Introduction
The role of sialic acid as a biological mask of
surface structures is well-documented (1,2). Sialic
acid plays an important role in the biological behavior of tumor
cells, with respect to cell recognition phenomena, metastasis
(3–5)
and cell adhesion (6), and influences
the clinical outcome of patients (7,8). We have
previously reported that sialylation of L-phyohemagglutinin (L-PHA)
reactive oligosaccharides is associated with worse prognosis in
patients with diffuse large B-cell lymphoma (7,8). Another
study has demonstrated that differential cell surface sialylation
of Burkitt lymphoma cell lines affects the cell adhesion to
fibronectin and collagen type IV (9).
In addition, differential cell surface sialylation is due to
differences in the mRNA expression of UDP-GlcNAc2-epimerase, which
is a key enzyme in sialic acid biosynthesis (9). Furthermore, Fas-induced apoptosis was
found to be regulated by sialylation of the Fas molecule in the
Jurkat T cell lymphoma cell line (10).
Doxorubicin is one of the chemotherapeutic agents
used in the therapy of human malignant lymphoma (11). In addition, etoposide, a topoisomerase
II inhibitor, is another chemotherapeutic agent used in malignant
lymphoma (12). Etoposide-induced
apoptosis has been reported to be regulated through caspase
activation, mitochondrial damage and cytochrome c release
(13–16). However, only a limited number of
studies have investigated the association between cell surface
sialylation and drug resistance in tumor cell lines (17). The aim of the present study was to
investigated whether cell surface sialylation may regulate
etoposide-induced apoptosis in a caspase-dependent manner in the
HBL-2 human malignant lymphoma cell line.
Materials and methods
Cell line
The HBL-2 human lymphoma cell line was established
in the laboratory of the Department of Diagnostic Pathology
(Fukushima Medical University, Fukushima, Japan) from a patient
suffering from diffuse large B-cell lymphoma. The study was
approved by the Ethics Committee of Fukushima Medical University
(Fukushima, Japan). The HBL-2 cells were grown in RPMI 1640
(Sigma-Aldrich, St. Louis, MO, USA) culture medium containing 15%
fetal calf serum (FCS) in a 5%CO2 atmosphere at 37°C
(18).
Reagents
Limax flavus agglutinin (LFA), a biotinylated
lectin, was purchased from EY Laboratories, Inc. (San Mateo, CA,
USA). Neuraminidase from Vibrio Cholerae was obtained from
Roche Diagnostics GmbH, Mannheim, Germany.
Flow cytometry
HBL-2 cells (5×105 cells) were suspended
in 100 µl phosphate-buffered saline (PBS), incubated at 4°C for 20
min with 5 µl biotinylated LFA lectin and washed twice with PBS.
The cells were then incubated at 4°C for 20 min with 5 µl
avidin-fluorescein isothiocyanate (Vector Laboratories, Inc.,
Burlingame, CA, USA) and washed twice with PBS. The fluorescent
intensities were analyzed using a FACScan device (BD Biosciences,
Mountain View, CA, USA). In order to analyze cell surface
sialylation, 6×106 cells were incubated at 37°C for 30
min in 200 µl RPMI 1640 medium containing 15% FCS and 40 µl of 1
U/ml Vibrio Cholerae neuraminidase prior to incubation with
biotinylated LFA lectin (9). Lectin
reactivity was then analyzed by flow cytometry.
LFA is a sialic acid-specific lectin, reacts with
the surfaces of HBL-2 cells, which was observed using flow
cytometry. LFA lectin reactivity is completely eliminated by
neuraminidase treatment, indicating that sialic acids on the cell
surface are removed (19).
Doxorubicin or etoposide-induced cell
death
Cell viability was assessed using a WST-1 Cell
Proliferation Assay kit (Roche Diagnostics) (10). HBL-2 cells were grown for 2 days and
then seeded in 96-well microtiter plates, at a density of
5×104 cells/well. Next, the cells were incubated for 24,
48 or 72 h at 37°C in 100 µl culture medium containing doxorubicin
(D1515; Sigma-Aldrich, St. Louis, MO, USA) at a final concentration
of 1.6 or 3.2 µM, or incubation with etoposide (40 µg/ml, for 24h;
E1383; Sigma-Aldrich) at a final concentration of 10 or 30 µg/ml.
Subsequent to the incubation, 10 µl WST-1 reagent was added to each
well and the plates were incubated for a further 1 or 2 h at 37°C.
Then the absorbance at 450 nm was measured with an i-MARK
microplate reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
To examine the extent of cell surface sialylation, 6×106
cells were incubated at 37°C for 30 min in 200 µl RPMI 1640
containing 15% FCS and 40 µl of 1 U/ml Vibrio Cholerae
neuraminidase, prior to incubation with doxorubicin or etoposide
(10).
Detection of apoptosis
The cultured cells were cytospun using a Cytospin2
(Thermo Fisher Scientific, Inc., Waltham, MA, USA), Japan. The
cells which adhered to the slide glass were stained using Giemsa
solution (Wako Pure Chemical Industries, Ltd., Chuo-ku, Japan), as
to the manufacturer's instructions. The apoptosis-associated
morphological changes, including nuclear fragmentation,
condensation and membrane changes were evaluated by light
microscope (BX51; Olympus Corporation, Tokyo, Japan).
Giemsa-stained cytospin cell preparations were evaluated to detect
any apoptosis-associated morphological changes, including nuclear
fragmentation, condensation and membrane changes.
Caspase3, caspase-8 and caspase-9
activities
Upon induction of apoptosis with etoposide, the
caspase-3, caspase-8 and caspase-9 activities were measured using a
colorimetric caspase activity assay kit (Apopcyto™; Medical &
Biological Laboratories Co., Ltd., Tokyo, Japan), according to the
manufacturer's instructions (20).
Statistical analysis
P-values were calculated based on Student's t-test
using Microsoft Office Excel software, version 2007 (Microsoft
Corporation, Redmond, WA, USA), and P<0.05 was considered to
indicate a statistically significant difference.
Results
Doxorubicin- and etoposide-induced
cell death
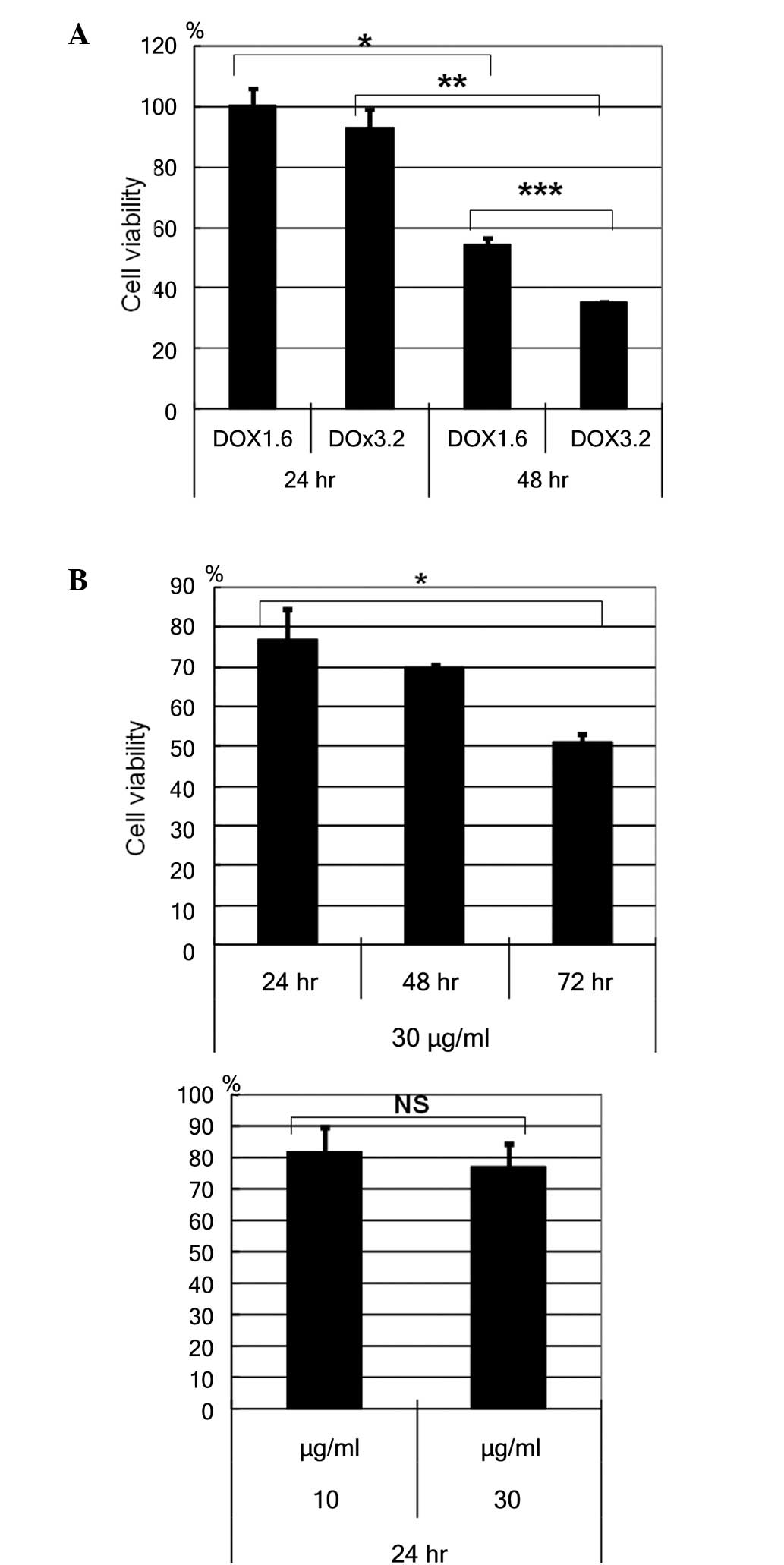
HBL-2 cells were incubated with or without
doxorubicin, at a concentration of 1.6 or 3.2 µM for the cell
viability assay using WST reagent (Fig.
1): The cell viability of HBL-2 cells reduced in a dose- and
time- dependent manner (Fig. 1A). The
time dependent reduction of cell viability was observed with 1.6 µM
(P=0.0002) and 3.2 µM (P<0.0001) doxorucin treatment. A dose
dependent reduction in cell viability was observed at 48h
(P=0.0001). In addition, HBL-2 cells were incubated with or without
etoposide, at a concentration of 10 or 30 µg/ml. The cell viability
of HBL-2 cells reducedd in a time-dependent manner, but not in a
dose-dependent manner (Fig. 1B).
Effect of neuraminidase treatment on
doxorubicin- or etoposide-induced cell death
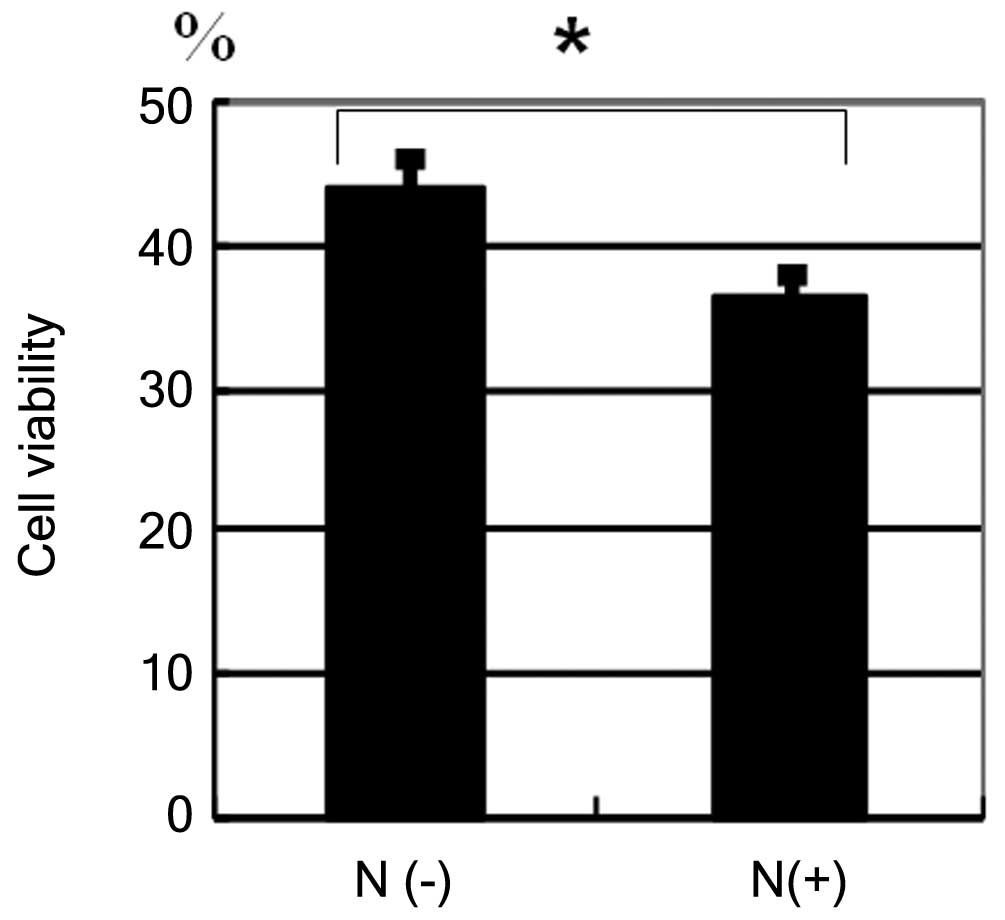
HBL-2 cells were incubated with etoposide at a
concentration of 30 µg/ml (Fig. 2).
After 72 h, the reduction in cell viability of
neuraminidase-pretreated cells was greater when compared with cells
that were not pretreated with neuraminidase (Fig. 2, P=0.0021).
Detection of apoptosis
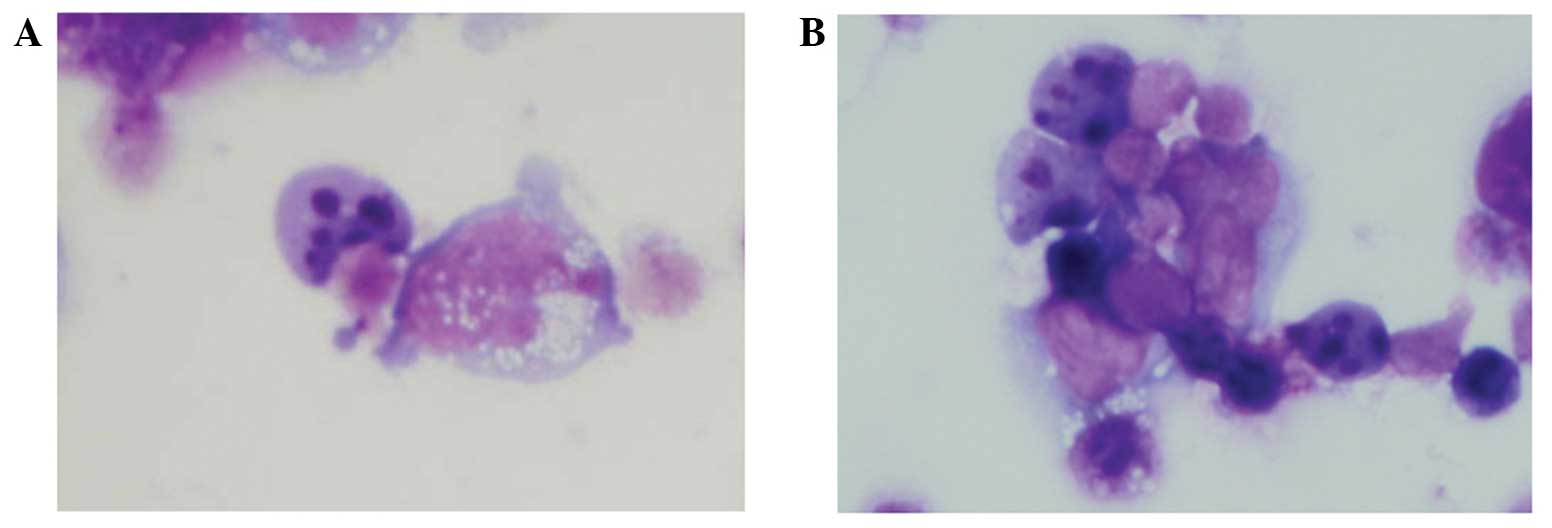
HBL-2 cells that were incubated with etoposide for
24 h demonstrated an apoptotic morphology (including nuclear
fragmentation) on Giemsa-stained samples (Fig. 3A). The apoptotic morphology was also
observed in HBL-2 cells pretreated with neuraminidase (Fig. 3B).
Caspase-3, caspase-8 and caspase-9
activities
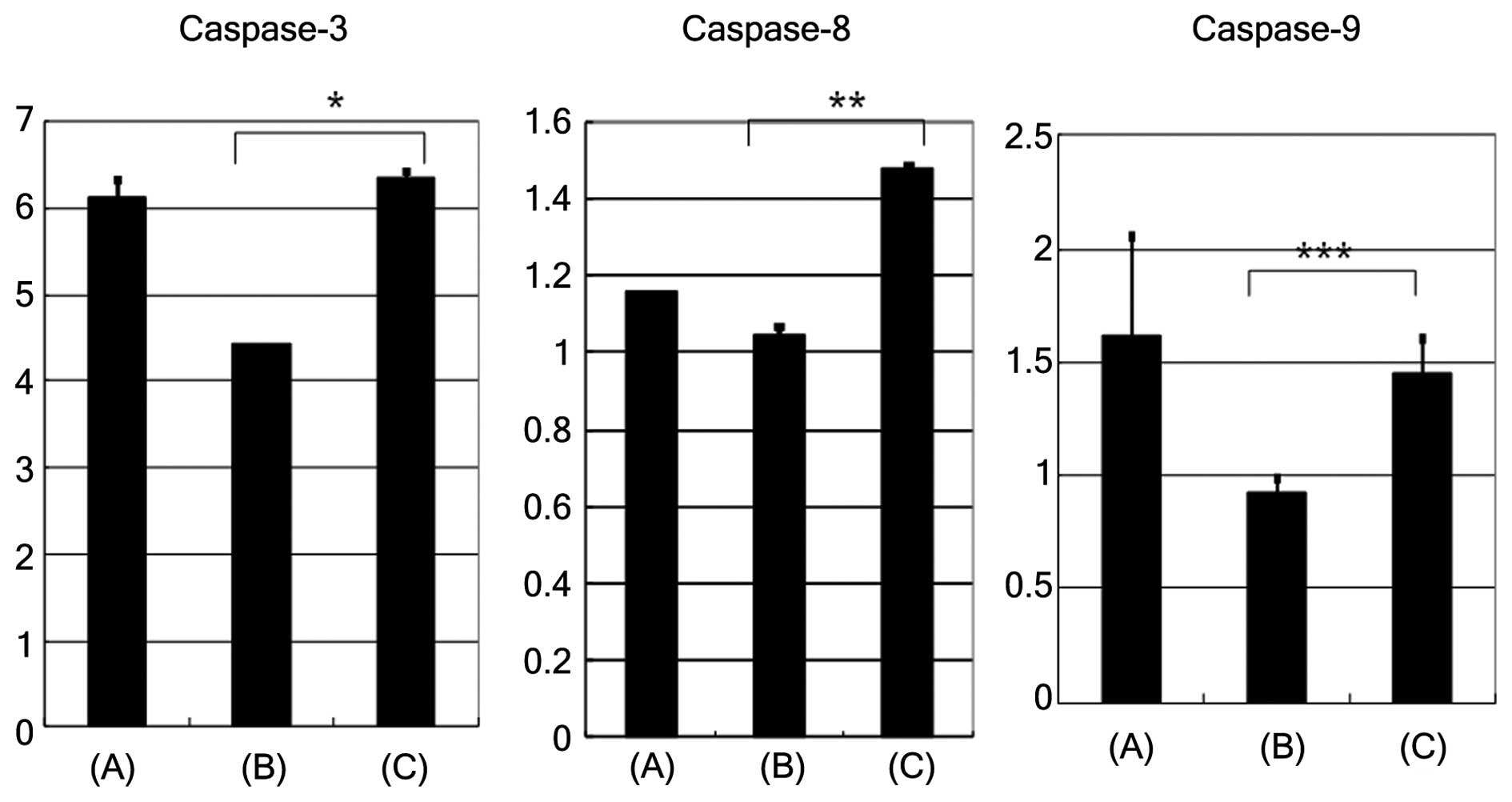
Upon induction of apoptosis with etoposide, the
caspase-3, caspase-8 and caspase-9 activities (activity/h/mg of
protein) were found to be higher in cells pretreated with
neuraminidase, compared with cells that were not pretreated
(Fig. 4; caspase-3, P=0.0011;
caspase-8, P=0.0014; caspase-9, P>0.05). The removal of sialic
acid by neuraminidase pretreatment resulted in enhancement of
caspase-3, -8 and 9 activity compared with the absence of
neuraminidase pre-treatment.
Discussion
Etoposide is a topoisomerase II inhibitor that has
been widely used to couple DNA damage to apoptosis (21). Topoisomerase II is able to unknot and
untangle DNA molecules by passing an intact helix through a
transient double-stranded break (13). Inhibitors, such as etoposide,
stabilize the complex formed by topoisomerase II and form
non-repairable DNA double-strand breaks. Subsequently, cells are
able to recognize these DNA damages and eliminate the injured cells
by apoptosis.
Etoposide-induced apoptosis has previously been
found to be dependent on caspase activation and mediated by
mitochondrial damage (22).
Doxorubicin-induced apoptosis is also mediated by caspase-9
activation (23). Scaffidi et
al identified two different types of Fas-induced apoptosis
signaling pathways (24). Type I
apoptosis is mediated by a death domain and activation of caspase-8
in various cell lines (24). By
contrast, type II apoptosis is mediated by activation of caspase-9
and loss of mitochondrial membrane potential (Δψm). The activities
of the death domain and caspase-8 in type I apoptosis are lower
compared to those in type II apoptosis (24). In the present study, etoposide-induced
apoptosis was found to be mediated by the activation of both
caspase-8 and caspase-9. Therefore, although the detailed
underlying mechanisms remain unclear, cell surface sialylation
inhibited the pathway of apoptosis depending on the activation of
caspase-8, caspase-9 and caspase-3. In addition, sialic acids may
regulate both type I and type II apoptosis.
Cell surface sialylation in human lymphoma cells has
been reported to be involved in modulating sensitivity towards
Fas-mediated apoptotic cell death (10). Previous studies have suggested that
alteration in the extent of sialylation in the cell surface of
tumor cells appeared to be closely associated with cell adhesion,
metastasis and the clinical outcome of the patients (7). Our previous study revealed that
α2,6-sialic acid residues or sialylation in L-PHA reactive
oligosaccharides appeared to be closely associated with worse
prognosis in patients suffering from human diffuse large B-cell
lymphoma (8). Differential cell
surface sialylation regulates lymphoma cell adhesion to the
extracellular matrix (9). In the
present study, cell surface sialylation inhibited etoposide-induced
apoptosis and this drug-induced apoptosis was caspase-3
dependent.
In conclusion, the results of the present study may
provide new scientific foundation on the cell survival mechanism in
drug-induced apoptosis and sialylation of cell surface glycans,
which appeared to inhibit the anticancer drug effect in lymphoma.
The current authors speculate that altered cell surface sialylation
may be useful for more effective cell killing in human malignant
lymphoma.
References
|
1
|
Varki A: Sialic acids as ligands in
recognition phenomena. FASEB J. 11:248–255. 1997.PubMed/NCBI
|
|
2
|
Kelm S and Schauer R: Sialic acids in
molecular and cellular interactions. Int Rev Cytol. 175:137–240.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Altevogt P, Fogel M, Cheingsong-Popov R,
Dennis J, Robinson P and Schirrmacher V: Different patterns of
lectin binding and cell surface sialylation detected on related
high- and low-metastatic tumor lines. Cancer Res. 43:5138–5144.
1983.PubMed/NCBI
|
|
4
|
Yogeeswaran G and Salk PL: Metastatic
potential is positively correlated with cell surface sialylation of
cultured murine tumor cell lines. Science. 212:1514–1516.
1981.PubMed/NCBI
|
|
5
|
Abe M, Suzuki O, Tasaki K, Tominaga K and
Wakasa H: Analysis of lectin binding properties on human Burkitt's
lymphoma cell lines that show high spontaneous metastasis to
distant organs in SCID mice: The binding sites for soybean
agglutinin lectin masked by sialylation are closely associated with
metastatic lymphoma cells. Pathol Int. 46:977–983. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dennis J, Waller C, Timpl R and
Schirrmacher V: Surface sialic acid reduces attachment of
metastatic tumour cells to collagen type IV and fibronectin.
Nature. 300:274–276. 1982. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Suzuki O, Nozawa Y, Kawaguchi T and Abe M:
Phaseolus vulgaris leukoagglutinating lectin-binding
reactivity in human diffuse large B-cell lymphoma and its relevance
to the patient's clinical outcome: Lectin histochemistry and lectin
blot analysis. Pathol Int. 49:874–880. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Suzuki O, Nozawa Y, Kawaguchi T and Abe M:
Alpha-2,6-sialylation of L-PHA reactive oligosaccharides and
expression of N-acetylglucosaminyltransferase V in human diffuse
large B cell lymphoma. Oncol Rep. 10:1759–1764. 2003.PubMed/NCBI
|
|
9
|
Suzuki O, Nozawa Y, Kawaguchi T and Abe M:
UDP-GlcNAc2-epimerase regulates cell surface sialylation and cell
adhesion to extracellular matrix in Burkitt's lymphoma. Int J
Oncol. 20:1005–1011. 2002.PubMed/NCBI
|
|
10
|
Suzuki O, Nozawa Y and Abe M: Sialic acids
linked to glycoconjugates of Fas regulate the caspase-9-dependent
and mitochondria-mediated pathway of Fas-induced apoptosis in
Jurkat T cell lymphoma. Int J Oncol. 23:769–774. 2003.PubMed/NCBI
|
|
11
|
Halaas JL, Moskowitz CH, Horwitz S, et al:
ADR-CHOP-14 in patients with diffuse large B-cell lymphoma:
Feasibility and preliminary efficacy. Leuk Lymphoma. 46:541–547.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zinzani PL, Gherlinzoni F, Storti S,
Zaccaria A, Pavone E, Moretti L, Gentilini P, Guardigni L, De Renzo
A, Fattori PP, et al: Randomized trial of 8-week versus 12-week
VNCOP-B plus G-CSF regimens as front-line treatment in elderly
aggressive non-Hodgkin's lymphoma patients. Ann Oncol.
13:1364–1369. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen GL, Yang L, Rowe TC, Halligan BD,
Tewey KM and Liu LF: Nonintercalative antitumor drugs interfere
with the breakage-reunion reaction of mammalian DNA topoisomerase
II. J Biol Chem. 259:13560–13566. 1984.PubMed/NCBI
|
|
14
|
Burden DA and Osheroff N: Mechanism of
action of eukaryotic topoisomerase II and drugs targeted to the
enzyme. Biochim Biophys Acta. 1400:139–154. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Karpinich NO, Tafani M, Rothman RJ, Russo
MA and Farber JL: The course of etoposide-induced apoptosis from
damage to DNA and p53 activation to mitochondrial release of
cytochrome c. J Biol Chem. 277:16547–16552. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mizumoto K, Rothman RJ and Farber JL:
Programmed cell death (apoptosis) of mouse fibroblasts is induced
by the topoisomerase II inhibitor etoposide. Mol Pharmacol.
46:890–895. 1994.PubMed/NCBI
|
|
17
|
Ma H, Zhou H, Song X, Shi S, Zhang J and
Jia L: Modification of sialylation is associated with multidrug
resistance in human acute myeloid leukemia. Oncogene. 34:726–740.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Suzuki O, Nozawa Y and Abe M: Regulatory
roles of altered N-and O-glycosylation of CD45 in
galectin-1-induced cell death in human diffuse large B cell
lymphoma. Int J Oncol. 26:1063–1068. 2005.PubMed/NCBI
|
|
19
|
Suzuki O, Nozawa Y and Abe M: The
regulatory roles of cell surface sialylation and N-glycans in human
B cell lymphoma cell adhesion to galectin-1. Int J Oncol.
28:155–160. 2006.PubMed/NCBI
|
|
20
|
Lin CF, Chen CL, Chang WT, Jan MS, Hsu LJ,
Wu RH, Tang MJ, Chang WC and Lin YS: Sequential caspase-2 and
caspase-8 activation upstream of mitochondria during ceramideand
etoposide-induced apoptosis. J Biol Chem. 279:40755–40761. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Boesen-de Cock JG, Tepper AD, de Vries E,
van Blitterswijk WJ and Borst J: Common regulation of apoptosis
signaling induced by CD95 and the DNA-damaging stimuli etoposide
and gamma-radiation downstream from caspase-8 activation. J Biol
Chem. 274:14255–14261. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chandra D, Choy G, Deng X, Bhatia B,
Daniel P and Tang DG: Association of active caspase 8 with the
mitochondrial membrane during apoptosis : Potential roles in
cleaving mitochondrion-endoplasmic reticulum cross talk in
etoposide-induced cell death. Mol Cell Biol. 24:6592–6607. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gamen S, Anel A, Perez-Galan P, Lasierra
P, Johnson D, Pineiro A and Naval J: Doxorubicin treatment
activates a Z-VAD-sensitive caspase, which causes deltapsim loss,
caspase-9 activity, and apoptosis in Jurkat cells. Exp Cell Res.
258:223–35. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Scaffidi C, Fulda S, Srinivasan A, Friesen
C, Li F, Tomaselli KJ, Debatin KM, Krammer PH and Peter ME: Two
CD95 (APO-1/Fas) signaling pathways. EMBO J. 17:1675–1687. 1998.
View Article : Google Scholar : PubMed/NCBI
|