Introduction
Gliomas account for ~81% of malignant intracranial
tumors, among which astrocytomas are the most common type (1). A recent study indicated that 14.5% of
patients with astrocytoma survived for five years from diagnosis
(2). However, the pathogenesis and
progression of astrocytoma remain poorly understood, and
therapeutic strategies for this disease are limited. Due to the
necessity to identify novel therapeutic approaches, further studies
are required, in order to examine the biological processes
underlying the development of astrocytoma (3).
microRNAs (miRNAs) are small, non-coding RNA
molecules of 20–23 nucleotides in length. They have been shown to
be involved in regulating the development and maintenance of
undifferentiated or incompletely differentiated cell types
(4–6).
Accumulating evidence indicates that dysregulation of specific
miRNAs may be associated with certain types of cancer, and that
miRNAs act as oncogenes or tumor suppressors of particular target
genes (7–10). miR-155, for example, acts as an
inhibitor of ovarian cancer-initiating cells by regulating claudin
1, thus disrupting the development of human ovarian cancer
(11). Furthermore, fragments of
miRNA have been detected and measured in the serum of patients with
cancer, demonstrating that they may be useful as biomarkers for
disease diagnosis. One such example is the upregulation of miR-23a,
27a and 24 in the serum of patients with hepatocellular carcinoma,
which are associated with tumor-suppressive activities (12). However, whether specific miRNAs are
dysregulated in astrocytoma, and whether they contribute to the
development and progression of this disease remains unclear.
Neuritin, encoded by the NRN1 gene, is a novel
member of the neurotrophic factor (NF) family, and is induced by
neuronal activity in the rat hippocampus (13). Within the central nervous system,
neuritin promotes neurite outgrowth and synaptic maturation,
protects motor neuron axons and regulates apoptosis of
proliferative neurons (13). Notably,
neuritin has also been shown to be expressed outside the nervous
system, including in invasive breast carcinoma and Kaposi's
sarcoma, where it is associated with apoptosis and tumorigenesis
(14,15). In accordance with the results of
previous studies, Zhang et al (16) demonstrated that neuritin was
overexpressed in the U251 human astrocytoma cell line, and that the
level of expression was positively correlated with tumor
malignancy. Furthermore, recent studies have demonstrated that
neuritin acts as a novel angiogenic factor, promoting tumor
angiogenesis (17), and that it
modulates neuronal migration (18).
However, the characteristics and mechanism of action of neuritin
regulation in astrocytomas, particularly at the epigenetic level,
have not yet been described.
The present study investigated whether miR-182
specifically targets NRN1, and influences cell proliferation and
migration ability in the U251 human astrocytoma cell line.
Materials and methods
Bioinformatics analysis of miR-182 and
the 3′-UTR of NRN1
The precursor miRNA (pre-miRNA) sequences, mature
miRNA sequences, chromosomal locations, and the length of miR-182
and the target gene, NRN1, were analyzed in multiple species
(human, rhesus and mouse) using the online research tool of the
miRBase Target database (http://www.mirbase.org) (19,20).
Cell culture
The U251 human astrocytoma cell line was grown in
Dulbecco's modified Eagle's medium (DMEM; Hyclone, Logan, UT, USA),
supplemented with 10% fetal bovine serum (FBS; PAA Lab., Inc.,
Morningside, Queensland, Australia), penicillin (100 U/ml),
streptomycin (100 U/ml) and 2 mM L-glutamine (Hyclone). U251 cells
were maintained at 37°C in a humidified atmosphere of air
containing 5% CO2.
Vector construction
For experiments measuring miRNA-182 expression, the
plasmid pRNAT-CMV32-cGFP-mir182 (pre-miRNA of miRNA-182),
oligonucleotide pairs for the pre-miRNA of miRNA-182 and linker
sequences with BamHI and XhoI sites, were chemically
synthesized (20). The sequences of
the oligonucleotides were as follows: Top strand,
5′-GTggatccCTGTTTGGCAATGGTAGAACTCACACTT
TTTGCCTCCAACTGACTCCTACATATTAGCATTAACAGc tcgagCC-3′ and bottom
strand, 5′-GGc tcg agC TGT TAA TGC TAA TAT GTA GGA GTC AGT TGG AGG
CAA AAA AGT GTG AGT TCT ACC ATT GCC AAA CAG
gga tcc AC-3′ (sequences corresponding to miRNA-182 seed sequences
are shown in bold and restriction enzyme sites are in lower case).
In order to build the expression plasmid, the pairs of
oligonucleotides were annealed and inserted into the multiple
cloning sites between the BamHI and XhoI restriction
sites in the pRNAT-CMV32-cGFP/Neo vector (GenScript, Piscataway,
NJ, USA). The negative control plasmid, pRNAT-CMV32-cGFP-
mir182-Mut, was similarly constructed, with 17 nucleotides in the
sequence corresponding to the miRNA-182 seed sequence, mutated from
TTTGGCAATGGTAGAAC to
TaTGcgAtTGGaAcAtg
(mutations shown in lower case). Transfection of 0.3 µg of
miRNA-182 or miRNA-182 mutant expression vector into the U251 cell
line was conducted using Lipofectamine 2000™ (Thermo Fisher
Scientific, Inc.,Waltham, MA, USA) transfection reagent, according
to the manufacturer's instructions.
RNA extraction and analysis by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA from each group of cells was isolated
using TRIzol™ reagent (Invitrogen Life Technologies, Carlsbad, CA,
USA) according to the manufacturer's instructions. RNA samples were
treated with DNase I (Sigma-Aldrich, St. Louis, MO, USA),
quantified and then reverse-transcribed into cDNA using the
ReverTra Ace-α first strand cDNA synthesis kit (Toyobo Co., Ltd.,
Osaka, Japan). RT-qPCR was conducted using a RealPlex4 real-time
PCR detection system (Eppendorf Co., Ltd., Hamburg, Germany), with
SYBR-Green real-time PCR Master Mix (Toyobo Co., Ltd.) as the
detection dye. The following PCR primer sequences used: Forward,
5′-ATCCTCGCGGTGCAAATAG-3′ and reverse, 5′-GAGCAAACAGTCCGAAAAGC-3′
for NRN1 and forward, 5′-CAGCCACCCGAGATTGAGCA-3′ and reverse,
5′-TAGTAGCGACGGGCGGTGTG-3′ for 18S rRNA. PCR was performed under
the following conditions: Denaturation at 95°C for 15 sec,
annealing at 58°C for 30 sec and extension at 72°C for 42 sec. The
comparative threshold cycle (Ct) was used to determine relative
NRN1 gene expression, normalized to that of 18S rRNA. For each
sample, the Ct values of the genes were normalized using the
formula ΔCt = Ctgenes - Ct18S rRNA. In order
to determine relative expression levels, the following formula was
used: ΔΔCt = ΔCtall groups - ΔCtblank control
group. The values used to plot the relative expression of
markers were calculated using the expression 2−ΔΔCt. The
cDNA of each gene was amplified with primers as previously
described (21).
Flow cytometric (FCM) analysis of cell
cycle progression
Each group of U251 cells was seeded at
3×105 cells per well in six-well plates, and cultured
until 85% confluence was reached. After washing with
phosphate-buffered saline (PBS) three times, cells were collected
by centrifugation (Allegra X-22R; Beckman Coulter, Brea, CA, USA)
at 1000 × g for 5 min. Cell pellets were resuspended in 1 ml of
PBS, fixed in 75% ice-cold ethanol, and stored in a freezer for
>48 h. Prior to FCM analysis, fixed cells were centrifuged and
washed twice with PBS, then resuspended in propidium iodide (PI)
staining solution (Sigma-Aldrich), containing 50 µl/ml PI and 250
µl/ml RNase A (Sigma-Aldrich, Carlsbad, CA, USA). The cell
suspension was incubated for 30 min at 4°C in darkness and analyzed
using FACS (FCM-500; Beckman Coulter). A total of 20,000 events
were acquired for the analysis, using CellQuest software (version
5.1; BD Biosciences, Franklin Lakes, NJ, USA).
Luciferase reporter assay
All steps of the luciferase reporter assay were
performed, as previously described (12,22).
Non-anthropogenic NIH-3T3 cells (Stem Cell Bank, Chinese Academy of
Sciences, Shanghai, China) were used to avoid the influence of
endogenous human miRNA-182. NIH-3T3 cells were seeded at
3×104 cells per well in 48-well plates and cotransfected
with 400 ng pRNAT-CMV32-cGFP-mir182, pRNAT-CMV32-cGFP or
pRNAT-CMV32-cGFP-mir182-mut, and 20 ng pGL3cm-CTTN-3UTR-WT or
pGL3cm-CTTN-3UTR-mut, and pGL-3 (Promega Corporation, Madison, WI,
USA), using Lipofectamine 2000 transfection reagent according to
the manufacturer's instructions. Following transfection for 48 h,
the luciferase activity was measured using the dual-luciferase
reporter assay system (Promega Corporation).
Northern blotting
All steps for the northern blotting analysis were
performed as previously described (18). For all groups, 20 µg of high-quality
total RNA was analyzed on a 7.5 M urea, 12% paraformaldehyde
denaturing gel, and transferred to a Hybond-N+ nylon
membrane (Amersham, Freiburg, Germany). Membranes were cross-linked
using UV light for 30s at 1200 mJ/m2. Hybridization was
performed with the miRNA-182 antisense StarFire probe,
5′-TTTGGCAATGGTAGAACTCACACT-3′ (IDT, Coralville, IA, USA), in order
to detect the 22-nucleotide miRNA-182 fragments, according to the
manufacturer's instructions. After washing, membranes were exposed
for 20–40 h to Kodak XAR-5 film (Sigma-Aldrich). As a positive
control, all membranes were hybridized with a human U6 snRNA probe,
5′-GCA GGG GCC ATG CTA ATC TTC TCT GTA TCG-3′. Exposure times for
the U6 control probe varied from between 15 to 30 min.
Western blotting
Total protein extracts of each group of cells were
resolved by 12% SDS-PAGE and transferred to PVDF membranes
(IPVH00010; Merck Millipore, Billerica, MA, USA). After blocking,
the PVDF membranes were washed 4 times for 15 min with
Tris-buffered saline with Tween-20® (TBST; Beyotime Institute of
Biotechnology, Haimen, China) at room temperature and incubated
with rabbit anti-human neuritin polyclonal antibody (1:500
dilution; cat no. Ab64186; Abcam, San Francisco, CA, USA).
Following extensive washing, membranes were incubated with the
secondary peroxidase-linked goat anti-rabbit IgG (1:1000 dilution;
cat no. sc-45101; Santa Cruz Biotechnology, Inc., Dallas, TX, USA)
for 1 h. After washing four times for 15 min with TBST at room
temperature, immunoreactivity was visualized using the enhanced
chemiluminescence ECL kit (Pierce Biotechnology, Inc., Rockford,
IL, USA), and membranes were exposed to Kodak XAR-5 film.
Transwell migration assay
All steps were performed as previously described
(23). Cells (2×105) were
resuspended in 200 µl of serum-free medium, and seeded on the top
chamber of the 8.0-µm pore, 6.5-mm polycarbonate transwell filters
(Corning, Lowell, MA, USA). Complete medium (600 µl) containing 10%
FBS was added to the bottom chamber. For the invasion assay,
inserts coated with Matrigel™ (product no. 356234; Shanghai
Hengyuan Macromolecular Materials Co., Ltd., Shanghai, China) were
used. Cells were allowed to migrate for 24, 48 or 72 h at 37°C in a
humidified incubator with 5% CO2. The cells that
attached to the lower surface of the membranes were fixed in 4%
paraformaldehyde at room temperature for 30 min and stained with
DAPI (C1002; Beyotime Institute of Biotechnology), and the number
of cells on the lower surface of the filter was counted under the
microscope (BX51TF; Olympus Corporation, Tokyo, Japan). A total of
five fields were counted from each transwell filter.
Cell Counting kit-8 (CCK8) cell
viability assay
Cell viability was assessed using a CCK8 assay
(Dojindo, Kumamoto, Japan). U251 cells transfected with miR-182 or
mutant miR-182, were seeded into 96-well plates. Following
transfection for 0 and 72 h, 10 µl of CCK8 was added to each well
and incubated at 37°C for 1 h. Cell viability was measured at 450
nm using an ELISA reader (BioTek, Winooski, VT, USA), according to
the manufacturer's instructions.
Statistical analysis
Each experiment was performed at least three times,
and data are presented as the mean ± standard error, where
applicable. Differences were evaluated using Student's t-test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Bioinformatics analysis of miR-182 and
the 3′-UTR of NRN1
Multi-species bioinformatics was performed using the
online research tool of the miRBase Target database (http://www.mirbase.org) (19,20). The
present study focused on human miR-182, which may target the 3′-UTR
of NRN1. This site is conserved, to varying degrees, across
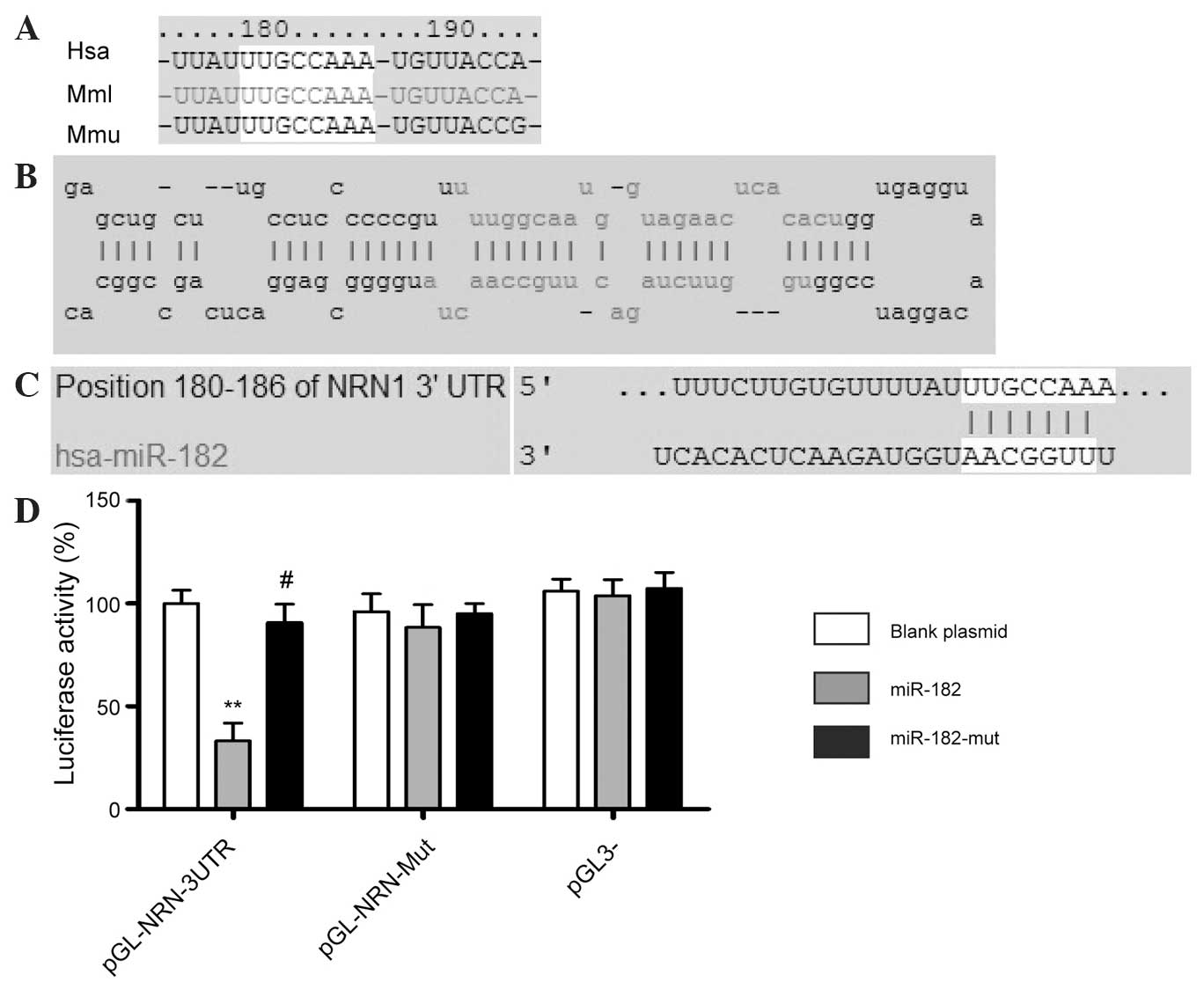
different species (Fig. 1).
Identification of miR-182 binding
sites in the 3′-UTR region of NRN1
A luciferase reporter assay was used to determine
whether NRN1 expression was regulated by mature miR-182. Plasmids
with a WT or mutant NRN1 mRNA 3′-UTR miR-182 binding site, or an
empty plasmid control were cotransfected with the miR-182
expression vector (WT miR-182, empty vector or mutant miR-182) into
NIH-3T3 cells. The results showed that the activity of the
luciferase reporter containing the WT NRN1 3′-UTR, was
significantly inhibited following transfection with WT miR-182,
while the luciferase activity of the reporter gene with the mutated
NRN1 3′-UTR was unchanged, indicating that miR-182 may target NRN1
mRNA by specifically binding to its 3′-UTR (Fig. 1D).
miR-182 influences the expression of
neuritin in the U251 human astrocytoma cell line
Subsequently, the regulation of neuritin expression
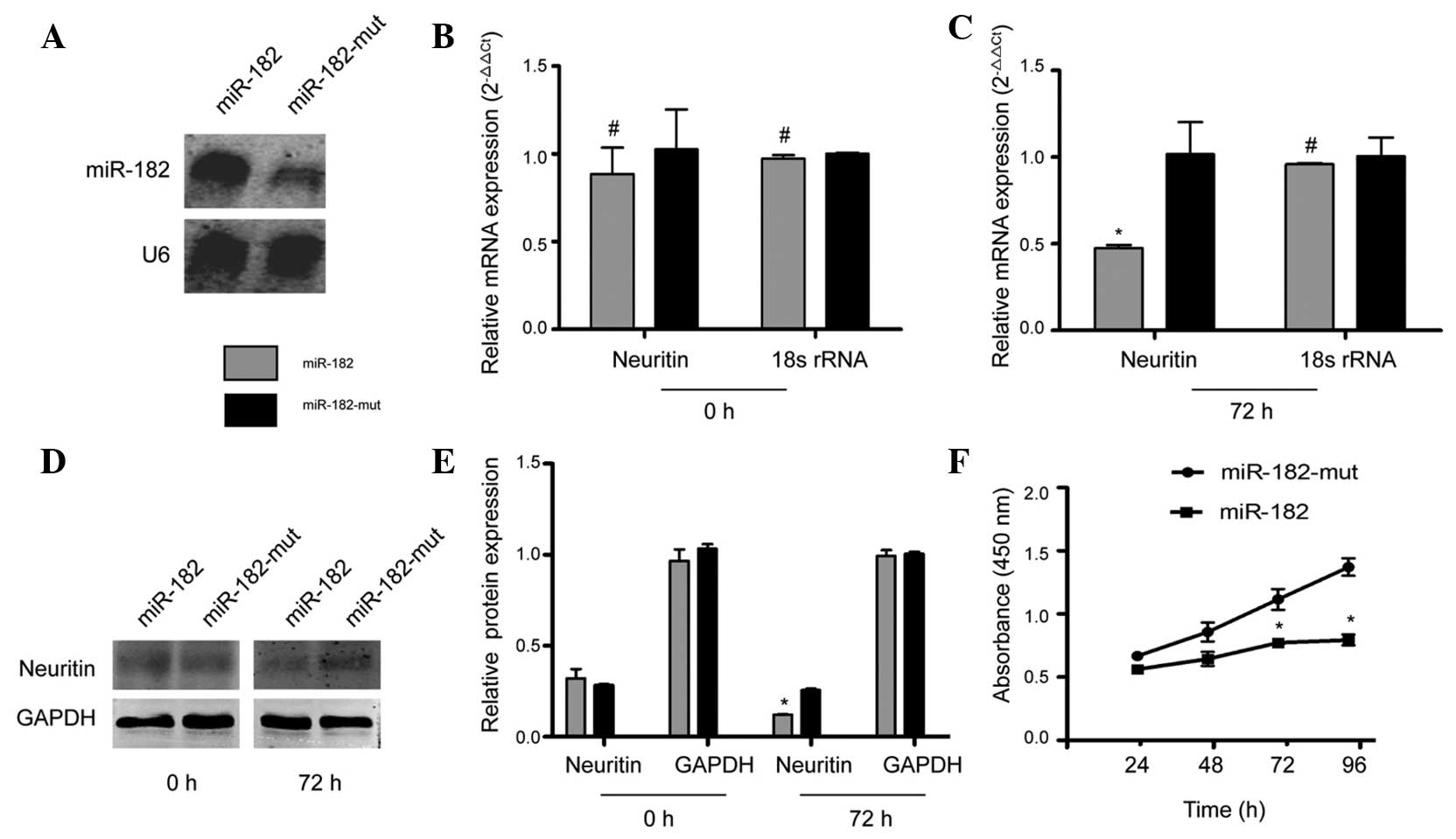
in U251 cells by exogenous miR-182 was examined. Northern blotting
results showed that the hybridization signal of mutant
miR-182-transfected U251 cells was weaker than that in the
miR-182-transfected cells (Fig. 2A).
These results were confirmed by RT-qPCR, which showed that at 72 h
following transfection with the miR-182 expression vector, NRN1
mRNA levels decreased significantly, compared with those of U251
cells transfected with mutant miR-182. The relative mRNA expression
is shown following normalization to that of 18S rRNA, which was
used as an internal control (Fig. 2B and
C). Furthermore, western blotting indicated that protein levels
of neuritin in mutant miR-182-transfected cells were ~2-fold higher
than levels in cells transfected with miR-182 (Fig. 2D and E). The CCK8 cell viability assay
indicated no difference in viability between miR-182-transfected
cells and mutant miR-182-transfected cells at 24 h. However, a
significant reduction in cell viability was observed in cells
transfected with miR-182 at 72 h and 96 h, compared with mutant
miR-182-transfected cells (Fig. 2F).
These data suggest that exogenous miR-182 downregulates NRN1
expression.
Alteration of cell cycle progression
and invasion ability of U251 cells following miR-182
transfection
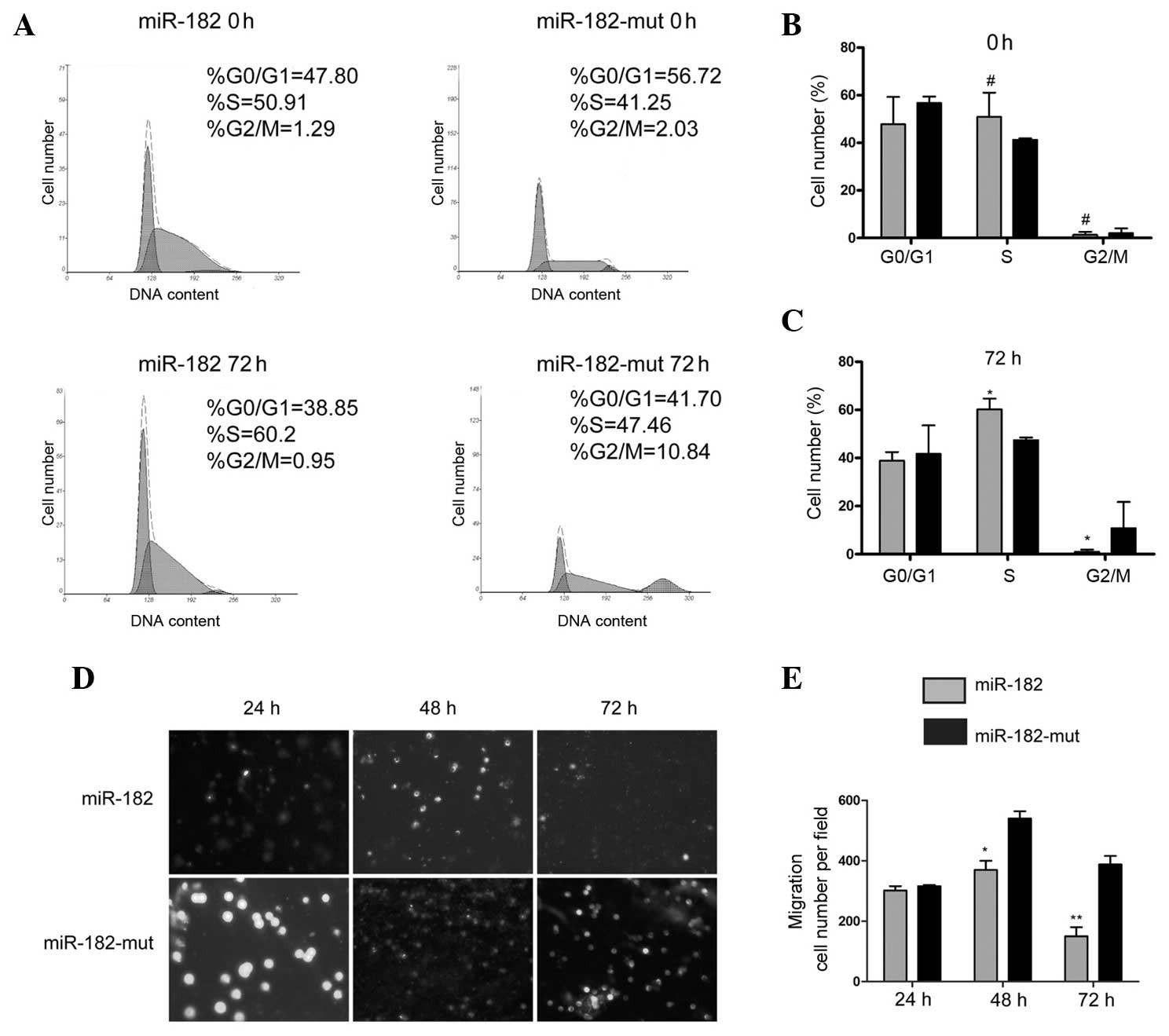
A flow cytometric (FCM) assay was performed in order
to detect changes in cell cycle progression. miR-182-transfected
and mutant miR-182-transfected U251 cells were stained with PI and
analyzed by FCM. The results showed no significant differences in
cell cycle distribution between miR-182-transfected and mutant
miR-182-transfected cells at 0 h (Fig.
3A). However, at 72 h following transfection, the number of
cells that were arrested in S phase of the cell cycle, and the
percentage of cells in the G2/M phase, significantly decreased in
the miR-182-transfected cells compared with the proportions in the
miR-182-mut-transfected cells (Fig.
3A–C). These results indicate that increased expression of
miR-182 significantly affects cell cycle progression in
vitro. The transwell migration invasion assay showed that the
number of invading cells was significantly reduced in
miR-182-transfected cells at 48 h and 72 h, compared with the
number of invading mutant miR-182-transfected cells, while no
significant difference in the number of invading cells was detected
between the two groups 24 h after transfection (Fig. 3D and E). These results suggested that
reduced expression of neuritin in the U251 cell line following
miR-182 transfection influences cell cycle progression and reduces
cell invasion and migration.
miR-182 inhibition of neuritin
promotes apoptosis in the U251 cell line
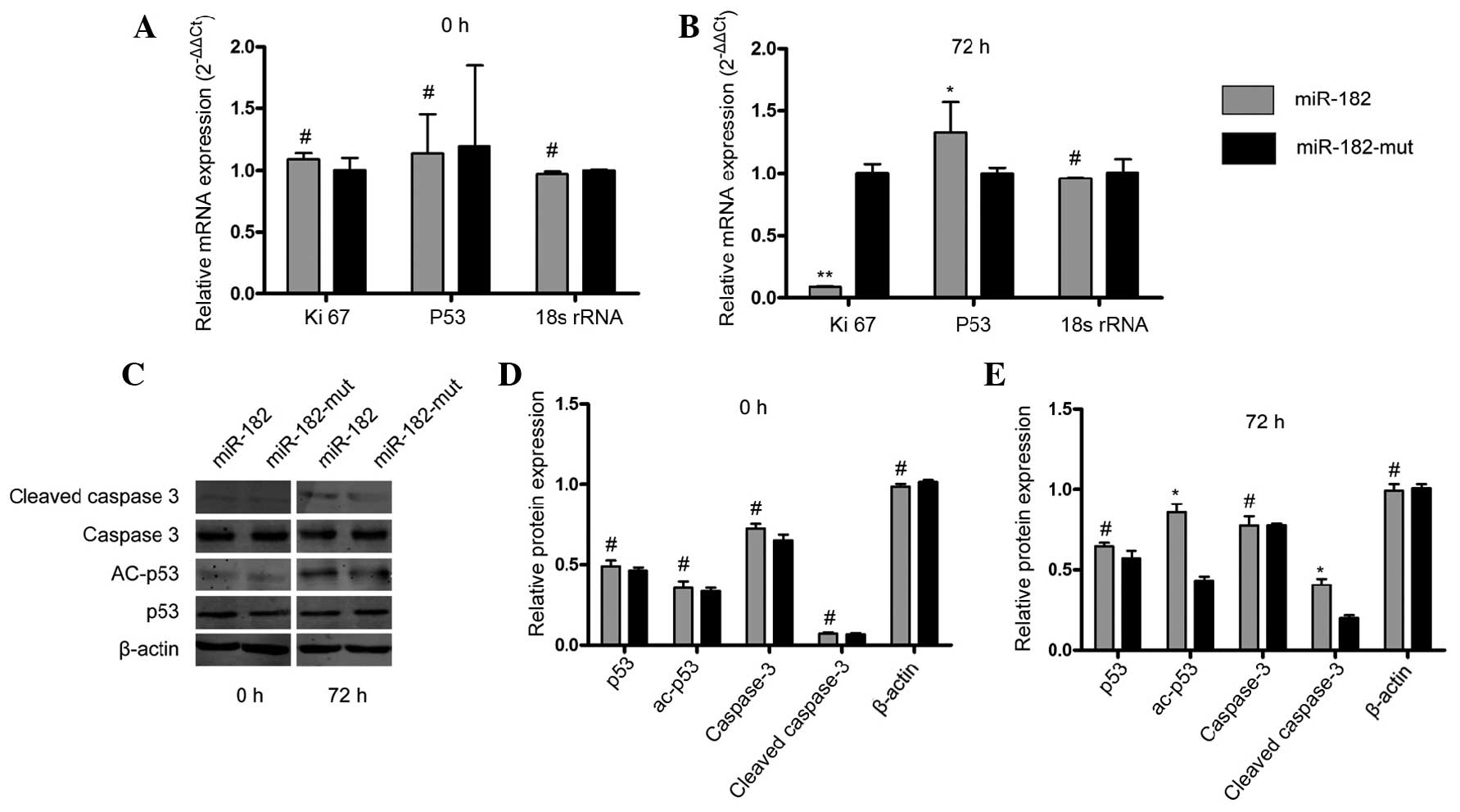
In order to determine whether inhibition of neuritin
by miR-182 influences cell proliferation and apoptosis in U251
cells, RT-qPCR and western blotting were performed. As shown in
Fig. 4A and B, the expression of
Ki-67 mRNA (23), a cell
proliferation-related protein, was inhibited when the miR-182
expression plasmid was transfected into U251 cells after 72 h,
compared with that in the mutant miR-182-transfected cells. In
addition, p53 mRNA levels were increased in miR-182-transfected
cells, compared with levels in mutant miR-182-transfected cells.
Western blotting demonstrated changes in the levels of
apoptosis-related proteins. It was shown that the protein levels of
cleaved caspase 3 and acetyled-p53 were increased significantly in
miR-182-transfected cells, compared with levels in mutant
miR-182-transfected cells at 72 h following transfection (Fig. 4C–E). These results indicated that
exogenous miR-182 mediates inhibition of neuritin, suppresses cell
proliferation and promotes apoptosis in astrocytoma cell lines.
Discussion
Astrocytomas are the most common type of glial
tumors, and are associated with a poor prognosis due to
infiltrative growth and a high migratory capacity (24,25). The
low median survival of patients with astrocytomas highlights the
requirement for a comprehensive understanding of this disease.
Neuritin is a new member of the NF family, which has
received little attention in terms of research into the development
of astrocytoma from an epigenetic perspective. A previous study
confirmed that neuritin is overexpressed in U251 cells and that its
expression is positively correlated with tumor malignancy (16). The present study used a U251 cell
line, transfected with an miR-182 vector, in order to detect
whether the expression of neuritin is regulated by miR-182, and to
examined its influence on cell proliferation and invasion in these
cells.
To the best of our knowledge, the current results
showed for the first time that miR-182 specifically targets
neuritin. It was demonstrated that the mRNA and protein levels of
neuritin were decreased in miR-182-transfected U251 cells compared
with mutated miR-182-transfected cells. In addition, flow cytometry
showed that the majority of the miR-182-transfected cells were
arrested in the S phase of the cell cycle, suggesting that miR-182
overexpression influences cell cycle progression and tumor growth
in vitro. Similarly, a transwell migration invasion assay
demonstrated that the number of invading cells was significantly
decreased in miR-182-transfected cells compared with mutant
miR-182-transfected cells, suggesting that miR-182 suppresses cell
invasion ability. Furthermore, RT-qPCR analysis showed that levels
of Ki-67 and p53 mRNA, two important factors associated with cell
proliferation and apoptosis, were decreased and increased,
respectively, following transfection with miR-182. Western blotting
also confirmed that the levels of two apoptosis-related proteins
increased significantly when cells were transfected with miR-182,
compared with transfection with mutant miR-182. These data
demonstrate that miR-182 targets neuritin and regulates U251 cell
proliferation and invasion. Inhibition or loss of miR-182 may lead
to neuritin overexpression and thus promote tumorigenesis.
In conclusion, the present study showed that miR-182
specifically targets neuritin, and inhibits the proliferation and
invasion of astrocytoma cells. Furthermore, miR-182-induced
neuritin inhibition promoted apoptosis, which is a possible
explanation for the observed suppression of cell proliferation and
invasion. Further studies and experiments in vivo are
required in order to fully explain the function of miR-182 and its
potential targets in the development and progression of
astrocytoma.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (grant nos. 81371410 and
81171205), the National Basic Research Program of China (973
Program; grant no. 2011CB707506), the National Natural Science
Foundation of China (grant no. 81202811), Project funded by China
Postdoctoral Science Foundation (grant no. 2014M550250) and
Shanghai Municipal Health Bureau Fund (grant no. 20124320).
Glossary
Abbreviations
Abbreviations:
|
CCK8
|
cell counting kit-8
|
|
FBS
|
fetal bovine serum
|
|
FCM
|
flow cytometric
|
|
miRNA
|
microRNA
|
|
NF
|
neurotrophic factor
|
|
NSCLC
|
non-small cell lung cancer
|
|
pre-miRNA
|
precursor miRNA
|
References
|
1
|
Wang W, Da R, Wang M, Wang T, Qi L, Jiang
H, Chen W and Li Q: Expression of brain-specific angiogenesis
inhibitor 1 is inversely correlated with pathological grade,
angiogenesis and peritumoral brain edema in human astrocytomas.
Oncol Lett. 5:1513–1518. 2013.PubMed/NCBI
|
|
2
|
Fuentes-Raspall R, Puig-Vives M,
Guerra-Prio S, Perez-Bueno F and Marcos-Gragera R: Population-based
survival analyses of central nervous system tumors from 1994 to
2008. An up-dated study in the temozolomide-era. Cancer Epidemiol.
38:244–247. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ostrom QT, Bauchet L, Davis FG, Deltour I,
Fisher JL, Langer CE, Pekmezci M, Schwartzbaum JA, Turner MC, Walsh
KM, et al: The epidemiology of glioma in adults: A state of the
science review. Neuro Oncol. 16:2014. View Article : Google Scholar
|
|
4
|
Lee RC, Feinbaum RL and Ambros V: The C.
elegans heterochronic gene lin-4 encodes small RNAs with antisense
complementarity to lin-14. Cell. 75:843–854. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lungu G, Stoica G and Ambrus A: MicroRNA
profiling and the role of microRNA-132 in neurodegeneration using a
rat model. Neurosci Lett. 553:153–158. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cui J, Li D, Zhang W, Shen L and Xu X:
Bioinformatics analyses combined microarray identify the
deregulated microRNAs in oral cancer. Oncol Lett. 8:218–222.
2014.PubMed/NCBI
|
|
7
|
Cheng W, Liu T, Wan X, Gao Y and Wang H:
MicroRNA-199a targets CD44 to suppress the tumorigenicity and
multidrug resistance of ovarian cancer-initiating cells. FEBS J.
279:2047–2059. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kong W, Yang H, He L, Zhao JJ, Coppola D,
Dalton WS and Cheng JQ: MicroRNA-155 is regulated by the
transforming growth factor beta/Smad pathway and contributes to
epithelial cell plasticity by targeting RhoA. Mol Cell Biol.
28:6773–6784. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schetter AJ, Leung SY, Sohn JJv, Zanetti
KA, Bowman ED, Yanaihara N, Yuen ST, Chan TL, Kwong DL, Au GK, et
al: MicroRNA expression profiles associated with prognosis and
therapeutic outcome in colon adenocarcinoma. Jama. 299:425–436.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lanza G, Ferracin M, Gafà R, Veronese A,
Spizzo R, Pichiorri F, Liu CG, Calin GA, Croce CM and Negrini M:
mRNA/microRNA gene expression profile in microsatellite unstable
colorectal cancer. Mol Cancer. 6:542007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Qin W, Ren Q, Liu T, Huang Y and Wang J:
MicroRNA-155 is a novel suppressor of ovarian cancer-initiating
cells that targets CLDN1. FEBS Lett. 587:1434–1439. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huang S, He X, Ding J, Liang L, Zhao Y,
Zhang Z, Yao X, Pan Z, Zhang P, Li J, et al: Upregulation of
miR-23a approximately 27a approximately 24 decreases transforming
growth factor-beta-induced tumor-suppressive activities in human
hepatocellular carcinoma cells. Int J Cancer. 123:972–978. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhou S and Zhou J: Neuritin, a
neurotrophic factor in nervous system physiology. Curr Med Chem.
21:1212–1219. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Raggo C, Ruhl R, McAllister S, Koon H,
Dezube BJ, Früh K and Moses AV: Novel cellular genes essential for
transformation of endothelial cells by Kaposi's sarcoma-associated
herpesvirus. Cancer Res. 65:5084–5095. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Putz U, Harwell C and Nedivi E: Soluble
CPG15 expressed during early development rescues cortical
progenitors from apoptosis. Nat Neurosci. 8:322–331. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang L, Zhao Y, Wang CG, Fei Z, Wang Y,
Li L, Li L and Zhen HN: Neuritin expression and its relation with
proliferation, apoptosis and angiogenesis in human astrocytoma. Med
Oncol. 28:907–912. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Han D, Qin B, Liu G, Liu T, Ji G, Wu Y and
Yu L: Characterization of neuritin as a novel angiogenic factor.
Biochem Biophys Res Commun. 415:608–612. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zito A, Cartelli D, Cappelletti G,
Cariboni A, Andrews W, Parnavelas J, Poletti A and Galbiati M:
Neuritin 1 promotes neuronal migration. Brain Struct Funct.
219:105–118. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cheng W, Liu T, Jiang F, Liu C, Zhao X,
Gao Y, Wang H and Liu Z: microRNA-155 regulates angiotensin II type
1 receptor expression in umbilical vein endothelial cells from
severely pre-eclamptic pregnant women. Int J Mol Med. 27:393–399.
2011.PubMed/NCBI
|
|
20
|
Zhang L, Liu T, Huang Y and Liu J:
microRNA-182 inhibits the proliferation and invasion of human lung
adenocarcinoma cells through its effect on human cortical
actin-associated protein. Int J Mol Med. 28:381–388.
2011.PubMed/NCBI
|
|
21
|
Liu T, Cheng W, Lai D, Huang Y and Guo L:
Characterization of primary ovarian cancer cells in different
culture systems. Oncol Rep. 23:1277–1284. 2010.PubMed/NCBI
|
|
22
|
He M, Xu Z, Ding T, Kuang DM and Zheng L:
MicroRNA-155 regulates inflammatory cytokine production in
tumor-associated macrophages via targeting C/EBPbeta. Cell Mol
Immunol. 6:343–352. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jiang F, Liu T, He Y, Yan Q, Chen X, Wang
H and Wan X: MiR-125b promotes proliferation and migration of type
II endometrial carcinoma cells through targeting TP53INP1 tumor
suppressor in vitro and in vivo. BMC Cancer. 11:4252011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu Y, Tang K, Yan W, Wang Y, You G, Kang
C, Jiang T and Zhang W: Identifying Ki-67 specific miRNA-mRNA
interactions in malignant astrocytomas. Neurosci Lett. 546:36–41.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hamadi A, Giannone G, Takeda K and Rondé
P: Glutamate involvement in calcium-dependent migration of
astrocytoma cells. Cancer Cell Int. 14:422014. View Article : Google Scholar : PubMed/NCBI
|