Introduction
The vascular niche is a major compartment of the
tumor microenvironment, and therefore may have a role in the
initiation, progression and metastasis of tumors (1,2). The
abundant blood supply provided by the vascular niche enables
physicians to use enhanced computed tomography (CT) during clinical
examinations. Enhanced CT is often used in order to distinguish
between malignant and benign lung lesions. The role of angiogenesis
in the pathogenesis of lung cancer is well recognized. Endothelial
cells (ECs) constitute the vast majority of vascular cells,
however, the origin of vascular ECs in the tumor microenvironment
remains controversial. It is believed that vascular ECs may be
derived from normal ECs adjacent to the tumor, from progenitor ECs
in the peripheral circulation or from undifferentiated cancer cells
(3).
The interaction between cancer cells and ECs may
underlie the process of angiogenesis within the tumor
microenvironment. Tumor cells may directly or indirectly promote
the phenotypic conversion of normal ECs (4). In addition, the ECs may be induced by
tumor cells to partake in angiogenesis, which in turn affects the
behavior of the tumor cells (5).
Cluster of differentiation (CD)31 and CD146 are
unique markers of normal ECs (6,7). By
contrast, CD105 is rarely expressed by normal ECs, but is strongly
expressed by activated and rapidly proliferating ECs (8). The glucose-regulated protein-78
(GRP-78) is expressed by tumor cells, whose roles are drawing
growing attention (9,10). Vascular endothelial growth factor
(VEGF) and basic fibroblast growth factor (bFGF), and their
corresponding receptors, are involved in the process of normal
physiological angiogenesis in developing human and mouse embryos
(11). However, the pivotal
mechanism and signaling pathways involved in EC phenotype
conversion and the cross-talk between lung cancer cells and ECs is
yet to be elucidated.
The present study aimed to investigate the
interaction between lung cancer cells and ECs, and to identify the
potential role of VEGF and bFGF in the angiogenesis of lung
cancer.
Materials and methods
Cell culture
The human lung adenocarcinoma A549 cell line was
provided by the Biomedical Ultrasonic and Gynecological Oncology
Laboratory, West China Second University Hospital (Sichuan, China).
The cell line was cultured in RPMI 1640 medium containing 10% fetal
bovine serum at 37°C in 5% CO2.
Primary culture of human umbilical vein
endothelial cells (HUVECs)
The primary culturing of the HUVECs was performed
according to the methodology described by Baudin (12). The present study was approved by the
Ethics Committee of the West China Hospital and written informed
consent was obtained from all patients. Umbilical cords were
donated from 5 women who gave birth naturally, in the Obstetrics
and Gynecology Department of West China Second University Hospital.
Informed consent was obtained from the patients. First, the HUVECs
were isolated from the umbilical vein vascular wall. The umbilical
cord, measuring 10–30 cm, was then washed using 1×
phosphate-buffered saline (PBS) several times to remove the
remnants of blood. Next, the umbilical cord was digested using 0.2%
collagenase II at 37°C for 15 min. The cell suspension was then
collected and centrifuged in a closed tube at 95 × g for 10 min.
The supernatant was discarded carefully. Next, the cell pellets
were suspended in 4 ml endothelial cell medium-2 (ECM-2). The cells
were then dissociated by gentle aspiration and repulsing using a
pipette. The samples were incubated at 37°C in 5% CO2.
The medium was replaced every 24 h. Subsequent to primary
culturing, the samples were passaged two to four times prior to
use.
Immunofluorescence (IF)
The HUVECs were seeded into 24-well plates at a
density of ~104 cells per well and cultured for two days
in ECM. Next, the cells were washed and fixed in 4%
paraformaldehyde at 4°C for 15 min, washed with 1× PBS, blocked
with 5% bovine serum albumin for 30 min and then incubated with the
primary antibody at room temperature for 60 min. The expression of
CD31, CD146, and GRP-78 was detected using monoclonal rabbit
anti-human antibodies (dilution, 1:100; ab180175, ab75769 and
ab108615, respectively; Abcam, Cambridge, UK) CD105 was detected
using a monoclonal mouse anti-human antibody (dilution, 1:100;
ab11414, Abcam). The negative control samples were concurrently
incubated with 1× PBS. The secondary antibodies were fluorescein
isothiocyanate-conjugated goat anti-rabbit and goat anti-mouse
(dilution, 1:50; ZF-0311 and ZF-0312, respectively; ZSGB-BIO,
Beijing, China). The nuclei were stained using DAPI (dilution,
1:1,000). The samples were viewed under an Olympus BX51
fluorescence microscope (Olympus, Tokyo, Japan) and analyzed using
Image-Pro Plus 6.0 software (IPP 6.0; Media Cybernetics Inc.,
Rockville, MD, USA).
Co-culture of the HUVECs and A549 cell
line
The co-culture of the HUVECs and A549 cells was
performed using the Millicell co-culture system (Millipore,
Billerica, MA, USA). In total, ~103 HUVECs were
inoculated in the upper chamber of the culture system (pore size, 4
μm), while the A549 cells were plated in the lower chamber.
Subsequent to a 12-h incubation with Dulbecco’s modified Eagle’s
medium (upper chamber, 0.2 ml; lower chamber, 1.25 ml), the old
medium was discarded, and new ECM-2 was added prior to an eight-day
culture period. The HUVECs were incubated alone in the upper
chamber, and the A549 cells incubated in the lower chamber were
used as a negative control.
The WST-1 assay (Roche, Basel, Switzerland),
performed as previously described (12), was used to quantify the rate of
proliferation. Briefly, for the HUVECs cultured alone, ~1,000
HUVECs were added to the upper well and ~1.25 ml ECM-2 (without any
cells) was added to the lower well. For the A549 cells cultured
alone, ~1,000 A549 cells were added to the lower well and 0.2 ml
ECM-2 was added to the upper well (without any cells). For the
co-culture assay, ~1,000 HUVECs were added to the upper well (the
volume was ~0.2 ml) and ~1,000 A549 cells were added to the lower
well (the volume was ~1.25 ml). For each well, 40 μl WST was added
to each well according to the specification and the cells were
incubated for 4 h. Next, 100-μl samples were obtained from each
well and transferred to 96-well plates. The absorbance was measured
by a Varioskan Flash microplate reader (Thermo Fisher Scientific,
Waltham, MA, USA) at a wavelength of 440 nm. The assay was repeated
three times and the mean absorbance was recorded. Optical density
(OD) was used to compare the rate of proliferation.
ELISA
For the collection of the conditioned media,
serum-free DMEM was used in this assay. Next, the supernatant was
harvested, centrifuged at 855 × g for 5 min and then stored at
−80°C. The expression of VEGF and bFGF in the conditioned medium of
the co-culture system and the single-culture groups was measured
using an ELISA kit (Life Technologies, Carlsbad, CA, USA). The
standard curves were constructed separately with the quantities of
VEGF and bFGF selected as 800, 400, 200, 100, 50 or 25 ng/ml, and
25, 12.5, 6.25, 3.125, 1.5625 and 0.78125 ng/ml, respectively. The
absorbance was measured at 450 nm.
Statistical analysis
The purity of the HUVECs was quantified by counting
the ratio of CD31- or CD146-positive cells. Five fields of vision
(magnification, ×40) per well and a total of three wells were
randomly selected for analysis. The intensities of CD105 and GRP-78
were analyzed using IPP 6.0. The mean OD (MOD) was defined as the
ratio of the sum of the integrated OD and the sum of the area. The
results are presented as the mean ± standard error of the mean. The
independent sample t-test was used. P<0.05 was used to indicate
a statistically significant difference.
Results
Isolation of HUVECs from the umbilical
vein
The isolated cells were incubated with CD31 and
CD146 antibodies and cultured according to the aforementioned
method. The proteins were positively stained in the cytoplasm. The
HUVECs demonstrated a high expression level of CD31 (range,
92.45–98.25%; mean, 96.15%) and CD146 (range, 96.20–98.00%; mean,
97.30%) compared with the control (Fig.
1A and B).
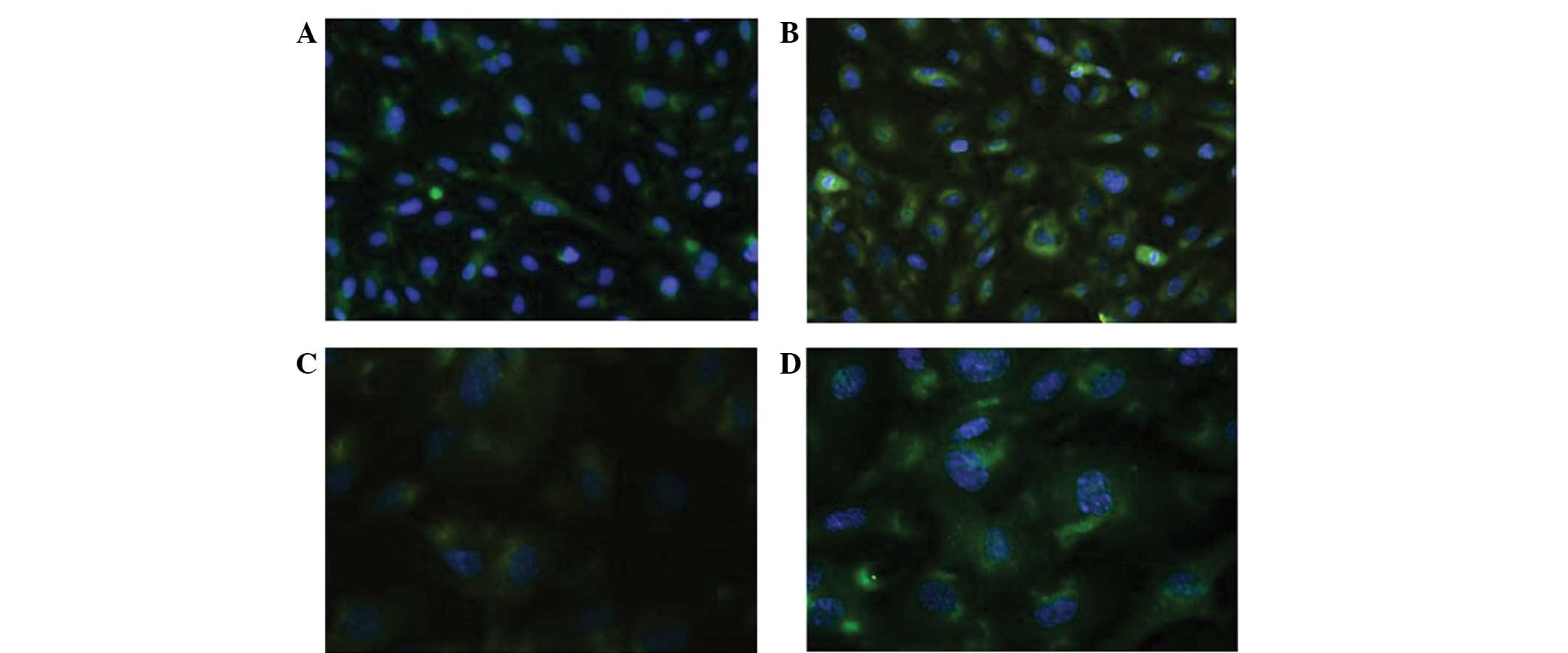
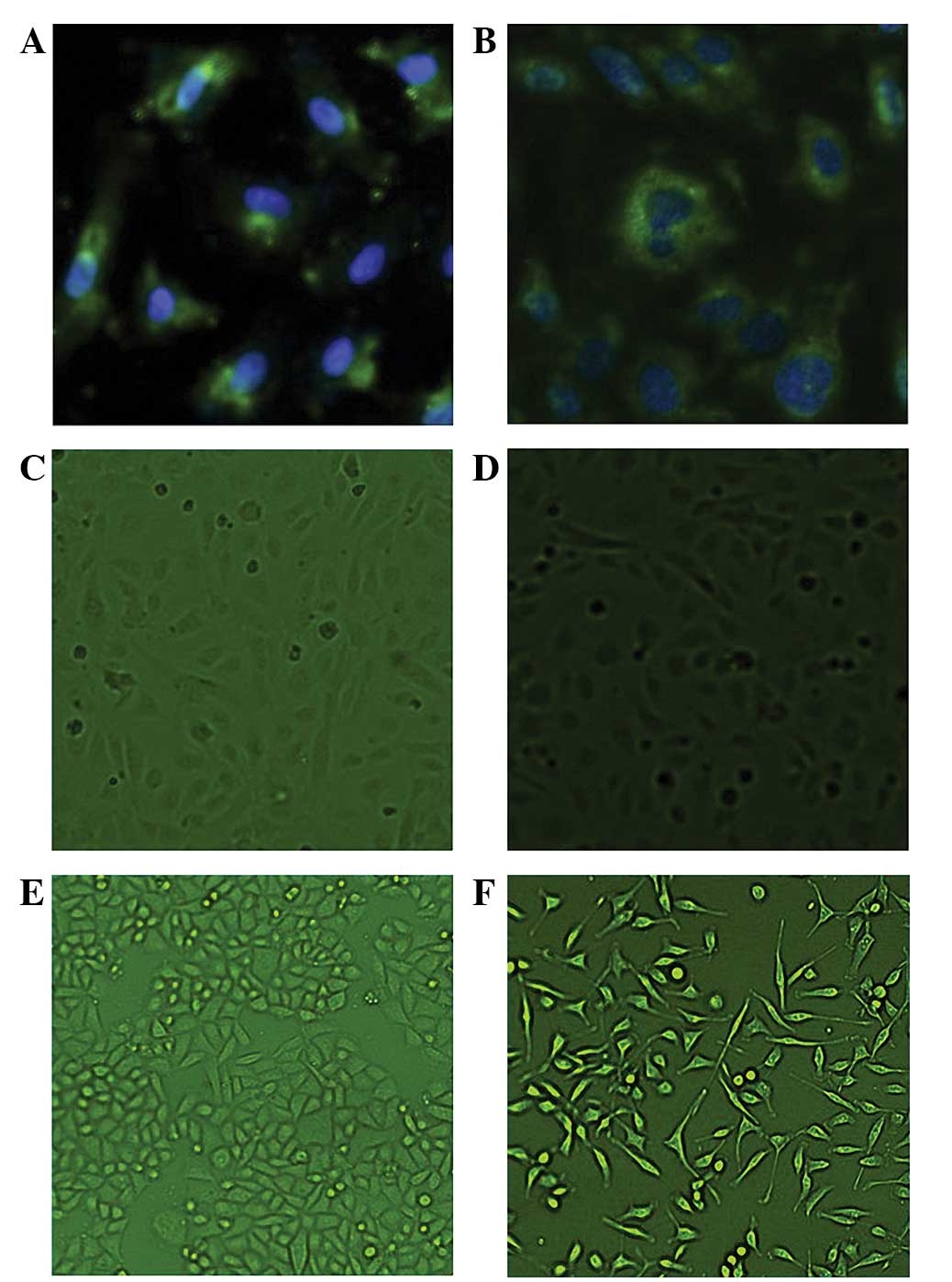 | Figure 1Immunofluorescence revealing the
characteristics of the human umbilical vein endothelial cells
(HUVECs) and the A549 cell line. (A) Cluster of differentiation
(CD)31 and (B) CD146 expression in HUVECs (magnification, ×40). (C)
The HUVECs that were cultured alone were teardrop-shaped
(magnification, ×10) (D) The HUVECs in the co-culture group
contained larger nuclei and elongated cell bodies, similar to the
vascular ECs (magnification, ×10). (E) The A549 cells were fusiform
or polygonal in shape when cultured alone (magnification, ×10). (F)
In the co-culture system, however, these cells exhibited larger
nuclei, with certain cells appearing binuclear or polynuclear. In
addition, the cells became stretched and scattered (magnification,
×10). |
Interaction between HUVECs and the A549
cell line
Morphological changes of the HUVECs
and the A549 cell line
The HUVECs in the co-culture system exhibited a
different morphology to those cultured alone. The HUVECs cultured
alone were teardrop-shaped, with large nuclei and elongated cell
bodies, similar in appearance to the vascular ECs cultured with the
A549 cells (Fig. 1C and D).
The A549 cells were fusiform or polygonal in shape
when cultured alone. In the co-culture system, however, the A549
cells exhibited relatively larger nuclei, with certain cells
demonstrating binuclear or even polynuclear forms. Furthermore, the
cells appeared stretched and scattered (Fig. 1E and F).
A549 cells promote the proliferation
of HUVECs in the co-culture system
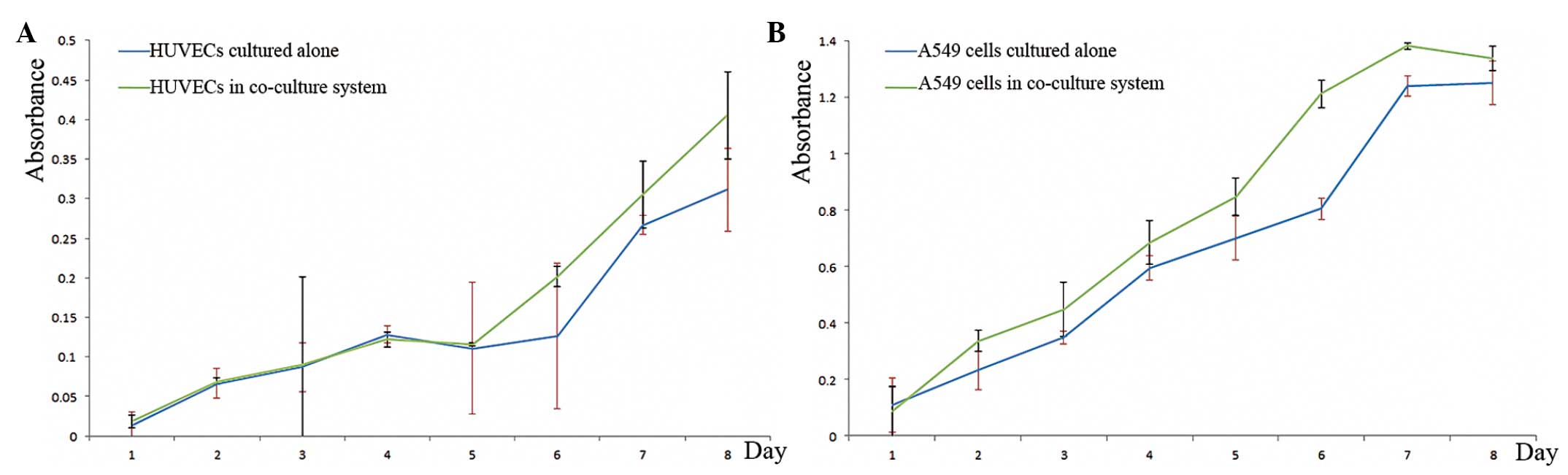
The proliferation rate of the HUVECs was analyzed
using the WST-1 assay, and the absorbance was measured. Compared
with the control group, the proliferation of the HUVECs in the
co-cultured system was significantly higher on day five
(absorbance, 0.8152 vs. 1.0274; P<0.001). However, between days
one and four there was no difference in terms of proliferation
between the groups (absorbance, 0.0741 vs. 0.0751; P=0.1845). On
average during the eight days, the absorbance of the HUVECs was
0.1660±0.1304 in the co-cultured system and 0.1389±0.1003 in the
control group (P=0.0448) (Fig.
2A).
HUVECs promote the proliferation of
A549 cells in the co-culture system
The proliferation of the A549 cell line was affected
by the HUVECs. The mean absorbance of the co-culture group was
significantly higher than that of the control group (0.7927±0.4877
vs. 0.6610±0.4304) (P=0.036)(Fig.
2B).
Phenotype conversion of HUVECs and the
A549 cell line
CD105 is a unique marker of activated endothelial
cells in the tumor microenvironment (8). The expression of CD105 in the HUVECs
was detected using IF, and the MOD was calculated following eight
days of co-culture. CD105 staining was positive in the cytoplasm of
the HUVECs in the co-cultured system and in the negative control
group. The MOD was significantly higher in the co-culture group
than in the control group (40.3247±3.3343 vs. 23.2515±5.2713)
(P=0.0027). The expression of CD105 was significantly higher in the
co-culture group than in the control group. Furthermore, the effect
of proliferation was downregulated (Fig. 3A).
GRP-78 is a molecular chaperone within the
endoplasmic reticulum, and its expression is associated with the
differentiation, invasion and drug-resistance of cancer cells
(9,10). In the present study, the expression
of GRP-78 was detected using IF, and the MOD was calculated. GRP-78
was expressed in the cytoplasm of the A549 cells. The MODs of the
co-culture and control groups were 58.5987±7.1013 and
39.1734±2.2089, respectively. The expression of GRP-78 was
significantly higher in the co-culture group than in the control
group (P=0.0397), and the effect of proliferation was eliminated
(Fig. 3B).
Contents of VEGF and bFGF in the
medium
In the Millicell co-culture system, the HUVECs and
A549 cells were unable to interact via direct contact, which
suggests that cytokines may be involved in the cellular
interactions. The expression levels of the cytokines, VEGF and
bFGF, were measured in the conditioned medium of the co-cultured
and control groups. The HUVECs that were cultured alone secreted
extremely small amounts of VEGF and bFGF. The levels of VEGF were
measured at 53.34±6.52 and 21.47±1.63 ng/ml (P<0.0001) and the
levels of bFGF were 5.06±0.24 and 2.95±0.28 ng/ml (P<0.001) in
the medium of the A549 single-cultured and co-culture groups,
respectively.
In order to investigate the role of VEGF and bFGF
without interference, HUVECs were cultured alone with serum-free
DMEM. In total, 50 ng/ml VEGF or 1 ng/ml bFGF was artificially
added to the medium, as recommended by Bai et al (13). The proliferation of the HUVECs was
measured using a WST-1 assay. Compared with the HUVECs cultured in
serum-free DMEM, the proliferation of the HUVECs in the VEGF(+) or
bFGF(+) group was significantly higher (P<0.001). When the two
factors were added consecutively, the effect upon HUVEC
proliferation was significantly greater than that observed
following the single addition of either factor alone (proliferation
curves not shown).
Discussion
Lung cancer is the leading cause of cancer-related
mortality worldwide, and is therefore known for its high rates of
morbidity and mortality. The highly progressive nature of the
disease and its ability to metastasize make it incurable, and for
any of its subtypes, the five-year survival rate is only ~15%
(14). Overall, non-small cell lung
cancer (NSCLC) accounts for 85% of all types of lung cancer
(3). The rapid proliferation and
metastatic nature of NSCLC cells relies upon support from tumor
blood vessels in the form of angiogenesis (15). As tumor ECs (TECs) differ from
normal ECs, tumor blood vessels demonstrate abnormal morphology.
The interactions between TECs are aberrant, which leads to the
formation of complex tumor blood vessels and uneven vessel
diameters (16). In addition, TECs
are unable to form normal monolayers, which leads to an incomplete
barrier function of the tumor blood vessels and the occurrence of
leakiness (17).
Due to the difficultly of isolating and culturing
TECs from tumor tissues, few studies have focused on them.
Furthermore, it has been suggested that the cells may lose their
unique features following isolation. For these reasons, TECs are
usually replaced by HUVECs. For a long time, TECs were considered
to be phenotypically and cytogenetically normal. Following their
successful isolation, it was realized that they differ from normal
ECs in phenotype and express 46 unique tumor endothelial markers
(18). In addition, TECs were
identified to be karyotypically aneuploid, unlike normal ECs, which
are diploid (19).
In the present study, the normal HUVECs expressed
CD31 and CD146, which are two unique markers of normal ECs
(20). The HUVECs exhibited a
phenotype conversion when cultured with A549 cells. The phenotype
of the co-cultured HUVECs became similar to that of the TECs, with
a significant upregulation of CD105. CD105 (also known as endoglin)
is an accessory protein belonging to the transforming growth
factor-β receptor family, which is expressed in activated vascular
ECs and has a key role in angiogenesis (7). The function of CD105 makes it
important during embryonic development, and genetic mutations of
this protein have been revealed to lead to Osler-Weber-Rendu
syndrome (21). In solid tumors,
the overexpression of CD105 is correlated with metastases and
decreased survival (22).
Cancer cells can affect the phenotype and
proliferation of TECs in a co-culture system, but TECs may also in
turn affect tumor cells. The present study identified that A549
cells demonstrate morphological and phenotypic changes. When
cultured with the HUVECs, the proliferation of the A549 cells
increased. In addition, GRP-78 expression was detected in the A549
cells. GRP-78 was selected as a novel biomarker, as its level is
associated with the differentiation, metastasis, chemoresistance
and prognosis of tumor cells (10).
Angiogenesis is known to promote tumor progression and metastasis
by providing cells with the nutrients and oxygen necessary for
growth and metastasis (23). The
upregulation of GRP-78 in the A549 cells indicated that these cells
were more prone to metastasis or progression to an advanced stage.
This result coincided with that of a previous study (24). Despite this, the mechanism by which
HUVECs affect A549 cells is yet to be elucidated.
Angiogenesis is a key process involved in
physiological and pathological environments. A number of factors,
including the key mediators, VEGF and FGF, take part in tumor
angiogenesis (25). The present
study demonstrated that HUVECs secrete extremely small amounts of
VEGF. The expression of VEGF in the medium containing the A549
cells, however, was significantly higher than that observed in the
co-culture group. Therefore, it was hypothesized that VEGF may act
in a paracrine manner to affect the growth and proliferation of
HUVECs. Under physiological conditions, angiogenesis is tightly
regulated by a balance between anti- and pro-angiogenic factors.
However, cancer cells are able to unbalance the related factors,
and therefore promote angiogenesis. The VEGF secreted by cancer
cells destroys the balance between anti- and pro-angiogenic
factors, which promotes angiogenesis. VEGF is a homodimeric
glycoprotein, which consists of two identical 23-kDa subunits. VEGF
was first identified in the medium of an animal tumor model
(26). VEGF is closely associated
with lung diseases, such as pulmonary hypertension, acute
respiratory distress syndrome, asthma and emphysema. In particular,
high levels of VEGF have been identified in cases of lung
adenocarcinoma (27). This
association is highlighted by the fact that bevacizumab, a
humanized monoclonal antibody against VEGF, is approved by the Food
and Drug Administration to treat advanced NSCLC (25).
In addition to VEGF, FGF is a factor involved in
cancer angiogenesis. FGF, which is a heparin-binding factor, can be
divided into acidic FGF (aFGF) and basic FGF (bFGF) (28). bFGF can affect smooth muscle cells
and ECs. Furthermore, it acts as a chemoattractant during the
proliferation of ECs, which in turn promotes angiogenesis (29). The present study demonstrated that
extremely small amounts of bFGF were secreted by the HUVECs, but
that high levels were identified in the medium containing the A549
cells. In addition, only a small amount of bFGF was required for
proliferation compared with VEGF (1 vs. 50 ng/ml). Therefore, it
can be concluded that bFGF not only directly promotes HUVEC
proliferation, but also acts indirectly by inducing the action of
VEGF (28,29). In a previous study, bFGF was induced
by hypoxia-inducible factor-α (29). As a result, the hypoxia induced an
upregulation in the expression of bFGF in the tumor
microenvironment, which promoted angiogenesis and resulted in the
formation of a number of immature blood vessels. Due to the
presence of immature blood vessels, cancer tissues are usually
hypoxic (15). The hypoxic
environment in turn promotes cancer cells to secrete more bFGF.
Therefore, an antibody against bFGF may be more useful than one
against VEGF. Although brivanib, a novel, orally available and
selective receptor tyrosine kinase inhibitor that targets bFGF and
VEGF receptors, is currently under clinical evaluation, further
clinical trials are required (30).
Using a co-culture system, the present study
examined the interplay between the A549 lung cancer cell line and
tumor HUVECs. It was revealed that lung cancer cells may affect
HUVECs in a paracrine manner. The secretion of VEGF and bFGF by
cancer cells is found to potentially have key roles in promoting
the proliferation of HUVECs. The A549 cells were also affected by
the HUVECs at the same time. The upregulation of GRP-78 in cancer
cells reflects that these tumor cells become more invasive and
prone to metastasis. Angiogenesis is a very complicated mechanism
and is likely to involve more factors. Additional research into
angiogenesis and its potential underlying mechanisms is
required.
Acknowledgements
The authors would like to thank staff at the
Department of Thoracic Surgery and at the Biomedical Ultrasonics
and Gynecological Oncology Laboratory, West China Hospital, Sichuan
University. The present study was funded by a grant from the
National Science Foundation (no. 81272595).
References
|
1
|
Nyberg P, Salo T and Kalluri R: Tumor
microenvironment and angiogenesis. Front Biosci. 13:6537–6553.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fan F, Schimming A, Jaeger D and Podar K:
Targeting the tumor microenvironment: focus on angiogenesis. J
Oncol. 2012:2812612012. View Article : Google Scholar
|
|
3
|
Herbst RS, Heymach JV and Lippman SM: Lung
Cancer. N Engl J Med. 359:1367–1380. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Witz IP: Tumor-microenvironment
interactions: dangerous liaisons. Adv Cancer Res. 100:203–229.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Galligioni E and Ferro A: Angiogenesis and
antiangiogenic agents in non-small cell lung cancer. Lung Cancer.
34(Suppl 4): S3–S7. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Takase Y, Kai K, Masuda M, Akashi M and
Tokunaga O: Endoglin (CD105) expression and angiogenesis status in
small cell lung cancer. Pathol Res Pract. 206:725–730. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dallas NA, Samuel S, Xia L, et al:
Endoglin (CD105): a marker of tumor vasculature and potential
target for therapy. Clin Cancer Res. 14:1931–1937. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tanaka F, Otake Y, Yanagihara K, et al:
Correlation between apoptotic index and angiogenesis in non-small
cell lung cancer: comparison between CD105 and CD34 as a marker of
angiogenesis. Lung Cancer. 39:289–296. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang Y, Wang W, Wang S, et al:
Down-regulation of GRP78 is associated with the sensitivity of
chemotherapy to VP-16 in small cell lung cancer NCI-H446 cells. BMC
Cancer. 8:3722008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee AS: GRP78 induction in cancer:
therapeutic and prognostic implications. Cancer Res. 67:3496–3499.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ballas MS and Chachoua A: Rationale for
targeting VEGF, FGF, and PDGF for the treatment of NSCLC. Onco
Targets Ther. 4:43–58. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Baudin B, Bruneel A, Bosselut N and
Vaubourdolle M: A protocol for isolation and culture of human
umbilical vein endothelial cells. Nat Protoc. 2:481–485. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bai Y, Leng Y, Yin G, et al: Effects of
combinations of BMP-2 with FGF-2 and/or VEGF on HUVECs angiogenesis
in vitro and CAM angiogenesis in vivo. Cell Tissue Res.
356:109–121. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Burstein HJ and Schwartz RS: Molecular
origins of cancer. N Engl J Med. 358:5272008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hida K, Kawamoto T, Ohga N, Akiyama K,
Hida Y and Shindoh M: Altered angiogenesis in the tumor
microenvironment. Pathol Int. 61:630–637. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
McDonald DM and Baluk P: Significance of
blood vessel leakiness in cancer. Cancer Res. 62:5381–5385.
2002.PubMed/NCBI
|
|
17
|
Hashizume H, Baluk P, Morikawa S, et al:
Openings between defective endothelial cells explain tumor vessel
leakiness. Am J Pathol. 156:1363–1380. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
St Croix BS, Rago C, Velculescu V, et al:
Genes expressed in human tumor endothelium. Science. 289:1197–1202.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hida K, Hida Y, Amin DN, et al:
Tumor-associated endothelial cells with cytogenetic abnormalities.
Cancer Res. 64:8249–8255. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tormin A, Li O, Brune JC, et al: CD146
expression on primary nonhematopoietic bone marrow stem cells is
correlated with in situ localization. Blood. 117:5067–5077. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Santibanez JF, Letamendia A,
Perez-Barriocanal F, et al: Endoglin increases eNOS expression by
modulating Smad2 protein levels and Smad2-dependent TGF-beta
signaling. J Cell Physiol. 210:456–468. 2007. View Article : Google Scholar
|
|
22
|
Lebrin F, Deckers M, Bertolino P and Ten
Dijke P: TGF-beta receptor function in the endothelium. Cardiovasc
Res. 65:599–608. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bergers G and Benjamin LE: Tumorigenesis
and the angiogenic switch. Nat Rev Cancer. 3:401–410. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dong D, Stapleton C, Luo B, et al: A
critical role for GRP78/BiP in the tumor microenvironment for
neovascularization during tumor growth and metastasis. Cancer Res.
71:2848–2857. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shojaei F: Anti-angiogenesis therapy in
cancer: current challenges and future perspectives. Cancer Lett.
320:130–137. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Senger DR, Galli SJ, Dvorak AM, Perruzzi
CA, Harvey VS and Dvorak HF: Tumor cells secrete a vascular
permeability factor that promotes accumulation of ascites fluid.
Science. 219:983–985. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bonnesen B, Pappot H, Holmstav J and Skov
BG: Vascular endothelial growth factor A and vascular endothelial
growth factor receptor 2 expression in non-small cell lung cancer
patients: relation to prognosis. Lung Cancer. 66:314–318. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Rusnati M and Presta M: Interaction of
angiogenic basic fibroblast growth factor with endothelial cell
heparan sulfate proteoglycans. Int J Clin Lab Res. 26:15–23. 1996.
View Article : Google Scholar
|
|
29
|
Przybylski M: A review of the current
research on the role of bFGF and VEGF in angiogenesis. J Wound
Care. 18:516–519. 2009. View Article : Google Scholar
|
|
30
|
Dempke WC and Zippel R: Brivanib, a novel
dual VEGF-R2/bFGF-R inhibitor. Anticancer Res. 30:4477–4483.
2010.PubMed/NCBI
|