Introduction
Lung cancer, the most common type of malignant
carcinoma, is the leading cause of cancer-associated mortality
worldwide (1). Despite the
availability of various therapeutic strategies, chemotherapy plays
a major role in the treatment of lung cancer (2). Thus far, alternative strategies to
chemotherapy have not resulted in significant improvement in the
survival rate of postoperative patients with lung cancer.
Therefore, the development of novel agents with selectivity against
critical apoptotic targets may provide a rational approach to the
treatment of cancer. Over the last decade, an abundance of
pharmacological evidence regarding the anticancer properties of
conventional natural Chinese products has been obtained (3,4). A number
of these anticancer agents have exerted their therapeutic action by
inducing apoptosis in human malignant cells (5–7).
Dracorhodin perchlorate (DP) is a synthetic analogue
of the anthocyanin red pigment dracorhodin, a major constituent of
traditional Chinese medicine, ‘dragon's blood’ (also known as
Daemonorops draco) (8–10). A previous study demonstrated that DP
inhibited the activation of the phosphoinositide 3-kinase/Akt and
nuclear factor-κB signaling pathways, and upregulated the
expression of p53 in gastric cancer cells (11). Furthermore, DP was found to inhibit
cell growth and trigger apoptosis in melanoma and breast cancer
cells (12,13). In addition, DP induced apoptosis
through the caspase pathways and generated reactive oxygen species
(ROS) in cervical cancer cells (14).
However, the effect of DP on human lung cancer cells has yet to be
reported.
Apoptosis is a type of programmed cell death that
occurs in multicellular organisms, and mitochondrial dysfunction is
considered to be the central executioner of the apoptosis pathway.
Mitochondrial membrane permeability is regulated by various
proteins, including p53, B cell lymphoma-2 (Bcl-2) and
Bcl-2-activated X protein (Bax) (15,16).
Depolarization of the mitochondrial membrane potential (MMP)
stimulates mitochondria to release apoptosis-inducing factor (AIF)
and various other proapoptotic molecules that eventually result in
the activation of caspase-3 (17,18). Thus,
AIF and caspase-3 are able to trigger chromatin condensation and
DNA degradation in order to induce programmed cell death.
The present study aimed to examine the possible
inhibitory effect of DP and the mechanism of DP-induced apoptosis
in SK-MES-1 lung cancer cells.
Materials and methods
Materials
The human lung squamous carcinoma cell line,
SK-MES-1, was purchased from the Cell Bank of the Chinese Academy
of Sciences (Shanghai, China). DP, which was purchased from the
Tonglian Pharmaceutical Co., Ltd. (Shanghai, China), was dissolved
in dimethyl sulfoxide (DMSO; Shengong Biotechnology Co., Shanghai,
China) to make a stock solution. Furthermore, fetal bovine serum
(FBS) was purchased from Gibco Life Technologies (Carlsbad, CA,
USA). The following reagents were purchased from Sigma-Aldrich (St.
Louis, MO, USA):
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT),
Hoechst 33342, Dulbecco's modified Eagle's medium (DMEM) and
Rhodamine 123 mitochondrial-specific fluorescent dye. The Cell
Cycle Analysis [propidium iodide (PI) + RNase A; catalog number
(cat. no.) C1052] and the Reactive Oxygen Species Assay kits were
purchased from Beyotime Institute of Biotechnology (Shanghai,
China). In addition, the BCA Protein Assay and the Annexin
V-fluorescein isothiocyanate (FITC) Apoptosis Detection kits were
purchased from Nanjing KeyGen Biotech. Co. Ltd. (Nanjing, China).
Polyclonal goat anti-rabbit antibodies against β-actin, (dilution,
1:2000; cat. no. 4967S) Bax (dilution, 1:1000; cat. no. 2772S),
Bcl-2 (dilution, 1:1000; cat. no. 2876S), pro-caspase-3 (dilution,
1:1000; cat. no. 9662P), poly(ADP-ribose) polymerase (PARP;
dilution, 1:1000; cat. no. 9542S), AIF (dilution, 1:1000; cat. no.
4642S), p53 (dilution, 1:1000; cat. no. 9282S) and phosphorylated
retinoblastoma (pRb; dilution, 1:1000; cat. no. 9306S), as well as
horseradish peroxidase (HRP)-conjugated secondary antibodies
(anti-rabbit IgG; dilution, 1:2000; cat. no. 7074P2), were
purchased from Cell Signaling Technology, Inc. (Shanghai, China). A
western blot detection kit was purchased from EMD Milipore
(Billerica, MA, USA).
Cell culture and treatments
SK-MES-1 human lung squamous carcinoma cells were
cultured in DMEM supplemented with 10% FBS and maintained at 37°C
in a 5% CO2 humidified atmosphere. Cells were treated
with DP dissolved in DMSO with a final DMSO concentration of 1%.
DMSO-treated cells were used as the controls in all the
experiments. The DMSO concentration was maintained at a
concentration of <0.01% in all the cell cultures and did not
exert any detectable effect on the rate of cell growth or
death.
Cell growth inhibition assay
The inhibition of cell growth was determined by
performing an MTT assay, as previously described (13). Briefly, SK-MES-1 cells were seeded in
96-well plates at a density of 1×104 cells/well, and
treated with various concentrations of DP (0, 10, 20, 40, 80 and
160 µM) for 24 h. Following treatment, the MTT reagent was added
(100 µl/ml) and the cells were incubated at 37°C for an additional
4 h. Next, 150 µl DMSO was added to dissolve the formazan crystals
and absorbance was read in a microplate reader (Varioskan Flash;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) at a wavelength
of 570 nm. The viable cell number was directly proportional to the
production of formazan. The growth assay was repeated three times
and the half maximal inhibitory concentration (IC50)
values were calculated using GraphPad Prism software (version 5.0;
GraphPad Software, Inc., La Jolla, CA, USA). The percentage of
inhibition was calculated as follows: Inhibitory ratio (%) =
(Acontrol / Asample)/Acontrol ×
100%, where Asample and Acontrol are the
absorbance values of the treated and control groups, respectively,
following incubation.
Flow cytometric cell cycle
analysis
A PI cell cycle detection kit (PI + RNase A) and
flow cytometry were used for cell cycle analysis. Briefly, SK-MES-1
cells were seeded in a 6-well plate at a density of
6×105 cells/well and treated with DP at concentrations
of 0, 40 and 80 µM. Following treatment for 24 h, the cells were
harvested and fixed in 500 µl 70% ice-cold ethanol at 4°C for 2 h.
Next, the samples were washed with phosphate-buffered saline (PBS),
and incubated with RNase A and PI staining solution, according to
the manufacturer's instructions. Subsequent to staining, the
samples were analyzed by performing flow cytometry (EPICS® XL™;
Beckman Coulter, Brea, CA, USA).
Flow cytometric apoptosis analysis
using Annexin V-FITC/PI staining
SK-MES-1 cells were seeded in a 6-well plate at a
density of 3×105 cells/well and treated with DP at
concentrations of 0, 40 and 80 µM. Following treatment for 24 h,
the cells were collected and washed with PBS. Subsequently, the
apoptotic cell death rate was examined by performing Annexin V-FITC
and PI double-staining using the Annexin V-FITC apoptosis detection
kit, according to the manufacturer's instructions. After staining
with Annexin V-FITC/PI, the samples were analyzed using flow
cytometry.
Nuclei fragmentation observed by
Hoechst 33342 staining
To visualize apoptotic cell death and nuclear
morphology, SK-MES-1 cells were stained with Hoechst 33342.
Briefly, SK-MES-1 cells were seeded into a 6-well flat bottom plate
at a density of 6×105 cells/well and treated with DP at
concentrations of 0, 40 and 80 µM. Following treatment for 24 h,
the cells were collected, washed and allowed to dry on slides.
Subsequently, the nuclei were stained with Hoechst 33342 for 10 min
and the apoptotic cells displaying fragmented or condensed nuclei
were observed under a fluorescence microscope (BH2-RFL-T3; Olympus
Corporation, Tokyo, Japan).
Western blot analysis
To determine the underlying mechanism of the
apoptotic effect of DP, western blot analysis was performed for a
number of apoptosis-associated proteins. First, SK-MES-1 cells were
seeded in a 10 cm culture dish at a density of 60%, treated with DP
at concentrations of 0, 40 and 80 µM. Following treatment for 24 h,
the cells were collected and washed with PBS. The cells were
centrifuged at 1,000 × g for 5 min; the cell pellets were then
resuspended in radioimmunoprecipitation assay lysis buffer and
lysed on ice using ultrasound (200 W for 15 sec). Following
centrifugation at 12,000 × g for 10 min, the supernatant fluids
were collected and the protein contents of the supernatant were
determined using a BCA Protein Assay kit, with the protein samples
stored at −20°C. Subsequently, the protein lysates were separated
by performing electrophoresis on a 10% sodium dodecyl
sulphate-polyacrylamide gel and transferred to a polyvinylidene
fluoride membrane (GE Healthcare, Piscataway, NJ, USA). The
membranes were then soaked in blocking buffer (5% skimmed milk) for
2 h. To probe for all the apoptosis-associated proteins, the
membranes were incubated overnight at 4°C with the relevant
antibodies, followed by appropriate HRP-conjugated secondary
antibodies, and enhanced chemiluminescence detection was performed.
Gel-Pro Analyzer software (GelPro32, version 4.0; Media
Cybernetics, Inc., Rockville, MD, USA) was used to extract valuable
qualitative and quantitative data from the electrophoretic gels to
document and store the western blot data.
Flow cytometric determination of the
MMP
Flow cytometry was performed to evaluate
perturbations in the MMP in SK-MES-1 cells treated with Rhodamine
123. Briefly, SK-MES-1 cells were seeded in a 6-well plate at a
density of 3×105 cells/well and treated with DP at
concentrations of 0, 40 and 80 µM. Following treatment for 24 h,
the cells were collected, washed with PBS and incubated with 10 µM
Rhodamine 123 at a temperature of 37°C for 20 min. The stained
cells were washed and resuspended in 200 µl PBS prior to
determining the MMP level using flow cytometry.
Flow cytometric measurement of
intracellular ROS generation
A 2′,7′-dichlorofluorescin-diacetate (DCFH-DA)
detection kit was used to measure the ROS levels, according to the
manufacturers instructions. Briefly, SK-MES-1 cells were treated
with DP at concentrations of 0, 40 and 80 µM. Following treatment
for 24 h, the cells were collected, washed in DMEM without FBS and
incubated with 10 mM DCFH-DA at a temperature of 37°C for 15 min.
The stained cells were washed and resuspended in 200 µl DMEM.
Intracellular ROS mediate the oxidation of DCFH to the fluorescent
compound DCF; thus, the generation of ROS was analyzed using flow
cytometry.
Statistical analysis
All the data are expressed as the mean ± standard
deviation of at least three independent experiments. For
statistical analysis, comparisons between results from different
groups were analyzed using SPSS software for Windows (version 17.0;
SPSS, Inc., Chicago, IL, USA). Comparisons were performed using
one-way analysis of variance, followed by Dunnett's test. P<0.05
was considered to indicate a statistically significant
difference.
Results
DP time- and dose-dependently inhibits
SK-MES-1 cell growth
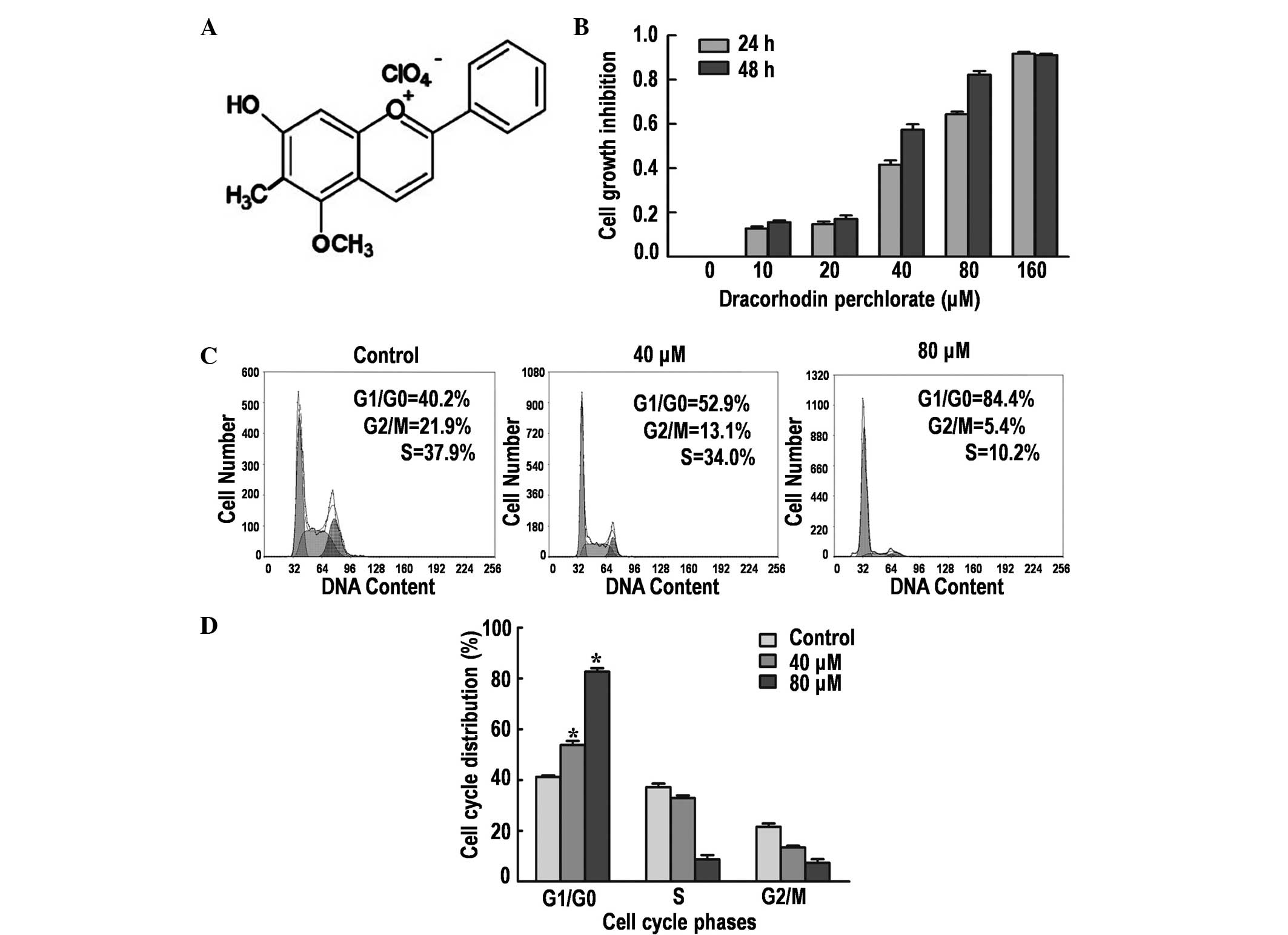
DP (Fig. 1A) is a
synthetic analogue of the antimicrobial anthocyanin red pigment
dracorhodin, a compound isolated from the exudates of the fruit of
Daemonorops draco (9). The
antiproliferative effect of DP on SK-MES-1 cells was determined by
performing an MTT assay. Treatment with DP for 24 and 48 h reduced
the cell viability in a time- and dose-dependent manner (Fig. 1B). The IC50 values were ~50
and ~30 µM following treatment for 24 and 48 h, respectively. Thus,
24-h treatments with 40 and 80 µM DP were selected for the
subsequent experiments.
DP induces G1/G0
phase arrest in SK-MES-1 cells
Cell cycle arrest is one of the major causes of cell
growth inhibition. Therefore, the induction of cell cycle arrest
was analyzed using PI staining and flow cytometry. The results
demonstrated that DP treatment caused significant cell cycle arrest
at the G1/G0 phase in a dose-dependent manner
(P<0.05; Fig. 1C and D). The
percentage of cells accumulated in the G1/G0
phase were 40.2, 52.9 and 84.4% following treatment with 0, 40 and
80 µM DP for 24 h, respectively. In addition, a corresponding
decrease in G2/M and S phase cells was observed, in part
caused by the induction of G1/G0 phase cell
cycle arrest.
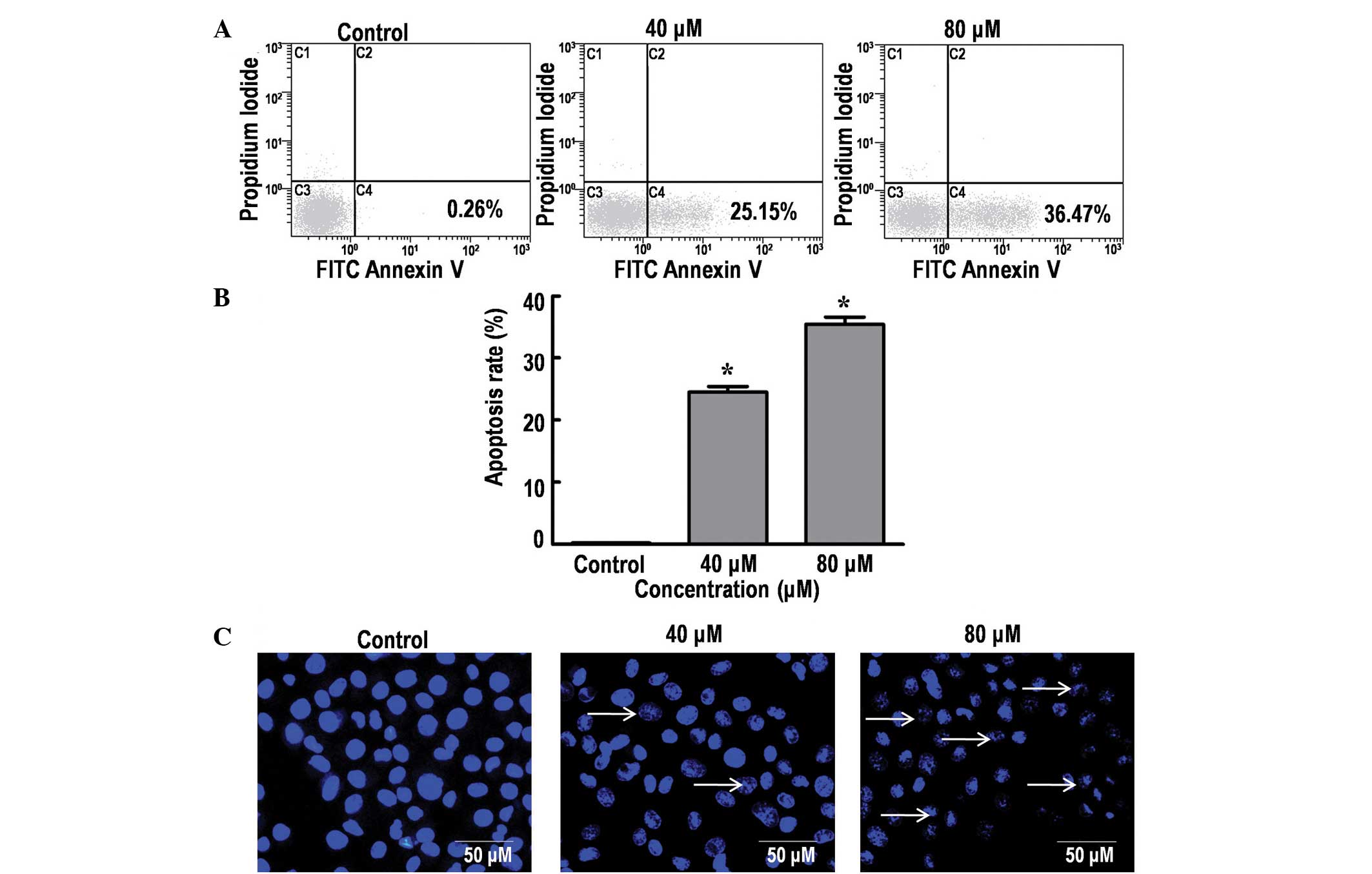
DP induces apoptosis in SK-MES-1
cells
The effect of DP on cell apoptosis was analyzed
using Annexin V-FITC/PI and Hoechst 33342 staining. The results
indicated a significant increase in the percentage of dead cells in
a dose-dependent manner, from 0.26% (control group; 0 µM DP) to
25.15 and 36.47% following treatment with 40 and 80 µM DP for 24 h,
respectively (P<0.05; Fig. 2A and
B). DNA fragmentation is an important characteristic of
apoptosis that can be clearly identified using Hoechst staining
(19). Consistent with the
aforementioned results, treatment of the SK-MES-1 cells with 40 and
80 µM DP for 24 h resulted in a marked increase in nuclear
fragmentation (Fig. 2C). Thus, the
current data demonstrated that DP can induce apoptosis in SK-MES-1
cells in a dose-dependent manner.
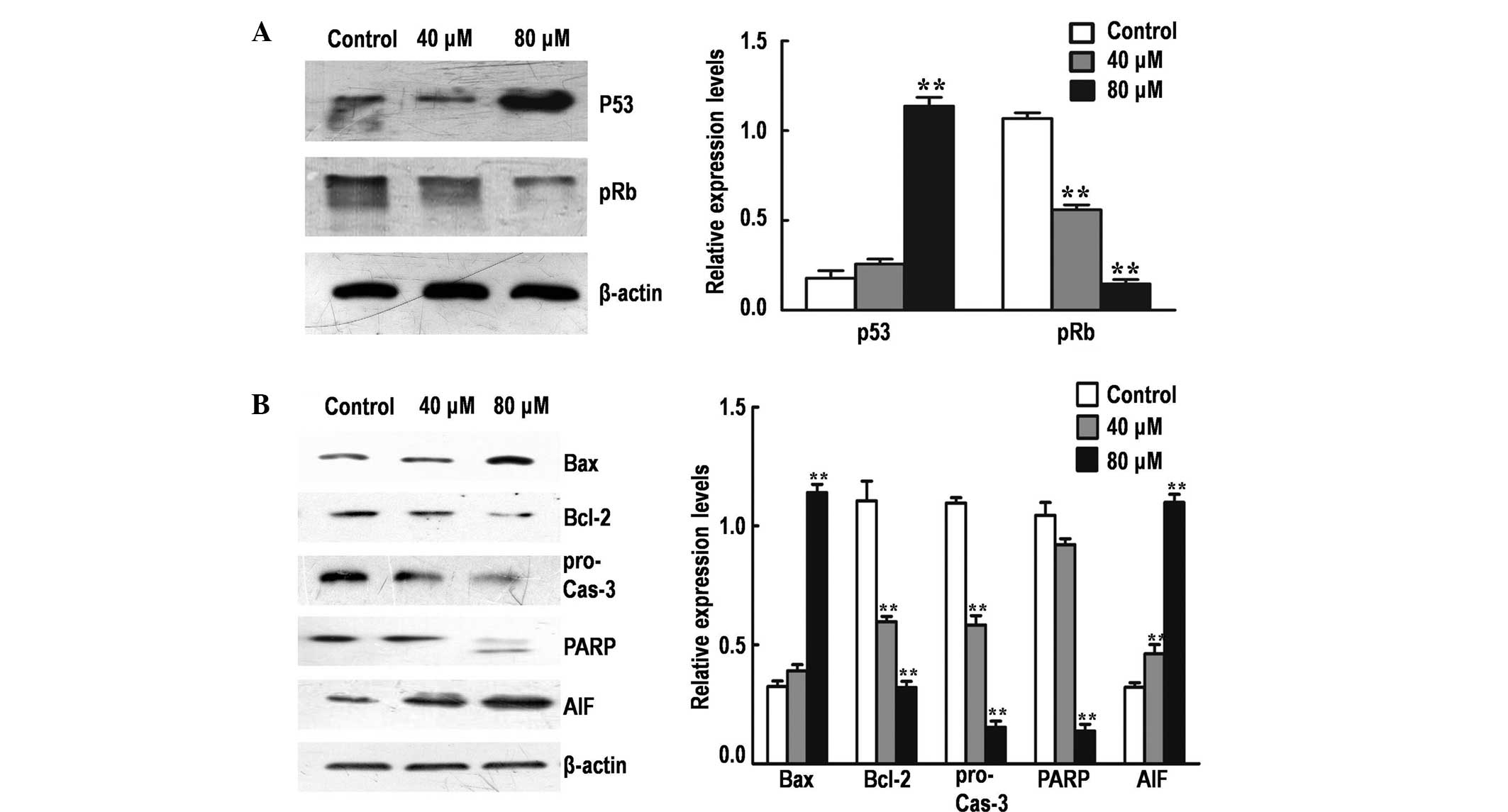
Effect of DP on the expression of
major cell cycle and mitochondrial apoptosis regulators
To elucidate the molecular mechanism underlying
G1/G0 phase arrest mediated by DP, the
protein expression levels of various major cell cycle regulatory
proteins (p53 and pRb) were detected by performing western blot
analysis. The treatment of SK-MES-1 cells with DP for 24 h resulted
in the significant upregulation of p53 and the significant
downregulation of pRb in a dose-dependent manner (P<0.05;
Fig. 3A).
To investigate the rate of mitochondrial apoptosis
in SK-MES-1 cells, western blot analysis was performed to determine
the effect of DP treatment on the protein expression levels of
various major mitochondrial apoptosis regulatory proteins (Bax,
Bcl-2, caspase-3, PARP and AIF; Fig.
3B). The Bax/Bcl-2 expression ratio was significantly increased
following treatment with DP (P<0.05), accompanied by activation
of procaspase-3 and cleavage of PARP in a dose-dependent manner. In
addition, the expression of AIF was significantly increased
following treatment (P<0.05), possibly due to the increase in
mitochondrial permeability, resulting in the release of AIF from
the mitochondria into the cytosol. These results indicate that DP
can induce apoptosis in SK-MES-1 cells via the mitochondrial
pathway.
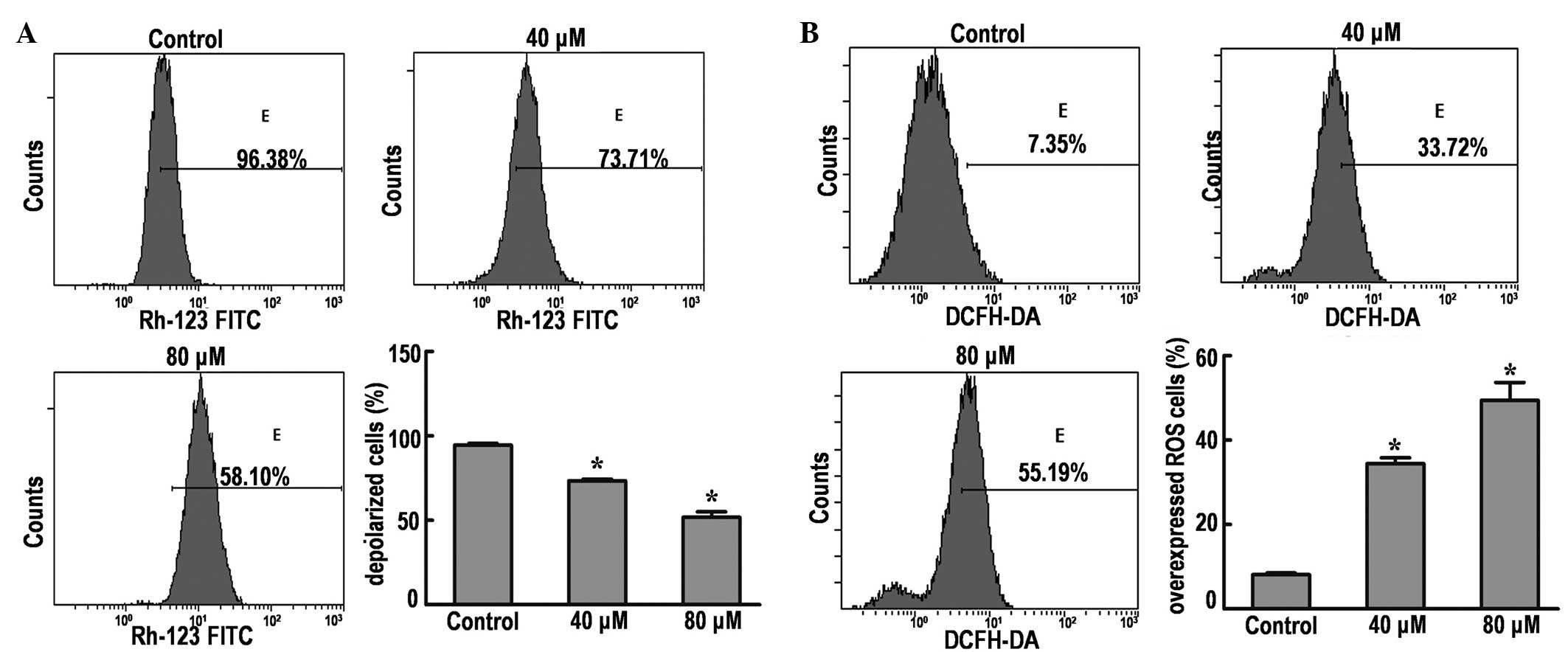
DP causes disruption of the MMP in
SK-MES-1 cells
Depolarization of the MMP is a characteristic
feature of apoptosis; therefore, the effects of DP on the MMP of
SK-MES-1 cells were examined by flow cytometry using Rhodamine 123
staining. The results demonstrated that the MMP was significantly
decreased from 96.38% (control group) to 73.71 and 58.10% in cells
treated with 40 and 80 µM DP, respectively (P<0.05; Fig. 4A). These results indicate that
dissipation of the MMP may be one of mechanisms through which DP
induces apoptosis.
DP induces increased generation of ROS
in SK-MES-1 cells
Intracellular ROS generation in SK-MES-1 cells was
evaluated by flow cytometry using DCFH-DA (Fig. 4B). The ROS level in SK-MES-1 cells
treated with 40 and 80 µM DP was significantly increased from 7.35%
(control group) to 33.72 and 55.19%, respectively.
Discussion
A number of studies have recently identified that DP
has a broad spectrum of cytotoxicity towards various human cancer
cell lines of different origins (11–13). In
the present study, DP was demonstrated to significantly inhibit the
growth of SK-MES-1 human lung cancer cells in a dose-dependent
manner.
Cell cycle regulation and apoptosis are considered
to be major causes of cell growth inhibition (20). The cell cycle is controlled at
different checkpoints. These checkpoints ensure that specific
processes have been precisely completed at each stage of the cell
cycle before allowing progress to the next phase of the cycle. The
loss of key checkpoints is a characteristic of cancer cells that
results in anomalous proliferation and the promotion of oncogenic
transformation (21). The results of
the current study indicated that the treatment of lung cancer cells
with DP induced G1/G0 phase arrest in a
dose-dependent manner.
In addition to cell cycle arrest, DP exhibited a
cytotoxic effect in the present study, inducing apoptotic cell
death in lung cancer cells. These apoptotic effects are consistent
with the results of a number of previous studies, which observed
that DP inhibited abnormal proliferation by the induction of
apoptosis in various types of cancer cells, including melanoma,
gastric cancer and prostate cancer cells (11,12,22).
p53 is an important factor in the regulation of cell
cycle progression, checkpoint activation, apoptosis and repair of
DNA damage (15,23,24). Once
activated, p53 can activate its downstream transcription factor,
which can form complexes with cell cycle-dependent protein kinase
and inhibit the activity of Rb protein. Rb is critical in the
G1/S phase transition, regulating the expression of
genes necessary for cell cycle progression (25). The present study determined that
treatment with DP significantly upregulated the p53 protein
expression levels. This upregulation was accompanied by a
significant decrease in the protein expression levels of pRb. These
data indicated that DP may disrupt cell cycle progression via the
activation of p53 and the inhibition of Rb.
The p53 tumor suppressor is crucial for regulating
the expression of various genes that mediate apoptosis.
Furthermore, it is well-established that p53 activates apoptosis
via the regulation of mitochondrial integrity, resulting in the
release of downstream cytokines and, ultimately, the activation of
caspases (15,23,24). The
Bcl-2 protein family is a well-known family of apoptosis regulating
proteins that act via the mitochondrial pathway. This family
includes anti-apoptotic proteins, such as Bcl-2, and proapoptotic
proteins, such as Bax (17). These
proteins function together to regulate mitochondrial membrane
permeability and modulate the release of apoptogenic proteins that
promote cell death (26–28). In the present study, the expression
levels of proteins involved in the mitochondrial pathway were
detected by western blot analysis. The current data demonstrated
that the expression of Bax significantly increase, while the
expression of Bcl-2 significantly decreased, indicating that DP may
induce apoptosis via the mitochondrial pathway.
An increase in mitochondrial permeability can result
in the release of proapoptotic molecules, leading to the activation
of other downstream caspases and, ultimately, the activation of
caspase-3 (29–31). Caspases play a key role in the process
of apoptosis and frequently catalyze the specific cleavage of
numerous pivotal cellular proteins (18). For instance, the cleavage of PARP,
which is a DNA repair enzyme, is performed by caspases, in
particular caspase-3, and is the hallmark of apoptosis (18,32). The
data presented in the current study clearly demonstrated the
cleavage of PARP following treatment with DP. These results
indicate that the intrinsic mitochondrial-mediated caspase
activation pathway is involved in the DP-mediated apoptosis of
SK-MES-1 cells.
Dysfunction of the mitochondrial membrane allows the
release of AIF from mitochondria into the cytosol. AIF is a
mitochondrial intermembrane flavoprotein that can induce apoptosis
in a caspase-independent manner. AIF condenses chromatin and
fragmented DNA in order to trigger programmed cell death (33,34).
Previous studies have indicated that AIF is required for cell death
following certain cell stresses (33,34). This
has been clarified by the injection of anti-AIF antibodies or
knockout of the AIF gene, which alleviated the progression
of apoptosis (33,34). The current study demonstrated that
treatment with DP significantly increased the expression of AIF,
indicating that DP may partially induce the apoptosis of SK-MES-1
cells via a caspase-independent pathway.
Increasing the expression of Bax appears to result
in increased mitochondrial membrane permeability and dissipation of
the MMP (35). Flow cytometry data
obtained in the present study supports this hypothesis, with a
significant reduction in MMP observed in the treatment group cells.
Thus, the results of the current study indicate that DP treatment
may disrupt the integrity of mitochondria by increasing the
Bax/Bcl-2 ratio.
A number of studies have indicated that ROS are
downstream mediators of p53-dependent apoptosis. However, ROS may
transmit a signal for apoptosis as opposed to being a consequence
of the cellular changes accompanied by apoptosis (36), as p53 affects the mitochondrial
apoptotic pathway and mitochondria are the major target of ROS. In
addition, ROS generation may alter the redox status of cells, thus,
altering the sensitivity of cells to apoptotic stimuli and
ultimately triggering subsequent apoptotic events (37–40). In
the current study, DP treatment significantly increased the
generation of ROS in SK-MES-1 cells. Our future studies will aim to
summarize the association between the current findings, and the
cell cycle and apoptosis. Understanding the specific mechanism of
ROS generation may provide a novel method for the development of
therapeutic agents that are capable of selectively inducing
apoptosis in healthy or neoplastic cells.
In conclusion, the current study revealed that
treatment of SK-MES-1 cells with DP induced: Mitotic arrest;
caspase-dependent apoptosis via an increase in the Bax/Bcl-2 ratio
and caspase-3 expression, causing cleavage of PARP; and
caspase-independent apoptosis via an increase in AIF expression.
Therefore, DP may be a potential compound for the development of
future lung cancer therapeutic strategies.
Acknowledgements
The present study was supported by grants from the
Science and Technology Services of Jilin Province Scientific and
Technological Project (no. 20140521) and the Natural Science
Foundation of China (no. 81272472).
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Johnson DH: Evolution of cisplatin-based
chemotherapy in non-small cell lung cancer: A historical
perspective and the eastern cooperative oncology group experience.
Chest. 117 (4 Suppl 1):133S–137S. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cragg GM and Newman DJ: Plants as a source
of anti-cancer agents. J Ethnopharmacol. 100:72–79. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Amin AR, Kucuk O, Khuri FR and Shin DM:
Perspectives for cancer prevention with natural compounds. J Clin
Oncol. 27:2712–2725. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Russo M, Palumbo R, Mupo A, et al:
Flavonoid quercetin sensitizes a CD95-resistant cell line to
apoptosis by activating protein kinase Calpha. Oncogene.
22:3330–3342. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xia MY, Wang MW, Cui Z, et al: Dracorhodin
perchlorate induces apoptosis in HL-60 cells. J Asian Nat Prod Res.
8:335–343. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ferreira CG, Epping M, Kruyt FA and
Giaccone G: Apoptosis: Target of cancer therapy. Clin Cancer Res.
8:2024–2034. 2002.PubMed/NCBI
|
|
8
|
Brockmann H and Junge H: Die konstitution
des dracorhodins, eines neuen farbstoffes aus dem ‘drachenblut’.
Eur J Inorg Chem. 76:751–763. 1943.(In German).
|
|
9
|
Rao GS, Gerhart MA, Lee RT III, et al:
Antimicrobial agents from higher plants. Dragon's blood resin. J
Nat. Prod. 45:646–648. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gao WF, Zheng H, Wang YS, et al: Synthesis
of dracorhodin. Chin J Pharma. 20:247–250. 1989.(In Chinese).
|
|
11
|
Rasul A, Ding C, Li X, et al: Dracorhodin
perchlorate inhibits PI3K/Akt and NF-κB activation, up-regulates
the expression of p53, and enhances apoptosis. Apoptosis.
17:1104–1119. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xia M, Wang M, Tashiro S, Onodera S,
Minami M and Ikejima T: Dracorhodin perchlorate induces A375-S2
cell apoptosis via accumulation of p53 and activation of caspases.
Biol Pharm Bull. 28:226–232. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yu JH, Zheng GB, Liu CY, Zhang LY, Gao HM,
Zhang YH, Dai CY, Huang L, Meng XY, Zhang WY and Yu XF: Dracorhodin
perchlorate induced human breast cancer MCF-7 apoptosis through
mitochondrial pathways. Int J Med Sci. 10:1149–1156. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xia M, Wang D, Wang M, Tashiro S, Onodera
S, Minami M and Ikejima T: Dracorhodin perchlorate induces
apoptosis via activation of caspases and generation of reactive
oxygen species. J Pharmacol Sci. 95:273–283. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fridman JS and Lowe SW: Control of
apoptosis by p53. Oncogene. 22:9030–9040. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Adams JM and Cory S: Life-or-death
decisions by the Bcl-2 protein family. Trends Biochem Sci.
26:61–66. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Saelens X, Festjens N, Vande Walle L, van
Gurp M, van Loo G and Vandenabeele P: Toxic proteins released from
mitochondria in cell death. Oncogene. 23:2861–2874. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Porter AG and Jänicke RU: Emerging roles
of caspase-3 in apoptosis. Cell Death Differ. 6:99–104. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Araki M, Iida Y, Taketani S, Watanabe K,
Ohta T and Saito T: Characterization of photoreceptor cell
differentiation in the rat retinal cell culture. Dev Biol.
124:239–247. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
King KL and Cidlowski JA: Cell cycle
regulation and apoptosis. Annu Rev Physiol. 60:601–617. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pan J, She M, Xu ZX, et al:
Farnesyltransferase inhibitors induce DNA damage via reactive
oxygen species in human cancer cells. Cancer Res. 65:3671–3681.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
He Y, Ju W, Hao H, Liu Q, Lv L and Zeng F:
Dracorhodin perchlorate suppresses proliferation and induces
apoptosis in human prostate cancer cell line PC-3. J Huazhong Univ
Sci Technolog Med Sci. 31:215–219. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Budram-Mahadeo V, Morris PJ and Latchman
DS: The Brn-3a transcription factor inhibits the pro-apoptotic
effect of p53 and enhances cell cycle arrest by differentially
regulating the activity of the p53 target genes encoding Bax and
p21(CIP1/Waf1). Oncogene. 21:6123–6131. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Vogelstein B, Lane D and Levine AJ:
Surfing the p53 network. Nature. 408:307–310. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Das SK, Hashimoto T, Shimizu K, et al:
Fucoxanthin induces cell cycle arrest at
G0/G1 phase in human colon carcinoma cells
through up-regulation of p21WAF1/Cip1. Biochim Biophys Acta.
1726:328–335. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huang DC and Strasser A: BH3-Only
proteins-essential initiators of apoptotic cell death. Cell.
103:839–842. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Frenzel A, Grespi F, Chmelewskij W and
Villunger A: Bcl2 family proteins in carcinogenesis and the
treatment of cancer. Apoptosis. 14:584–596. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Danial NN: BCL-2 family proteins: critical
checkpoints of apoptotic cell death. Clin Cancer Res. 13:7254–7263.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Schuler M, Bossy-Wetzel E, Goldstein JC,
et al: p53 induces apoptosis by caspase activation through
mitochondrial cytochrome c release. J Biol Chem. 275:7337–7342.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Toshiyuki M and Reed JC: Tumor suppressor
p53 is a direct transcriptional activator of the human bax gene.
Cell. 80:293–299. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Greiner M, Cárdenas S, Parra C, et al:
Adrenalectomy regulates apoptotic-associated genes in rat
hippocampus. Endocrine. 15:323–333. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Slee EA, Adrain C and Martin SJ:
Executioner caspase −3, −6, and −7 perform distinct, non-redundant
roles during the demolition phase of apoptosis. J Biol Chem.
276:7320–7326. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Norberg E, Orrenius S and Zhivotovsky B:
Mitochondrial regulation of cell death: processing of
apoptosis-inducing factor (AIF). Biochem Biophys Res Commun.
396:95–100. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Candé C, Cohen I, Daugas E, et al:
Apoptosis-inducing factor (AIF): a novel caspase-independent death
effector released from mitochondria. Biochimie. 84:215–22. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kroemer G: The proto-oncogene Bcl-2 and
its role in regulating apoptosis. Nat Med. 3:614–620. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Johnson TM, Yu ZX, Ferrans VJ, et al:
Reactive oxygen species are downstream mediators of p53-dependent
apoptosis. Proc Natl Acad Sci USA. 93:11848–11852. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cai J and Jones DP: Mitochondrial redox
signaling during apoptosis. J Bioenerg Biomembr. 31:327–334. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Simon HU, Haj-Yehia A and Levi-Schaffer F:
Role of reactive oxygen species (ROS) in apoptosis induction.
Apoptosis. 5:415–418. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Akgul C, Moulding DA and Edwards SW:
Molecular control of neutrophil apoptosis. FEBS Lett. 487:318–322.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cimino F, Esposito F, Ammendola R and
Russo T: Gene regulation by reactive oxygen species. Curr Top Cell
Regul. 35:123–148. 1997.PubMed/NCBI
|