Introduction
Lymphoma is one of the most common types of
malignant tumors in children and the incidence of this disease is
increasing (1). Non-Hodgkin's
lymphoma accounts for ~80–85% of the total number of lymphoma
cases, and exhibits a high degree of malignancy, rapid progression
and a poor prognosis in children (2).
A novel strategy for the treatment of lymphoma, is the induction of
apoptosis of lymphoma cells. However, the majority of currently
available apoptosis inducers contain large quantities of heavy
metals and thus produce a number of intolerable side effects
(3,4).
Therefore, the identification of novel inducers of apoptosis in
lymphoma cells, which exhibit higher efficiency and lower toxicity,
is required.
Embelin is a small molecule inhibitor of X-linked
inhibitor of apoptosis (XIAP), which is used in traditional Chinese
medicine (5–7). Previous studies have shown that Embelin
inhibits proliferation, promotes apoptosis, increases sensitivity
to chemotherapeutic agents and reduces drug resistance in tumor
cells (8–12). Therefore, Embelin has been identified
as a promising antitumor agent. However, to date there have been
few studies conducted, examining the effect of Embelin on acute T
cell lymphoma. The present study investigated the inhibitory effect
of Embelin on human acute T cell lymphoma Jurkat cells, in addition
to the mechanisms underlying this effect.
Materials and methods
Cell culture
Human acute T cell lymphoma Jurkat cells (Chinese
Academy of Medical Sciences, Beijing, China) were cultured in RPMI
1640 medium (Applied Biosystems China Ltd., Beijing, China),
containing 100 U/ml penicillin and 100 µg/ml streptomycin (Life
Technologies, Grand Island, NY, USA) at 37°C with 5%
CO2. Cells were treated with 0.25% trypsin (Applied
Biosystems China Ltd.,) for passage, and cells in the logarithmic
growth phase were selected for subsequent experiments. Embelin was
purchased from Sigma-Aldrich (St. Louis, MO, USA). Caspase
inhibitors, z-DEVD-fmk and Ac-LEHD-CHO, were obtained from Promega
Corporation (Madison, WI, USA).
MTS/PMS method
The MTS/PMS method was used to evaluate the
inhibition rate of tumor cell proliferation. Jurkat cells were
divided into three groups that were treated with various
concentrations of Embelin (5, 10 or 20 mM; Hebei Medical
University, Haisen Pharmaceutical Co., Ltd., Hebei, China). The
control group was incubated with an equal quantity of RPMI 1640
medium. RPMI 1640 medium with no cells was used as a blank control
group, and the cell viability of each sample was measured 3 times,
and the mean of the 3 values was calculated. Jurkat cells in the
logarithmic growth phase were cultured in RPMI 1640 medium
containing 10% fetal bovine serum (Hangzhou Sijiqing Biological
Engineering Materials Co., Ltd., Zhejiang, China). The cell
concentration was adjusted to 1×105/ml, and cells were
inoculated into a 96-well culture plate; each well containing 90 µl
of culture medium. Plates were incubated at 37°C with 5%
CO2. Following incubation for 24 h, 10 µl Embelin at
various concentrations (5, 10 or 20 mmol/ml) was added to the
appropriate wells. Culture medium containing 0.01% dimethyl
sulfoxide (DMSO; Tianjin Yongda Chemical Reagents Development
Center, Tianjin, China) was added to the control group for the MTS
assay.
In subsequent experiments, the caspase-3 inhibitor,
z-DEVD-fmk, and the caspase-9 inhibitor, Ac-LEHD-CHO, were added to
cells, to achieve a final concentration of 10 µl. The different
concentrations of Embelin were added 1 h later, and cells were then
cultured for 24 h. The MTS/PMS mixture (20 µl) was added to the
cells, and plates were cultured for a further 3–4 h. Morphological
changes were observed under an inverted microscope (Olympus
Corporation, Tokyo, Japan) under 40X magnification. A fully
automated enzyme mark measuring instrument (Titertek Multiskan,
North Ryde, Australia) was used to detect the absorbance of the
samples at 570 nm wavelength. Cell viability was calculated as
follows: Viability (%) = [experimental group (OD) - blank group
(OD)]/[control group (OD) - blank group (OD)] × 100.
Phosphatidylserine ectropion [Annexin
V/propidium iodide (PI)] analysis
Phosphatidylserine ectropion (Annexin V/PI) analysis
were used to analyze rates of cell apoptosis. Culture medium (2 ml)
with various concentrations of Embelin (5, 10 or 20 mM) were added
to Jurkat cells in the logarithmic growth phase as the drug
treatment groups. Complete culture medium was used for the control
group. After 24 h, the cells were collected, centrifuged at 850 ×
g for 5 min and then washed three times with phosphate
buffer solution. Centrifugation (5 min) was performed following
each wash. Cells were resuspended in 2 ml of phosphate buffer
solution for 5 min. Subsequently, 5 µl Annexin V-FITC (Lianke
Biological Engineering Co., Ltd., Zhejiang, China) and 10 µl
propidium iodide (Lianke Biological Engineering Co., Ltd.) were
added, and samples were incubated for 10 min at room temperature.
Cells were stained in darkness at 4°C for 30 min, and flow
cytometry (Beckman Coulter, Inc., USA) was then used to analyze the
rate of apoptosis. In the results of the apoptosis analysis, the
left upper, right upper, left lower and right lower quadrants
represent necrotic cells, late apoptotic cells, normal cells and
early apoptotic cells, respectively. The right lower quadrant was
selected for evaluation of the levels of apoptosis in Jurkat
cells.
Western blotting
Western blotting was performed as described
previously (13). Briefly, cells
treated with 5, 10, or 20 mM Embelin for 24 h were lysed, and
protein concentrations were determined using a Lowry protein assay
kit (Sigma Corporation of America, New York, NY, USA). Protein
samples (50 µg) were separated using SDS-PAGE (Millipore
Corporation, Billerica, MA, USA), transferred to a PVDF membrane,
blocked in 5% dried non-fat milk for 1 h at 25°C, and then
incubated with primary rabbit polyclonal antibodies to XIAP, (cat.
no, sc-11426; 1:1,000), and Caspase 3, 8 and 9 (cat. no. sc-7148;
1:500), and mouse monoclonal antibody to PARP (cat. no. sc-56196;
1:1,000) were purchased from Santa Cruz Biotechnology, Inc.(Dallas,
TX, USA) and primary mouse monoclonal antibody to GADPH (cat. no.
2118S; 1:10,000) and primary rabbit polyclonal antibodies to Bcl-2
(cat. no. 2870P; 1:1,000), Bcl-XL (cat. no. 2764P; 1:1,000) and Bax
(cat. no. 2774S, 1:1,000) were purchased from Cell Signaling
Technology, Inc. (CST, CA, USA) overnight at 4°C. Membranes were
washed 3 times for 5 min with TBST and incubated for 1 h with
fluorochrome-labeled secondary antibodies against rabbit or mouse
(1:10,000; IRDye 800-LI-COR for rabbit antibodies or IRDye
700-LI-COR for mouse antibodies; LI-COR Biosciences, Ltd.,
Cambridge, UK). Following 4 washes with TBST, expression of the
target proteins were analyzed by imaging the membrane with a LI-COR
Odyssey infrared imager (LI-COR Biosciences, Ltd.). GAPDH was used
as an internal control.
Statistical analysis
Data are reported as the mean ± standard error of
the mean. A one-way analysis of variance (ANOVA) was performed to
determine the significance between groups. Tukey's method was used
for multiple comparisons. P<0.05 was considered to indicate a
statistically significant difference. SPSS software, version 13.0
(SPSS, Inc., Chicago, IL, USA) was used for data analysis.
Results
Effect of Embelin on the proliferation
of Jurkat cells
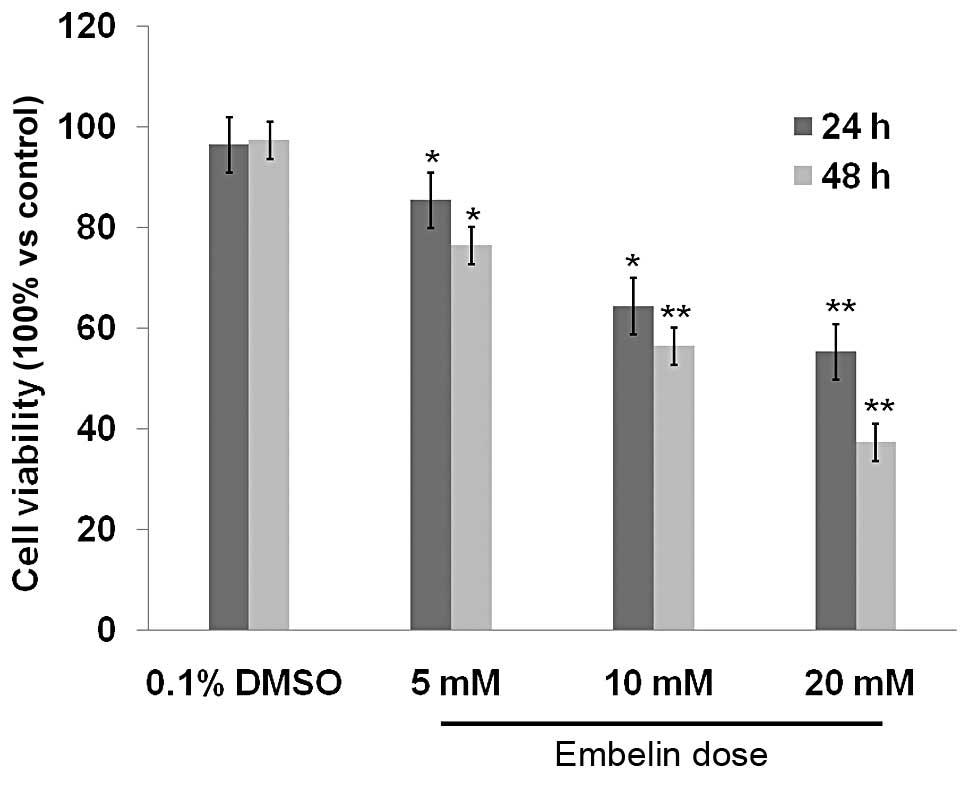
The results of the MTS assay showed that all
concentrations of Embelin used in the present study, significantly
inhibited the proliferation of Jurkat T cell lymphoma cells in a
dose- and time-dependent manner, compared with proliferation of
cells in the control group. As shown in Fig. 1, following treatment with 5, 10 and 20
mM for 48 h, cell viability was ~82.31, 58.65 and 37.62%
respectively, which was significantly reduced compared with that in
the control group and the 0.1% DMSO control group (P<0.01). It
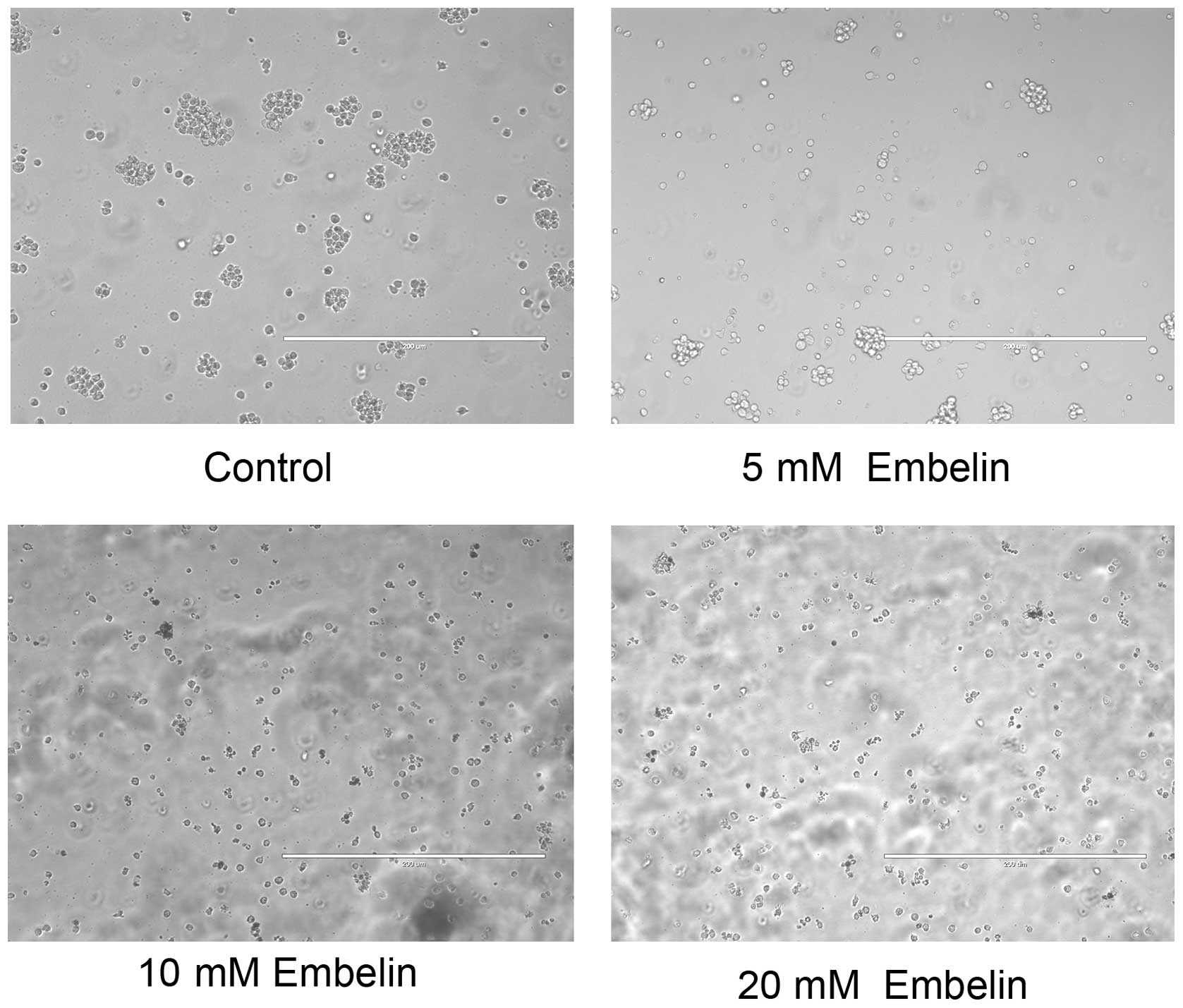
was also shown that the number of viable cells visible under the
microscope was significantly reduced (Fig. 2). These results demonstrated the
potency of Embelin in inhibiting the growth of T cell lymphoma
cancer cells in vitro.
Effect of Embelin on the rate of
apoptosis in Jurkat cells
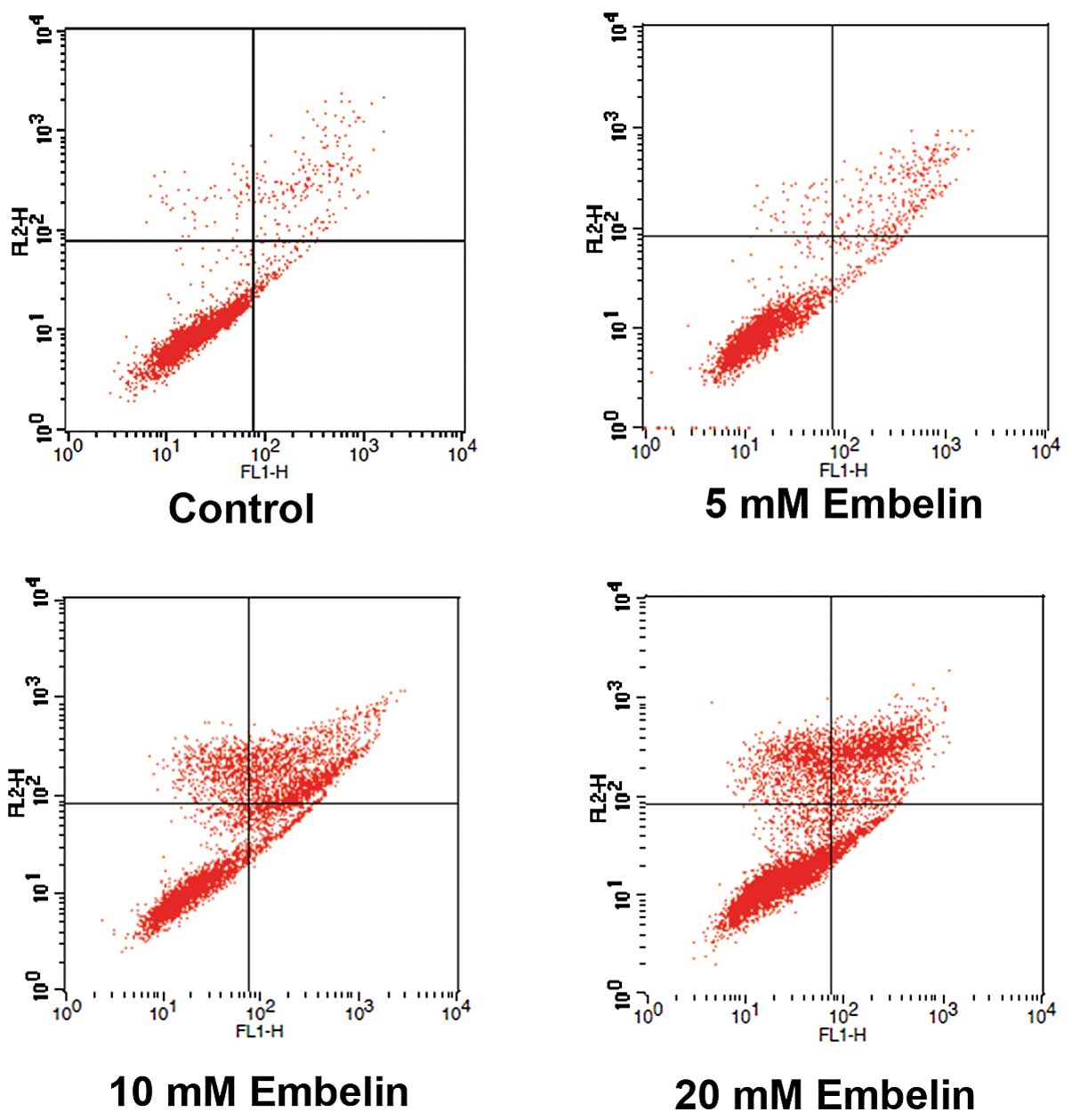
Induction of apoptosis is the primary mechanism of
action of Embelin. Therefore the mechanism underlying the induction
of cell death by Embelin was examined using flow cytometry. The
results showed that all concentrations of Embelin used,
significantly increased the rates of apoptosis of Jurkat T cell
lymphoma cells, compared with rates in the control group
(P<0.05). The percentage of apoptotic cells following treatment
with 5, 10 and 20 mM Embelin, was ~9.65, 32.45 and 40.79%
respectively. An association between higher drug concentrations and
increased apoptosis was also observed (P<0.05; Fig. 3, Table
I). These data suggest that Embelin induces apoptosis of Jurkat
cells in a dose-dependent manner.
 | Table I.Percentage of apoptotic Jurkat cells
following treatment with various doses of Embelin. |
Table I.
Percentage of apoptotic Jurkat cells
following treatment with various doses of Embelin.
| Group | Healthy cells
(%) | Apoptotic cells
(%) | Dead cells (%) |
|---|
| Control (DMSO) |
95.94±0.35 |
3.05±0.73 |
1.01±0.04 |
| Embelin 5
mmol/ml |
87.35±0.94 |
9.65±1.20a |
2.31±0.61 |
| Embelin 10
mmol/ml |
58.24±1.53 |
32.45±3.08a |
12.45±2.04 |
| Embelin 20
mmol/ml |
50.16±0.69 |
40.79±2.78a |
13.64±2.21 |
Effect of Embelin on activity of the
caspase pathway in Jurkat cells
In order to examine the molecular pathways
underlying the effects of Embelin on Jurkat cells, western blot
analysis was performed to measure the activity of
apoptosis-related, caspase proteins. The results showed that
treatment of Jurkat cells with Embelin for 48 h resulted in
cleavage of caspase-3, as demonstrated by the appearance of a 19
kDa intermediate. Furthermore, treatment of Jurkat cells with
Embelin also resulted in significantly increased levels of cleaved
caspase-9, with no concomitant change in the levels of procaspase-8
(Fig. 4A and B). Based on these
results, it was hypothesized that Embelin triggers Jurkat cell
apoptosis through the intrinsic but not the extrinsic pathway.
Notably, treatment with the caspase-3 inhibitor, z-DEVD-fmk, and
the caspase-9 inhibitor, Ac-LEHD-CHO, reduced the inhibitory
effects of Embelin on Jurkat T cell lymphoma cells (P<0.05;
Fig. 4C). Activation of caspases
during apoptosis partly accounts for the cleavage of certain
cellular substrates, such as poly ADP-ribose polymerase (PARP).
Therefore PARP has been adopted as a visual marker of caspase-3
activity in the intrinsic apoptosis pathway. As shown in Fig. 4, the levels of cleaved PARP fragment
(the active form) were significantly increased following the
exposure of cells to Embelin for 48 h, further clarifying the
activity of caspase-3 in Jurkat cells. These results also suggested
that Embelin induces apoptosis of Jurkat cells via the intrinsic
apoptotic pathway.
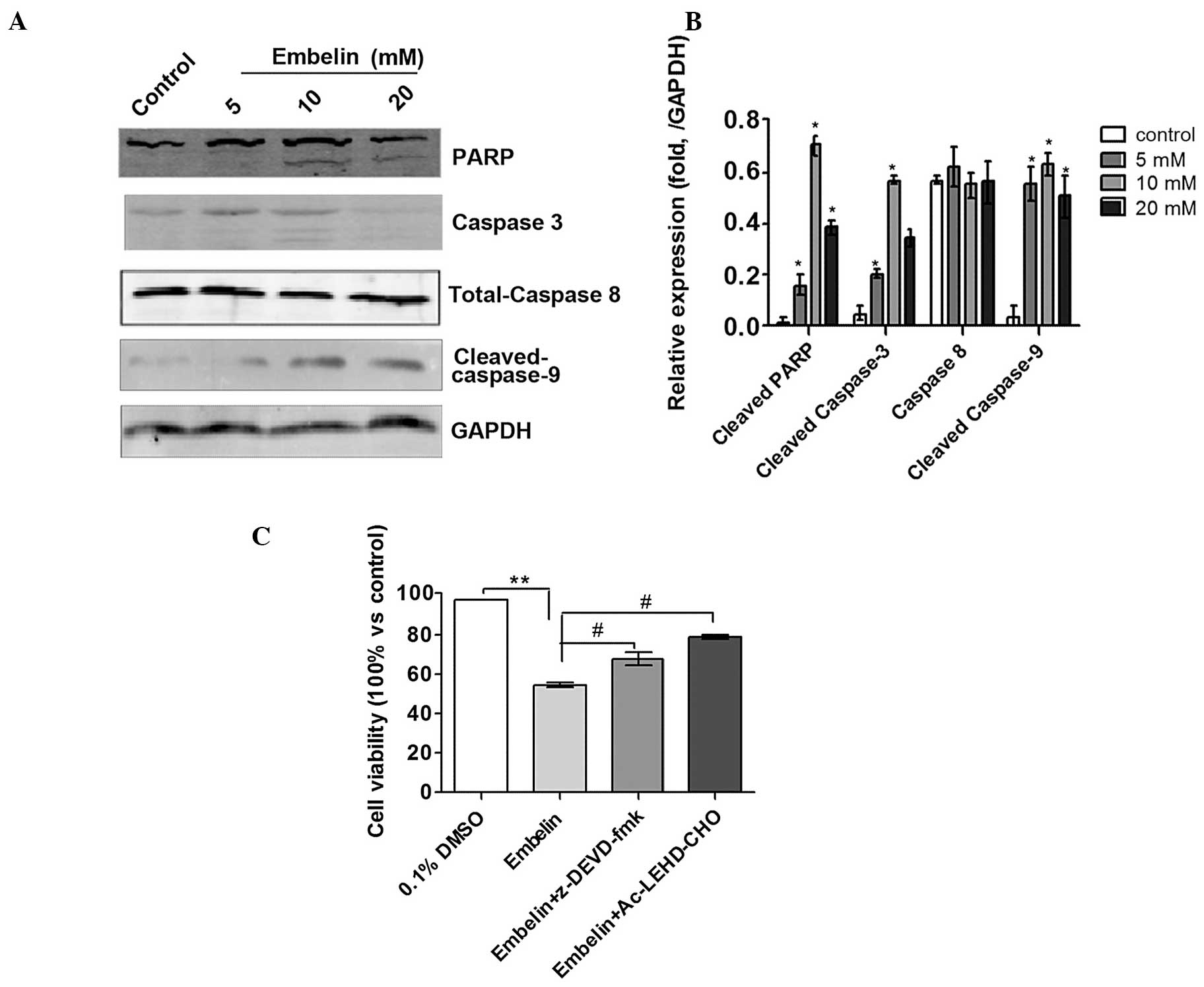 | Figure 4.Effect of Embelin on the activity of
the caspase pathway and effect of the caspase inhibitors,
z-DEVD-fmk and Ac-LEHD-CHO, on Embelin-treated Jurkat cells. (A)
Effect of Embelin on the expression of caspase 3, 8 and 9, and
PARP. Experiments were repeated in triplicate. (B) Quantitative
analysis of the results presented in (A). (C) Effect of Embelin
combined with caspase inhibitors, z-DEVD-fmk and Ac-LEHD-CHO, on
the viability of Jurkat cells. **P<0.01, vs. 0.1% DMSO control
group and #P<0.05, vs. 20 mM Embelin treatment group.
PARP, poly ADP ribose polymerase; DMSO, dimethyl sulfoxide. |
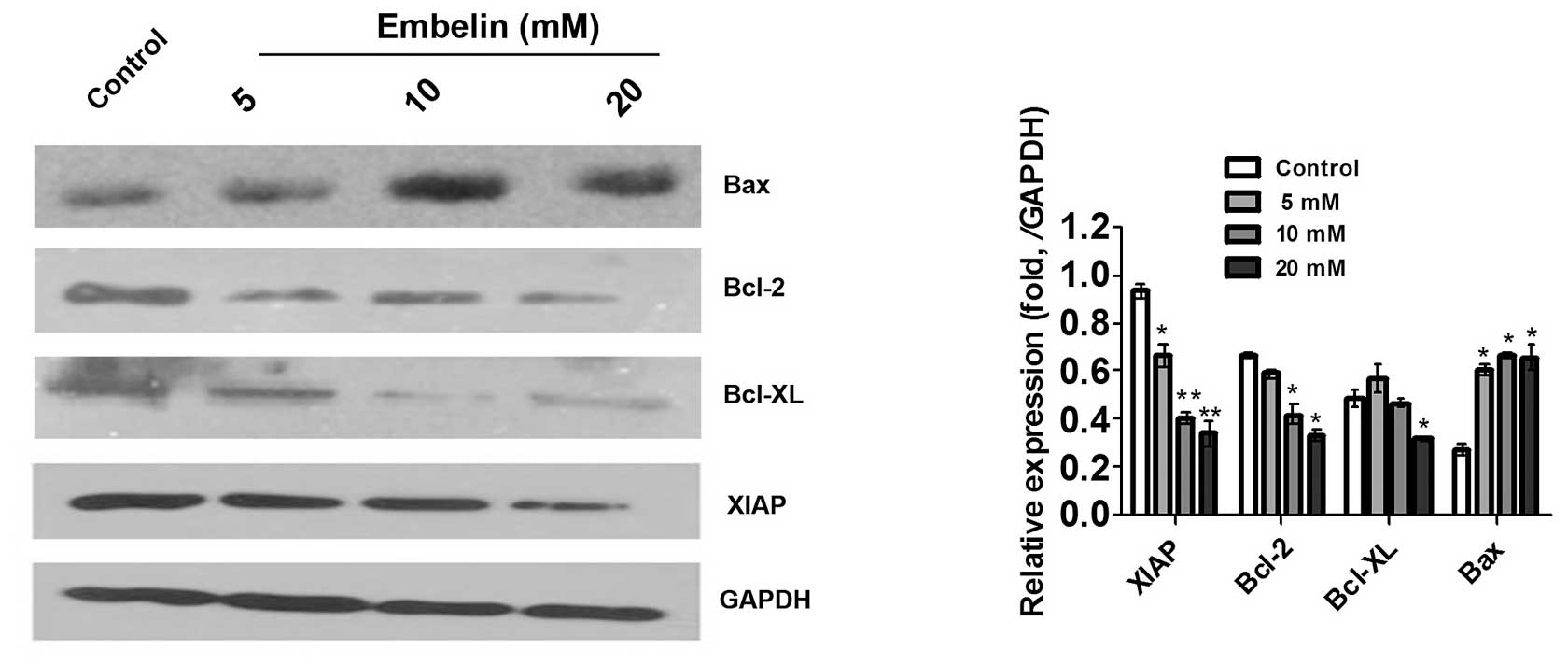
Effect of Embelin on the expression of
apoptosis-related proteins in Jurkat cells
Embelin is a small molecule inhibitor of XIAP that
is used in traditional Chinese medicine. The results of the present
study showed that 5–20 mM Embelin downregulated the expression of
XIAP in Jurkat cells (Fig. 5),
suggesting that Embelin induces apoptosis of Jurkat cells, in part
through regulation of the expression of XIAP. Mitochondrial
integrity is regulated by the Bcl-2 family, which is composed of
proapoptotic members, including Bcl-2 and Bcl-xl, and antiapoptotic
members, such as Bax (14,15). Therefore, in the present study, the
expression of these Bcl-2 family members was detected in the Jurkat
cells, following treatment with various concentrations of Embelin
for 48 h. As shown in Fig. 5,
decreased expression of Bcl-2 and Bcl-xl accompanied by an
increased expression of Bax was observed in Jurkat cells. These
results demonstrated that regulation of the Bcl-2 family also is
involved in the induction of apoptosis in Jurkat cells by
Embelin.
Discussion
XIAP is a recently discovered key member of the IAP
family and is the strongest inhibitor of apoptosis among the IAP
proteins (16). XIAP functions to
inhibit the activity of caspases, and therefore regulates apoptosis
via multiple pathways (17).
Overexpression of the XIAP gene is detected in the majority of
tumor cell lines and is closely associated with the progression,
recurrence, prognosis and resistance to chemotherapy of tumor cells
(18). XIAP overexpression has also
been demonstrated in a number of types of highly malignant tumor
cell lines and is associated with resistance to chemotherapy of
tumor cells (19). Embelin is a small
molecule inhibitor of XIAP, used in traditional Chinese medicine
that binds the Smac binding site in the BIR3 domain of XIAP, thus
blocking the inhibitory effects of XIAP on caspases, increasing the
level of active caspases, and thereby promoting apoptosis (20,21).
The present study examined the effects of Embelin on
human acute T cell lymphoma Jurkat cells. Embelin inhibited the
proliferation of acute lymphoma Jurkat cells in a dose-dependent
manner. Furthermore, the caspase-3 inhibitor, z-DEVD-fmk, and the
caspase-9 inhibitor, Ac-LEHD-CHO, inhibited the induction of
apoptosis by Embelin. Western blotting results demonstrated that
XIAP protein expression in the Embelin treatment groups was
significantly decreased, compared with that in the control group
(P<0.05). In addition, it was shown that Embelin treatment led
to the activation of caspase-3 and PARP. Furthermore, as the
concentration of Embelin increased, the levels of caspase-3 and
PARP cleavage fragments also increased. Caspase-3 is a member of
the caspase family, and functions as the executor of apoptosis; its
activation thus being important for cell apoptosis. Activation of
caspase-3 is primarily responsible for the cleavage of PARP and
ultimately results in cell apoptosis (22,23).
The current study also confirmed that Embelin
induces apoptosis by inhibiting the expression of XIAP and
activating the caspase pathway. In addition, it was shown that
following Embelin treatment, there was a dose-dependent increase in
the level of the caspase-9 cleavage fragment in the Embelin
treatment groups (5 and 10 mM Embelin) compared with the control
group, however this effect did not increase further with 20 mM
Embelin treatment, while no significant changes in the level of
caspase-8 were detected. The present study also demonstrated that
Embelin upregulated the proapoptotic protein, Bax, and
downregulated the expression of the antiapoptotic proteins, Bcl-2
and Bcl-xl, indicating that Embelin induces apoptosis via
activation of the mitochondrial pathway. These results suggested
that Embelin may induce apoptosis of Jurkat cells by inhibiting
XIAP, Bcl-2 and Bcl-xl, and by increasing the expression of Bax,
thus activating the caspase cascade reaction.
In conclusion, the current study showed that Embelin
inhibits the growth and induces apoptosis of Jurkat cells in
vitro, and that these effects are associated with its effect on
XIAP and the intrinsic apoptotic pathway. These results suggest
that Embelin may be a candidate for the treatment of T cell
lymphoma.
Acknowledgements
This study was supported by the Natural Science
Foundation of China (grant no. 81173611) and the Major
Infrastructure Projects of Hebei Science and Technology (grant no.
2011011) and was organized by The Fourth Hospital of Hebei Medical
University, China.
References
|
1
|
Andrade AF, Borges KS, Castro-Gamero AM,
Silveira VS, Suazo VK, Oliveira JC, Moreno DA, de Paula Queiroz RG,
Scrideli CA and Tone LG: Zebularine induces chemosensitization to
methotrexate and efficiently decreases AhR gene methylation in
childhood acute lymphoblastic leukemia cells. Anticancer Drugs.
25:72–81. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Stiff PJ, Unger JM, Cook JR, Constine LS,
Couban S, Stewart DA, Shea TC, Porcu P, Winter JN, Kahl BS, et al:
Autologous transplantation as consolidation for aggressive
non-Hodgkin's lymphoma. N Engl J Med. 369:1681–1690. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Plosker GL and Figgitt DP: A review of its
use in non-Hodgkin's lymphoma and chronic lymphocytic leukaemia.
Drugs. 63:803–843. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Till KJ, Coupland SE and Pettitt AR:
Motility and trafficking in B-cell non-Hodgkin's lymphoma (Review).
Int J Oncol. 45:5–12. 2014.PubMed/NCBI
|
|
5
|
Moreno-Martínez D, Nomdedeu M,
Lara-Castillo MC, Etxabe A, Pratcorona M, Tesi N, Díaz-Beyá M,
Rozman M, Montserrat E, Urbano-Ispizua A, et al: XIAP inhibitors
induce differentiation and impair clonogenic capacity of acute
myeloid leukemia stem cells. Oncotarget. 12:4337–4346. 2014.
|
|
6
|
Wang A, Zhang B, Zhang J and Wu W and Wu
W: Embelin-induced brain glioma cell apoptosis and cell cycle
arrest via the mitochondrial pathway. Oncol Rep. 29:2473–2478.
2013.PubMed/NCBI
|
|
7
|
Dai Y, Jiao H, Teng G, Wang W, Zhang R,
Wang Y, Hebbard L, George J and Qiao L: Embelin reduces
colitis-associated tumorigenesis through limiting IL-6/STAT3
signaling. Mol Cancer Ther. 13:1206–1216. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yao GH, Luan JF, Ye D, Yan JM, Lei QH, Zhu
PY and Jin J: Effects of triptolide on proliferation and apoptosis
of Jurkat cell line in acute T lymphocytic leukemia. Zhongguo Shi
Yan Xue Ye Xue Za Zhi. 16:506–509. 2008.(In Chinese). PubMed/NCBI
|
|
9
|
Hu R, Wu B, Zhang GJ, Wang HT, Zhu K, Yang
W and Liu ZG: Effect of Embelin on proliferation, differentiation
and apoptosis of HL-60 cells. Zhonghua Xue Ye Xue Za Zhi.
31:442–445. 2010.(In Chinese). PubMed/NCBI
|
|
10
|
Marsh JL, Jackman CP, Tang SN, Shankar S
and Srivastava RK: Embelin suppresses pancreatic cancer growth by
modulating tumor immune microenvironment. Front Biosci (Landmark
Ed). 19:113–125. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cheng YJ1, Jiang HS, Hsu SL, Lin LC, Wu
CL, Ghanta VK and Hsueh CM: XIAP-mediated protection of H460 lung
cancer cells against cisplatin. Eur J Pharmacol. 627:75–84. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Deveraux QL, Takahashi R, Salvesen GS and
Reed JC: X-linked IAP is a direct inhibitor of cell-death
proteases. Nature. 388:300–304. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao L, Shan B, Du Y, Wang M, Liu L and
Ren FZ: Periplocin from Cortex periplocae inhibits cell growth and
down-regulates survivin and c-myc expression in colon cancer in
vitro and in vivo via beta-catenin/TCF signaling. Oncol Rep.
24:375–383. 2010.PubMed/NCBI
|
|
14
|
Danial NN: Bcl-2 family proteins: Critical
checkpoints of apoptotic cell death. Clin Cancer Res. 13:7254–7263.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Silke J and Vucic D: IAP family of cell
death and signaling regulators. Methods Enzymol. 545:35–65. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kuwana T and Newmeyer DD: Bcl-2-family
proteins and the role of mitochondria in apoptosis. Curr Opin Cell
Biol. 15:691–699. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bao W, Zhu F, Duan Y, Yang Y and Cai H:
HtrA1 resensitizes multidrug-resistant hepatocellular carcinoma
cells by targeting XIAP. Biomed Pharmacother. 70:97–102. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen FH, Qu HY and Sui GJ: Effects of
apoptosis protein XIAP inhibito on the apoptosis and sensitivity of
chemotherapy in A549 cell. Chin J Lung Cancer. 11:368–372. 2008.(In
Chinese).
|
|
19
|
Nikolovska-Coleska Z, Xu L, Hu Z, et al:
Discovery of embelin as a cell-permeable, small-molecular weight
inhibitor of XIAP through structure-based computational screening
of a traditional herbal medicine three-dimensional structure
database. J Med Chem. 47:2430–2440. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Irshad S, Ashworth A and Tutt A:
Therapeutic potential of PARP inhibitors for metastatic breast
cancer. Expert Rev Anticancer Ther. 11:1243–1251. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nikrad M, Johnson T, Puthalalath H,
Coultas L, Adams J and Kraft AS: The proteasome inhibitor
bortezomib sensitizes cells to killing by death receptor ligand
TRAIL via BH3-only proteins Bik and Bim. Mol Cancer Ther.
4:443–449. 2005.PubMed/NCBI
|
|
22
|
Wong RS: Apoptosis in cancer: From
pathogenesis to treatment. J Exp Clin Cancer Res. 30:872011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ulukaya E, Acilan C and Yilmaz Y:
Apoptosis: Why and how does it occur in biology. Cell Biochem
Funct. 29:468–480. 2011. View
Article : Google Scholar : PubMed/NCBI
|