Introduction
Tumor formation is driven by the activation of
oncogenes; however, cells harbor several barriers to tumor
development, including cell growth inhibition, premature senescence
and cell death (1,2). Ras-induced cellular stress is a widely
explored ex vivo model employed in order to study
oncogene-induced cellular barriers (3–6).
Overexpression of Ras activates p38-regulated/activated protein
kinase (PRAK) (7), which
phosphorylates p53 at serine 37, leading to premature cellular
senescence (8).
Ras homolog enriched in brain (Rheb) is a member of
Ras family (9), serving as the
upstream activator of mammalian target of rapamycin complex 1
(mTORC1) (10). Previous studies have
suggested that Rheb-mTORC1 may be the pivotal axis in regulating
cell growth in coordination with nutrient availability in the
environment (11,12). Rheb is overexpressed in numerous types
of cancer (13) and may be a critical
target for farnesyltransferase inhibitors (FTIs) therapy in
non-small-cell lung cancer cells (14).
A previous study reported that PRAK phosphorylated
Rheb at serine 130 and that this post-translational modification of
Rheb attenuated its guanine nucleotide binding activities, leading
to its inactivation and subsequent cell growth inhibition (15). However, the role of Rheb
phosphorylation in cancer development remains to be elucidated.
Therefore, the aim of the present study was to determine the effect
of Rheb phosphorylation on tumor growth in vitro and in
vivo. In addition, tissue samples were obtained from 70
hepatocellular carcinoma (HCC) patients in order to determine any
associations between Rheb phosphorylation and the
clinicopathological characteristics of patients.
Materials and methods
Materials
Mouse monoclonal antibodies against hemagglutinin
(cat. no. sc-7392), c-Myc (cat. no. sc-40) and HRas (cat.
no. sc-29) were obtained from Santa Cruz Biotechnology Inc.
(Dallas, TX, USA). Anti-phospho-Rheb (S130) antibodies were raised
against an S130 phospho-modified peptide of Rheb by Abmart Co.,
Ltd. (Shanghai, China). Rabbit anti-Rheb polyclonal antibody was
from Proteintech (Wuhan, Hubei, China). Rabbit polyclonal antibody
against p-PRAK(T182) was a gift from Professor Jiahuai Han. Mouse
anti-ribosomal protein S6 monoclonal antibody (cat. no. 2317),
rabbit anti-p-S6 (S235/S236) polyclonal antibody (cat. no. 2211),
rabbit anti-Erk polyclonal antibody (cat. no. 9102) and rabbit
anti-Erk (T202/Y204) polyclonal antibody (cat. no. 4370) were
purchased from Cell Signaling Technology, Inc. (Beverly, MA, USA).
For western blot analysis, antibodies were used at a dilution of
1:3,000 for exogenous proteins and 1:1,000 for endogenous proteins
(if not otherwise indicated).
All mutant constructs of PRAK were created through
polymerase chain reaction (PCR) mutagenesis and were verified by
DNA sequencing. The sense primers used were as follows: for PRAK-KM
mutation, 5′-gaacggtttgcgctgaugattcttcttgatcg-3′; for PRAK-DN,
5′-caaggtgacttgatggcaccccagttcac-3′. The antisense primers used
were the exact reverse complementary sequence of the sense primers.
DNA sequencing was performed by Sangon Biotech (Shanghai,
China).
Cell culture and transfection
Human embryonic kidney 293 (HEK293) cells and mouse
embryonic fibroblast (MEF) were purchased from ATCC (Manassas, VA,
USA). Cells were cultured in Dulbecco's modified Eagle's medium
(DMEM) containing 10% fetal bovine serum (FBS) (Thermo Fisher
Scientific, Inc., Waltham, MA, USA), as previously described
(15). Calcium phosphorylate
transfection was used to transfect DNA into HEK293 cells for
lentiviral package. Lipofectamine 2000 (Thermo Fisher Scientific,
Inc.) was used to transfect DNA into HEK293 for gene
overexpression. Protein expression was determined by western blot
analysis.
In vitro kinase assay
Glutathione S-transferase-Rheb was purified from
BL21(DE3) competent Escherichia coli (New England Biolabs,
Ipswich, MA, USA) and subjected to a kinase assay in kinase buffer
(25 mM Tris, pH 7.5; 10 mM MgCl2; 2 mM DTT; 5 mM
β-glycerophosphate; and 0.1 mM Na3VO4) at
30°C for 30 min. Tris, MgCl2, dithiothreitol,
β-glycerophosphate and Na3VO4 were all from
Sigma-Aldrich (Shanghai, China).
Xenograft assay
PRAK+/+ and PRAK−/− mouse
embryonic fibroblasts (MEFs), HRas and adenovirus early region 1A
(E1A)-expressing lentiviral plasmids were kindly provided by
Professor Jiahuai Han (Xiamen University, Xiamen, China). A tumor
formation assay was performed as previously described (8). Briefly, MEFs were infected with HRas-
and adenovirus E1A-expressing lentiviruses. Lentiviruses were
packaged in HEK293 cells, and at 48 h post-transfection, the cell
media were collected and filtered through a 0.45 µm membrane
(Millipore, Billerica, MA, USA), and then mixed with fresh medium
(1:1) and 4 ng/ml polybrene (Sigma-Aldrich). Transformed MEFs
(2×106) were injected subcutaneously into the flanks of
female immunodeficient athymic nude mice (age, 6–8 weeks; Biomodel
Organisms, Shanghai, China) in 100 µl phosphate-buffered saline
(PBS; Thermo Fisher Scientific, Inc.). Mice were housed under
specific pathogen-free condition, caged individually and given ad
libitum access to food and water. All animal experiments were
conducted with the approval of the Institutional Animal Care and
Use Committee of Fujian Medical University (Fujian, China). Tumors
were measured at least every 2 days without anaesthetizing. Mice
were sacrificed using carbon dioxide for euthanasia when the tumor
diameter was >1 cm.
Patient samples
A total of 70 HCC patients were enrolled in the
present study at the Liver Center of Fujian Province (Mengchao
Hepatobiliary Hospital of Fujian Medical University; Fujian,
China), between January 2011 and July 2012. HCC and corresponding
adjacent normal liver tissue samples were obtained from patients
who underwent hepatectomy surgery. Patients were not included in
the present study if they had previously received preoperative
chemotherapy or radiation therapy. Peritumoral liver tissues were
obtained from regions >3 cm from the tumor site. Immediately
following surgery, tissue samples were fixed in neutral-buffered
formalin and embedded in paraffin (Sinopharm Chemical Reagent,
Shanghai, China) for immunohistochemical studies. HCC diagnoses
were confirmed through pathological studies; in addition, the
peritumoral liver tissue samples were all confirmed to be normal.
Clinical information was collected from patient records. Tumor
stage was determined using the Barcelona Clinic Liver Cancer (BCLC)
staging system (16) and tumor
differentiation was graded according to the Edmondson grading
system (17). The present study was
approved by the Institute Research Ethics Committee of Fujian
Medical University. Written informed consent was obtained from each
patient according to the committee regulations.
Tissue microarrays (TMAs) and
immunohistochemistry
According to the protocol described by Kononen et
al (18) for TMA, a modified
method was developed for the preparation of paraffin TMAs, as
previously described (19). HCC and
corresponding peritumoral tissue from 70 patients was
formalin-fixed and paraffin-embedded. Next, 3 tissue cores
(diameter, 0.75 mm) from each sample block were exhumed by a holing
needle and then arrayed on the recipient block.
Immunohistochemistry was performed using the Elivision™ Plus
two-step system (Maixin Biotech Co., Ltd., Fuzhou, China) according
to the manufacturer's instructions, as previously described
(19). TMA slides were blocked with
10% normal goat serum (Maixin Biotech Co., Ltd.) at 37°C for 30 min
and incubated with a 1:500 dilution of rabbit polyclonal anti-human
phospho-Rheb antibody for 1 h at 37°C followed by three washes with
PBS containing 0.05% Triton X-100 (Beyotime Institute of
Biotechnology, Shanghai, China). The slides were incubated with
polymerized horseradish peroxidase-conjugated anti-mouse/rabbit
immunoglobulin G (Maixin Biotech Co., Ltd.), followed by
3,3′-diaminobenzidine; slides were then counterstained with
hematoxylin (Harbin Green Specimen Technology Development, Harbin,
China). Negative control sections were incubated with pre-immune
serum. The percentage of hepatocytes or tumor cells showing
cytoplasmic staining was scored for each case as previously
described (20,21). Cells were scored broadly, according to
the staining intensity and the distribution of stained cells.
Staining intensities were scored as follows: No staining, 0; weak
staining, 1; moderate staining, 2; and strong staining, 3. The
distribution of stained cells was scored as follows: 0%, 0;
<25%, 1; 25–50%, 2; and >50%, 3. The final staining score was
obtained by adding the scores of staining intensity and
distribution score of stained cells. A score of 0–2 points was
considered negative and scores of 3–6 points were considered
positive. The staining results were evaluated by two independent
pathologists (Professor Shengbing Zang and Miss Xueting Fang) who
were blinded to the clinicopathological features of the samples;
all differences in interpretation were re-evaluated to reach a
consensus. Tissue scoring was performed in triplicate for each
tumor and high levels of homogeneity for staining intensity and
percentage of positive cells were obtained.
Statistical analysis
Statistically significant differences between and
among groups were assessed using the χ2 test with SPSS
17.0 software (SPSS, Inc., Chicago, IL, USA). P<0.05 was
considered to indicate a statistically significant difference
between values.
Results
Rheb phosphorylation was induced
through Ras overexpression in a PRAK-dependent manner
In order to explore the role of Rheb phosphorylation
in the antitumor barrier, it was investigated whether Rheb
phosphorylation was induced by certain oncogenes. Ras
overexpression is known to activate PRAK (8) and PRAK was reported to directly
phosphorylate Rheb at serine 130 (15); therefore, it was hypothesized that
Rheb phosphorylation may be induced by Ras overexpression. The
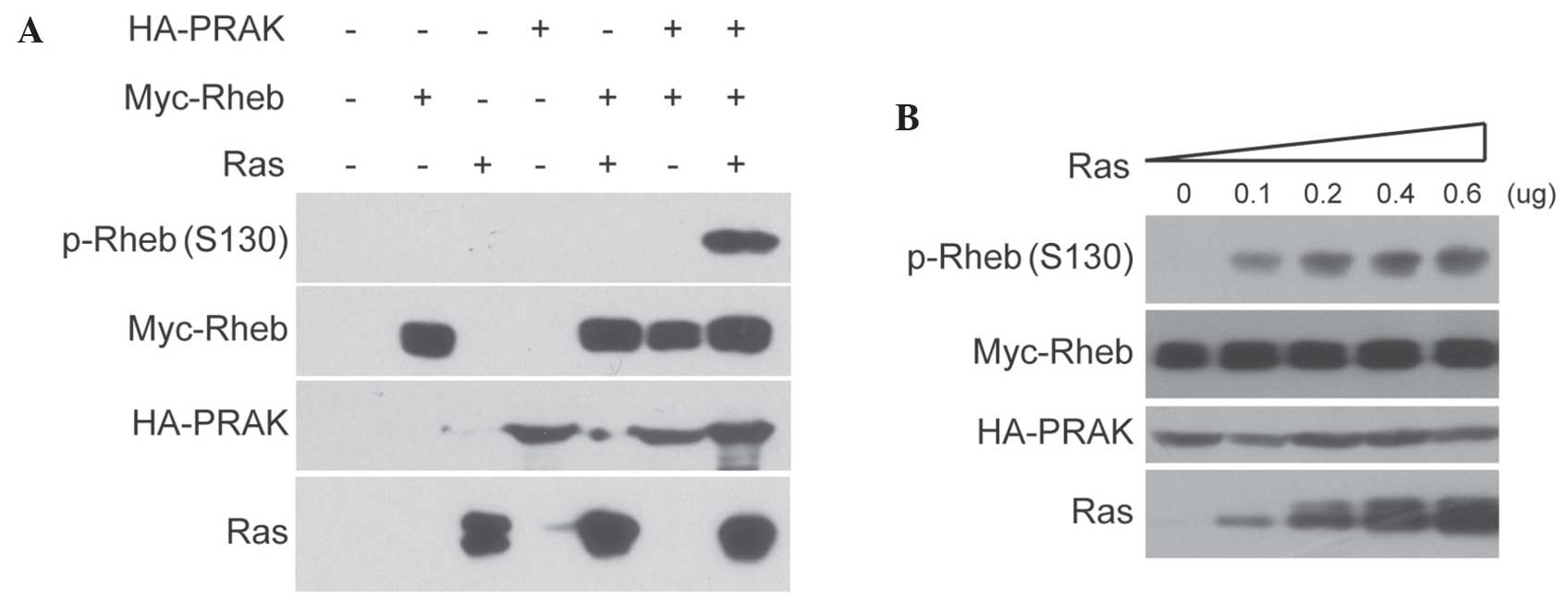
results of the present study confirmed that the co-expression of
Ras with PRAK and Rheb in HEK293 cells markedly increased the
signal of Rheb phosphorylation at serine 130 (Fig. 1A). In addition, the results revealed
that Ras-induced Rheb phosphorylation was dose-dependent (Fig. 1B). By contrast, little Rheb
phosphorylation was detected in cells co-expressing Ras and Rheb
without PRAK. Subsequently, it was investigated whether PRAK
activity was required for Ras-induced Rheb phosphorylation. As
shown in Fig. 2A, kinase-dead PRAK
mutant (PRAK-KM) and dominant-negative PRAK mutant (PRAK-DN) were
able to abolish Ras-induced Rheb phosphorylation. In addition, the
kinase assay demonstrated that PRAK-KM abolished PRAK activity on
Rheb in vitro (Fig. 2B). These
data indicated that Rheb phosphorylation may be induced through Ras
overexpression, which is dependent on PRAK and requires its kinase
activity.
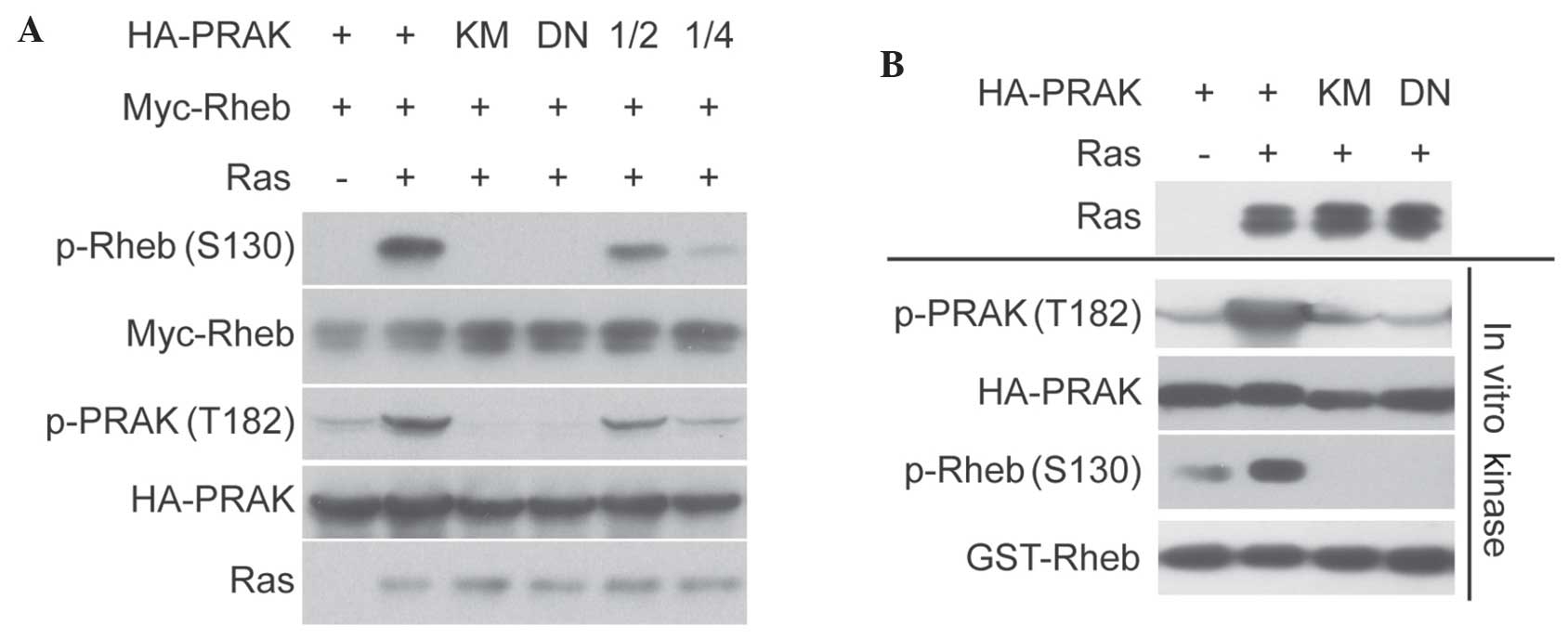 | Figure 2.Rheb phosphorylation induced by Ras
overexpression requires PRAK kinase activity. (A) HA-PRAK,
HA-PRAK-KM or PRAK-DN were co-transfected with Myc-Rheb and HRas
into HEK293 cells. Phosphorylation of Rheb was determined. (B)
HA-PRAK, HA-PRAK-KM or HA-PRAK-DN were co-transfected with HRas
into HEK293 cells. HA-PRAK was then pulled down and incubated with
glutathione S-transferase-Rheb for in vitro kinase assay.
Phosphorylation of Rheb was determined. Rheb, Ras homolog enriched
in brain; PRAK, p38-regulated/activated protein kinase; KM,
kinase-dead mutant HA-PRAK; DN, dominant-negative mutant HA-PRAK;
p-, phosphorylated; HA, hemagglutinin; 1/2, 0.1 µg HA-PRAK for DNA
transfection; 1/4, 0.05 µg HA-PRAK. |
Rheb phosphorylation is involved in
PRAK-mediated tumor suppression
Ras overexpression is a cellular model for tumor
development and PRAK is known to act as tumor suppressor in this
process (8). In addition, Rheb
phosphorylation was reported to attenuate its activity on mTORC1
activation (15), which is pivotal
for tumor growth. Therefore, it was proposed that PRAK-mediated
Rheb phosphorylation may also be associated with PRAK-mediated
tumor suppression. In the present study, PRAK+/+ and
PRAK−/− MEFs were infected with HRas and E1A. Cells were
then injected into nude mice and tumor formation was monitored
periodically (at least every 2 days). The results revealed that
tumor formation of PRAK+/+ MEFs was slower compared with
that of PRAK−/− MEFs (Fig. 3A
and B). In addition, the phosphorylation of Rheb, ribosomal
protein S6 and extracellular signal-regulated kinase (ERK) were
determined in tumor tissues. As shown in Fig. 3C, phosphorylation of Rheb was detected
in tumors derived from PRAK+/+ MEFs, but not in those
from PRAK−/− cells. By contrast, phosphorylation of S6
in PRAK+/+ tumors was decreased compared with that in
PRAK−/− tumors, which supported the hypothesis that Rheb
phosphorylation may inhibit mTORC1 activity. Phosphorylation of
ERK1/2 was unchanged. In conclusion, these data indicated that Rheb
phosphorylation may be involved in PRAK-mediated tumor
suppression.
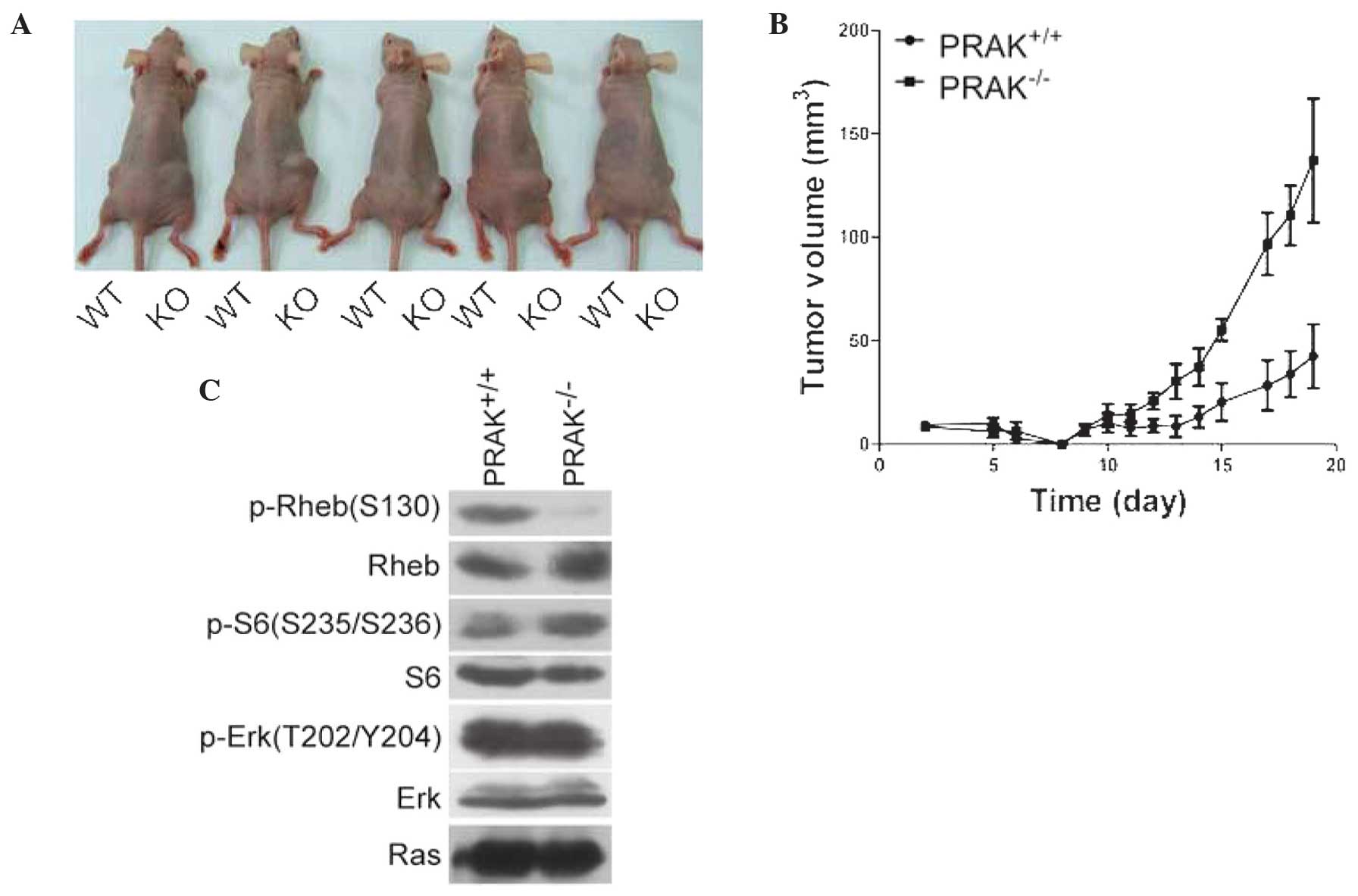 | Figure 3.Rheb phosphorylation is involved in
PRAK-mediated tumor suppression. (A) Primary MEF cells from E13.5
WT (PRAK+/+) and KO (PRAK−/−)embryos were
transduced with HRas and E1A, then inoculated subcutaneously into
six nude mice (left flank, PRAK+/+; right flank,
PRAK−/−). Image of only 5 mice was captured at 24 days
post-inoculation, since 1 mouse died before. (B) Tumor volumes were
monitored daily. (C) Following ~25 days, mice were sacrificed and
tumors were harvested. Phosphorylation of Rheb, S6 and Erk were
determined. Rheb, Ras homolog enriched in brain; PRAK,
p38-regulated/activated protein kinase; MEF, mouse embryonic
fibroblasts; WT, wild type; KO, knock-out; p-, phosphorylated. |
Rheb phosphorylation is compromised in
human HCC tissues
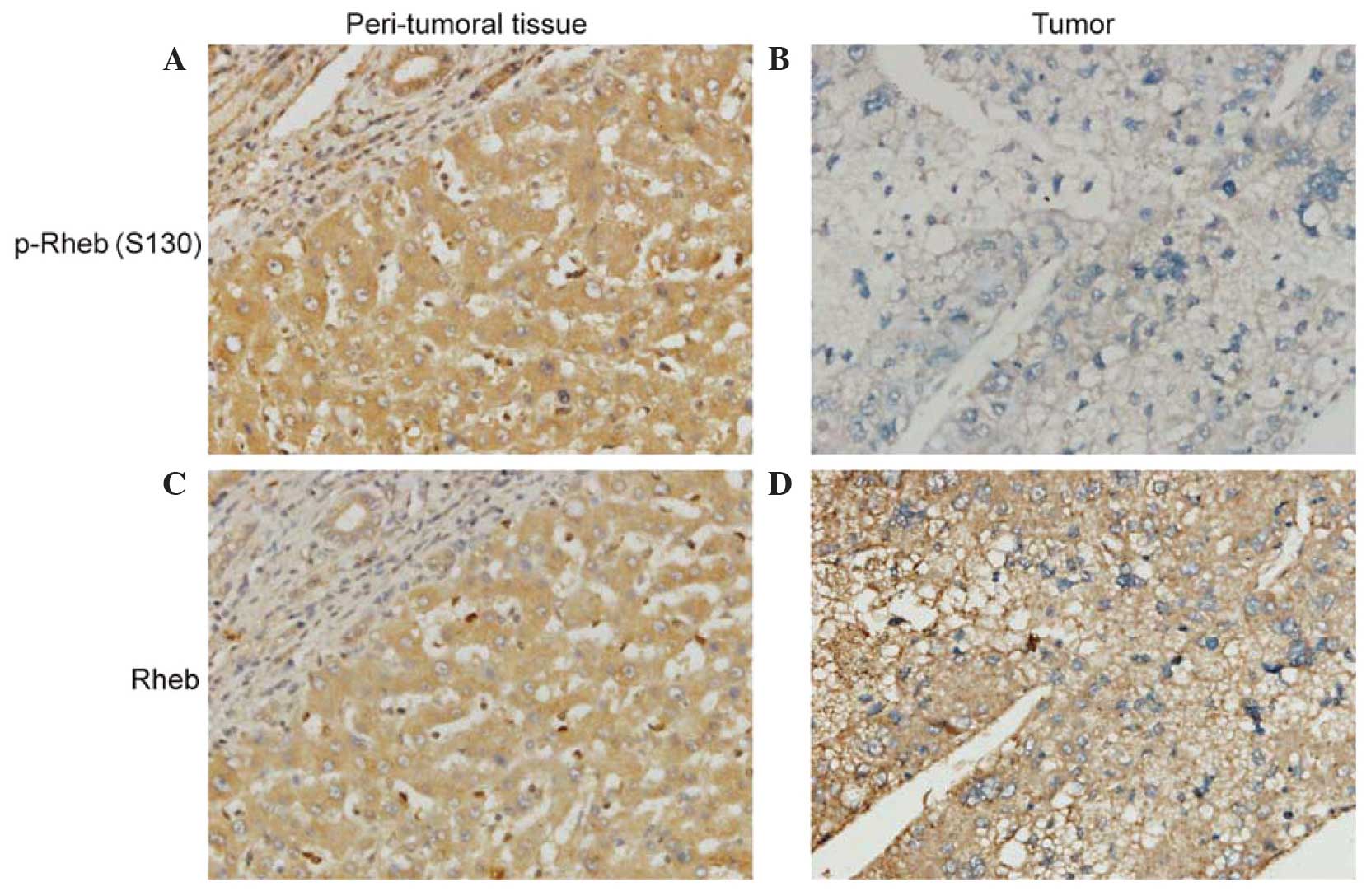
In order to investigate Rheb phosphorylation in
cancer development, immunohistochemistry was performed on TMAs
containing 70 HCC tissue samples and their peritumoral liver tissue
samples in order to assess Rheb phosphorylation levels. Cytoplasmic
staining of Rheb phosphorylation was moderate or strong in
peritumoral liver tissue samples (Fig.
4A), while the tumor cells of certain HCC tissue samples
demonstrated loss of expression of Rheb phosphorylation (Fig. 4B). The presence of Rheb
phosphorylation in HCC tissue samples (62.86%) was significantly
reduced compared with that in peritumoral liver tissue samples
(98.57%) (c2=28.679, P<0.001). In addition, the expression of
Rheb in HCC tissue samples was examined. The results demonstrated
that peritumoral liver tissue samples (Fig. 4C) and HCC tissue samples (Fig. 4D) exhibited moderate or strong
cytoplasmic staining of Rheb. The presence of Rheb in HCC tissue
samples (92.86%) was not significantly different from that in
peritumoral liver tissue samples (98.57%) (c2=2.786, P=0.10).
Furthermore, the association between Rheb phosphorylation and
selected clinicopathological parameters of HCC patients was
evaluated. As shown in Table I, Rheb
phosphorylation did not vary significantly with gender, age, tumor
number, serum α-fetoprotein levels, hepatitis B virus infection,
histological differentiation or the presence of cirrhosis or portal
vein thrombus. However, it was demonstrated that patients with low
Rheb phosphorylation were prone to larger tumors (>5 cm) and
advanced BCLC stage. In conclusion, these data indicated that low
Rheb phosphorylation was associated with poor proliferation and
progression of HCC.
 | Table I.Associations between Rheb
phosphorylation and clinicopathologic parameters. |
Table I.
Associations between Rheb
phosphorylation and clinicopathologic parameters.
|
|
| p-Rheb
expression |
|
|
|---|
|
|
|
|
|
|
|---|
| Variable | n | Positive, n (%) | Negative, n (%) | χ2 | P-value |
|---|
| Gender |
|
|
|
|
|
| Male | 61 | 40 (65.6) | 21 (34.4) | 1.500 | 0.221 |
|
Female | 9 | 4 (44.4) | 5 (55.6) |
|
|
| Age (years) |
|
|
|
|
|
| ≤50 | 33 | 19 (57.6) | 14 (42.4) | 0.746 | 0.388 |
|
>50 | 37 | 25 (67.6) | 12 (32.4) |
|
|
| Tumor size (cm) |
|
|
|
|
|
| ≤5 | 17 | 15 (88.2) | 2 (11.8) | 6.194 | 0.013 |
|
>5 | 53 | 29 (54.7) | 24 (45.3) |
|
|
| Tumor number |
|
|
|
|
|
|
Single | 54 | 34 (63.0) | 20 (37.0) | 0.001 | 0.973 |
|
Multiple | 16 | 10 (62.5) | 6 (37.5) |
|
|
| α-fetoprotein
(ng/ml) |
|
|
|
|
|
|
≤400 | 30 | 21 (70.0) | 9 (30.0) | 1.147 | 0.284 |
|
>400 | 40 | 23 (57.5) | 17 (42.5) |
|
|
| Cirrhosis |
|
|
|
|
|
|
Yes | 48 | 31 (64.6) | 17 (35.4) | 0.195 | 0.659 |
| No | 22 | 13 (59.1) | 9 (40.9) |
|
|
| HBsAg |
|
|
|
|
|
|
Positive | 60 | 36 (60.0) | 24 (40.0) | 1.469 | 0.226 |
|
Negative | 10 | 8 (80.0) | 2 (20.0) |
|
|
|
Differentiation |
|
|
|
|
|
|
Well/moderate | 37 | 25 (67.6) | 12 (32.4) | 0.746 | 0.388 |
|
Poor | 33 | 19 (57.6) | 14 (42.4) |
|
|
| BCLC stage |
|
|
|
|
|
|
0/A | 12 | 12 (100.0) | 0 (0.0) | 8.558 | 0.003 |
|
B/C | 58 | 32 (55.2) | 26 (44.8) |
|
|
| Portal vein
thrombus |
|
|
|
|
|
|
Yes | 39 | 25 (64.1) | 14 (35.9) | 0.059 | 0.809 |
| No | 31 | 19 (61.3) | 12 (38.7) |
|
|
Discussion
Tumor formation is a consequence of oncogene
activation and tumor suppressor inactivation; when activated,
oncogenes drive cell growth and proliferation. In order to maintain
cellular homeostasis, cells develop barriers to tumor development,
most of which are mediated by tumor suppressors (22). Inactivation of tumor suppressors
deregulates intracellular barriers and leads to epigenetic and
metabolic reprogramming in favor of cancer development (1,23).
Unveiling the mechanisms underlying barrier establishment and
deregulation may therefore further current understanding of the
process of tumor development and may aid the development of novel
anticancer therapies.
Ectopic expression of activated Ras was reported to
evoke a fabricated network of intracellular signaling pathways. Ras
promotes cell proliferation through the Ras-phosphoinostide
3-kinase-mTORC1 (24) and
Ras-mitogen-activated protein kinase (25) pathways. In addition, Ras has been
reported to elicit cellular antitumorigenic defenses through the
Ras-PRAK-p53 signaling pathway (8).
Inactivation of PRAK was demonstrated to attenuate its antitumor
barrier, promoting tumor formation (15). In addition to p53, Rheb was also
identified as a substrate of PRAK (15). However, the role of PRAK-mediated Rheb
phosphorylation in cancer development remained to be elucidated.
The present study demonstrated that PRAK-mediated Rheb
phosphorylation was induced by Ras overexpression and that Rheb
phosphorylation may be involved in PRAK-mediated tumor suppression.
Therefore, it was proposed that Rheb phosphorylation may be an
alternative mechanism underlying PRAK-mediated tumor suppression in
addition to p53 activation.
In conclusion, although Rheb overexpression was
observed in skin tumors (13),
prostate cancer (21,26) and lymphomas (27), the present study did not identify any
significant differences in Rheb expression between cancer and
peritumoral tissue in HCC. However, it was revealed that Rheb
phosphorylation levels in tumor tissue samples were markedly
reduced compared with those in peritumoral tissue samples. In
addition, the clinicopathological data indicated that Rheb
phosphorylation was associated with the poor proliferation and
progression of HCC, supporting the hypothesis that Rheb
phosphorylation may be an intracellular barrier to cancer
development.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China (no. 31100995), the
Natural Science Foundation of Fujian Province (no. 2011J01187) and
the Scientific Research Project of Education Department of Fujian
Province (no. 2013B009).
References
|
1
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bae HJ, Chang YG, Noh JH, Kim JK, Eun JW,
Jung KH, Kim MG, Shen Q, Ahn YM, Kwon SH, et al: DBC1 does not
function as a negative regulator of SIRT1 in liver cancer. Oncol
Lett. 4:873–877. 2012.PubMed/NCBI
|
|
3
|
Dimauro T and David G: Ras-induced
senescence and its physiological relevance in cancer. Curr Cancer
Drug Targets. 10:869–876. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Serrano M, Lin AW, McCurrach ME, Beach D
and Lowe SW: Oncogenic ras provokes premature cell senescence
associated with accumulation of p53 and p16INK4a. Cell. 88:593–602.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Braig M, Lee S, Loddenkemper C, Rudolph C,
Peters AH, Schlegelberger B, Stein H, Dörken B, Jenuwein T and
Schmitt CA: Oncogene-induced senescence as an initial barrier in
lymphoma development. Nature. 436:660–665. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ferbeyre G: Barriers to Ras
transformation. Nat Cell Biol. 9:483–485. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
New L, Jiang Y and Han J: Regulation of
PRAK subcellular location by p38 MAP kinases. Mol Biol Cell.
14:2603–2616. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sun P, Yoshizuka N, New L, Moser BA, Li Y,
Liao R, Xie C, Chen J, Deng Q, Yamout M, et al: PRAK is essential
for ras-induced senescence and tumor suppression. Cell.
128:295–308. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yamagata K, Sanders LK, Kaufmann WE, Yee
W, Barnes CA, Nathans D and Worley PF: Rheb, a growth factor- and
synaptic activity-regulated gene, encodes a novel Ras-related
protein. J Biol Chem. 269:16333–16339. 1994.PubMed/NCBI
|
|
10
|
Inoki K, Li Y, Xu T and Guan KL: Rheb
GTPase is a direct target of TSC2 GAP activity and regulates mTOR
signaling. Genes Dev. 17:1829–1834. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Babcock JT and Quilliam LA: Rheb/mTOR
activation and regulation in cancer: Novel treatment strategies
beyond rapamycin. Curr Drug Targets. 12:1223–1231. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wazir U, Newbold RF, Jiang WG, Sharma AK
and Mokbel K: Prognostic and therapeutic implications of mTORC1 and
Rictor expression in human breast cancer. Oncol Rep. 29:1969–1974.
2013.PubMed/NCBI
|
|
13
|
Lu ZH, Shvartsman MB, Lee AY, Shao JM,
Murray MM, Kladney RD, Fan D, Krajewski S, Chiang GG, Mills GB, et
al: Mammalian target of rapamycin activator RHEB is frequently
overexpressed in human carcinomas and is critical and sufficient
for skin epithelial carcinogenesis. Cancer Res. 70:3287–3298. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zheng H, Liu A, Liu B, Li M, Yu H and Luo
X: Ras homologue enriched in brain is a critical target of
farnesyltransferase inhibitors in non-small cell lung cancer cells.
Cancer Lett. 297:117–125. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zheng M, Wang YH, Wu XN, Wu SQ, Lu BJ,
Dong MQ, Zhang H, Sun P, Lin SC, Guan KL, et al: Inactivation of
Rheb by PRAK-mediated phosphorylation is essential for
energy-depletion-induced suppression of mTORC1. Nat Cell Biol.
13:263–272. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Llovet JM, Di Bisceglie AM, Bruix J,
Kramer BS, Lencioni R, Zhu AX, Sherman M, Schwartz M, Lotze M,
Talwalkar J and Gores GJ: Panel of Experts in HCC-Design Clinical
Trials: Design and endpoints of clinical trials in hepatocellular
carcinoma. J Natl Cancer Inst. 100:698–711. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Edmondson HA and Steiner PE: Primary
carcinoma of the liver: A study of 100 cases among 48,900
necropsies. Cancer. 7:462–503. 1954. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kononen J, Bubendorf L, Kallioniemi A,
Bärlund M, Schraml P, Leighton S, Torhorst J, Mihatsch MJ, Sauter G
and Kallioniemi OP: Tissue microarrays for high-throughput
molecular profiling of tumor specimens. Nat Med. 4:844–847. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shengbing Z, Feng LJ, Bin W, Lingyun G and
Aimin H: Expression of KiSS-1 gene and its role in invasion and
metastasis of human hepatocellular carcinoma. Anat Rec (Hoboken).
292:1128–1134. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Eom M, Han A, Lee MJ and Park KH:
Expressional difference of RHEB, HDAC1, and WEE1 proteins in the
stromal tumors of the breast and their significance in
tumorigenesis. Korean J Pathol. 46:324–330. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kobayashi T, Shimizu Y, Terada N, Yamasaki
T, Nakamura E, Toda Y, Nishiyama H, Kamoto T, Ogawa O and Inoue T:
Regulation of androgen receptor transactivity and mTOR-S6 kinase
pathway by Rheb in prostate cancer cell proliferation. Prostate.
70:866–874. 2010.PubMed/NCBI
|
|
22
|
Prieur A and Peeper DS: Cellular
senescence in vivo: A barrier to tumorigenesis. Curr Opin Cell
Biol. 20:150–155. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Menendez JA and Alarcón T: Metabostemness:
a new cancer hallmark. Front Oncol. 4:2622014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shaw RJ and Cantley LC: Ras, PI(3)K and
mTOR signalling controls tumour cell growth. Nature. 441:424–430.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Aksamitiene E, Kiyatkin A and Kholodenko
BN: Cross-talk between mitogenic Ras/MAPK and survival PI3K/Akt
pathways: A fine balance. Biochem Soc Trans. 40:139–146. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nardella C, Chen Z, Salmena L, Carracedo
A, Alimonti A, Egia A, Carver B, Gerald W, Cordon-Cardo C and
Pandolfi PP: Aberrant Rheb-mediated mTORC1 activation and Pten
haploinsufficiency are cooperative oncogenic events. Genes Dev.
22:2172–2177. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mavrakis KJ, Zhu H, Silva RL, Mills JR,
Teruya-Feldstein J, Lowe SW, Tam W, Pelletier J and Wendel HG:
Tumorigenic activity and therapeutic inhibition of Rheb GTPase.
Genes Dev. 22:2178–2188. 2008. View Article : Google Scholar : PubMed/NCBI
|