Introduction
Adipose-derived stem cells (ASCs) comprise a
plastic-adherent, proliferative, multipotent cell population
isolated from the stromal vascular fraction (SVF) of adipose tissue
(1). ASCs are capable of self-renewal
and are able to differentiate into various types of cell
populations, including adipocytes, osteoblasts, chondrocytes,
myoctyes and neurons. Thus, ASCs are valuable sources of stem cells
for use in regenerative medicine and cosmetic applications
(2). As with embryonic stem cells
(ESCs) and induced pluripotent stem cells (iPSCs), ASCs contribute
to tissue repair and skin rejuvenation, as well as bone and
cartilage formation. These therapeutic effects of ASCs are
considered to result from the differentiation of ASCs into multiple
cell types, as well as the paracrine action of the diverse
cytokines and growth factors secreted by ASCs onto adjacent tissues
(3,4).
Although ASCs are considered to be a promising
therapeutic tool with various potential clinical applications, the
effect of ASCs on tumor growth, metastasis and recurrence are
controversial. A large number of studies have indicated that ASCs
promote the proliferation of cancer cells. For example, when ASCs
were co-injected with tumor cells, they promoted the tumorigenic
growth of H460 human non-small cell lung cancer cells and U87MG
malignant glioma cells in nude mice (5). A similar tumorigenic effect was observed
in human prostate (6) and breast
(7) cancer cells in mice.
Furthermore, co-culture of ASCs enhanced the proliferation of human
squamous cell carcinoma cells (8) and
the migration of breast cancer cells in a xenograft model (9). However, conversely, numerous studies
have reported suppressive effects of ASCs on tumorigenesis. For
example, human mesenchymal stromal cells were demonstrated to
reduce lung metastasis, the growth of breast cancer cells (10), the proliferation of pancreatic cancer
cells (11), and LNCaP or PC3
prostate cancer cell-derived tumor growth in mice (12) by inducing cell death.
Melanoma is a malignant tumor of melanocytes. The
occurrence or melanoma has increased steadily between 2002 and
2011, with an average incidence increase of 1.4% per year in the
USA (13). Melanoma is a dangerous
disease entity if not detected in the early stages, as recurrence
is common (14). Linkage analysis has
revealed that families with melanoma possess mutations in the
p14ARF and p16INK4a tumor suppressor
proteins. Loss-of-function of these proteins may promote the
activation of cyclin-dependent kinase 4/6 (CDK4/6) via
destabilization of p53 and inhibition of retinoblastoma (RB)
protein (15), leading to
acceleration of the G1-S transition and re-entry into the cell
cycle (16). In addition, BRAF gene
mutations have recently been shown to indicate a predisposition to
melanoma (16). However, the detailed
molecular mechanisms regarding the development of melanoma have
remained poorly understood.
Recently, the effects of ASCs and their soluble
factors on the viability of various types of tumor have been
investigated; however, their effect has yet to be sufficiently
addressed in melanoma cells. Therefore, the present study aimed to
investigate whether ASC-conditioned medium (ASC-CM) treatment
affected the proliferation and migration of B16 melanoma cells. In
addition, the regulatory effect of ASC-CM on the tumorigensis of
B16 melanoma cells was evaluated in a mouse model.
Materials and methods
ASC-CM preparation
ASCs were provided by Dr Jong-Hyuk Sung (Yonsei
University, Seoul, Korea) (17). The
cells (4×105) were seeded onto a 100-mm cell culture
dish (BD Biosciences, Bedford, MA, USA) with the control medium
[Dulbecco's modified Eagle's medium (DMEM) supplemented with 10%
fetal bovine serum (FBS) (both from Invitrogen Life Technologies,
Carlsbad, CA, USA)] and incubated overnight at 37°C, prior to
culture in DMEM/F-12 serum-free media (Gibco Life Technologies,
Grand Island, NY, USA). Following three days of culture at 37°C,
ASC-CM was collected, centrifuged at a speed of 300 × g for 5 min
and filtered through a 0.22-mm syringe filter (EMD Millipore,
Bedford, MA, USA). The filtered ASC-CM was then applied to B16
melanoma cells (Korean Cell Line Bank, Seoul, Korea) at varying
dilutions (0, 10, 50 and 100%).
Cell culture
B16 melanoma cells were cultured in a growth medium
of DMEM containing 10% FBS and 1% cellgro® penicillin/streptomycin
solution (Mediatech, Inc., Manassas, VA, USA). The cells were grown
in a humidified incubator at 37°C and 5% CO2.
Cell proliferation assay
The B16 melanoma cells (1×103 cells/well)
were cultured in 96-well plates. Following one day of culture, the
media was replaced with DMEM containing 10% FBS and concentrated
ASC-CM. Incubation with 10X ASC-CM occurred for varying
time-periods (24, 48 and 72 h), while incubation with various
dilutions of ASC-CM (1, 5 and 10X) was performed for 72 h. At the
end of the indicated time-periods, cell proliferation was measured
using a Cell Counting Kit-8 (CCK-8 kit; Dojindo Molecular
Technologies, Inc., Gaithersburg, MD, USA), in accordance with the
manufacturer's instructions. In brief, 10 µl CCK-8 solution was
added to the cells and incubated for 3 h. Absorbance was
subsequently measured at a wavelength of 450 nm using a microplate
reader (SpectraFluor, Tecan Austria GmbH, Grödig, Austria). The
relative number of cells was calculated using comparable standard
curves of the obtained optical density values.
Cell cycle analysis
The B16 melanoma cells were seeded in a medium
containing 10% FBS at 37°C and allowed to grow to 60% confluence.
The cells were starved by incubation with serum-free medium for 24
h, and were then continuously cultured for 24 h with or without 10X
ASC-CM. Subsequently, the cells were harvested, washed twice with
phosphate-buffered saline (PBS; Gibco-BRL, Carlsbad, CA, USA) and
fixed in ice-cold 70% ethanol (Merck & Co., Kenilworth, NJ,
USA). The fixed cells were then washed once with PBS, incubated
with RNase A (1 µg/µl; Qiagen, Inc., Valencia, CA, USA) for 1 h at
37°C and treated with propidium iodide (50 µg/ml; Sigma-Aldrich,
St. Louis, MO, USA) for DNA staining. The distribution of the cell
cycle phases was analyzed using a FACScan™ flow cytometer (BD
Biosciences, San Jose, CA, USA).
Immunoblot analysis
Cell lysates were prepared in lysis buffer
containing 20 mM Tris (pH 7.5), 5 mM EDTA, 10 mM
Na4P2O7, 100 mM NaF, 2 mM
Na3VO4, 1% NP-40 (all from Sigma-Aldrich), 1
mM phenylmethylsulfonyl fluoride, 10 mg/ml aprotinin and 10 mg/ml
leupeptin (ICN Biomedicals, Asse-Relegem, Belgium). The proteins
were separated by 12% SDS-PAGE and then transferred onto
nitrocellulose membranes (EMD Millipore) for immunoblot analysis.
The membranes were incubated with rabbit polyclonal antibodies
against CDK4 (IgG; catalog no. sc-260; Santa Cruz Biotechnology,
Inc., Dallas, TX, USA), cyclin D1 (IgG; catalog no. 2407-0508;
Bio-Rad AbD Serotec, Ltd., Kidlington, UK), p27 (IgG; catalog no.
sc-528; Santa Cruz Biotechnology, Inc.), cyclin E (catalog no.
2925; Cell Signaling Technology, Inc., Danvers, MA, USA) and CDK2
(IgG; catalog no. sc-163; Santa Cruz Biotechnology, Inc.), with a
goat polyclonal IgG anti-β-actin antibody (catalog no. sc-1615;
Santa Cruz Biotechnology, Inc.) used as the internal control, at
room temperature for 3 h. All primary antibodies were used at a
dilution of 1:1,000. The membranes were then incubated with goat
anti-rabbit IgG (catalog no. sc-2004; Santa Cruz Biotechnology,
Inc.) or donkey anti-goat IgG (catalog no. sc-2020; Santa Cruz
Biotechnology, Inc.) secondary antibodies conjugated with
horseradish peroxidase, which were used at a dilution of 1:5,000,
at room temperature for 1 h. The bands were visualized with a
ChemiDoc™ MP imaging system (Bio-Rad Laboratories, Inc., Hercules,
CA, USA) and quantified using Image Lab™ software version 3.0
(Bio-Rad Laboratories, Inc.).
Cell migration analysis
Confluent B16 melanoma cells were maintained in
serum-free medium for 24 h and a wound was formed using a razor
blade. The cells were washed with PBS prior to the application of
control medium or 10X ASC-CM. Images of the wounded area were
captured at 12 and 24 h using phase-contrast microscopy (Eclipse
TS100, Nikon Instruments, Inc., Melville, NY, USA). The wound
closure was estimated by marking four randomly selected points
along each wound and measuring the horizontal distance of the
migrating cells from the initial wound.
Animals and xenograft model
Six-week-old C57BL/6 male mice (n=20) were obtained
from Orient Bio, Inc. (Seongnam, Korea) and acclimatized for one
week; the animals were housed in an air-conditioned room at 25°C
with a 12 h dark/light cycle. All animals received human care with
unlimited access to mouse chow and water. Animal experimental
procedures were conducted in accordance with the guidelines of the
INHA Institutional Animal Care and Use Committee of the Medical
School of Inha University (Incheon, Korea). Mice were randomly
divided into four groups (n=5; two control and two ASC-CM groups)
and B16 melanoma cells (1×106) were inoculated into the
dorsal flank of each mouse. When the tumors had reached a volume of
~50–100 mm3, the mice were administered with a single
intratumoral injection of 10X-concentrated ASC-CM (100 µl) or
vehicle (PBS; control group), daily or every other day (n=5), a
total of five times. Tumor axes were measured using calipers and
tumor volume was calculated using the following equation: Volume =
0.5 × long axis × (short axis)2. After 20 days, the mice
were sacrificed by anaesthetizing with ketamine and rompun mixture,
and their tumors were excised.
Immunohistochemistry
Tumor specimens were fixed with 10% buffered
formaldehyde and embedded in paraffin (Sigma-Aldrich).
Immunohistochemical staining was performed using 8 µm-thick
sections of the tumor samples following deparaffinization, as
previously described (18). Primary
antibodies against Ki-67 (Abcam, Cambridge, UK) and CD34 (Santa
Cruz Biotechnology, Inc., Santa Cruz, CA, USA) were applied.
Unbound primary antibodies were washed off and the sections were
incubated with a secondary biotinylated immunoglobulin G antibody.
The stained cells were visualized using avidin-biotin complex (ABC)
solution followed by incubation with a 3,3-diaminobenzidine
tetrahydrochloride solution using a Vectastain Elite ABC kit
(Vector Laboratories, Inc., Burlingame, CA, USA).
Statistical analysis
Statistical analyses were performed using an
unpaired Student's t-test. All data are presented as the mean ±
standard deviation and P<0.05 was considered to indicate a
statistically significant value.
Results
ASC-CM suppresses the proliferation of
B16 melanoma cells
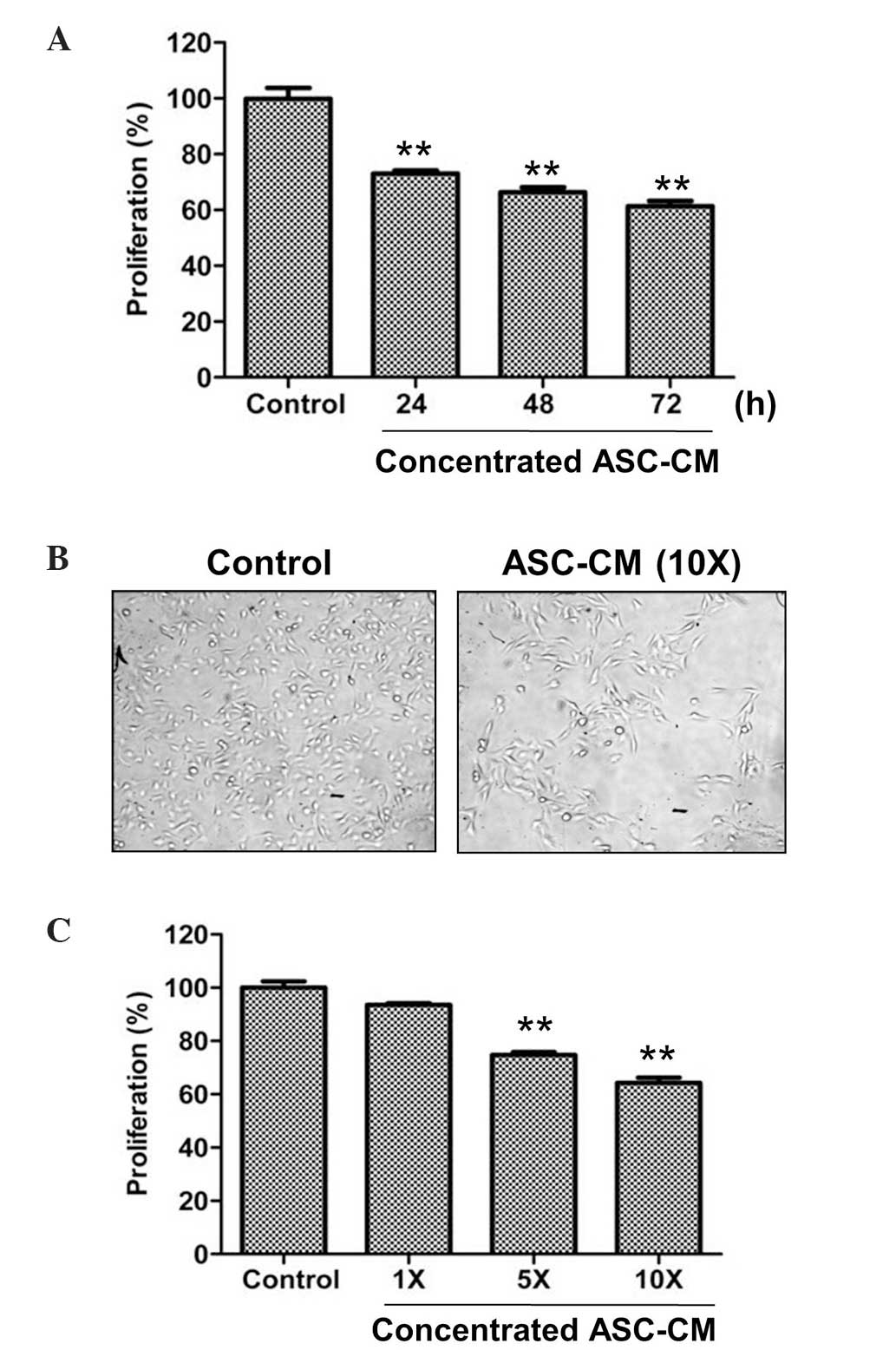
The present study initially investigated the effect
of ASC-CM on the proliferation of B16 melanoma cells in
vitro using a CCK-8 assay. Incubation of the B16 melanoma cells
with 10X ASC-CM significantly suppressed cell proliferation in a
time-dependent manner, compared with the control cells incubated in
basal medium without ASC-CM (P<0.01; Fig. 1A). Following a 72-h incubation period,
cells treated with ASC-CM exhibited a ~40% reduction in viability.
In agreement with the CCK-8 findings, microscopic analysis
identified retarded cell growth in 10X ASC-CM-treated cells
compared with that of the control (Fig.
1B). Furthermore, the anti-proliferative effect of ASC-CM on
melanoma cells was clarified by incubation with various
concentrations of ASC-CM (1, 5 and 10X), demonstrating a
significant dose-dependent decrease in proliferation (P<0.05;
Fig. 1C). These data indicated that
culture medium from ASCs may effectively suppress the proliferation
of B16 melanoma cells.
ASC-CM-induces cell cycle arrest in
B16 melanoma cells at G1 phase without induction of cell death
To elucidate the mechanism by which ASC-CM exerts
its anti-proliferative effect, the present study analyzed the cell
cycle of B16 melanoma cells following incubation with 10X ASC-CM
for 24 h. ASC-CM treatment significantly increased the number of
cells in the G1/S phase by ~2.5-fold (P<0.01), while reducing
the proportion of cells in the S and G2/M phases. These data
indicated that the anti-proliferative effect of ASC-CM is induced
by G1 phase arrest (Fig. 2A and B).
Furthermore, no detectable difference in the proportion of cells in
the sub-G1 phase was observed, indicating that ASC-CM-induced cell
growth retardation is not associated with the induction of cell
death. In addition, western blot analysis demonstrated that ASC-CM
treatment downregulated cyclin D1 protein expression, which
promotes G1/S transition, without detectable changes in the
expression of cyclin E or p27, a CDK inhibitor (Fig. 2C). Thus, ASC-CM appears to reduce the
proliferation of B16 melanoma cells by inducing G1 arrest via
modulation of cell cycle regulators.
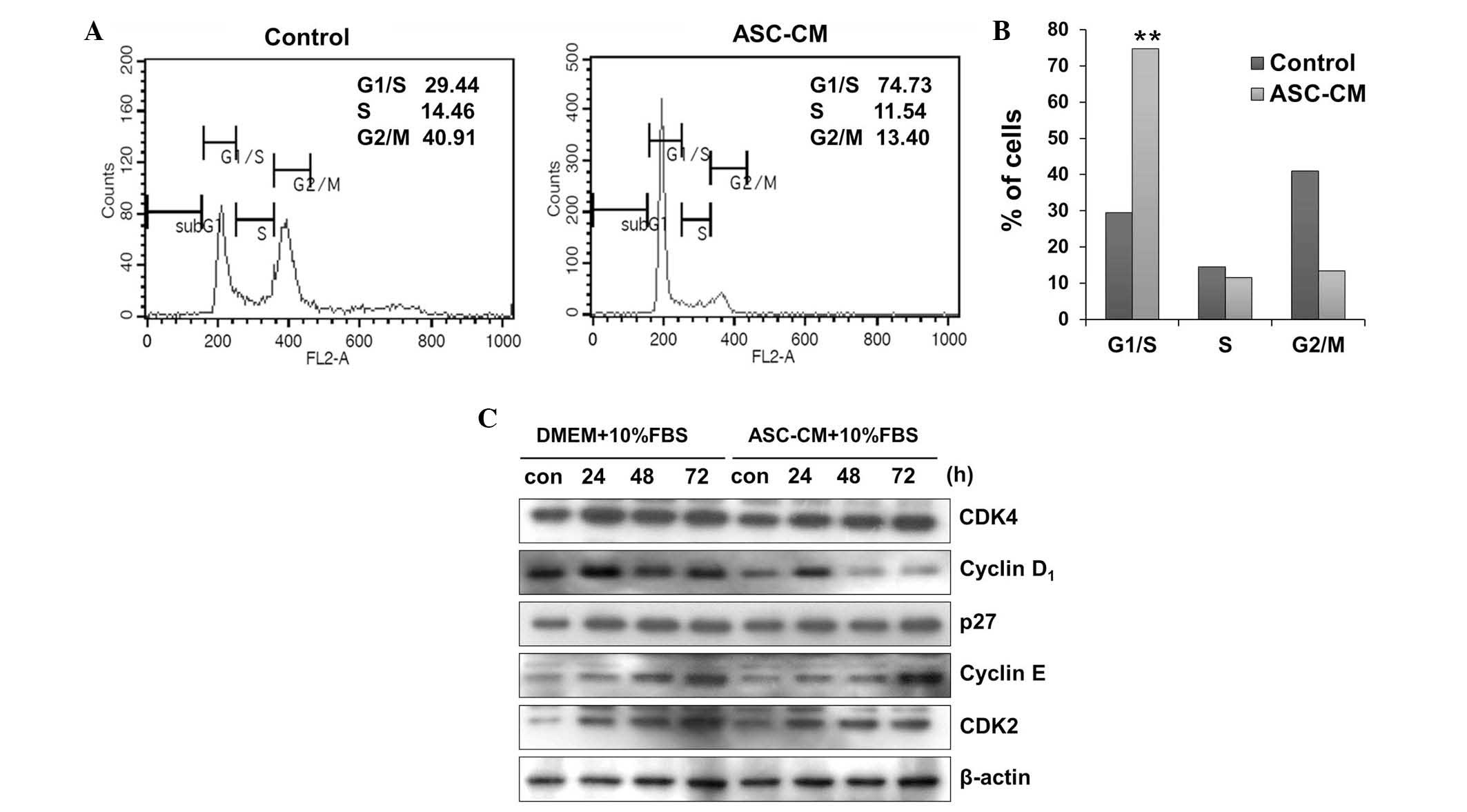 | Figure 2.ASC-CM induces cell cycle arrest in
B16 cells at G1 phase. Cell cycle analysis of B16 melanoma cells
incubated with ASC-CM. (A) Cells were incubated with 10X ASC-CM for
24 h and stained with propidium iodide. Cell cycle distribution was
analyzed by flow cytometry. (B) Relative number of cells in the
G1/S, S, and G2/M phases were compared between the control group
and the ASC-CM-treated cells. A representative result from three
independent experiments is shown. **P<0.01 vs. control. (C)
Cells were treated with or without ASC-CM (10X) and the protein
expression levels of key cell cycle regulatory proteins, including
cyclin D1, p27, cyclin E and CDK2, were determined by performing an
immunoblot analysis. β-actin was used as the loading control.
ASC-CM, adipose-derived stem cell-conditioned medium; DMEM,
Dulbecco's modified Eagle's medium; FBS, fetal bovine serum; con,
control; CDK, cyclin-dependent kinase. |
ASC-CM decreases the migration of B16
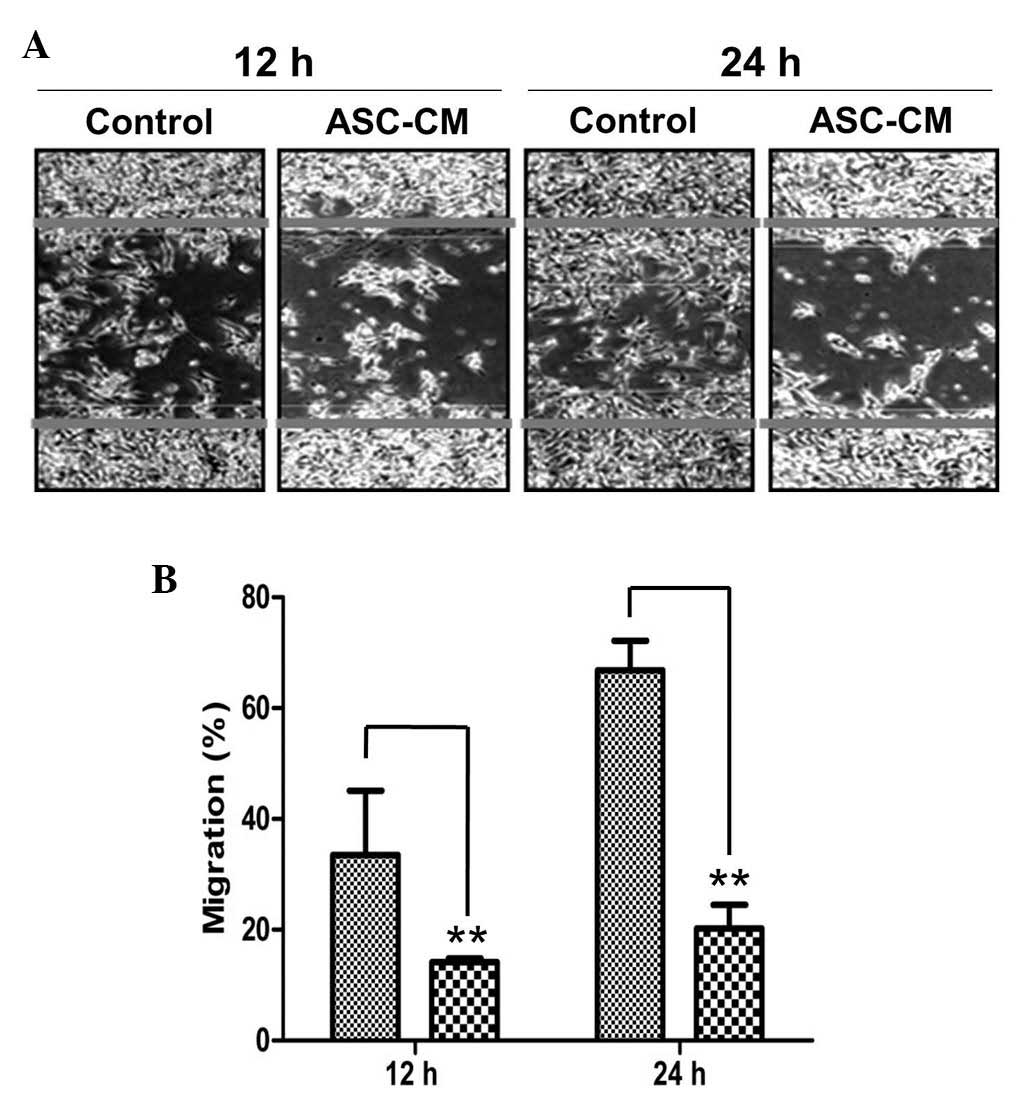
melanoma cells
In addition to facilitating normal embryonic
development, melanocyte migration is a critical step in the
malignant transformation of metastatic melanomas. To determine
whether ASC-CM was able to regulate the migration of B16 melanoma
cells, a wound migration assay was conducted in B16 cells incubated
with or without ASC-CM. Following wounding, the melanocytes were
incubated for 12 or 24 h in medium containing ASC-CM. Subsequent
phase contrast microscopy revealed that migration was markedly
reduced in the ASC-CM-treated cells (Fig.
3A). Subsequently, migration was quantified by measuring the
horizontal distance of the migrating cells from the initial wound.
Under physiological conditions, cell migration was calculated as
~30 and ~65% following incubation for 12 and 24 h, respectively.
However, migration was significantly retarded in the ASC-CM-treated
cells, resulting in <20% migration following 12 and 24 h of
incubation(P<0.01; Fig. 3B). The
results of the present study indicate that, as well as reducing
cell proliferation, ASC-CM is able to effectively suppress the
migration of B16 melanoma cells.
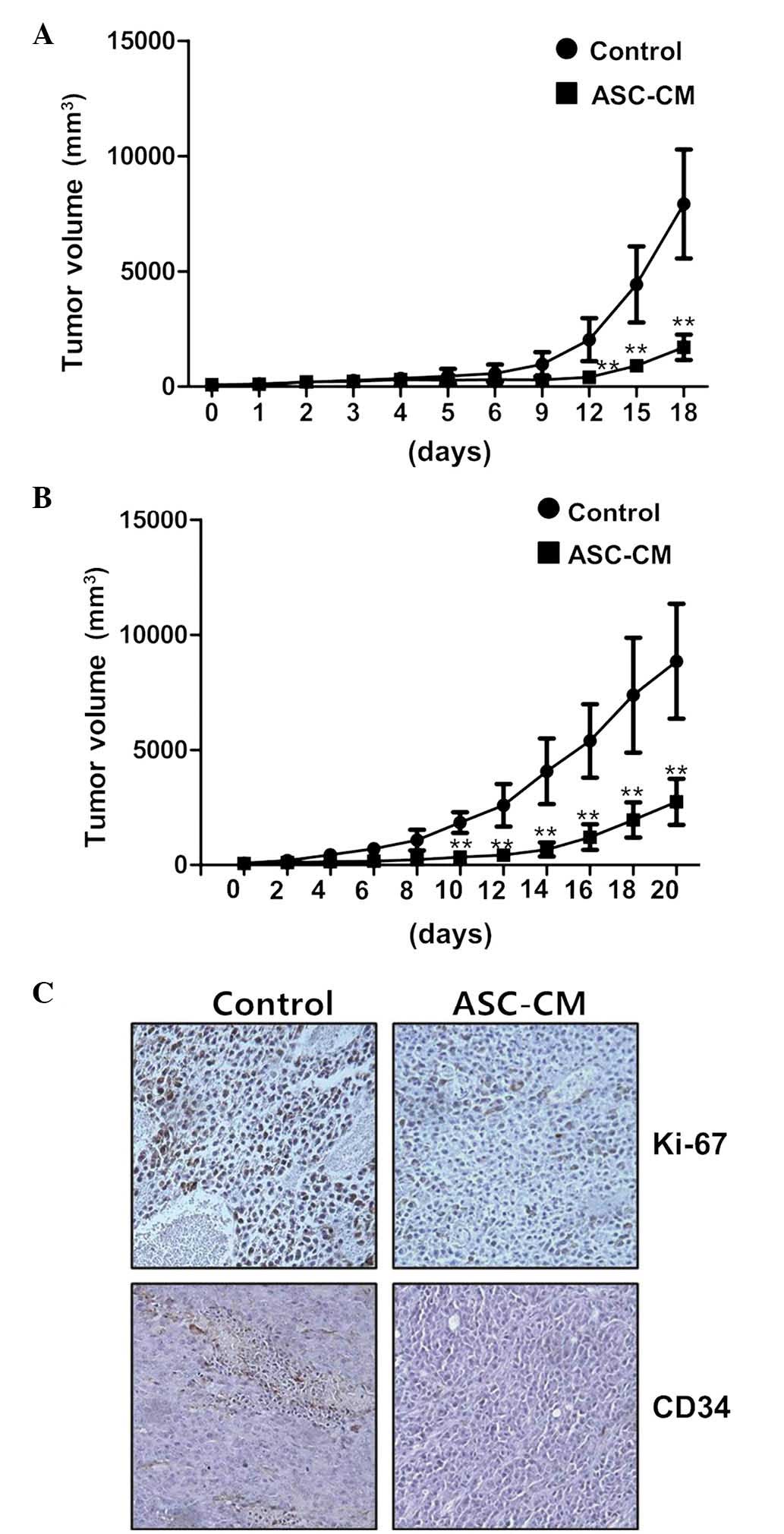
ASC-CM suppresses B16 melanoma cell
tumor growth in a mouse xenograft model
Considering the aforementioned in vitro
findings, the effects of ASC-CM were investigated in an in
vivo mouse transplantation model. Following subcutaneous
injection of B16 melanoma cells (1×106 cells) into the
right flank of C57BL/6 mice and the growth of a tumor mass to a
volume of ~50–100 mm3, 100 µl of 10X ASC-CM was
administered intratumorally, daily or every other day, a total of
five times. By measuring the tumor volume, it was identified that
tumor growth was significantly retarded in the ASC-CM-treated
groups compared with that of the control group following treatment
daily or every other day, exhibiting a 71.4 and 73.7% reduction in
size at day 18, respectively (P<0.01; n=5 per group; Fig. 4A and B). In addition,
immunohistochemistry revealed that the numbers of actively
proliferating Ki-67 positive cells and CD34-positive endothelial
cells were markedly decreased in the ASC-CM-treated group,
indicating that ASC-CM exerts anti-tumorigenic activity in
vivo (Fig. 4C). Collectively,
these in vivo transplantation data support the hypothesis of
a role for ASC-CM in the efficient inhibition of tumor growth in
mice, as well as in an in vitro cell culture model.
Discussion
Adipose tissue consists of populations of
heterogeneous cell types, including ASCs, endothelial cells,
preadipocytes, pericytes, hematopoietic-lineage cells and
fibroblasts, termed the SVF (19).
ASCs are able to differentiate into mesodermal cell types (1), neuronal-like cells of ectodermal lineage
(20–23) and cells of epidermal lineage (24), demonstrating their multi-lineage
potential. This has resulted in the use of ASCs an alternative to
pluripotent ESCs in the laboratory and for clinical use. A
promising area for ASC therapeutics is in the regeneration of
various tissues, including cardiovascular, bone, cartilage, nerve
and liver tissues.
Previously, it was demonstrated that ASCs and ASC-CM
have skin-regenerative potential (4,17,25–28).
Studies identified that direct ASC transplantation or the
application of ASC-CM resulted in a significant acceleration in
wound-healing, the promotion of hair regeneration, a reduction in
undesirable pigmentation and anti-wrinkle effects (4,17,25–28). These
skin-regenerative results arise primarily from paracrine effects,
including anti-apoptotic effects and angiogenesis of ASC secretory
factors, in addition to their building-block function. Furthermore,
ASCs and their soluble factors have been reported to increase the
proliferation and migration of normal keratinocytes and fibroblasts
(4,17). Notably, ASC-CM administration appeared
to inhibit melanin synthesis in B16 melanoma cells by
downregulating tyrosinase and tyrosinase-related proteins (27). However, the effects of ASC-CM on cell
proliferation and migratory ability have yet to be sufficiently
addressed. The present study aimed to investigate whether ASC-CM
treatment affected the proliferation and migration of B16 melanoma
cells and evaluate the ability of ASC-CM to regulate a B16 melanoma
cell tumor mass in a mouse xenograft model.
To date, only a small number of studies have
described the molecular mechanisms underlying the tumorigenesis of
melanoma. Linkage analysis on melanoma families revealed that the
development of melanoma may be initiated by loss-of-function
mutations in the two tumor suppressor proteins, p14ARF
and p16INK4a, alternative splicing products coded by the
CDKN2A gene (29). p14ARF
stabilizes p53 by rapidly degrading mouse/human double minute-2
(30,31). By contrast, p16INK4a
enhances binding of RB protein with transcription factors of the
E2F family by suppressing CDK4/6 activity (15). Mutations in p14ARF and
p16INK4a proteins may promote G1-S phase transition, as
well as re-entry into the cell cycle, thus, potentiating abnormal
cell proliferation (16). In
addition, BRAF gene mutations have recently been identified to
indicate a predisposition to melanoma. Furthermore, the
mitogen-activated protein kinase and phosphatidylinositide 3-kinase
signaling pathways are also activated in the transcription of
critical cell proliferation genes in the majority of cases of
melanoma (32). In the present study,
it was identified that treatment with concentrated ASC-CM inhibited
the proliferation of B16 melanoma cells by ~40% and that this
anti-proliferative effect was induced by cell cycle arrest at the
G1 phase, as revealed by flow cytometry. This cell cycle arresting
effect of ASC-CM was clarified by the detection of decreased cyclin
D1 expression levels, indicating that there may be merit in the
development of ASC-CM into a therapeutic agent in the future. The
present study also performed a wound migration assay, which
indicated that the migration of melanocytes was inhibited following
treatment with ASC-CM, indicating that ASC-CM exhibited an
anti-tumorigenic effect on the transformed melanocytes. In addition
to these in vitro evidences, ASC-CM was demonstrated to
decrease tumor growth in a mouse model, indicating that ASCs or
ASC-CM may be promising candidates for future therapeutic
strategies targeting melanoma.
ASCs produce various growth factors and adipokines,
thus, the cells function as an endocrine or paracrine organ that is
considered to perform a pivotal role in maintaining a suitable
microenvironment for the regulation of tumor initiation,
progression and metastasis (27,33–35). ASCs
express a number of biological proteins involved in the promotion
of cell viability, including transforming growth factor β 1
(27,33) and progranulin (34). However, numerous studies have
demonstrated the anti-proliferative or anti-tumorigenic effects of
ASCs. For example, preconditioned endothelial progenitor cells have
been observed to reduce melanoma metastasis by cell-cell
interactions induced by secreted protein, acidic and rich in
cysteine, as well as endocytosis (9).
Human ESCs have been reported to secrete soluble factors that
arrest the growth of human epithelial cancer cell lines (10). For example, angiopoietin-1 produced
from ASCs appears to exhibit an inhibitory effect on neointimal
formation in the rat femoral artery (36). However, the profile of cytokines
released from ASCs may be modulated by external factors; for
example, hepatocyte growth factor was released from ASCs following
an exposure to basic fibroblast growth factor or epidermal growth
factor (EGF), while ASCs secreted various hematopoietic factors and
proinflammatory cytokines following lipopolysaccharide treatment
(37). These results indicate that
environmental factors are crucial for determination of the proteins
that are released from ASCs. Furthermore, a recent study
demonstrated that varying expression levels of protein factors are
released in accordance with ASC subtype. Specifically, monocyte
chemotactic protein-1, eotaxin, IL-1 receptor agonist, IL-6,
granulocyte-macrophage colony-stimulating factor and vascular EGF
were more abundant in conditioned medium obtained from the visceral
omental ASC fraction compared with that of the abdominal
subcutaneous ASC fraction, indicating heterogeneity in the
secretory ability of visceral and subcutaneous fat tissue (35). These findings indicate that the set of
factors secreted from ASCs may be determined by the cell origin and
the environmental factors during the culturing process.
Controversy remains surrounding the interpretation
of studies exploring the induction versus inhibition of cancer
following stem cell transplantation, therefore, hindering their
clinical use. Despite the controversial interpretations regarding
their effects on various types of cancer, freshly isolated ASCs and
cosmetics using ASC-CM are commercially available and are currently
undergoing clinical trials in certain countries.
In conclusion, the present study shows that ASC-CM
attenuates the proliferation and migration of B16 melanoma cells in
culture and in a mouse xenograft model, suggesting ASC-CM may be a
useful material for anti-cancer therapeutics. Further knowledge
must be acquired regarding the safety of ASCs and ASC-CM prior to
their use in future clinical applications.
Acknowledgements
The present study was supported by grants from the
Korean Health Technology Research and Development Project of the
Korean Ministry of Health and Welfare (grant no. A120266) and from
Inha University.
References
|
1
|
Zuk PA, Zhu M, Mizuno H, et al:
Multilineage cells from human adipose tissue: Implications for
cell-based therapies. Tissue Eng. 7:211–228. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hokugo A, Sorice S, Parhami F, et al: A
novel oxysterol promotes bone regeneration in rabbit cranial bone
defects. J Tissue Eng Regen Med. Aug 29–2013.(Epub ahead of print).
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fu X, Fang L, Li H, et al: Adipose tissue
extract enhances skin wound healing. Wound Repair Regen.
15:540–548. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kim WS, Park BS, Sung JH, et al: Wound
healing effect of adipose-derived stem cells: A critical role of
secretory factors on human dermal fibroblasts. J Dermatol Sci.
48:15–24. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yu JM, Jun ES, Bae YC and Jung JS:
Mesenchymal stem cells derived from human adipose tissues favor
tumor cell growth in vivo. Stem Cells Dev. 17:463–473. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Prantl L, Muehlberg F, Navone NM, et al:
Adipose tissue-derived stem cells promote prostate tumor growth.
Prostate. 70:1709–1715. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Eterno V, Zambelli A, Pavesi L, et al:
Adipose-derived Mesenchymal Stem Cells (ASCs) may favour breast
cancer recurrence via HGF/c-Met signaling. Oncotarget. 5:613–633.
2014.PubMed/NCBI
|
|
8
|
Koellensperger E, Gramley F, Preisner F,
Leimer U, Germann G and Dexheimer V: Alterations of gene expression
and protein synthesis in co-cultured adipose tissue-derived stem
cells and squamous cell-carcinoma cells: Consequences for clinical
applications. Stem Cell Res Ther. 5:652014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rowan BG, Gimble JM, Sheng M, et al: Human
adipose tissue-derived stromal/stem cells promote migration and
early metastasis of triple negative breast cancer xenografts. PLoS
One. 9:e895952014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sun B, Roh KH, Park JR, et al: Therapeutic
potential of mesenchymal stromal cells in a mouse breast cancer
metastasis model. Cytotherapy. 11:289–298. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cousin B, Ravet E, Poglio S, et al: Adult
stromal cells derived from human adipose tissue provoke pancreatic
cancer cell death both in vitro and in vivo. PLoS One. 4:e62782009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Takahara K, Ii M, Inamoto T, et al:
Adipose-derived stromal cells inhibit prostate cancer cell
proliferation inducing apoptosis. Biochem Biophys Res Commun.
446:1102–1107. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
National Cancer Institute, . Surveillance,
Epidemiology, and End Results Program. SEER Stat Fact Sheets:
Melanoma of the Skin. http://seer.cancer.gov/statfacts/html/melan.htmlApril
25–2015
|
|
14
|
Squires MH III and Delman KA: Current
treatment of locoregional recurrence of melanoma. Curr Oncol Rep.
15:465–472. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Koh J, Enders GH, Dynlacht BD and Harlow
E: Tumour-derived p16 alleles encoding proteins defective in
cell-cycle inhibition. Nature. 375:506–510. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sheppard KE and McArthur GA: The
cell-cycle regulator CDK4: an emerging therapeutic target in
melanoma. Clin Cancer Res. 19:5320–5328. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Won CH, Yoo HG, Kwon OS, et al: Hair
growth promoting effects of adipose tissue-derived stem cells. J
Dermatol Sci. 57:134–137. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ma YL, Peng JY, Zhang P, Liu WJ, Huang L
and Qin HL: Immunohistochemical analysis revealed CD34 and Ki67
protein expression as significant prognostic factors in colorectal
cancer. Med Oncol. 27:304–309. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kokai LE, Marra K and Rubin JP: Adipose
stem cells: Biology and clinical applications for tissue repair and
regeneration. Transl Res. 163:399–408. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Safford KM, Safford SD, Gimble JM, Shetty
AK and Rice HE: Characterization of neuronal/glial differentiation
of murine adipose-derived adult stromal cells. Exp Neurol.
187:319–328. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ashjian PH, Elbarbary AS, Edmonds B, et
al: In vitro differentiation of human processed lipoaspirate cells
into early neural progenitors. Plast Reconstr Surg. 111:1922–1931.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kingham PJ, Kalbermatten DF, Mahay D,
Armstrong SJ, Wiberg M and Terenghi G: Adipose-derived stem cells
differentiate into a Schwann cell phenotype and promote neurite
outgrowth in vitro. Exp Neurol. 207:267–274. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xu Y, Liu L, Li Y, et al: Myelin-forming
ability of Schwann cell-like cells induced from rat adipose-derived
stem cells in vitro. Brain Res. 1239:49–55. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Trottier V, Marceau-Fortier G, Germain L,
Vincent C and Fradette J: IFATS collection: Using human
adipose-derived stem/stromal cells for the production of new skin
substitutes. Stem Cells. 26:2713–2723. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim WS, Park BS, Kim HK, et al: Evidence
supporting antioxidant action of adipose-derived stem cells:
Protection of human dermal fibroblasts from oxidative stress. J
Dermatol Sci. 49:133–142. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kim WS, Park BS, Park SH, Kim HK and Sung
JH: Antiwrinkle effect of adipose-derived stem cell: Activation of
dermal fibroblast by secretory factors. J Dermatol Sci. 53:96–102.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kim WS, Park SH, Ahn SJ, et al: Whitening
effect of adipose-derived stem cells: A critical role of TGF-beta
1. Biol Pharm Bull. 31:606–610. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kim WS, Park BS and Sung JH: The
wound-healing and antioxidant effects of adipose-derived stem
cells. Expert Opin Biol Ther. 9:879–887. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tsao H, Chin L, Garraway LA and Fisher DE:
Melanoma: From mutations to medicine. Genes Dev. 26:1131–1155.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kamijo T, Weber JD, Zambetti G, Zindy F,
Roussel MF and Sherr CJ: Functional and physical interactions of
the ARF tumor suppressor with p53 and Mdm2. Proc Natl Acad Sci USA.
95:8292–8297. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang Y, Xiong Y and Yarbrough WG: ARF
promotes MDM2 degradation and stabilizes p53: ARF-INK4a locus
deletion impairs both the Rb and p53 tumor suppression pathways.
Cell. 92:725–734. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Eggermont AM, Spatz A and Robert C:
Cutaneous melanoma. Lancet. 383:816–827. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kim BS, Kang KS and Kang SK: Soluble
factors from ASCs effectively direct control of chondrogenic fate.
Cell Prolif. 43:249–261. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tsuruma K, Yamauchi M, Sugitani S, et al:
Progranulin, a major secreted protein of mouse adipose-derived stem
cells, inhibits light-induced retinal degeneration. Stem Cells
Transl Med. 3:42–53. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Perrini S, Ficarella R, Picardi E, et al:
Differences in gene expression and cytokine release profiles
highlight the heterogeneity of distinct subsets of adipose
tissue-derived stem cells in the subcutaneous and visceral adipose
tissue in humans. PLoS One. 8:e578922013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Takahashi M, Suzuki E, Kumano S, et al:
Angiopoietin-1 mediates adipose tissue-derived stem cell-induced
inhibition of neointimal formation in rat femoral artery. Circ J.
77:1574–1584. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kilroy GE, Foster SJ, Wu X, et al:
Cytokine profile of human adipose-derived stem cells: Expression of
angiogenic, hematopoietic, and pro-inflammatory factors. J Cell
Physiol. 212:702–709. 2007. View Article : Google Scholar : PubMed/NCBI
|