Introduction
Oral tongue squamous cell carcinoma (OTSCC) is one
of the most common types of malignant tumors of the oral and
maxillofacial region, comprising 32.3% of all cases of oral cancer.
Although the pathogenesis of OTSCC remains unclear, it has been
suggested that it may involve the mutation and abnormal expression
of multiple genes (1). The
prognosis for patients with OTSCC is relatively poor and the risk
of relapse is high, which may be attributable to the highly
invasive nature of OTSCC cells, the frequent movements of the
tongue and the rich blood supply to the tongue. Therefore, early
lymph node and late distant metastases are extremely common in
tongue cancer.
With recent advancements in molecular biology,
molecular genetics and related disciplines, study regarding
potential treatments for OTSCC has focused on gene therapy
(2). The first tumor suppressor
gene with phosphatase activity identified in humans was the
phosphatase and tensin homolog (PTEN) gene. Studies have
shown that the PTEN gene undergoes significant mutations and
deletions in a variety of tumors, including melanoma, breast,
prostate and endometrial cancer, resulting in a loss of protein
expression or dysfunction, thereby contributing to tumor
development (3,4). Additional study has indicated that
mutations and deletions in the PTEN gene also promote the
growth and development of gliomas and head and neck cancers
(5).
The epithelial-mesenchymal transition (EMT) refers
to the process whereby skin-derived precursor cells undergo
phenotypic changes during the embryonic and tumor progression
stages. E-cadherin, a 120-kDa transmembrane glycoprotein, interacts
with α-, β- and γ-catenins, as well as the E-cadherin/catenin
complex, to then associate with the actin microfilament system of
the cell, regulating tissue and morphological changes. Thus, the
expression and functional status of the E-cadherin/catenin complex
within the tumor influences cell separation and adhesion, mediating
tumor invasion (6). Vimentin is
often considered a marker for tumors of mesenchymal origin, and
vimentin expression is increased in numerous epithelial tumors and
is closely associated with tumor invasion. SNAIL is a zinc finger
protein that binds to the promoter of the E-cadherin gene, inducing
tumor cell EMT (7). Vimentin, SNAIL
and E-cadherin are closely associated with EMT and may be useful
indicators of EMT. Recent study has examined the ability of EMT to
induce tumor invasion and metastasis (8); however, thus far, no reports have
investigated the impact of the PTEN tumor suppressor gene on
EMT in OTvSCC (9).
In the present study, PTEN was overexpressed
in SCC-4 cells, and the effects of PTEN expression on the
proliferation and apoptosis of OTSCC cells was examined. In
addition, the correlation between the invasiveness of
PTEN-transfected OTSCC cells and EMT-associated markers was
investigated.
Materials and methods
Reagents and antibodies
SCC-4 cells were provided by the Ninth People’s
Hospital of Shanghai Jiaotong University (Shanghai, China) and
originally purchased from the Cell Bank of the Chinese Academy of
Sciences (Shanghai, China). Mouse anti-human vimentin polyclonal
antibodies were purchased from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA). Mouse anti-human E-cadherin monoclonal
antibodies, rabbit anti-human Akt polyclonal antibodies, rabbit
anti-phospho-Akt polyclonal antibodies and rabbit anti-human
Bcl-2-interacting mediator of cell death (BIM) polyclonal
antibodies were obtained from Jiamay Biotech (Beijing, China).
Primer synthesis and DNA sequencing were performed by Wuhan Ying Qi
Biotechnology Co., Ltd. (Wuhan, China), MTT and dimethyl sulfoxide
were purchased from Promega Corporation (Madison, WI, USA), and
tetramethylethylenediamine and sodium dodecyl sulfate were
purchased from Sigma-Aldrich (St. Louis, MO, USA).
Cell culture
The OTSCC SCC-4 cell line was maintained at 37°C in
a humidified incubator with an atmosphere of 5% CO2 in
Dulbecco’s modified Eagle’s medium (DMEM)/F12 medium (Hyclone;
Thermo Fisher Scientific, Rockford, IL, USA) supplemented with 10%
fetal bovine serum (Thermo Fisher Scientific) without any
antibiotics.
Human tissue specimens
A total of 40 human tissue specimens were collected
from individuals who underwent surgery at the Department of
Surgery, First Affiliated Hospital of Liaoning Medical University
(Jinzhou, China) between January 2007 and December 2010. Clinical
information is summarized in Table
I. All patients provided written informed consent and were
assessed for PTEN expression. This study was approved by the
human ethics committee of Liaoning Medical University.
 | Table IClinicopathological features of
OTSCC. |
Table I
Clinicopathological features of
OTSCC.
| Clinicomorphological
parameters | n (%) |
|---|
| Age, years |
| <20 | 6 (15.0) |
| 20–40 | 14 (35.0) |
| >40 | 20 (50.0) |
| Gender |
| Female | 26 (65.0) |
| Male | 14 (35.0) |
| Normal oral
tissuesa |
| Female | 10 (66.7) |
| Male | 5 (33.4) |
| OTSCC tissues |
| Female | 18 (72.0) |
| Male | 7 (28.0) |
Immunohistochemistry
Immunohistochemistry was performed using a
Streptavidin-Biotin Complex (SABC) kit (Wuhan Boster Biological
Technology, Ltd., Wuhan, China) according to the manufacturer’s
instructions. Briefly, the tissue sections were deparaffinized in
xylene for 20 min and then dehydrated in graded alcohol solutions,
followed by detection using the avidin-biotin complex method by
SABC kit. The endogenous peroxidase activity was blocked by
immersing the sections in 3% H2O2 in methanol
for 30 min. For antigen retrieval, the sections were heated in 0.01
M citrate buffer (pH 6.0; Shanghai Xin Biological Technology Co.,
Ltd., Shanghai, China) for 15 min. The sections were then treated
with 10% normal rabbit serum for 30 min, followed by incubation
with mouse anti-human PTEN monoclonal antibodies [1:100 dilution;
Santa Cruz Biotechnology (Shanghai) Co., Ltd., Shanghai, China] at
4°C overnight. Following incubation with a biotin-conjugated
secondary antibody, incubation was performed with streptavidin
solution at 37°C for 20 min, followed by incubation with SABC
reagents at 37°C for 30 min. The tissues were stained with
3,3′-diaminobenzidine (Chinese sales platform ELISA kits, Shanghai,
China). Negative and positive controls were conducted in each run
of immunohistochemistry. A total of five to six fields from each
tissue section was selection, and 100 cells from each field were
counted (Countstar automated cell counter, Biomen Biosystems Co.,
Ltd., Guangzhou, China) at a final magnification of ×400 (Olympus
BX43; Shanghai Zeshi Photoelectric Technology Co., Ltd., Shanghai,
China). The evaluation was performed by two independent
pathologists, without any prior knowledge of each patient’s
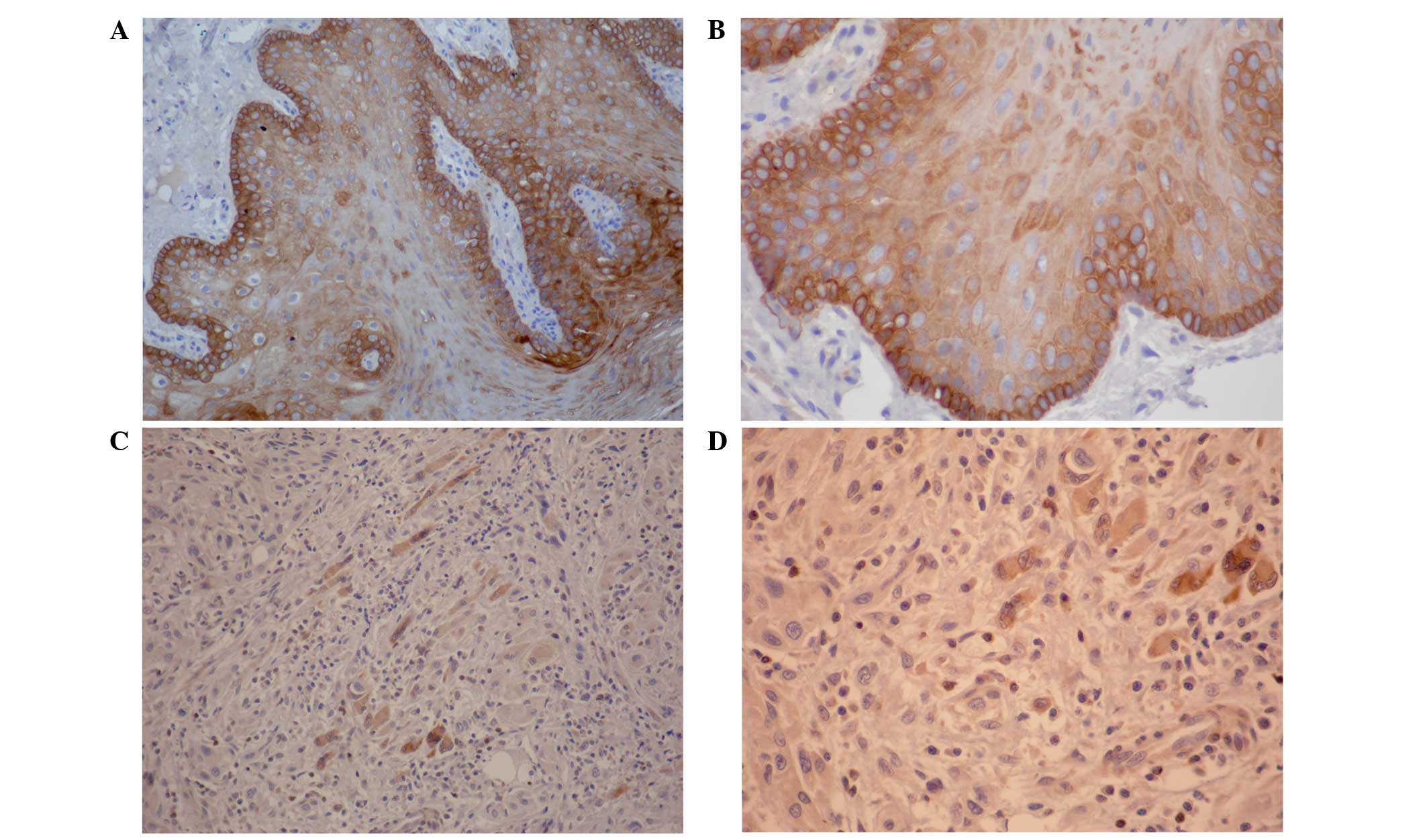
clinical information (Fig. 1).
Expression of PTEN mRNA by reverse
transcription-polymerase chain reaction (RT-PCR)
Total RNA was extracted from SCC-4 cells using the
TRIzol method (Gibco-BRL, Carlsbad, CA, USA). PTEN total RNA
was amplified using RT-PCR. The amplification system and conditions
were based on the manufacturer’s instructions stated in the Takara
One-Step RNA PCR kit (Takara Bio, Inc., Shiga, Japan). Primer
Premier 5.0 software (PREMIER Biosoft, Palo Alto, CA, USA) was used
to design primers for the PTEN gene based on sequences
retrieved from GenBank. The upstream and downstream primers were
5′-GCCGAATTCGACTTTTGTAATTTGTGTA-3′ and
5′-CCGCTCGAGCAGTCGCTGCAACCATCCA-3′, respectively, with EcoRI
restriction sites introduced to the 5′ ends for nucleotide
protection. For RT-PCR, each reaction was carried out as follows:
Denaturation at 94°C for 5 min; 60 cycles of 94°C for 60 sec, 60°C
for 60 sec and 72°C for 1.5 min; and extension at 72°C for 10
min.
Transfection with the PTEN eukaryotic
expression plasmids
SCC-4 cells growing at the logarithmic growth phase
were seeded onto six-well plates and transfections were performed
when the cells reached 70–80% confluence, using the Lipofectamine
2000 reagent kit (Invitrogen Life Technologies, Carlsbad, CA, USA)
according to the manufacturer’s instructions. Experiments were
carried out using three groups of cells: Cells transfected with
phosphorylated (p)-enhanced green fluorescent protein (EGFP)-PTEN
recombinant plasmid; cells transfected with pEGFP-N1 empty plasmid;
and untransfected control cells. The intracellular expression of
GFP was observed under a fluorescence microscope (Olympus BX43) at
24, 48 and 72 h following transfection. At 48 h following
transfection, the cells were also cultured in DMEM selection medium
containing 800 μg/ml G418. Cloned SCC-4 cells exhibiting stable
expression of PTEN were then filtered for amplification and
culture, and stable cell lines in the logarithmic growth phase were
used for follow-up tests.
Western blotting
Following transfection, cells were subjected to
total protein extraction. Protein content was measured against
bovine serum albumin, which was used as the standard. Proteins were
separated by polyacrylamide gel electrophoresis on 10% gels,
transferred to polyvinylidene fluoride membranes and blocked for 1
h in 5% skimmed milk. Following washing with Tris-buffered saline
and Tween 20, membranes were incubated with the primary antibodies
(Akt, 1:1,000; phospho-Akt, 1:1,000; BIM, 1:1,000; and PTEN, 1:500)
and then incubated overnight at 4°C (PTEN) or room temperature
(Akt, phospho-Akt and BIM). Next, the membranes were washed and
incubated with a sheep anti-mouse polyclonal horseradish
peroxidase-conjugated secondary antibody (Jiamay Biotech) for 1–2 h
at room temperature. The Biospectrum imaging system (Beijing Dequan
Development Trading Co., Ltd., Beijing, China) was used for image
capture. The optical density of each band was measured using ImageJ
software (National Institutes of Health).
Cell proliferation assays
Following transfection, cells at the logarithmic
growth phase were used for cell proliferation assays. The cell
concentration was adjusted to 1×104 cells/ml, and 100 μl
cell suspension was added to each well of a 96-well plate (n=42
wells/cell group). Every day between day one and seven, cells were
counted to measure proliferation and MTT assays were performed
according to standard instructions to confirm cell viability (n=3
wells/day per cell group for each of the proliferation and MTT
assays).
Statistical analysis
All statistical analyses were performed using SPSS
software, version 13.0 (SPSS, Inc., Chicago, IL, USA). Differences
between all three groups were determined using analysis of variance
tests, while differences between two groups were analyzed by
Student’s t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Loss of PTEN protein expression in
OTSCC
The expression of PTEN protein in OTSCC was
primarily cytoplasmic, with infrequent nuclear localization
(Fig. 1). PTEN expression was
observed in 15 out of 15 (100%) normal oral tissues; however, loss
of PTEN expression was apparent in all 25 OTSSC specimens (Fig. 1). When comparing the expression of
PTEN in OTSCC with normal oral tissue, the frequency of PTEN loss
was statistically significant (P<0.001). Notably, the expression
of PTEN was not associated with age, gender or histological grade
(data not shown).
PTEN mRNA expression levels in each cell
group
The results of 1% agarose gel electrophoresis
revealed that the three groups of cells appeared in ~1,200-bp
bands. However, the pEGFP-PTEN-SCC-4 group was found to exhibit
significantly thicker bands (Fig.
2D).
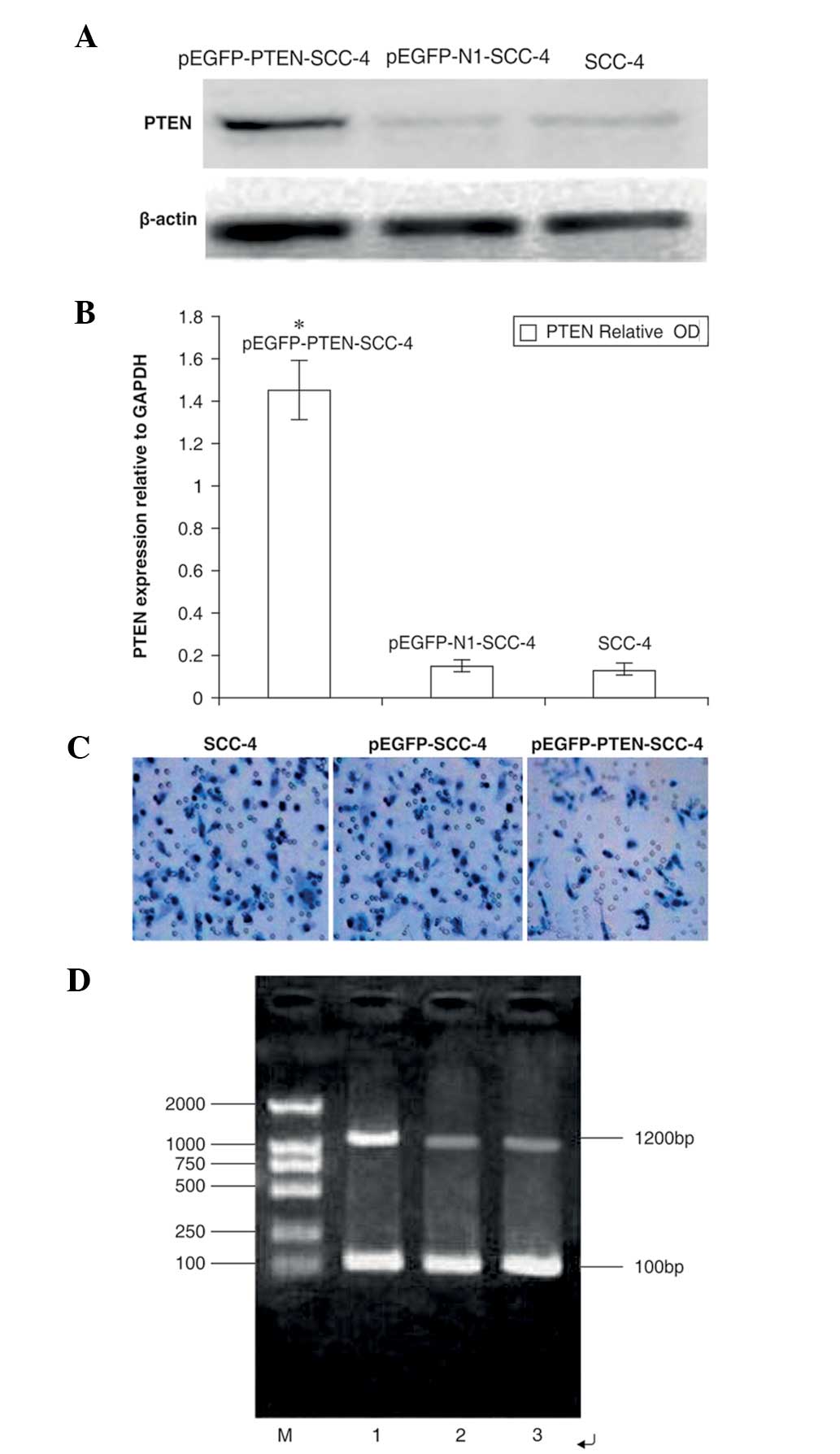 | Figure 2Effects of PTEN vector transfection on
PTEN expression in SCC-4 cells. (A) Untransfected cells (lane 1)
and cells transfected with empty vector (lane 2) or PTEN (lane 3)
were subjected to western blotting using anti-PTEN antibodies.
β-Actin was used as a loading control. (B) Quantification of
western blots to measure PTEN protein expression. The OD of PTEN
(relative to GAPDH) was as follows: 1.45±0.14, 0.17±0.02 and
0.15±0.03 in the pEGFP-PTEN-SCC-4, pEGFP-N1-SCC-4 and SCC-4 groups,
respectively. (C) Effects of PTEN expression on the invasion of
SCC-4 cells. Untransfected SCC-4 cells and cells transfected with
pEGFP-SCC-4 or pEGPF-PTEN-SCC-4 were subjected to Transwell
invasion assays. The number of cells with penetrated membranes in
the pEGPF-PTEN-SCC-4 group was evidently less than in the SCC-4 and
pEGFP-SCC-4 groups. (D) Reverse transcription-polymerase chain
reaction for PTEN gene mRNA expression. M, DNA maker; 1,
pEGFP-PTEN-SCC-4; 2, pEGFP-N1-SCC-4; and 3, SCC-4. PTEN,
phosphatase and tensin homolog; pEGFP, phosphorylated enhanced
green fluorescent protein; SCC, squamous cell carcinoma; OD,
optical density. |
PTEN expression in SCC-4 cells
Western blotting results revealed that the
transfected group exhibited a significantly increased brightness of
bands when compared with the other groups. The optical density of
PTEN protein expression (relative to GAPDH) in PTEN-transfected
cells was 1.07±0.15, which was identified to be significantly
different when compared with that of cells transfected with the
empty vector and untransfected cells (0.62±0.11 and 0.57±0.08,
respectively; P<0.05; Fig. 2A and
B). These results indicated that transfection with the
PTEN-containing plasmid induced overexpression of exogenous
PTEN.
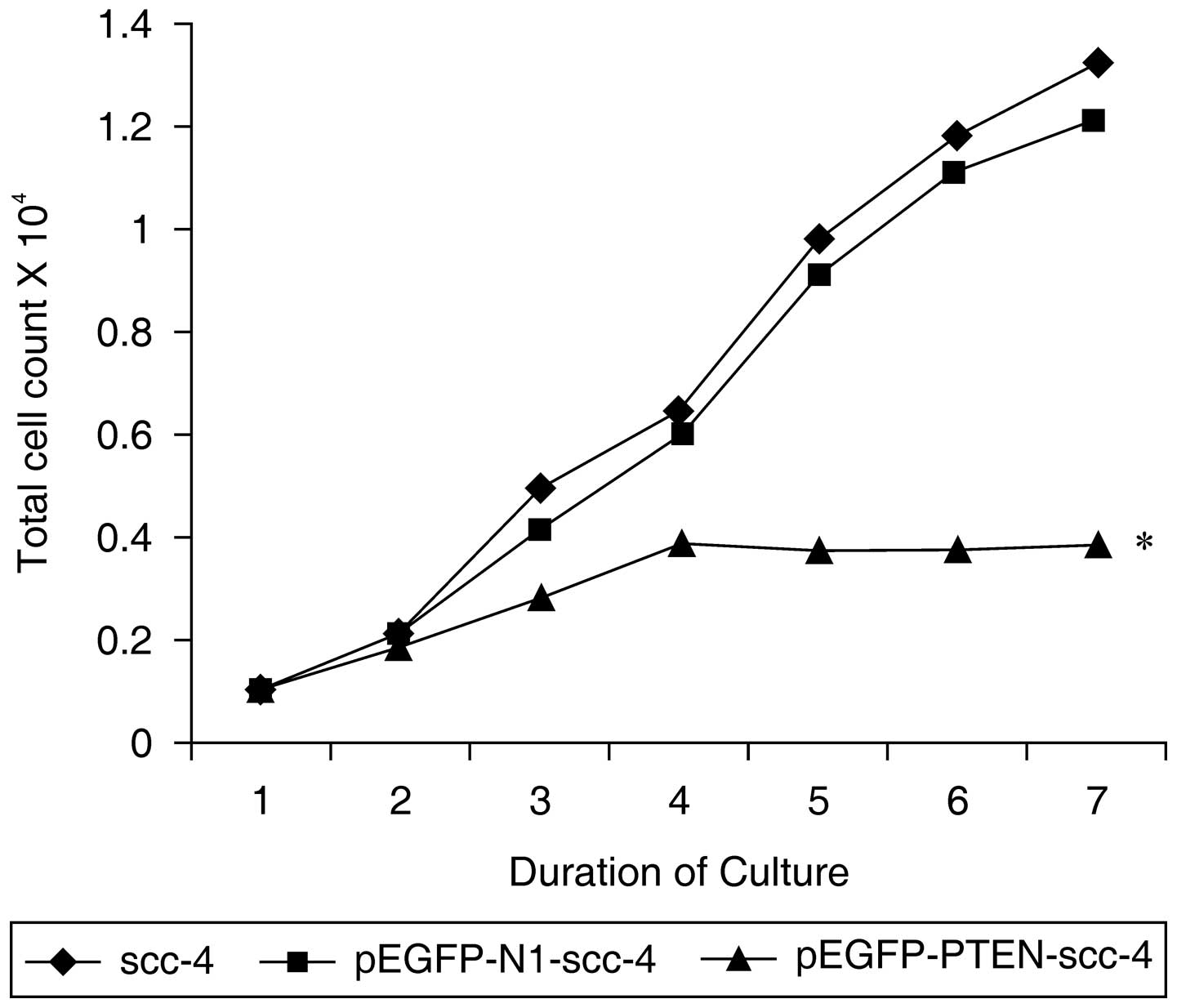
PTEN overexpression suppresses SCC-4 cell
proliferation
The effects of PTEN overexpression on SCC-4 cell
proliferation were investigated. Notably, transfection with the
pEGFP-PTEN-SCC-4 vector resulted in a significant reduction in cell
proliferation when compared with that of the pEGFP-N1-SCC-4 and
SCC-4 groups following the third day of culture (P<0.01;
Fig. 3). These results indicated
that PTEN expression suppresses SCC-4 cell proliferation.
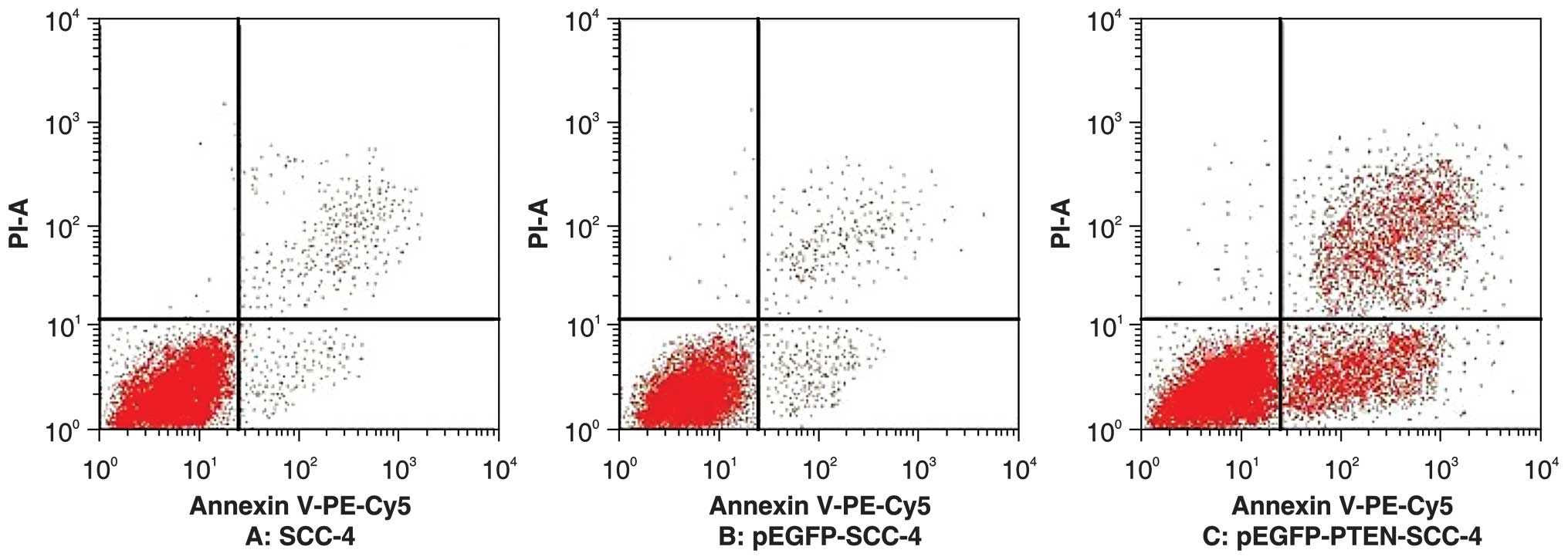
Overexpression of PTEN induces apoptosis
in SCC-4 cells
Flow cytometry analysis of Annexin
V-phycoerythrin-Cy5/propidium iodide double-staining indicated that
the percentage of apoptotic cells was significantly higher in
PTEN-GFP-transfected cells when compared with that of cells
transfected with the empty vector and untransfected control cells
(48.1±2.6, 1.2±0.7 and 1.4±0.9%, respectively) 48 h following
transfection. No significant differences in apoptotic rate between
the control and empty vector groups were identified (t=0.3;
P>0.05). However, a significant difference was identified
between the apoptotic rate of the experimental group and that of
the other two groups (t1=30.2; t2=29.4; P<0.01) (Fig. 4).
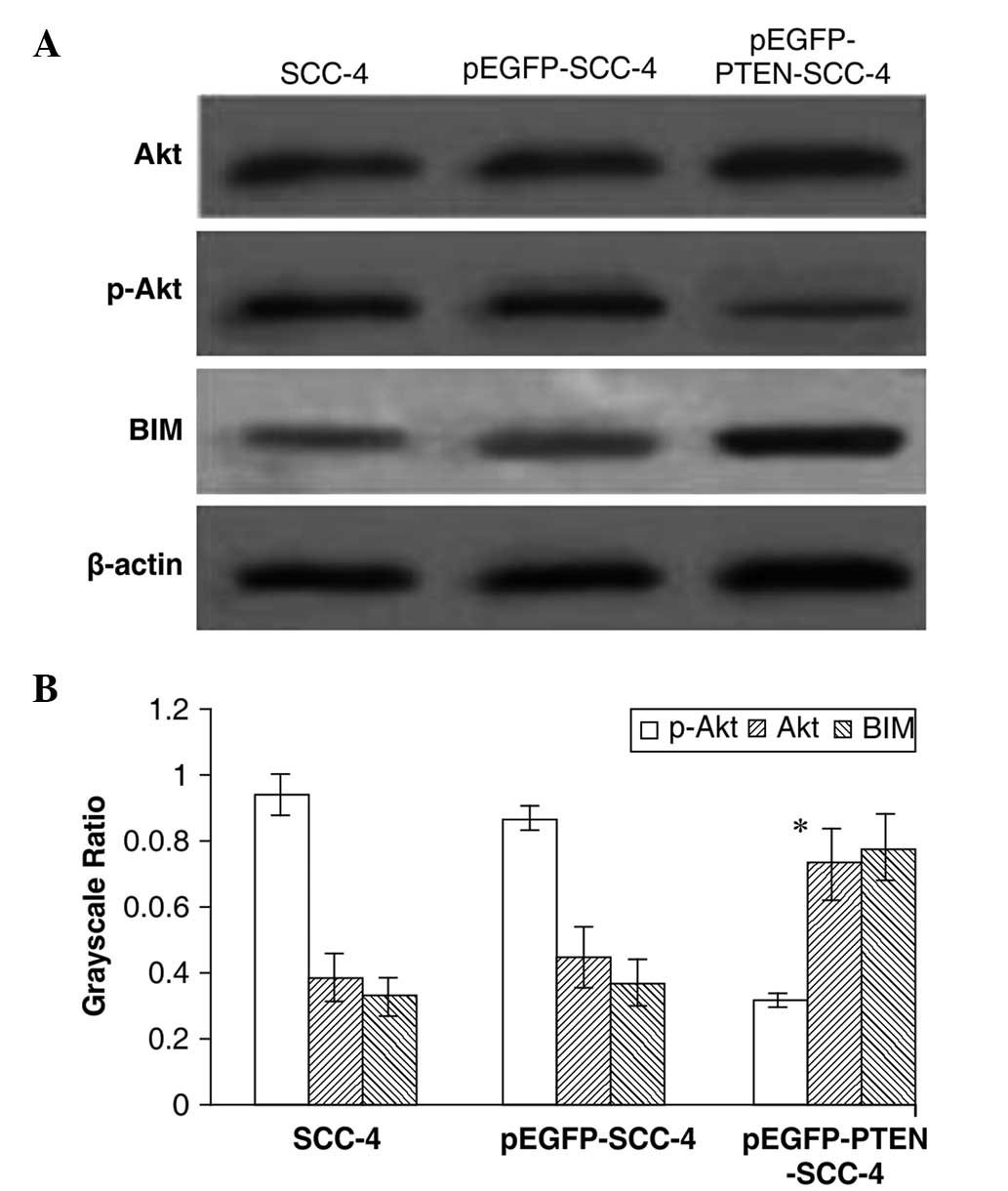
Effects of PTEN expression on Akt,
phospho-Akt and BIM levels in tongue cancer
No significant differences in total Akt expression
were identified among transfected and untransfected cells following
western blotting. However, while phospho-Akt levels were 0.94±0.13
for SCC-4, 0.87±0.04 for pEGFP-SCC-4 and 0.32±0.02 for
pEGFP-PTEN-SCC-4 (P>0.05), while phospho-Akt levels were
significantly reduced in PTEN-transfected cells (P<0.05). The
expression of BIM, a pro-apoptotic, BH3-only protein member of the
Bcl-2 family which is critical in apoptosis (10), was 0.33±0.06 for SCC-4, 0.37±0.07
for pEGFP-SCC-4 and 0.78±0.10 for pEGFP-PTEN-SCC-4; however, it was
found to be significantly upregulated in PTEN-transfected cells
(P<0.01) (Fig. 5).
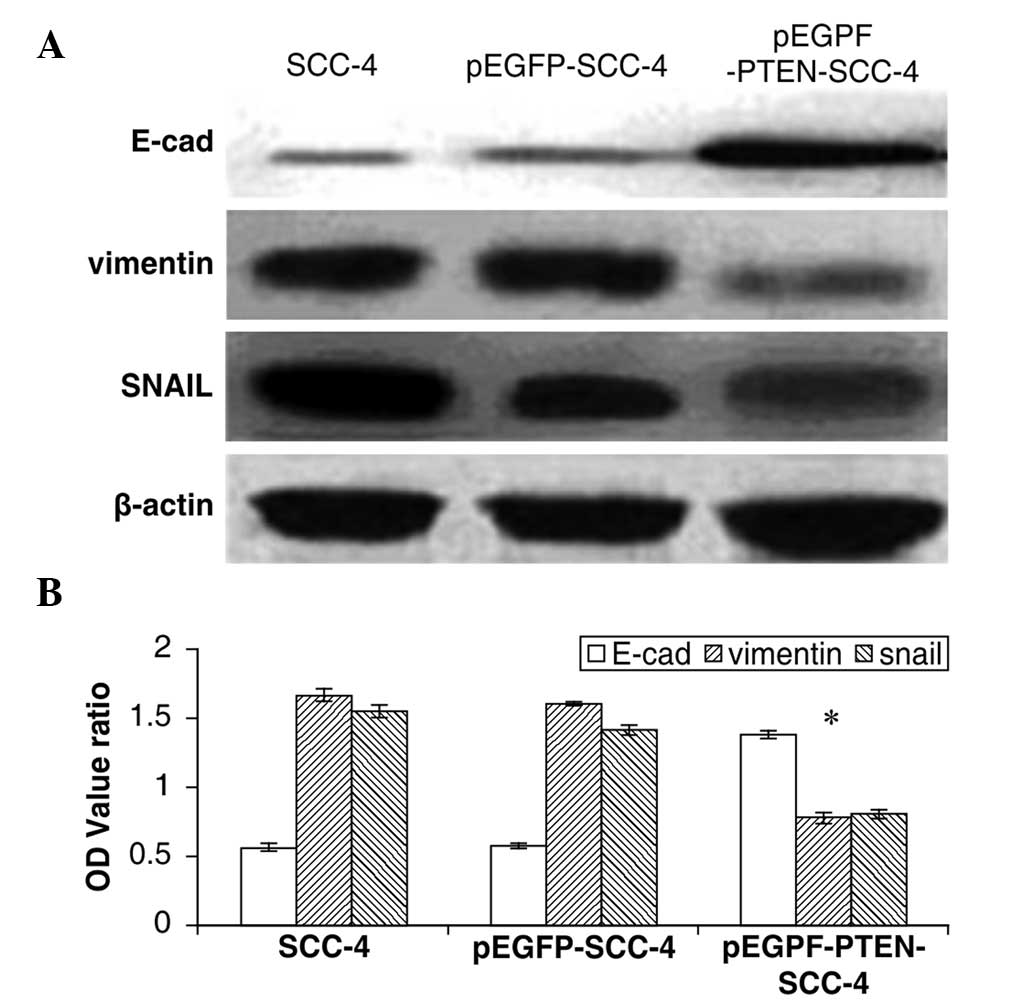
Overexpression of PTEN inhibits SCC-4
cell invasion
Cells in the control group (SCC-4), empty vector
group (pEGFP-SCC-4) and transfected group (pEGFP-PTEN-SCC-4) were
cultured in Transwell invasion chambers for 36 h. For the 30
visually selected fields from each group, the numbers of cells
invading through the membrane were 82±5, 80±4 and 42±5,
respectively (Fig. 2C). These
results demonstrated that PTEN expression caused a significant
reduction in SCC-4 invasion when compared with the control
(P<0.01). In addition, a statistically significant difference
was identified when comparing the expression of the three proteins
in the transfected and control groups (P<0.05). However, no
significant differences were identified between the protein
expression of the empty vector and control groups (P>0.05). The
results of the western blotting revealed that PTEN expression
significantly induced the expression of E-cadherin (SCC-4,
0.556±0.022; pEGFP-SCC-4, 0.573±0.013; and pEGFP-PTEN-SCC-4,
1.375±0.026) and suppressed the expression of SNAIL (SCC-4,
1.554±0.041; pEGFP-SCC-4, 1.412±0.036; and pEGFP-PTEN-SCC-4,
0.801±0.027) and vimentin (SCC-4, 1.667±0.045; pEGFP-SCC-4,
1.593±0.013; and pEGFP-PTEN-SCC-4, 0.778±0.032) (P<0.01)
(Fig. 6). These results suggested
that PTEN may block OTSCC cell invasion by inhibiting the EMT
process.
Discussion
PTEN is a tumor suppressor gene with dual
phosphatase activity. However, its mechanism of action is not fully
understood (11). At present, PTEN
is considered to convert dephosphorylated phosphatidylinositol
(3,4,5)-triphosphate (PIP3) to
phosphtidylinositol (4,5)-bisphosphate, thereby blocking
PIP3-mediated activation of protein kinase B/Akt and
suppressing the growth and development of tumors (12). In addition, PTEN has been shown to
function in the nucleus and thus may be important in
transcriptional regulation, however, its nuclear targets remain
unclear (13).
In the present study, PTEN expression was detected
in OTSCC specimens and the effects of PTEN expression in SCC-4
cells transfected with a PTEN expression vector were investigated.
Using this model, PTEN expression was found to exert a tumor
suppressor function, which significantly reduced the proliferation
capacity of SCC-4 cells, thus confirming the function of PTEN in
the malignant behavior of OTSCC.
Deletion of the BIM gene may lead to
tumorigenesis (14). Previous
studies have shown that numerous anticancer drugs, including those
for lung (15) and ovarian
(12) cancer, induce tumor cell
apoptosis via the increased expression of BIM. In the current
study, the results of the western blotting indicated that BIM
expression increased following transfection with a PTEN
expression vector in SCC-4 cells, suggesting that PTEN expression
may affect the phosphatidylinositide 3-kinase (PI3K)/Akt signaling
pathways via upregulation of BIM. The results of this study also
confirmed that it is possible to induce apoptosis of SCC-4 cells
via in vitro PTEN transfection, possibly through negative
regulation of the PI3K/Akt signaling pathway and increased
expression of the transcription factor Akt and the pro-apoptotic
protein BIM, thereby enhancing SCC-4 cell apoptosis. Since the
PI3K/Akt pathway involves a variety of additional factors, further
targets for gene therapy are available.
It has been demonstrated that PTEN suppresses tumor
development by promoting apoptosis of tumor cells and regulating
the cell cycle, reducing the invasiveness of tumor cells in
esophageal cancer and melanoma (16). Similarly, in the current study, PTEN
expression was reduced in advanced tumors and tumors undergoing
lymph node metastasis.
Loss of E-cadherin and upregulation of vimentin are
hallmarks of the EMT. In addition,
E-cadherinlow/vimentin may be used as an indicator of
tumor prognosis, whereby a small ratio indicates poor prognosis
(18). Furthermore, the
transcription factor SNAIL is important in EMT. Häyry et al
(19) revealed that in 73 cases of
OTSCC, SNAIL expression and depth of invasion were found to
significantly correlate, demonstrating that SNAIL directly affects
tumor invasion and metastasis. Thus, the differential expression of
these three proteins in OTSCC cells indicates the varying degrees
of EMT. In addition to the results of the current study regarding
cell invasion, these results show that PTEN regulates the
expression of E-cadherin vimentin and SNAIL, indicating the
involvement of PTEN in EMT. Thus, when considering the invasive
abilities of the different groups of cells, we hypothesize that the
invasive ability of OTSCC cells is associated with the EMT
process.
PTEN gene deletion induces EMT via the
PI3K/Akt signaling pathway (20),
thereby increasing the invasive ability of tumor cells.
Additionally, Leslie et al (21) reported that colorectal cancer cells
become spindle-shaped following treatment with LY294002, a
PI3K/Akt-specific inhibitor, an effect which was accompanied by a
reduced expression of E-cadherin and increased invasiveness of the
cells, further supporting the role of the PTEN/E-cadherin signaling
axis in EMT.
In conclusion, the PTEN gene is closely associated
with the development of SCC. Expression of the PTEN gene may
inhibit the growth of tongue SCC cells. This may present a possible
line of gene therapy. However, OTSCC is a solid tumor and its
structure and biological characteristics are extremely complex.
Therefore, the SCC-4 cell line does not fully reflect the tumor
itself and cannot present the complete genetic characteristics of
OTSCC. Thus, PTEN must be studied using animal models to elucidate
its detailed mechanism of action.
Acknowledgements
This study was supported by The First Affiliated
Hospital of Liaoning Medical University (Jinzhou, China), and the
generous donations of cancer patients and their families. The
authors would like to thank Professor Shu Lin Gao and Professor Qin
Yu for valuable support, and Editage/Cactus Communications Pvt.
Ltd. (Mumbai, India) for the editing of the study.
References
|
1
|
Zhang Z, Pan J, Li L, Wang Z, Xiao W and
Li N: Survey of risk factors contributed to lymphatic metastasis in
patients with oral tongue cancer by immunohistochemistry. J Oral
Pathol Med. 40:127–134. 2011.
|
|
2
|
Nagabhushan Kalburgi S, Khan NN and Gray
SJ: Recent gene therapy advancements for neurological diseases.
Discov Med. 15:111–119. 2013.
|
|
3
|
Barbosa M, Henrique M, Pinto-Basto J,
Claes K and Soares G: Prostate cancer in Cowden syndrome: somatic
loss and germline mutation of the PTEN gene. Cancer Genet.
204:224–225. 2011.
|
|
4
|
Feng ZZ, Chen JW, Yang ZR, Lu GZ and Cai
ZG: Expression of PTTGI and PTEN in endometrial carcinoma:
correlation with tumorigenesis and progression. Med Oncol.
29:304–310. 2012.
|
|
5
|
Huang SH and O’Sullivan B: Oral cancer:
Current role of radiotherapy and chemotherapy. Med Oral Patol Oral
Cir Bucal. 18:e233–e240. 2013.
|
|
6
|
Vasko V, Espinosa AV, Scouten W, et al:
Gene expression and functional evidence of
epithelial-to-mesenchymal transition in papillary thyroid carcinoma
invasion. Proc Natl Acad Sci USA. 104:2803–2808. 2007.
|
|
7
|
Tang CH and Tsai CC: CCL2 increases MMP-9
expression and cell motility in human chondrosarcoma cells via the
Ras/Raf/MEK/ERK/NF-κB signaling pathway. Biochem Pharmacol.
83:335–344. 2012.
|
|
8
|
Esteller M: Cancer epigenomics: DNA
methylomes and histone-modification maps. Nat Rev Genet. 8:286–298.
2007.
|
|
9
|
Tshering Vogel DW, Zbaeren P and Thoeny
HC: Cancer of the oral cavity and oropharynx. Cancer Imaging.
10:62–72. 2010.
|
|
10
|
Yip KW, Li A, Li JH, et al: Potential
utility of BimS as a novel apoptotic therapeutic molecule. Mol
Ther. 10:533–544. 2004.
|
|
11
|
Keniry M and Parsons R: The role of PTEN
signaling perturbations in cancer and in targeted therapy.
Oncogene. 27:5477–5485. 2008.
|
|
12
|
Wu B, Wang X, Chi ZF, et al: Ursolic
acid-induced apoptosis in K562 cells involving upregulation of PTEN
gene expression and inactivation of the PI3K/Akt pathway. Arch
Pharm Res. 35:543–548. 2012.
|
|
13
|
Gu T, Zhang Z, Wang J, Guo J, Shen WH and
Yin Y: CREB is a novel nuclear target of PTEN phosphatase. Cancer
Res. 71:2821–2825. 2011.
|
|
14
|
Rachmiel A, Aizenbud D and Peled M:
Enhancement of bone formation by bone morphogenetic protein-2
during alveolar distraction: an experimental study in sheep. J
Periodontol. 75:1524–1531. 2004.
|
|
15
|
Liu H, Liang SL, Kumar S, Weyman CM, Liu W
and Zhou A: Statins induce apoptosis in ovarian cancer cells
through activation of JNK and enhancement of Bim expression. Cancer
Chemother Pharmacol. 63:997–1005. 2009.
|
|
16
|
Ou Y, Ma L, Ma L, Huang Z, Zhou W, Zhao C,
Zhang B, Song Y, Yu C and Zhan Q: Overexpression of cyclin B1
antagonizes chemotherapeutic-induced apoptosis through PTEN/Akt
pathway in human esophageal squamous cell carcinoma cells. Cancer
Biol Ther. 14:45–55. 2013.
|
|
17
|
Ma WJ, Lv GD, Tuersun A, et al: Role of
microRNA-21 and effect on PTEN in Kazakh’s esophageal squamous cell
carcinoma. Mol Biol Rep. 38:3253–3260. 2011.
|
|
18
|
Iwatsuki M, Mimori K, Fukagawa T, et al:
The clinical significance of vimentin-expressing gastric cancer
cells in bone marrow. Ann Surg Oncol. 17:2526–2533. 2010.
|
|
19
|
Häyry V, Mäkinen LK, Atula T, et al: Bmi-1
expression predicts prognosis in squamous cell carcinoma of the
tongue. Br J Cancer. 102:892–897. 2010.
|
|
20
|
Pickhard AC, Margraf J, Knopf A, Stark T,
et al: Inhibition of radiation induced migration of human head and
neck squamous cell carcinoma cells by blocking of EGF receptor
pathways. BMC Cancer. 11:3882011.
|
|
21
|
Leslie NR, Yang X, Downes CP and Weijer
CJ: PtdIns(3,4,5) P(3)-dependent and -independent roles for PTEN in
the control of cell migration. Curr Biol. 17:115–125. 2007.
|