Introduction
Cervical cancer accounts for 15% of cancers in
females aged <65 years old in developing countries (1). Long-term high-risk human papillomavirus
(hrHPV) infection must be present, but is insufficient to cause
cervical cancer by itself according to the International Agency for
Research on Cancer (IARC). Certain other risk factors account for
the development of cervical cancer, including an active sexual
history, weakened immune system function and smoking (2). The induction of cervical carcinogenesis
by persistent hrHPV infection is a multistep process, consisting of
persistent infection, different stages of cervical intraepithelial
neoplasia (CIN) lesions and ultimately, cervical cancer (3). Although persistent hrHPV infection is an
important prerequisite for CIN, the vast majority (90%) of viruses
have been cleared by the hosts' immune system, without medical
intervention, as observed on the three-year follow-up examination,
and only 1–2% of the remaining 10% become chronic resulting in
cervical cancer (3).
To date, >180 types of HPV are known to exist
(4). HPV16, 18, 31, 33, 35, 39, 45,
51, 52, 56, 58, 59, 68 and 69 are classified as the hrHPVs
(5). HPV6, 11, 40, 42, 54, 55, 61,
62, 64, 71, 72, 81, 83 and 84 are classified as the low-risk HPVs
(6). HPV16 and 18 are the most common
types, accounting for ~70% of cervical cancers around the world
(7).
HrHPV infection starts with the contact of the virus
with the basement membrane, which is often exposed by
micro-abrasions on the cervical surface. Suitable receptors
increase the probability of hrHPV infection. Studies have found
several receptors involved in this process. Interaction between the
virus' capsid protein and the cell receptors promotes virus capsid
conformational changes, thus aiding the cell entry process
(8). Tissue-specific heparin sulfate
proteoglycan (HSPG), as one of the members of the glycosaminoglycan
family, is a receptor of major capsid protein L1 of HPV (9,10). HSPG
could aid HPV16 in binding to the extracellular matrix via
laminin-332 (11).
HPV DNA is double-stranded, containing 7,900 base
pairs, which are arranged in a circle (12). The genome consists of 8 open reading
frames, 6 early genes (E1, E2, E4, E5, E6 and E7) encoding the
early protein and 2 late genes (L1 and L2) encoding the late
protein. E1 protein aids in viral replication utilizing the host
replication machinery. E5, E6 and E7 proteins are considered to be
associated with virus immune evasion. The product of E5 could
downregulate the histocompatibility leucocyte antigen (HLA)
expression of the infected cells, facilitating virus immune escape
(13,14). E6 and E7 are considered as tumorigenic
genes; their products could bind to tumor suppressor protein p53
and arrest cell cycle, impairing the infected cells apoptosis and
enhancing their transformation (15).
The L1 and L2 proteins are responsible for forming the structural
components of the viral capsid. E2 exhibits a regulatory function
for E6 and E7 transcription. HPV genome integration into the host
genome would disrupt the regulatory function of E2, leading to the
uncontrolled expression of E6 and E7, promoting the progression of
the disruption of the normal cell cycle and carcinogenesis
(16).
Furthermore, the expression of E6 and E7 is
fundamental in hrHPV infection and CIN progression. The expression
of these oncoproteins inhibits the immune system response to hrHPV
and aids in the persistency of the infection. The oncoproteins
could downregulate TLR9 expression, a virus DNA sensor, which is
necessary to activate antigen-presenting cells (17), but they could also reduce the
expression of transporter-associated antigen processing 1, blocking
the activation of specific T lymphocytes. It has been confirmed
that neoplastic cervical keratinocytes (KCs) expressing high levels
of E6 and E7 oncoproteins could escape the attack from cytotoxic T
cells (CTLs) (18).
HPV infection regulating the host immune
response
According to a previous study, the incubation period
of HPV infection is ~10 years. The immune response of the host
plays a crucial role in the progression or regression of hrHPV
infection of the uterine cervix. An effective immune response
promotes spontaneous clearance of the virus, while a compromised
immune response often starts up the pathological process,
developing it into the higher grade (19). HrHPV infection causes several events,
which are essential for CIN progression and carcinogenesis
(Fig. 1).
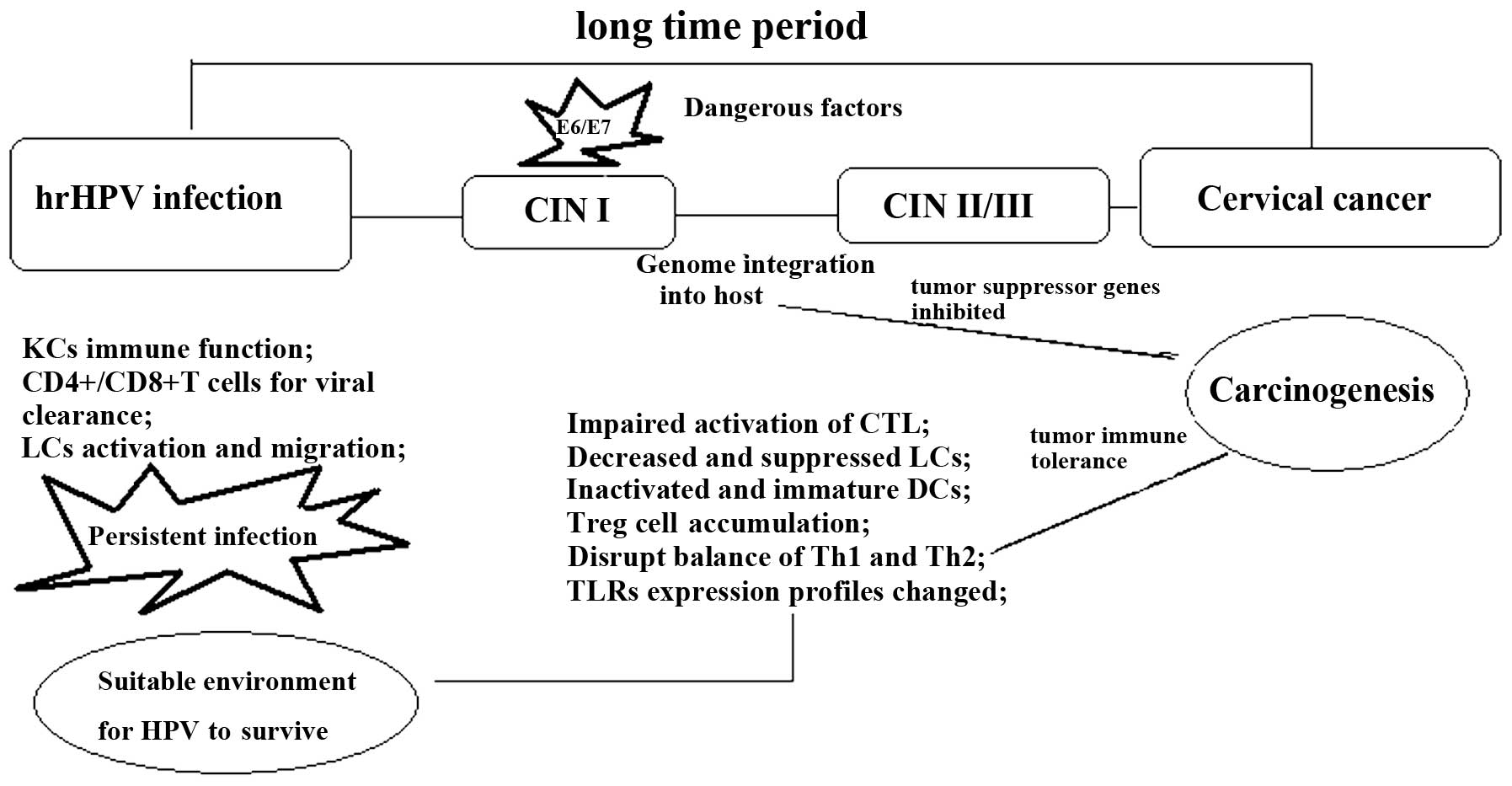 | Figure 1.There are several stages from HPV
infection to cervical cancer. Immune system modification is the
most important factor to promote lesion progression. Genome
integration would initiate immune cell activation and
differentiation profiles changes. These changes are characterized
by the inactivated CD4+/CD8+ T cells, Treg
cell upregulation, M2 cell generation, the immature DCs and reduced
anti-inflammatory cytokine infiltration. hrHPV, high-risk human
papilloma virus; CIN, cervical intraepithelial neoplasia; CD,
cluster of differentiation; LC, Langerhans cell; CTL, cytotoxic T
cell; DC, dendritic cell; Th, T-helper cell; TLR, toll-like
receptor. |
HrHPV infection promotes immune cell migration to
the dermis. In the squamous epidermis, macrophages, Langerhans
cells (LC), KCs, T lymphocytes, dendritic cells (DC), natural
killer cells (NK) and B lymphocytes play important roles during the
immune response to infection. HrHPV infection could cause the
immune system to become more tolerant to the infection, thus
creating a microenvironment susceptible to further infection and
facilitating CIN progression. The mechanisms that have been
proposed and proved are as follows: Firstly, hrHPV remains silent
for a long time; its duplication and assembly do not cause
cytolysis or the cytopathic death of the host cells (20). Secondly, hrHPV inhibits interferon
(IFN) synthesis through E6 and E7 oncoproteins interfering with IFN
signaling pathways (21). Thirdly,
hrHPV infection induces regulatory T cell (Treg) infiltration and
interleukin (IL)-10 or transforming growth factor β (TGF-β)
production. Fourthly, the infected cells express low levels of MHC
class I, resulting in impaired CTL function (22). Fifthly, they could induce an
accumulation of ineffective CD4 and CD8 T lymphocytes in stage
II/III CINs (23).
Additionally, KCs are the major cell type in the
epidermis, with certain immune functions, such as cytokine
secretion. The compromised innate immune defense in KCs is an
important reason for the evasion of the hrHPV infection from the
immune response, ultimately resulting in persistent virus infection
and the development into pre-neoplastic lesions (24).
Innate immunity affected by hrHPV
infection
Usually, once the hrHPV contacts the mucosal
epithelium, the innate immunity system mediated by the epithelial
barrier starts to conflict with it. LCs are immature DCs, and in
the transformation zone their numbers are significantly decreased.
The mechanism proposed for this is the direct interaction of E7
with CCAAT/enhancer-binding protein β, a transcription factor of
chemokine (C-C motif) ligand 20 (CCL20) (which has a decisive role
in the migration of LC precursors into the epidermis), thereby
inhibiting the transcription of CCL20 and thus hindering LC
recruitment (25).
Toll-like receptors (TLRs) are an important type of
pattern recognition receptor located at the endolysosomal
compartments, sensing the bacteria/virus and triggering the
associated innate immune response. Several of these receptors have
been investigated. HrHPV18 E6 and E7 downregulate TLR9 (specific
for the nucleic acid) expression at the infection site, an
important strategy for its escaping from immunosurveillance
(26). In contrast to TLR9, the
TLR3/5/8 pathways are activated in hrHPV-infected KCs. High
expression levels of TLR8 in cervical cancer cells are associated
with the upregulated expression of B-cell lymphoma-2 and vascular
endothelial growth factor in these cells (27). A previous study found that there is
marked TLR4 expression in hrHPV-positive cells and that its
expression is associated with the virus type, as well as the
histopathological grade. Higher TLR4 expression is found in
HPV16+ cells compared with HPV18+ cells, and
in cervical cancer compared with CIN. Additionally, it has been
demonstrated that TLR4 overexpression is correlated with the
apoptotic resistance of the HPV-infected cells (28).
It is known that tumor-associated macrophages, also
termed M2 macrophages, can promote cancer cell proliferation and
migration, angiogenesis and the restriction of immune defenses
(29). During cervical lesion
progression, the number of M2 macrophages is significantly
increased (30). The aggregation of
M2 is a key event for the pathological process of
carcinogenesis.
HrHPV infection compromises NK cell activation. NK
cells predominate at the initial stage of the infection and in the
low-grade lesions. Levels of NK-activating receptors, such as
NKp30, NKp45, NKp46, NKG2D and NKp80, are significantly decreased
in HPV16 cervical cancer. These receptors are closely associated
with the low cytotoxic activity of NK cells, facilitating lesion
progression and carcinogenesis (31).
hrHPV-infected tissues or cervical cancer cells could obtain their
immune evasion abilities by making NK cells more tolerant to the
hrHPV infection and cervical carcinogenesis (32).
Immune cell changes in the immune response
to hrHPV
Compromised adaptive immunity is the foundation for
the progression from hrHPV infection to cervical cancer. There are
different immune cell profiles for the different stages of the
disease progression in CIN and carcinogenesis. The changes and
modifications induced by hrHPV infection are involved in hrHPV
infection adapting the immune system to create a suitable
microenvironment for persistent infection and lesion
progression.
HPV infection induces compromised T
cell activation
T cell activation is important in hrHPV infection.
It has been proven that in patients with HIV infection, compromised
CD4+ T cells function as the inducer for HPV-associated
cancer occurrence (20). For these
patients, the induction of inefficient CD4+ T cells by
hrHPV infection is the factor promoting CIN lesion progression
(33). CTL is the main agent in
cancer specific immunity, and it recognizes the antigens with the
assistance of MHC/HLA class I. HPV16 E5 could suppress CTL through
the downregulation of MHC/HLA class I expression, as
aforementioned.
Distorted equilibrium between type 1 T-helper cells
(Th1) and Th2 cell is another property of cellular immunity during
hrHPV infection (34). There is a
marked Th2 cytokine profile in intraepithelial and invasive
cervical lesions. Increased Th2 cytokine (IL-10) and reduced Th1
cytokine (IFN-γ, IL-12, IL-2 and tumor necrosis factor-α) levels
have been detected in cervical exudates of hrHPV+
patients. This indicated that the reduced Th1 response and
increased Th2 response lead to cellular immunity suppression and
cervical lesion progression (35,36).
Another study proved that the production of certain Th2 cytokines
is also decreased during this process, indicating that the Th1 and
Th2 phenotypes, but particularly the Th1 phenotype, may be
suppressed by hrHPV infection. One of the mechanisms by which hrHPV
regulates T cell activation is based on E6 and E7 oncoprotein
expression, which would upregulate the expression of adhesion
molecules, such as intercellular adhesion molecule 1, vascular cell
adhesion molecule 1 and E-selectin (classical immunosuppressive
molecules), in the infected cells (37).
Overall, compromised T cell activation and the
distorted equilibrium between Th1 and Th2 presents hrHPV with the
ability to escape from the human immune defense.
Compromised DC activation induced by
hrHPV infection
DCs, as one of the important adaptive immune cells,
recognize the special patterns of pathogens through TLRs and
present the antigens to T cells through MHC molecules to initiate
the antigen-specific immune response. A high level of stromal DCs
is usually associated with a higher frequency of infection
regression. Along with hrHPV infection, immature DCs usually
prevent the activation of a correct immune response by CTLs and
facilitate lesion progression (38).
There is an extremely low distribution or complete absence of
functional DCs in patients with cervical cancer. HrHPV
E6-expressing cells or hrHPV+ cancer cells can inhibit
the differentiation of monocytes into fully functional DCs
(39). A low percentage of
plasmacytoid DCs in the peripheral blood is significantly
associated with persistent hrHPV infection (40). Furthermore, it has been demonstrated
that in hrHPV+ patients, decreased expression levels of
CD80 and CD86 in DCs have a positive correlation with increased CIN
grades (41).
One of the mechanisms involved in the induction of
compromised DC activation in hrHPV infection was proposed as the
stimulation of programmed death 1 (PD-1)/PD-1 ligand (PD-L1)
(CD279/CD274) pathway activation by chronic hrHPV infection in DCs.
PD-1/PD-L1 activation is associated with impaired DC-mediated
immunity to hrHPV infection, thus creating the immune tolerance
microenvironment for persistent infection and CIN progression
(41).
Interplay between Treg cells and hrHPV
infection
Tregs
(CD4+/CD25+/Foxp3+ T cells) are
the inducer of immune tolerance. There is increased
Foxp3+ Treg infiltration following hrHPV infection.
Recent studies have highlighted the role of hrHPV-specific Treg
generation induced by hrHPV infection and CIN progression (42). Tregs specific for the E6 and E7
antigens have been detected in high-grade squamous intraepithelial
lesion (HSIL)-infiltrating lymphocytes (43), and there are several mechanisms that
contribute to their generation and recruitment. Firstly, improper
activation of the immune response induced by hrHPV infection
provides the possibility of the toleration of T cell generation.
Secondly, the weakened innate immune function elicited by the
infected KCs creates an immunosuppressive microenvironment,
promoting HPV-specific Treg expansion (22).
The regulation of hrHPV infection and CIN
progression by Treg cells is mainly based on cytokine production.
For example, TGF-β is produced by Treg, and its level has a
positive correlation with lesion progression (36). IL-10 is another significant
Treg-producing cytokine whose presence would decrease
CD8+ T cell infiltration and increase the amount of
intratumoral Foxp3+ Treg cells, a critical event for CIN
lesion progression (44,45). There is also a notable interaction
between IL-10 and HPV-induced lesion progression (46). In hrHPV infected cells, E2 protein
binds to the regulatory region of the human IL-10 gene (−2054 nt)
and induces its expression; IL-10 can create an immune-tolerant
microenvironment, providing the possibility of progressive cervical
disease in turn (47).
Immune-related markers of CIN and cervical
cancer
Immune markers (immune cells, cytokines, chemokines
and soluble receptors) during persistent hrHPV infection and CIN
progression have not been well studied (48). The percentage of Tregs is not only
used as a marker of tumor immune status, but is also considered as
a possible marker of immune destruction induced by hrHPV infection.
Foxp3, as the only definitive marker of Tregs, exhibits gradually
increased expression from CIN I to CIN III, which indicates that
Foxp3 is involved in the development of cervical cancer (49). However, the exact mechanism of such
requires further investigation.
High mobility group box 1 protein (HMGB1), a
DNA-binding protein, is critical for host immune suppression. There
is a direct correlation between HMGB1 expression and the malignant
potential of CIN. A previous study also proved that high levels of
HMGB1 expression can be used as an early prognostic marker in
recurrent cervical cancer patients (50). Patients with greater HMGB1 expression
have a higher rate of hrHPV infection recurrence than those with
weak HMGB1 expression (51).
Mechanisms employed by HMGB1 to induce the immune tolerance to
hrHPV infection are proposed to be as follows: HMGB1 suppresses the
human immune function by upregulating Tregs and promoting IL-10
production. It also inhibits the function of T cells by
downregulating NF-κB signaling and polarizing Th1 cells to Th2
cells. HMGB1 is becoming a useful biomarker for the evaluation of
hrHPV infection persistency and CIN progression (52).
p16INK4a expression is induced by the
integration of hrHPV DNA into the host genome; it is the
cyclin-dependent kinase inhibitor and is overexpressed in CIN
(53). p16INK4a expression
has a positive correlation with the CIN grade, and could be used to
discriminate between different CIN grades in cervical biopsies.
Generally, hrHPV L1 capsid protein is expressed in
every stage of CIN, with a negative association with the CIN grades
(54). The abortion of hrHPV cycles
in HSIL results in the L1 capsid protein being non-detectable
(55). From the immunological
perspective, hrHPV L1 is the inducer for the CD8+ and
CD4+ T cell response against hrHPV infection (56). Therefore, reduced L1 capsid expression
is a significant mechanism for the immune evasion of higher grade
lesions.
Preventive and therapeutic HPV vaccines
The majority of clinical trials concerning
HPV-targeted therapies are based on eliciting cell-mediated immune
responses with vaccines. Finding a target to generate a specific
immune response is the strategy for the treatment of hrHPV
infection, as well as cervical cancer (57). The preventive vaccination against
HPV16 and 18 has now become widely used (58). Antigen-specific immunotherapy is one
of the effective methods to elicit immune responses to hrHPV.
Listeria monocytogenes (LM) has been used to eliminate
palpable, vascularized tumors in several mouse models due to its
ability to generate CD8+ tumor-infiltrating lymphocytes.
Two vaccines, LM-LLO-E7 and LM-ActA-E7, have been created by a
truncated listeriolysin O (LLO) fused to E7 and a fragment of the
ActA protein fused to E7, respectively. These vaccines overcome
central tolerance by expanding low avidity CD8+ T cells
specific for E7 (59). A novel fusion
protein, HPV16 E7-HBcAg-Hsp65 (VR111), which could elicit an
E7-specific CD8+ T cell response, is a novel potential
preventive vaccine (60).
Therapeutic vaccines aim to clear hrHPV infections
and the hrHPV-related cervical lesions, and are mainly targeted to
oncoproteins E6 and E7, the only viral proteins expressed in
cervical cancer and precursor lesions. Previously, E6/E7
oncoproteins were used as the target of the therapeutic vaccine,
which could increase the lesion-infiltrating CD4+ and
CD8+ T cells in the intraepithelial neoplasia lesions
(61). In a study, a novel HPV16 E6
and E7 gene plasmid containing oligomannose liposomes (OML-HPV) was
been generated for the immunotherapy of cervical cancer. With the
stimulation of OML-HPV, HPV16 E6-specific CTLs could be generated
from peripheral blood mononuclear cells for HPV16+
cervical carcinoma patients (62).
During the analysis of the HPV vaccines, it has been
found that the DC-based HPV vaccine is a promising tool to prevent
and treat hrHPV infection, as well as cervical cancer. DCs
generated from peripheral blood monocytes with IL-4 and
granulocyte-macrophage colony-stimulating factor were treated with
HPV16 mE7, which showed the significantly increased expression of
co-stimulatory molecules CD80 and CD40, and the marked production
of IL-12p70 and IFN-γ. This subset of DCs could upregulate
E7-specific CD8+ T cell responses in patients with
hrHPV-associated cervical cancer (63). HPV16 E7 polypeptide is another
DC-based HPV vaccine. DC loaded with hrHPV16 E7 polypeptide in
combination with CpG-ODN2006 also showed distinguished
immunotherapeutic activity. When SCID mice were inoculated with
such antigen-loaded DCs, they exhibit decreased tumor size,
increased IgG and IFN-γ levels and increased CTL activity (64). Another way to activate the DC immune
response is via genetic modification. A genetically-modified DC
vaccine expressing HPV16 E6/E7 fusion oncoproteins was able elicit
the action of E6/E7-specific CTLs against cervical carcinoma CaSki
cells and induce their apoptosis. These elicited specific
protective immunity functions by the HPV-16 E6/E7 fusion protein
produced an effective approach against cervical cancer cell growth
(65).
Different types of vaccine have been investigated;
one of the most important mechanisms for their utilization is based
on their initiating the damaged immune response by hrHPV. Schematic
representation of this has been presented in Fig. 2.
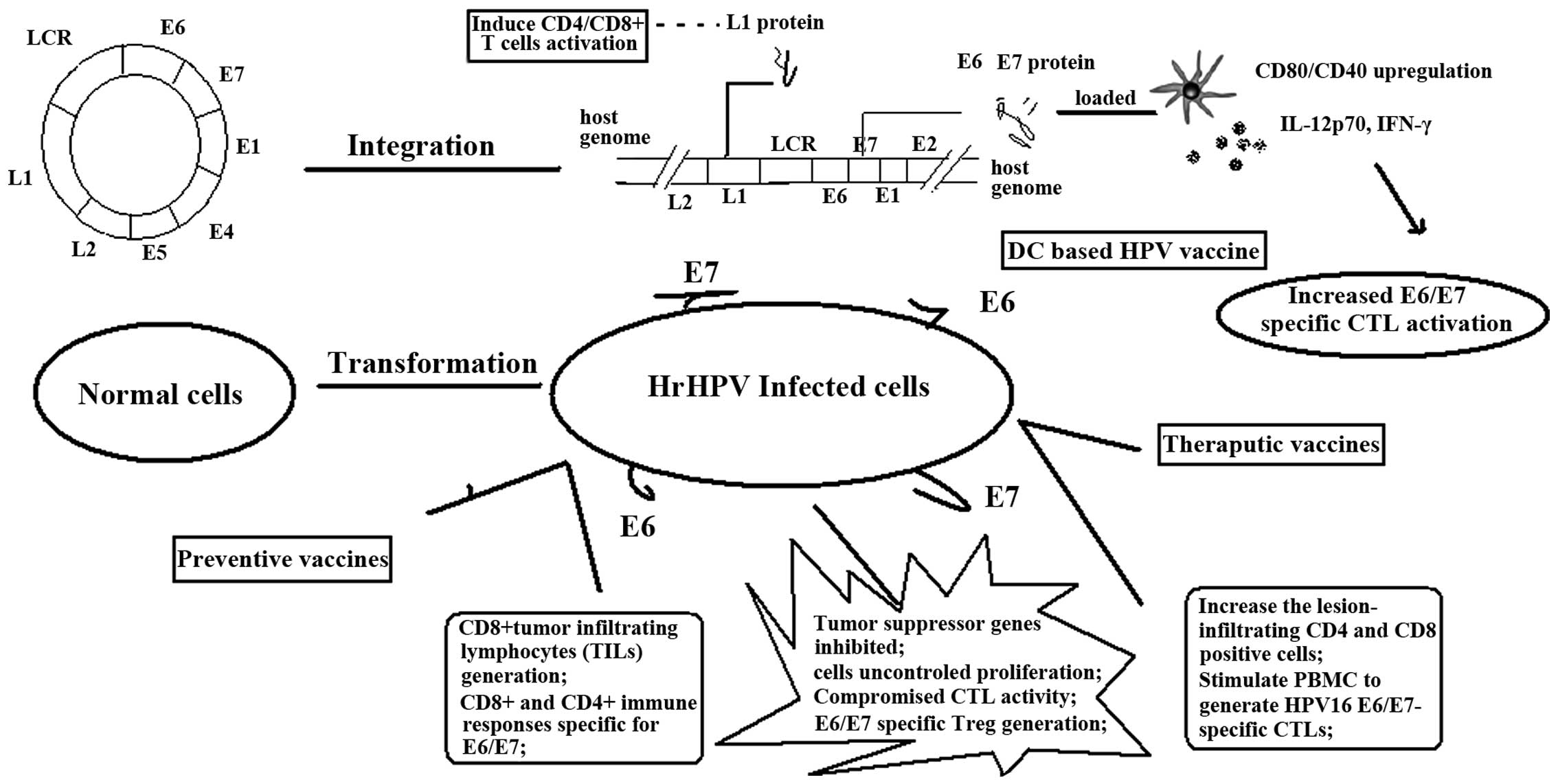 | Figure 2.Mechanisms for vaccines in the
prevention and therapy of HPV infection and CIN progression.
Vaccines aim to improve the HPV oncoprotein-specific immune
response, such as cytokine production, CTL initiation and immune
system modification, contributing to the treatment of every stage
from HPV infection to CIN to cervical cancer. CD, cluster of
differentiation; IL, interleukin; IFN, interferon; DC, dendritic
cell; hrHPV, high-risk human papilloma virus; CTL, cytotoxic T
cell; PBMC, peripheral blood mononuclear cell; Treg, regulatory T
cell; LCR, long control region. |
Conclusion
The present review investigated the association
between hrHPV and cervical cancer. The immune system plays an
important role from HPV infection to CIN and from CIN to cervical
cancer. A compromised immune response is the prerequisite for
disease progression. One unique feature of HPV infection is that it
can affect the immune system in such as way that it presents a much
more tolerant state, which facilitates persistent hrHPV infection
and cervical lesion progression. The counteraction of hrHPV
infection by vaccines is an important tool to stop the course of
the infection or oncogenesis. To date, numerous preventive and
therapeutic vaccines have been discovered, with the aim of
enhancing the damaged immune response to clear the virus and the
tumor cells.
References
|
1
|
Parkin DM and Bray F: Chapter 2: The
burden of HPV-related cancers. Vaccine. 24 (Suppl 3):(3): 11–25.
2006. View Article : Google Scholar
|
|
2
|
Almonte M, Albero G, Molano M, et al: Risk
factors for human papillomavirus exposure and co factors for
cervical cancer in Latin America and the Caribbean. Vaccine.
26:L16–L36. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Deligeoroglou E, Giannouli A,
Athanasopoulos N, et al: HPV infection: Immunological aspects and
their utility in future therapy. Infect Dis Obstet Gynecol.
2013:5408502013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bernard HU, Burk RD, Chen Z, et al:
Classification of papillomaviruses (PVs) based on 189 PV types and
proposal of taxonomic amendments. Virology. 401:70–79. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bouvard V, Baan R, Straif K, et al: A
review of human carcinogens-Part B: Biological agents. Lancet
Oncol. 10:321–322. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Coutlée F, Rouleau D, Petignat P, et al:
Enhanced detection and typing of human papillomavirus (HPV) DNA in
anogenital samples with PGMY primers and the Linear array HPV
genotyping test. J Clin Microbiol. 44:1998–2006. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Smith JS, Lindsay L, Hoots B, et al: Human
papillomavirus type distribution in invasive cervical cancer and
high-grade cervical lesions: A meta-analysis update. Int J Cancer.
121:621–632. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cerqueira C, Liu Y, Kühling L, et al:
Heparin increases the infectivity of Human Papillomavirus type 16
independent of cell surface proteoglycans and induces L1 epitope
exposure. Cell Microbiol. 15:1818–1836. 2013.PubMed/NCBI
|
|
9
|
Surviladze Z, Dziduszko A and Ozbun MA:
Essential roles for soluble virion-associated heparan sulfonated
proteoglycans and growth factors in human papillomavirus
infections. PLoS Pathog. 8:e10025192012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Raff AB, Woodham AW, Raff LM, et al: The
evolving field of human papillomavirus receptor research: A review
of binding and entry. J Virol. 87:6062–6072. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Horvath CA, Boulet GA, Renoux VM, Delvenne
PO and Bogers JP: Mechanisms of cell entry by human
papillomaviruses: An overview. Virol J. 7:112010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Asiaf A, Ahmad ST, Mohammad SO and Zargar
MA: Review of the current knowledge on the epidemiology,
pathogenesis and prevention of human papillomavirus infection. Eur
J Cancer Prev. 23:206–224. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Venuti A, Paolini F, Nasir L, et al:
Papillomavirus E5: the smallest oncoprotein with many functions.
Mol Cancer. 10:1402011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Campo MS, Graham SV, Cortese MS, et al:
HPV-16 E5 down-regulates expression of surface HLA class I and
reduces recognition by CD8 T cells. Virology. 407:137–142. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Faridi R, Zahra A, Khan K and Idrees M:
Oncogenic potential of human papillomavirus (HPV) and its relation
with cervical cancer. Virol J. 8:2692011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zuna RE, Allen RA, Moore WE, Mattu R and
Dunn ST: Comparison of human papillomavirus genotypes in high-grade
squamous intraepithelial lesions and invasive cervical carcinoma:
Evidence for differences in biologic potential of precursor
lesions. Mod Pathol. 17:1314–1322. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hasan UA, Zannetti C, Parroche P, et al:
The human papillomavirus type 16 E7 oncoprotein induces a
transcriptional repressor complex on the Toll-like receptor 9
promoter. J Exp Med. 210:1369–1387. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim H, Kwon B and Sin JI: Combined
stimulation of IL-2 and 4-1BB receptors augments the antitumor
activity of E7 DNA vaccines by increasing Ag-specific CTL
responses. PLoS One. 8:e837652013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bedoya AM, Jaramillo R, Baena A, et al:
Location and density of immune cells in precursor lesions and
cervical cancer. Cancer Microenviron. Jan 31–2012.(Epub ahead of
print). PubMed/NCBI
|
|
20
|
Stanley MA and Sterling JC: Host responses
to infection with human papillomavirus. Curr Probl Dermatol.
45:58–74. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Crosbie EJ, Einstein MH, Franceschi S and
Kitchener HC: Human papillomavirus and cervical cancer. Lancet.
382:889–899. 2013.PubMed/NCBI
|
|
22
|
Piersma SJ: Immunosuppressive tumor
microenvironment in cervical cancer patients. Cancer Microenviron.
4:361–375. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Alves DB, Tozetti IA, Gatto FA, et al: CD4
and CD8 T lymphocytes and NK cells in the stroma of the uterine
cervix of women infected with human papillomavirus. Rev Soc Bras
Med Trop. 43:425–429. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Feller L, Wood NH, Khammissa RA, et al:
HPV modulation of host immune responses. SADJ. 65:266–268.
2010.PubMed/NCBI
|
|
25
|
Le Borgne M, Etchart N, Goubier A, et al:
Dendritic cells rapidly recruited into epithelial tissues via
CCR6/CCL20 are responsible for CD8+ T cell cross priming
in vivo. Immunity. 24:191–201. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hasan U: Human papillomavirus (HPV)
deregulation of Toll-like receptor 9. Oncoimmunology. 3:e272572014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang Y, Yang H, Barnie PA, et al: The
expression of Toll-like receptor 8 and its relationship with VEGF
and Bcl-2 in cervical cancer. Int J Med Sci. 11:608–613. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang Y, Weng Y, Shi Y, et al: Expression
and Functional Analysis of Toll-like Receptor 4 in Human Cervical
Carcinoma. J Membr Biol. 247:591–599. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hammes LS, Tekmal RR, Naud P, Edelweiss
MI, et al: Macrophages, inflammation and risk of cervical
intraepithelial neoplasia (CIN) progression-clinicopathological
correlation. Gynecologic Oncology. 105:157–165. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lepique AP, Daghastanli KR, Cuccovia IM
and Villa LL: HPV16 tumor associated macrophages suppress antitumor
T cell responses. Clinical Cancer Research. 15:4391–4400. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Garcia-Iglesias T, Del Toro-Arreola A,
Albarran-Somoza B, et al: Low NKp30, NKp46 and NKG2D expression and
reduced cytotoxic activity on NK cells in cervical cancer and
precursor lesions. BMC Cancer. 9:1862009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jimenez-Perez MI, Jave-Suarez LF,
Ortiz-Lazareno PC, et al: Cervical cancer cell lines expressing
NKG2D-ligands are able to down-modulate the NKG2D receptor on NKL
cells with functional implications. BMC Immunol. 13:72012.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kobayashi A, Weinberg V, Darragh T and
Smith-McCune K: Evolving immunosuppressive microenvironment during
human cervical carcinogenesis. Mucosal Immunol. 1:412–420. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bais AG, Beckmann I, Lindemans J, et al: A
shift to a peripheral Th2-type cytokine pattern during the
carcinogenesis of cervical cancer becomes manifest in CIN/III
lesions. J Clin Pathol. 58:1096–1100. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Scott ME, Shvetsov YB, Thompson PJ, et al:
Cervical cytokines and clearance of incident human papillomavirus
infection: Hawaii HPV cohort study. Int J Cancer. 133:1187–1196.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Peghini BC, Abdalla DR, Barcelos AC,
Teodoro L, Murta EF and Michelin MA: Local cytokine profiles of
patients with cervical intraepithelial and invasive neoplasia. Hum
Immunol. 73:920–926. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lee YS, Lee CW, Song MJ, et al:
Cell-mediated immune response to human papillomavirus 16 E7 peptide
pools in patients with cervical neoplasia. Acta Obstet Gynecol
Scand. 90:1350–1356. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sheu BC, Chang WC, Lin HH, Chow SN and
Huang SC: Immune concept of human papillomaviruses and related
antigens in local cancer milieu of human cervical neoplasia. J
Obstet Gynaecol Res. 33:103–113. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Iijima N, Goodwin EC, Dimaio D and Iwasaki
A: High-risk human papillomavirus E6 inhibits monocyte
differentiation to Langerhans cells. Virology. 444:257–262. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Strickler HD, Martinson J, Desai S, et al:
The relation of plasmacytoid dendritic cells (pDCs) and regulatory
T-cells (Tregs) with HPV persistence in HIV-infected and
HIV-uninfected women. Viral Immunol. 27:20–25. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yang W, Song Y, Lu YL, Sun JZ and Wang HW:
Increased expression of programmed death (PD)-1 and its ligand
PD-L1 correlates with impaired cell-mediated immunity in high-risk
human papillomavirus-related cervical intraepithelial neoplasia.
Immunology. 139:513–522. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Adurthi S, Krishna S, Mukherjee G, et al:
Regulatory T cells in a spectrum of HPV-induced cervical lesions:
Cervicitis, cervical intraepithelial neoplasia andsquamous cell
carcinoma. Am J Reprod Immunol. 60:55–65. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
de Vos van Steenwijk PJ, Piersma SJ,
Welters MJ, et al: Surgery followed by persistence of high-grade
squamous intraepithelial lesions is associated with the induction
of a dysfunctional HPV16-specific T-cell response. Clin Cancer Res.
14:7188–7195. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ali KS, Ali HY and Jubrael JM:
Concentration levels of IL-10 and TNF α cytokines in patients with
human papilloma virus (HPV) DNA+ and DNA-cervical
lesions. J Immunotoxicol. 9:168–172. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bermudez-Morales VH, Gutierrez LX,
Alcocer-Gonzalez JM, Burguete A and Madrid-Marina V: Correlation
between IL-10 gene expression and HPV infection in cervical cancer:
a mechanism for immune response escape. Cancer Invest.
26:1037–1043. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bolpetti A, Silva JS, Villa LL and Lepique
AP: Interleukin-10 production by tumor infiltrating macrophages
plays a role in Human Papillomavirus 16 tumor growth. BMC Immunol.
11:272010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bermudez-Morales VH, Peralta-Zaragoza O,
Alcocer Gonzalez JM, Moreno J and Madrid-Marina V: IL-10 expression
is regulated by HPV E2 protein in cervical cancer cells. Mol Med
Rep. 4:369–375. 2011.PubMed/NCBI
|
|
48
|
Koshiol J, Sklavos M, Wentzensen N, et al:
Evaluation of a multiplex panel of immune-related markers in
cervical secretions: a methodologic study. Int J Cancer.
134:411–425. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zeng C, Yao Y, Jie W, et al: Up-regulation
of Foxp3 participates in progression of cervical cancer. Cancer
Immunol Immunother. 62:481–487. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Sheng X, Du X, Zhang X, et al: Clinical
value of serum HMGB1 levels in early detection of recurrent
squamous cell carcinoma of uterine cervix: Comparison with serum
SCCA, CYFRA21-1 and CEA levels. Croat Med J. 50:455–464. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Huang LF, Yao YM, Zhang LT, Dong N, Yu Y
and Sheng ZY: The effect of high-mobility group box 1 protein on
activity of regulatory T cells after thermal injury in rats. Shock.
31:322–329. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Pang X, Zhang Y, Wei H, et al: Expression
and effects of high-mobility group box 1 in cervical cancer. Int J
Mol Sci. 15:8699–8712. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Halloush RA, Akpolat I, Jim Zhai Q,
Schwartz MR and Mody DR: Comparison of ProEx C with p16INK4a and
Ki-67 immunohistochemical staining of cell blocks prepared from
residual liquid-based cervicovaginal material: A pilot study.
Cancer. 114:474–480. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Izadi-Mood N, Sarmadi S, Eftekhar Z,
Jahanteegh HA and Sanii S: Immunohistochemical expression of p16
and HPV L1 capsid proteins as predictive markers in cervical
lesions. Arch Gynecol Obstet. 289:1287–1292. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Lee SJ, Lee AW, Kang CS, et al:
Clinicopathological implications of human papilloma virus (HPV) L1
capsid protein immunoreactivity in HPV16-positive cervical
cytology. Int J Med Sci. 11:80–86. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Pinto LA, Edwards J, Castle PE, et al:
Cellular immune responses to human papillomavirus (HPV)-16 L1 in
healthy volunteers immunized with recombinant HPV-16 L1 virus-like
particles. J Infect Dis. 188:327–338. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Chen Z, Kamath P, Zhang S, St John L,
Adler-Storthz K and Shillitoe EJ: Effects on tumor cells of
ribozymes that cleave the RNA transcripts of human papillomavirus
type 18. Cancer Gene Ther. 3:18–23. 1996.PubMed/NCBI
|
|
58
|
Knoff J, Yang B, Hung CF and Wu TC:
Cervical cancer: development of targeted therapies beyond molecular
pathogenesis. Curr Obstet Gynecol Rep. 3:18–32. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Souders NC, Sewell DA, Pan ZK, et al:
Listeria-based vaccines can overcome tolerance by expanding low
avidity CD8+ T cells capable of eradicating a solid
tumor in a transgenic mouse model of cancer. Cancer Immu.
7:22007.
|
|
60
|
Zhou CM, Zhang GX and Ma XX:
Characterization and evaluation of the immune responses elicited by
a novel human papillomavirus (HPV) therapeutic vaccine: HPV
16E7-HBcAg-Hsp65 fusion protein. J Virol Methods. 197:1–6. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Davidson EJ, Boswell CM, Sehr P, et al:
Immunological and clinical responses in women with vulval
intraepithelial neoplasia vaccinated with a vaccinia virus encoding
human papillomavirus 16/18 oncoproteins. Cancer Res. 63:6032–6041.
2003.PubMed/NCBI
|
|
62
|
Mizuuchi M, Hirohashi Y, Torigoe T, et al:
Novel oligomannose liposome-DNA complex DNA vaccination efficiently
evokes anti-HPV E6 and E7 CTL responses. Exp Mol Pathol.
92:185–190. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Wang YT, Li W, Liu Q, Guan X and Hu J:
Dendritic cells treated with HPV16mE7 in a three-dimensional model
promote the secretion of IL-12p70 and IFN-γ. Exp Mol Pathol.
91:325–330. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Wang HL, Xu H, Lu WH, Zhu L, Yu YH and
Hong FZ: In vitro and in vivo evaluations of human
papillomavirus type 16 (HPV16)-derived peptide-loaded dendritic
cells (DCs) with a CpG oligodeoxynucleotide (CpG-ODN) adjuvant as
tumor vaccines for immunotherapy of cervical cancer. Arch Gynecol
Obstet. 289:155–162. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Wu XM, Liu X, Jiao QF, et al: Cytotoxic T
Lymphocytes elicited by dendritic cell-targeted delivery of human
papillomavirus Type-16 E6/E7 fusion gene exert lethal
effects on CaSki cells. Asian Pac J Cancer Prev. 15:2447–2451.
2014. View Article : Google Scholar : PubMed/NCBI
|














