
| ATG9A | ATG9 autophagy
related 9 homolog A (S. cerevisiae) | NM_001077198 | 11.4 | SAE1 | SUMO1-activating
enzyme subunit 1 | NM_005500 | −6.1 |
| MTUS1 | Microtubule
associated tumor suppressor 1 | NM_001001925 | 5.9 | RFESD | Rieske (Fe-S)
domain containing | NM_173362 | −5.9 |
| UBE2CBP |
Ubiquitin-conjugating enzyme E2C binding
protein | NM_198920 | 5.9 | GLYATL2 |
Glycine-N-acyltransferase-like 2 | NM_145016 | −5.7 |
| DNAJC8 | DnaJ (Hsp40)
homolog, subfamily C, member 8 | NM_014280 | 5.4 | C6ORF225 | Chromosome 6 open
reading frame 225 | NM_001033564 | −5.6 |
| C7ORF62 | Chromosome 7 open
reading frame 62 | NM_152706 | 5.2 | MRPS18A | Mitochondrial
ribosomal protein S18A | NM_018135 | −5.4 |
| LoVo |
| VP1 |
| IFI6 | Interferon,
α-inducible protein 6 | NM_022873 | 4.8 | WDD4A | RWD domain
containing 4A | NM_152682 | −7.3 |
| CCL5 | Chemokine (C-C
motif) ligand 5 | NM_002985 | 4.7 | HIBCH |
3-hydroxyisobutyryl-Coenzyme A
hydrolase | NM_014362 | −4.4 |
| IL6 | Interleukin 6
(interferon, β2) | NM_000600 | 4.7 | POU3F3 | POU class 3
homeobox 3 | NM_006236 | −4.1 |
| CCL20 | Chemokine (C-C
motif) ligand 20 | NM_004591 | 4.7 | ABCC6P1 | ATP-binding
cassette, sub-family C, member 6 pseudogene 1 | NR_003569 | −3.8 |
| TSPAN32 | Tetraspanin 32 | NM_139022 | 4.6 | LOC100131599 | Hypothetical
protein LOC100131599 | AK126221 | −3.6 |
| VP2 |
| LOC283482 | Hypothetical
LOC283482 | AK092513 | 6.3 | KRTAP21-1 | Keratin-associated
protein 21-1 | NM_181619 | −6.8 |
| RRP15 | Ribosomal RNA
processing 15 homolog (S. cerevisiae) | NM_016052 | 6.2 | MDM1 | Mdm1 nuclear
protein homolog (mouse) | NM_020128 | −6.1 |
| TRIM71 | Tripartite
motif-containing 71 | NM_001039111 | 5.9 | C2ORF76 | Chromosome 2 open
reading frame 76 | NM_001017927 | −5.8 |
| ADPRHL1 |
ADP-ribosylhydrolase like 1 | NM_138430 | 5.7 | OR4K17 | Olfactory receptor,
family 4, subfamily K, member 17 | NM_001004715 | −5.8 |
| GOLGA6L6 | Golgi autoantigen,
golgin subfamily a, 6-like 6 | NM_001145004 | 5.3 | PAQR5 | Progestin and
adipoQ receptor family member V | NM_001104554 | −5.6 |
| NS1 |
| TMEM50A | Transmembrane
protein 50A | NM_014313 | 11.7 | SAE1 | SUMO1 activating
enzyme subunit 1 | NM_005500 | −6.1 |
| TBCA | Tubulin folding
cofactor A | NM_004607 | 9.0 | RFESD | Rieske (Fe-S)
domain containing | NM_173362 | −5.9 |
| LOC723809 | Hypothetical
LOC723809 | NR_027374 | 8.8 | LOC100129954 | Hypothetical
LOC100129954 | XM_001714109 | −5.9 |
| PDZD8 | PDZ domain
containing 8 | NM_173791 | 6.8 | C9ORF38 | Chromosome 9 open
reading frame 38 | AF090921 | −5.4 |
| FEM1C | Fem-1 homolog c
(C. elegans) | NM_020177 | 6.3 | GORAB | Golgin,
RAB6-interacting | NM_152281 | −5.4 |
| Table IIIFold-change of
differentially-expressed genes associated with colorectal
cancer. |
Table III
Fold-change of
differentially-expressed genes associated with colorectal
cancer.
| | HCECs | LoVo cells |
|---|
| |
|
|
|---|
| Gene symbol | Genbank
accession | VP1 | VP2 | NS1 | VP1 | VP2 | NS1 |
|---|
| RAC3 | NM_005052 | - | - | 1.44 | - | - | 1.59 |
| MYCL1 | NM_005376 | - | - | 1.29 | - | - | 1.06 |
| APCDD1 | NM_153000 | - | 2.94 | - | 2.91 | - | - |
| APC | NM_001127511 | - | - | 1.41 | - | - | −2.38 |
| TCF7L2 | NM_030756 | - | - | - | - | 1.38 | 3.43 |
| VTI1A | NM_145206 | - | - | −1.17 | - | - | −1.23 |
| TP53INP2 | NM_021202 | 1.47 | - | - | - | - | - |
| TP53I11 | NM_001076787 | - | - | −3.42 | - | - | −1.34 |
| TP53INP1 | NM_033285 | - | - | - | 1.08 | - | - |
| CRCX7 | NM_020311 | 1.77 | - | - | 1.63 | - | - |
| BAX | NM_004324 | - | - | 1.23 | - | - | 1.05 |
| TMBIM1 | NM_022152 | - | - | - | 1.61 | - | - |
| LRP11 | NM_032832 | - | - | - | - | 3.04 | - |
| VEGFA | NM_001025370 | - | - | 1.19 | - | - | - |
| CCND1 | NM_053056 | - | - | 2.59 | - | - | - |
| FOS | NM_005252 | - | - | −1.82 | - | - | −1.83 |
| FOSB | NM_006732 | 1.66 | - | - | 1.27 | - | - |
| FOSL1 | NM_005438 | 2.21 | - | - | 1.98 | - | 2.27 |
| ID1 | NM_002165 | 1.24 | - | - | - | - | - |
| FZD4 | NM_012193 | 3.40 | - | - | 2.85 | - | - |
| FZD1 | NM_003505 | - | - | −1.14 | - | - | −1.11 |
| FZD10 | NM_007197 | 1.15 | - | - | - | - | - |
GO analysis
The differentially-expressed genes were classified
into different functional categories based on GO analysis for
biological process, molecular function and cellular components. The
primary GO categories for the upregulated genes in the HCECs and
LoVo cells transfected with pReceiver-M03-VP1 included immune
response, immune system process, defense response and response to
stimulus, and for the downregulated genes was primarily cellular
amino acid metabolic process. The predominant GO categories for the
upregulated genes in the HCECs and LoVo cells transfected with
pReceiver-M16-NS1 included organelle fission, nuclear division,
mitosis, the M-phase of the mitotic cell cycle, the mitotic cell
cycle, M-phase, cell cycle phase, cell cycle process and cell
division (Table IV).
| Table IVGO terms for the
differentially-expressed genes. |
Table IV
GO terms for the
differentially-expressed genes.
| Group | GO term | Genes, n | Genes, % | Fold
enrichment | P-value | FDR |
|---|
| Upregulation |
| VP1 in HCEC | GO:0006955~immune
response | 87 | 11.85 | 3.48 | 9.34E-25 | 0.000 |
| GO:0002376~immune
system process | 105 | 14.31 | 2.90 | 1.06E-23 | 0.000 |
| GO:0006952~defense
response | 64 | 8.72 | 2.87 | 5.31E-14 | 0.000 |
| GO:0050896~response
to stimulus | 197 | 26.84 | 1.55 | 3.60E-12 | 0.000 |
| VP2 in HCEC | No significant
enrichment | - | - | - | - | - |
| NS1 in HCEC |
GO:0048285~organelle fission | 20 | 4.49 | 4.33 | 1.90E-07 | 0.000 |
| GO:0000280~nuclear
division | 19 | 4.27 | 4.28 | 4.96E-07 | 0.000 |
|
GO:0007067~mitosis | 19 | 4.27 | 4.28 | 4.96E-07 | 0.000 |
| GO:0000087~M-phase
of mitotic cell cycle | 19 | 4.27 | 4.20 | 6.43E-07 | 0.001 |
| GO:0000278~mitotic
cell cycle | 24 | 5.39 | 3.21 | 1.72E-06 | 0.002 |
|
GO:0000279~M-phase | 22 | 4.94 | 3.31 | 3.23E-06 | 0.005 |
| GO:0022403~cell
cycle phase | 24 | 5.39 | 2.87 | 1.11E-05 | 0.019 |
| GO:0022402~cell
cycle process | 28 | 6.29 | 2.45 | 2.87E-05 | 0.049 |
| GO:0051301~cell
division | 18 | 4.04 | 3.02 | 1.02E-04 | 0.175 |
| VP1 in LoVo | GO:0006955~immune
response | 59 | 9.67 | 2.82 | 1.16E-12 | 0.000 |
| GO:0002376~immune
system process | 72 | 11.80 | 2.38 | 8.37E-12 | 0.000 |
| GO:0050896~response
to stimulus | 149 | 24.43 | 1.40 | 2.42E-06 | 0.004 |
| VP2 in LoVo | No significant
enrichment | - | - | - | - | - |
| NS1 in LoVo |
GO:0048285~organelle fission | 22 | 4.35 | 4.36 | 3.38E-08 | 0.000 |
|
GO:0007067~mitosis | 21 | 4.15 | 4.33 | 8.32E-08 | 0.000 |
| GO:0000280~nuclear
division | 21 | 4.15 | 4.33 | 8.32E-08 | 0.000 |
| GO:0000087~M-phase
of mitotic cell cycle | 21 | 4.15 | 4.26 | 1.12E-07 | 0.000 |
|
GO:0000279~M-phase | 25 | 4.94 | 3.45 | 2.75E-07 | 0.000 |
| GO:0000278~mitotic
cell cycle | 25 | 4.94 | 3.07 | 2.26E-06 | 0.004 |
| GO:0022403~cell
cycle phase | 26 | 5.14 | 2.85 | 4.98E-06 | 0.009 |
| GO:0022402~cell
cycle process | 30 | 5.93 | 2.41 | 1.99E-05 | 0.034 |
| GO:0051301~cell
division | 18 | 3.56 | 2.77 | 2.89E-04 | 0.492 |
| Downregulation |
| VP1 in HCEC | GO:0006520~cellular
amino acid metabolic process | 16 | 4.43 | 4.78 | 1.28E-06 | 0.002 |
| GO:0048037~cofactor
binding | 17 | 4.71 | 4.42 | 1.57E-06 | 0.002 |
| GO:0044106~cellular
amine metabolic process | 18 | 4.99 | 3.99 | 2.78E-06 | 0.004 |
| GO:0009308~amine
metabolic process | 20 | 5.54 | 3.38 | 7.38E-06 | 0.012 |
|
GO:0019752~carboxylic acid metabolic
process | 24 | 6.65 | 2.92 | 7.81E-06 | 0.013 |
| GO:0043436~oxoacid
metabolic process | 24 | 6.65 | 2.92 | 7.81E-06 | 0.013 |
| VP2 in HCEC | No significant
enrichment | - | - | - | - | - |
| NS1 in HCEC | GO:0043167~ion
binding | 175 | 25.36 | 1.32 | 1.35E-05 | 0.020 |
| GO:0046872~metal
ion binding | 171 | 24.78 | 1.32 | 1.75E-05 | 0.026 |
| GO:0043169~cation
binding | 171 | 24.78 | 1.31 | 3.08E-05 | 0.046 |
| VP1 in LoVo | GO:0006520~cellular
amino acid metabolic process | 14 | 4.27 | 4.58 | 1.17E-05 | 0.019 |
| GO:0009069~serine
family amino acid metabolic process | 6 | 1.83 | 17.06 | 2.19E-05 | 0.036 |
|
GO:0046394~carboxylic acid biosynthetic
process | 11 | 3.35 | 5.24 | 4.67E-05 | 0.078 |
| GO:0016053~organic
acid biosynthetic process | 11 | 3.35 | 5.24 | 4.67E-05 | 0.077 |
| GO:0008652~cellular
amino acid biosynthetic process | 7 | 2.13 | 10.14 | 5.98E-05 | 0.099 |
| VP2 in LoVo | No significant
enrichment | - | - | - | - | - |
| NS1 in LoVo | No significant
enrichment | - | - | - | - | - |
Pathway analysis
Significant pathways for the upregulated and
downregulated differentially-expressed genes are shown in Table V. No pathways or specified pathways
were found among the upregulated genes in the HCECs following
transfection with pReceiver-M33-VP2 or in the LoVo cells following
transfection with pReceiver-M33-VP2 or pReceiver-M16-NS1 (Table V). Similarly, no pathways were found
among the downregulated genes in the HCECs following transfection
with pReceiver-M03-VP1, pReceiver-M33-VP2 or pReceiver-M16-NS1, or
in the LoVo cells following transfection with pReceiver-M03-VP1 or
pReceiver-M33-VP2 compared with the cells transfected with the
control plasmids.
| Table VSignificant pathways for
differentially-expressed genes. |
Table V
Significant pathways for
differentially-expressed genes.
| Group | Pathway name | Genes, n | Genes | Fold change | P-value | FDR |
|---|
| Upregulation |
| VP1 in HCEC | Cell adhesion
molecules | 14 | CD274, CD86, F11R,
CDH1, ITGB8, ICAM1, HLA-A, HLA-B, HLA-C, HLA-E, HLA-F, HLA-G,
HLA-DRB5, PTPRC, SDC4 | 2.4 | 0.005 | 5.9 |
| Antigen processing
and presentation | 12 | B2M, CTSS, HSP70B,
HSP70B′, HLA-A, HLA-B, HLA-C, HLA-E, HLA-F, HLA-G, HLA-DRB5,
TAP1 | 2.7 | 0.010 | 1.1 |
| Cytokines and
inflammatory response | 7 | CSF-2, CSF-3, IL1A,
IL11, IL-6, IL-8, TNF | 4.6 | 0.002 | 3.2 |
| VP2 in HCEC | None | | | | | |
| NS1 in HCEC | Cell cycle | 21 | NDC80, SPC25, BUB1,
CDC20, CENPM, CCNA2, CCND1, KIF20A, KIF23, MCM5, PTTG1, PTTG2,
PSMB8, TUBA1A, TUBA4A, TUBB2C, TUBB, TUBB5, TUBBP2, TUBBP1,
UBE2E1 | 7.0 | 0.001 | 1.3 |
| Pathways in
cancer | 9 | BAX, APC, CCND1,
FGF10, FGF17, LAMA4, RAC3, VEGFA, WNT10B | 1.2 | 0.52 | 100 |
| Colorectal
cancer | 4 | BAX, APC, RAC3,
CCND1 | 2.0 | 0.32 | 99 |
| Focal adhesion | 6 | MYLPF, CAV1, CCND1,
LAMA4, RAC3, VEGFA | 1.5 | 0.34 | 99 |
| VP1 in LoVo | Cell adhesion
molecules | 15 | CD274, CD86, F11R,
ITGB8, ICAM1, HLA-A, HLA-B, HLA-C, HLA-E, HLA-F, HLA-G, HLA-DRB5,
SDC4, PVRL2, PVRL3 | 3.2 | 0.000 | 0.2 |
| Antigen processing
and presentation | 10 | B2M, CTSS, HLA-A,
HLA-B, HLA-C, HLA-E, HLA-F, HLA-G, HLA-DRB5, TAP1 | 3 | 0.009 | 10 |
| Cytokines and
inflammatory response | 4 | CSF-3, IL1A, IL-8,
TNF | 2.9 | 0.015 | 84 |
| VP2 in LoVo | None | | | | | |
| NS1 in LoVo | Not
significant | | | | | |
| Downregulation |
| VP1 in HCEC | Not
significant | | | | | |
| VP2 in HCEC | None | | | | | |
| NS1 in HCEC | Not
significant | | | | | |
| VP1 in LoVo | Not
significant | | | | | |
| VP2 in LoVo | Not
significant | | | | | |
| NS1 in LoVo | Cytoskeletal
regulation by Rho GTPase | 16 | ASPM, ENAH, IGFN1,
MYLK, MYH13, MYH6, PAK3, PAK2, RAC3, TTN, TUBB2A, TUBB3, TUBB,
TUBB5, TUBBP2, TUBBP1, | 2.2 | 0.013 | 13 |
| Wnt signaling
pathway | 22 | ARID1A, EP300,
INO80, SMARCB1, APC, ARRB2, DCHS1, FZD1, GNG3, MYH13, MYH6, NFATC3,
PPP3CB, PCDH18, PCDH7, PCDHA5, PCDHGA5, PCDHGB7, SVEP1, TTBK1,
TCF7L2, MTCL1 | 1.2 | 0.25 | 96 |
| Pathways in
cancer | 12 | BAX, EP300, BMP4,
FZD1, IGF1, PPARG, PIAS4, RAC3, RUNX1T1, TCEB2, TCF7L2, FOS | 0.78 | 0.91 | 100 |
| Colorectal
cancer | 6 | BAX, APC, RAC3,
FZD1, TCF7L2, FOS | 1.4 | 0.42 | 100 |
Confirmation of microarray results using
qPCR analysis
To verify the microarray analysis data, the
expression of the 12 differentially-expressed genes selected using
microarray analysis was confirmed by qPCR analysis in the different
groups. Consistent results were observed with regard to the nine
genes in the microarray and qPCR analysis data (Table VI).
| Table VIExpression of 12
differentially-expressed genes detected using microarray analysis
compared with qPCR analysis. |
Table VI
Expression of 12
differentially-expressed genes detected using microarray analysis
compared with qPCR analysis.
| Gene symbol | Groups | qPCR | Microarray |
|---|
| FOSB | VP1/HCEC | 1.6 | 1.7 |
| MYBPC2 | VP1/HCEC | −3.5 | −3.2 |
| APCDD1 | VP2/HCEC | 3.1 | 2.9 |
| NME4 | VP2/HCEC | −10.6 | −11.2 |
| RAC3 | NS1/HCEC | 1.7 | 1.4 |
| TP53I11 | NS1/HCEC | −4.0 | −3.4 |
| CRCX7 | VP1/LoVo | 1.5 | 1.6 |
| ABCC6P1 | VP1/LoVo | −3.7 | −3.8 |
| LRP11 | VP2/LoVo | 2.9 | 3.0 |
| KRTAP21-1 | VP2/LoVo | −6.9 | −6.8 |
| TCF7L2 | NS1/LoVo | 3.2 | 3.4 |
| FOS | NS1/LoVo | −1.6 | −1.8 |
Discussion
Despite our current understanding of the genetic
alterations associated with the progression of colon cancer, the
specific etiology of colorectal cancer has yet to be elucidated.
Epidemiological studies have indicated that environmental factors
and host immunological characteristics may contribute to the
initiation and progression of colon cancer. Infectious agents,
primarily viral infection, are acquired through the environment and
have the potential to alter numerous regulatory processes, which
may result in the development of colorectal cancer. Our previous
study showed that B19 infection may cause colon carcinoma (6). However, little is known regarding the
pathogenic mechanisms responsible for B19-induced
tumorigenesis.
B19 was discovered in 1974 and is the only
Parvoviridae family member that is known to be pathogenic in
humans. The genome of B19 has two large open reading frames
encoding a single non-structural protein, NS1, and two capsid
proteins, VP1 and VP2, which form an icosahedral capsid (5). The contribution of these viral
proteins to B19 infectivity have yet to be experimentally
demonstrated due to problems with in vitro culture and the
lack of an infectious clone. Due to the difficulty in culturing B19
in vitro, little experimental evidence exists regarding the
known and putative roles of B19 viral proteins in infectivity. In
the present study, plasmids containing VP1, VP2 and NS1 were
constructed and transfected into cultured HCECs and LoVo cells.
Through the analysis of differentially-expressed genes and their
functional enrichment, the present study aimed to identify
potential targets to enable further investigation of the function
of B19 in colon carcinoma, rather than to identify specific
signaling pathways or molecules leading to colon carcinoma in which
B19 participated.
Current understanding of the B19 viral proteins is
primarily based on studies of other parvoviruses. The B19 NS
protein is a multifunctional protein, for which sequence analysis
has revealed that, in addition to transregulation of the p6
promoter (13,14), NS contains motifs for nucleoside
triphosphate (NTP) binding and hydrolysis (15) associated with helicase activity,
thus indicating a role for the protein in B19 DNA replication. A
previous study has also indicated that the NTP-binding motifs of NS
play roles in the induction erythroid lineage cell apoptosis during
B19 infection (16). VP2 is the
major capsid protein, comprising 95% of the capsid and 58-kDa in
size (17). Previous studies in
insect cells have reported that VP2 can self-assemble into
virus-like particles (17) and that
it is capable of binding directly to blood group P antigen, which
is the cellular receptor of B19 (18). VP1 is the minor capsid protein,
which has an identical amino acid sequence to VP2, plus an extra
227 amino acids termed the VP1-unique region (VP1u)) at the amino
terminus (19). Previous studies
have shown that the VP1u, which is found on the exterior of the
capsid, contains the primary neutralizing epitopes of B19 (20–22).
Furthermore, a conserved phospholipase A2 motif has been identified
in the VP1u of members of the Parvoviridae family, including B19
(23,24). Two small 7.5- and 11-kDa proteins,
are encoded by the small abundant mRNA of B19 (25–27)
and are unique among those parvoviruses that have so far been
characterized. A number of proline-rich motifs are contained within
the 11-kDa protein and are conserved to the Src homology 3 binding
domain of eukaryotic proteins (28); however, the function of the 7.5- and
11-kDa proteins in B19 replication and/or pathogenesis has yet to
be elucidated.
In the present study, plasmids containing VP1, VP2
and NS1 were constructed for transfection into cultured HCECs and
LoVo cells. Hundreds of differentially-expressed genes were
identified in the HCECs and LoVo cells following VP1, VP2 and NS1
protein expression in different ontological pathways and functional
GO groups. GO analyses revealed that the significant VP1-related
ontology categories included that of immune response, immune system
process, defense response and response to stimulus, while
significant NS1-related ontology categories included organelle
fission, nuclear division, mitosis, M-phase of the mitotic cell
cycle, mitotic cell cycle, M-phase, cell cycle phase, cell cycle
process and cell division. Pathway expression analysis identified
that VP1-related pathways included cell adhesion molecules, antigen
processing and presentation, cytokines and inflammatory response.
Pathway expression analysis identified that NS1-related pathways
included cell cycle, pathways in cancer, colorectal cancer, the wnt
signaling pathway and focal adhesion. The functional GO categories
and pathways associated with VP1 and NS1 that were identified in
the present study were consistent with the functions of VP1 and NS1
reported previously (6,9,13,14,16–23,28–30).
This indicates that NS1 has a significant role in the pathogenesis
of B19 in colorectal carcinoma.
In conclusion, the present study identified twelve
differentially-expressed genes (BAX, EP300, BMP4, FZD1, IGF1,
PPARG, PIAS4, RAC3, RUNX1T1, TCEB2, TCF7L2 and FOS) that were found
to participate in general cancer pathways, and six genes (BAX, APC,
RAC3, FZD1, TCF7L2 and FOS) that were found to specifically
participate in colorectal cancer pathways. Furthermore, genes
associated with cancer, including MYCL1, APCDD1, VTI1A, TP53INP2,
TP53I11, TP53INP1, CRCX7, TMBIM1, LRP11, CCND1, FOSB, FOSL1, FZD4
and FZD10, were found to be differentially expressed. These may be
the primary genes involved in regulating the pathogenesis of B19 in
colorectal carcinoma. Moreover, NS1 may be the key molecule
involved in the pathogenesis of B19 in colorectal carcinoma.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China (no. 30873472) and the
China Postdoctoral Science Foundation (no. 20090461485).
References
|
1
|
Jemal A, Bray F, Center MM, et al: Global
cancer statistics. CA Cancer J Clin. 61:69–90. 2011.
|
|
2
|
Pagano JS, Blaser M, Buendia MA, et al:
Infectious agents and cancer: criteria for a causal relation. Semin
Cancer Biol. 14:453–471. 2004.
|
|
3
|
Newcomb PA, Bush AC, Stoner GL, et al: No
evidence of an association of JC virus and colon neoplasia. Cancer
Epidemiol Biomarkers Prev. 13:662–666. 2004.
|
|
4
|
Ricciardiello L, Laghi L, Ramamirtham P,
et al: JC virus DNA sequences are frequently present in the human
upper and lower gastrointestinal tract. Gastroenterology.
119:1228–1235. 2000.
|
|
5
|
Young NS and Brown KE: Parvovirus B19. N
Engl J Med. 350:586–597. 2004.
|
|
6
|
Li Y, Wang J, Zhu G, et al: Detection of
parvovirus B19 nucleic acids and expression of viral VP1/VP2
antigen in human colon carcinoma. Am J Gastroenterol.
102:1489–1498. 2007.
|
|
7
|
Kerr JR, Barah F, Cunniffe VS, et al:
Association of acute parvovirus B19 infection with new onset of
acute lymphoblastic and myeloblastic leukaemia. J Clin Pathol.
56:873–875. 2003.
|
|
8
|
Perreault N and Beaulieu JF: Use of the
dissociating enzyme thermolysin to generate viable human normal
intestinal epithelial cell cultures. Exp Cell Res. 224:354–364.
1996.
|
|
9
|
von Poblotzki A, Gigler A, Lang B, Wolf H
and Modrow S: Antibodies to parvovirus B19 NS-1 protein in infected
individuals. J Gen Virol. 76:519–527. 1995.
|
|
10
|
Huang da W, Sherman BT and Lempicki RA:
Bioinformatics enrichment tools: paths toward the comprehensive
functional analysis of large gene lists. Nucleic Acids Res.
37:1–13. 2009.
|
|
11
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009.
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
|
|
13
|
Doerig C, Hirt B, Antonietti JP and Beard
P: Nonstructural protein of parvoviruses B19 and minute virus of
mice controls transcription. J Virol. 64:387–396. 1990.
|
|
14
|
Raab U, Beckenlehner K, Lowin T, et al:
NS1 protein of parvovirus B19 interacts directly with DNA sequences
of the p6 promoter and with the cellular transcription factors
Sp1/Sp3. Virology. 293:86–93. 2002.
|
|
15
|
Momoeda M, Wong S, Kawase M, Young NS and
Kajigaya S: A putative nucleoside triphosphate-binding domain in
the nonstructural protein of B19 parvovirus is required for
cytotoxicity. J Virol. 68:8443–8446. 1994.
|
|
16
|
Moffatt S, Yaegashi N, Tada K, Tanaka N
and Sugamura K: Human parvovirus B19 nonstructural (NS1) protein
induces apoptosis in erythroid lineage cells. J Virol.
72:3018–3028. 1998.
|
|
17
|
Kajigaya S, Fujii H, Field A, et al:
Self-assembled B19 parvovirus capsids, produced in a baculovirus
system, are antigenically and immunogenically similar to native
virions. Proc Natl Acad Sci USA. 88:4646–4650. 1991.
|
|
18
|
Brown KE, Anderson SM and Young NS:
Erythrocyte P antigen: cellular receptor for B19 parvovirus.
Science. 262:114–117. 1993.
|
|
19
|
Ozawa K and Young N: Characterization of
capsid and noncapsid proteins of B19 parvovirus propagated in human
erythroid bone marrow cell cultures. J Virol. 61:2627–2630.
1987.
|
|
20
|
Saikawa T, Anderson S, Momoeda M, Kajigaya
S and Young NS: Neutralizing linear epitopes of B19 parvovirus
cluster in the VP1 unique and VP1–VP2 junction regions. J Virol.
67:3004–3009. 1993.
|
|
21
|
Rosenfeld SJ, Yoshimoto K, Kajigaya S, et
al: Unique region of the minor capsid protein of human parvovirus
B19 is exposed on the virion surface. J Clin Invest. 89:2023–2029.
1992.
|
|
22
|
Kawase M, Momoeda M, Young NS and Kajigaya
S: Most of the VP1 unique region of B19 parvovirus is on the capsid
surface. Virology. 211:359–366. 1995.
|
|
23
|
Lu J, Zhi N, Wong S and Brown KE:
Activation of synoviocytes by the secreted phospholipase A2 motif
in the VP1-unique region of parvovirus B19 minor capsid protein. J
Infect Dis. 193:582–590. 2006.
|
|
24
|
Zádori Z, Szelei J, Lacoste MC, et al: A
viral phospholipase A2 is required for parvovirus infectivity. Dev
Cell. 1:291–302. 2001.
|
|
25
|
Luo W and Astell CR: A novel protein
encoded by small RNAs of parvovirus B19. Virology. 195:448–455.
1993.
|
|
26
|
St Amand J and Astell CR: Identification
and characterization of a family of 11-kDa proteins encoded by the
human parvovirus B19. Virology. 192:121–131. 1993.
|
|
27
|
St Amand J, Beard C, Humphries K and
Astell CR: Analysis of splice junctions and in vitro and
in vivo translation potential of the small, abundant B19
parvovirus RNAs. Virology. 183:133–142. 1991.
|
|
28
|
Fan MM, Tamburic L, Shippam-Brett C,
Zagrodney DB and Astell CR: The small 11-kDa protein from B19
parvovirus binds growth factor receptor-binding protein 2 in vitro
in a Src homology 3 domain/ligand-dependent manner. Virology.
291:285–291. 2001.
|
|
29
|
Moffatt S, Tanaka N, Tada K, et al: A
cytotoxic nonstructural protein, NS1, of human parvovirus B19
induces activation of interleukin-6 gene expression. J Virol.
70:8485–8491. 1996.
|
|
30
|
Wan Z, Zhi N, Wong S, et al: Human
parvovirus B19 causes cell cycle arrest of human erythroid
progenitors via deregulation of the E2F family of transcription
factors. J Clin Invest. 120:3530–3544. 2010.
|