Introduction
Glioma, which occurs in the central nervous system
(CNS), is the most common primary malignant tumor, with an
unacceptably poor prognosis despite the application of a variety of
treatments, including surgery, radiotherapy and chemotherapy
(1). In recent years, accumulated
data have established that the immune network may be involved in
regulating glioma development and growth (2). Moreover, tumor cells have been killed
following direct immunization experiments using vaccines in mice,
indicating its role in specific protective immunity and tumor
destruction mediated by immune cells, such as microglia/macrophages
(3). These immunotherapies exert a
highly specific, long-term fatal effect on tumor cells by
stimulating and supplementing the body's antitumor immunity
(4,5),
with only minimal adverse reactions (6). However, in the past decade, the main
debate has focused on whether immunity exhibits an antitumor role
in the CNS or functions as a tumor growth promoter (7). Additionally, the functions and mechanism
of different immune cells in glioma require further
investigation.
Various pathological stimuli, including brain injury
(8), neurodegeneration (9), infection, inflammation (10) and brain tumors (11), can activate brain
macrophages/microglia. It has been observed that in CNS tumors,
activated microglia/macrophages can engulf tissue debris and
secrete growth-promoting factors, leading to regeneration (12). By contrast, the overactivation of
microglia/macrophages will lead to tissue damage in the CNS
(13). Following activation,
microglia/macrophages change their morphology, upregulate the
expression of certain membrane proteins and produce certain
cytokines, resulting in inflammation and tissue loss (14). The contrast in roles played by
activated microglia/macrophages may be due to the existence of
different subpopulations of microglia/macrophages (15).
Endothelial monocyte-activating polypeptide II
(EMAPII)+ macrophages secrete proinflammatory and
antiangiogenic cytokine in the parenchyma of the CNS, stimulate the
accumulation and cytokine production of macrophages, and trigger
the apoptosis of endothelial cells (16). Increased EMAPII expression is
considered to be a sensitive marker of neurotoxic lesions in rat
brains (17). ED1, which is mainly
found in phagocytosing macrophages and reactive microglia, is a
lysosomal membrane protein. Macrophages with ED1 can be observed in
the majority of brain tumors, as well as in gliomas (18). Cluster of differentiation 8 (CD8) was
traditionally recognized as a marker that is mainly expressed in
cytotoxic T cells and natural killer cells (19). More recently, studies have reported
the accumulation of CD8+ macrophages in various CNS
pathologies, such as ischemia, Wallerian degeneration and spinal
cord injury (19,20). CD8+ cells, no matter
whether lymphocytes or macrophages, may have important distinctive
functions in glioma. However, information on the role played by
different subsets of macrophages/microglia in glioma is relatively
lacking.
Further understanding of immune-tumor interactions
will benefit tumor therapies. Moreover, it has been demonstrated
that microglial cells are found in necrotic and intact areas of
brain tumor tissues (21). Thus, in
the present study, multiple antigens were used to study the
distribution and characteristics of tumor-associated
macrophages/microglia in rat C6 glioma.
Materials and methods
Rat brain tissue libraries
The C6 glioma cell-induced rat brain tissue was a
gift from Professor Hermann J. Schluesener (Institute of Brain
Research, Tübingen University, Tübingen, Germany). The C6 glioma
cells were injected into the brains of Sprague-Dawley rats, as
previously reported (22).
Immunohistochemistry
Brain sections were dewaxed and then boiled for 15
min in citrate buffer (2.1 g sodium citrate/l; pH 6) in a 600-W
microwave oven. 1% H2O2 in pure methanol was
used to inhibit endogenous peroxidase activity for 15 min.
Following blocking with 10% normal pig serum (Biochrom, Berlin,
Germany) for 15 min, the rat brain sections (0.5 µm) were incubated
with the following mouse monoclonal antibodies overnight at 4°C:
OX8 (1:100; AbD Serotec, Oxford, UK), ED1 (1:100; AbD Serotec),
EMAPII (1:100; BMA Biomedicals, Augst, Switzerland), W3/13 (1:100;
AbD Serotec), OX6 (1:100; AbD Serotec), OX62 (1:50; AbD Serotec) or
OX22 (1:100; AbD Serotec). Subsequently, the sections were
incubated with biotinylated swine anti-rabbit (DAKO, Hamburg,
Germany) immunoglobulin (Ig)G F(ab)2 antibody fragment or a
biotinylated rabbit anti-mouse IgG F(ab)2 antibody fragment (Dako,
Hamburg, Germany). Following washing with phosphate-buffered saline
with Tween 20 (Beijing SHX Biotechnology Co., Ltd., Beijing, China)
and further incubation with a Streptavidin-Avidin-Biotin complex
(Dako) for 30 min at room temperature, the sections were developed
with diaminobenzidine substrate (Fluka, Neu-Ulm, Germany) for
visualization. Finally, the tissue sections were counterstained
with hematoxylin. With regard to the negative controls, the primary
antibodies were omitted. Hematoxylin and eosin staining was
additionally applied to evaluate the lesions of the brain trauma
sections.
Light microscopic examination of
immunostained brain sections
The immunostaining was examined and the numbers of
positive cells at the brain tumor areas were quantified. For each
section, the numbers of OX8+, ED1+,
EMAPII+ or W3/13+ cells were counted in 5
non-overlapping high-power fields (HPF; ×400 magnification). Areas
around the pannecrotic border that had a maximum number of positive
cells were used as the selected HPF. Only positive cells with the
nucleus at the focal plane were counted, while positive
perivascular cells were not counted in each field studied. Counting
was performed by individuals who were blinded to the treatment. The
results of the numbers of positive cells per HPF are presented as
the arithmetic mean ± standard error of the mean.
Statistical analysis
Graph Pad Prism 4.0 (GraphPad Software, Inc., La
Jolla, CA, USA) was used for the statistical analysis. In-group and
between-group differences were studied by one-way analysis of
variance followed by Dunnett's multiple comparisons test. P<0.05
was used to indicate a statistically significant difference.
Results
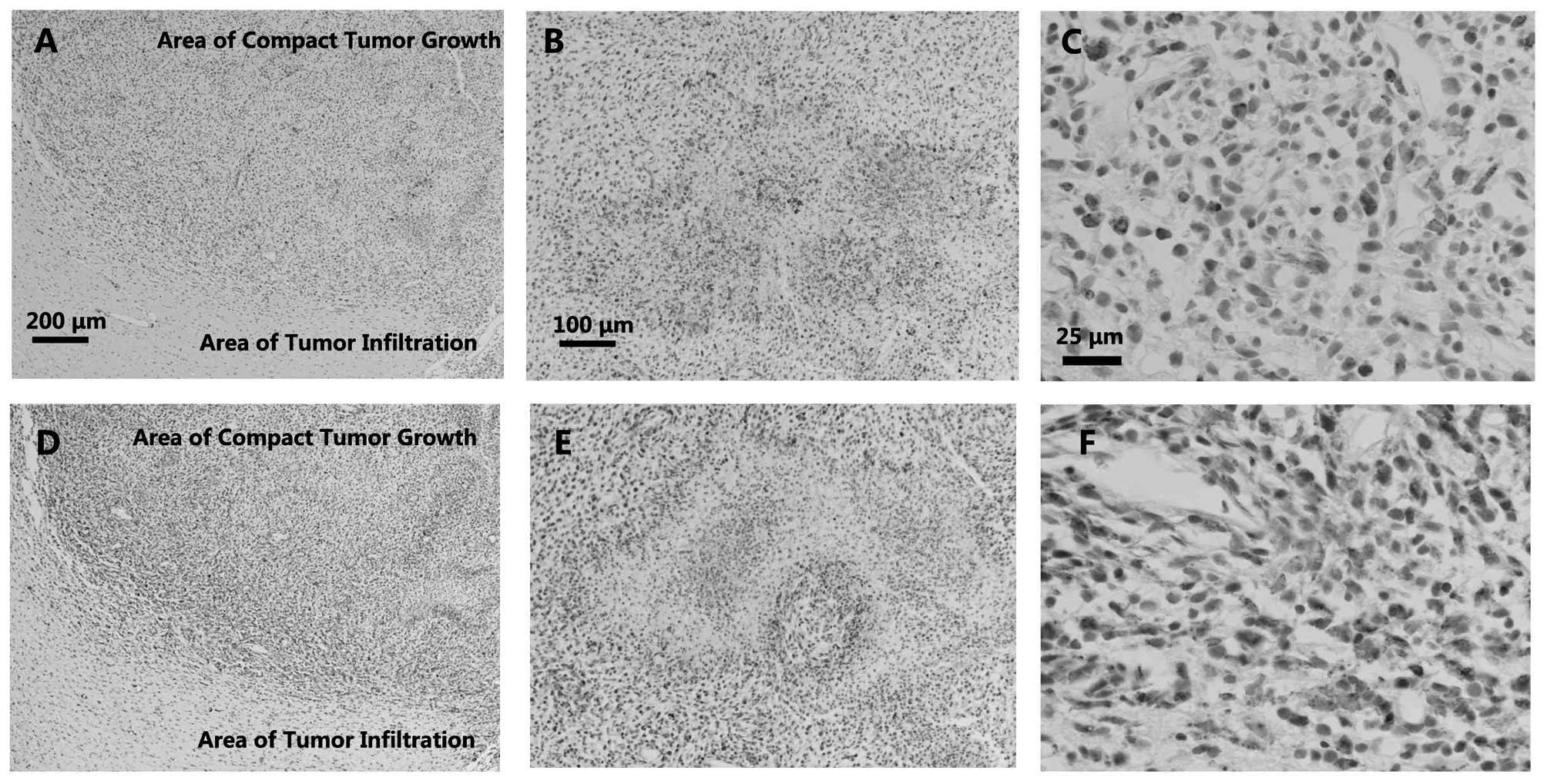
Activated microglia/macrophage
accumulation in brains of the rat C6 glioma model
The infiltration of reactive microglia/macrophages
was analyzed in rat C6 gliomas in this study by
immunohistochemistry with the antibodies ED1 and EMAPII. The ED1
antibody stains CD68, which is mainly found in phagocytosing
macrophages and reactive microglia (23). The upregulated expression of CD68
usually indicates an increase in phagocytical activity of the
marked cells. In the C6 gliomas, little infiltration of
ED1+ cells was observed in the areas of infiltrated
tumor growth (Fig. 1A). Slight
accumulation of ED1+ cells was observed in the border
areas (Fig. 1A), while relatively
strong infiltration was detected inside the areas of pannecrosis
(Fig. 1B) and in the areas of compact
tumor growth (Fig. 1A and C).
EMAPII, which is expressed by activated
microglial/macrophages in the CNS parenchyma, is a proinflammatory
cytokine (16). The expression of
EMAPII is considered a sensitive marker of microglia activation. In
the present study, similar to ED1 staining, an accumulation of
EMAPII+ cells was observed in the C6 tumor areas
(Fig. 1D and F). However, the number
of EMAPII+ cells was greater than that of the
ED1+ cells, and the EMAPII+ cells were
located more at the borders of the pannecrosis rather than inside
(Fig. 1E).
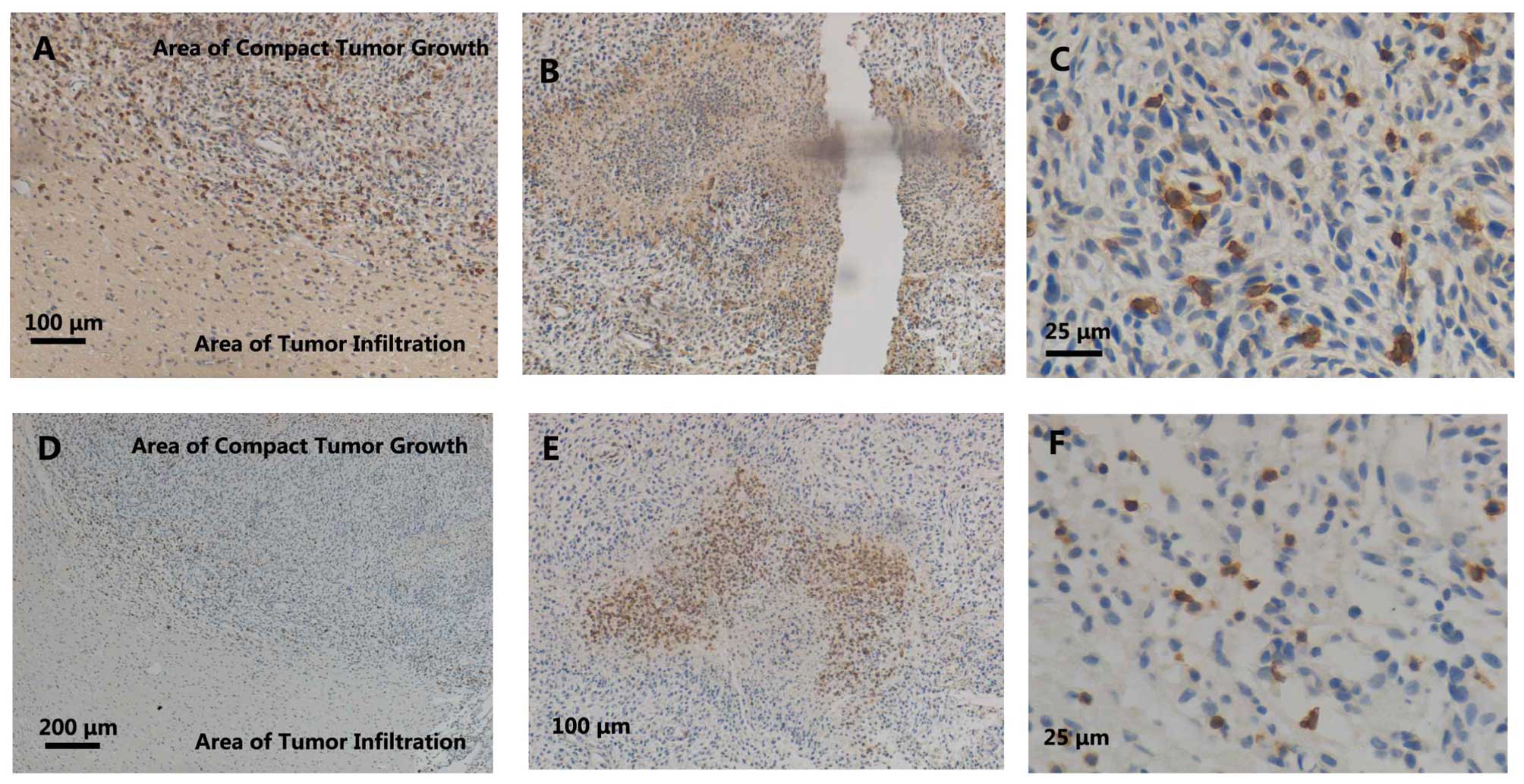
Single immunohistochemical staining of
CD8 in brains of the rat C6 glioma model
Next, CD8 expression was detected in the rat C6
glioma brains by immunohistochemistry. Little CD8 expression was
observed in the areas of infiltrating tumor growth (Fig. 2A). Slight infiltration of
CD8+ cells was observed in the border areas (Fig. 2A), while a relatively strong
accumulation was detected in areas of compact tumor growth
(Fig. 2A and C). Moreover, the
accumulation of CD8+ cells in perivasculature areas was
observed (Fig. 2D). Furthermore, a
strong infiltration of CD8+ cells was observed in areas
of compact tumor growth and at the edge of the pannecrosis
(Fig. 2B). However, inside the
pannecrosis areas (Fig. 2B) and in
normal neuronal areas (Fig. 2E), much
fewer CD8+ cells were observed. Morphologically, the
majority of the accumulated CD8+ cells were similar to
macrophage phenotypes, with amoeboid morphological characteristics
(Fig. 2C).
ED1 and EMAPII label activated
microglia/macrophages, and in the present study, the time course of
accumulation, the morphology and the distribution of
CD8+ cells were similar to those of the ED1+
and EMAPII+ cells. Similar to the majority of
ED1+ and EMAPII+ cells, the CD8+
cells showed amoeboid morphological characteristics in the
pannecrosis (Figs. 1C, 1F, 2C and 2D).
Moreover, the time course of CD8+ cell infiltration in
the tumor areas was parallel to that of the ED1+ and
EMAPII+ cells. In addition, the ED1+ and
EMAPII+ cells were located inside (Fig. 2B) and at the border (Fig. 2E) of pannecrosis areas, which was
similar for the CD8+ cells (Fig. 2B–D).
T cells accumulation in the brains of
the rat C6 glioma model
Since CD8+ is traditionally observed in T
cells, the present study further detected T cell accumulation in
the rat C6 gliomas using a pan-T-cell marker, W3/13. In contrast to
the CD8+ cells (Figs. 1A,
1D and 2A), the infiltration of
the W3/13+ cells was only seldomly observed in the
borders of the compact tumor growth areas (Fig. 2A). Furthermore, the W3/13+
cells exhibited smaller cell bodies (Fig.
2C) compared with the CD8+ cells (Fig. 2C). Therefore, the time course of
accumulation, morphology and distribution all suggested that the
major cellular source of CD8 was reactive macrophages/microglia,
but not T cells.
Discussion
In the present study, the early accumulation,
distribution and characteristics of CD8+,
ED1+, EMAPII+ and W3/13+ cells in
rat C6 glioma brains were analyzed. Significant ED1+
cell accumulation was observed mostly located to areas of compact
tumor growth and notably distributed inside the pannecrosis.
EMAPII+ cell accumulation was also detected mainly in
the areas of compact tumor growth, but only at the borders of the
pannecrosis. It is noteworthy that the morphology and distribution
of the CD8+ cells were similar to those of the reactive
ED1+ and EMAPII+ macrophages and microglia.
However, the morphology and localization of the CD8+
cells were different from those of the few infiltrating
W3/13+ T cells. These results were expected as they were
the same as those found previously (24). Thus, this observation means that it is
reasonable to speculate that the CD8+ cells were also
ED1+ and EMAPII+ macrophages/microglia. The
heterogeneity of activated macrophages and microglia in rat C6
gliomas represent diverse subpopulations with different but
relative functions, which may play a crucial role in the growth of
glioma.
EMAPII is a well-known proinflammatory and
antiangiogenic cytokine, which is expressed by activated
macrophages/microglia in the parenchyma of the CNS (16). The pathophysiological expression of
EMAPII induces the infiltration and activation of macrophages and
endothelial apoptosis (16).
Increased EMAPII expression is regarded as a sensitive marker of
neurotoxic lesions in the rat brain (17). ED1 is an indication of activated
macrophages expressed in the majority of tumor-associated
macrophages (25). The detection of
these two subsets may indicate a positive role in the formation of
necrosis.
CD8 has been classically viewed as a marker of a
certain subpopulation of lymphocytes (19). More recently, the accumulation of
CD8+ macrophages has been observed in several
pathologies of the CNS, such as ischemia (26), Wallerian degeneration (27), experimental autoimmune
encephalomyelitis (26), bornavirus
encephalitis (28), spinal cord
injury (20) and glomerulonephritis
(29). In the present study, nearly
all CD8+ cells shared a similar morphology and
distribution with reactive ED1+ and EMAPII+
macrophages and/or microglia, but were not similar to the few
infiltrating W3/13+ T cells. This similar morphology and
distribution of reactive CD8+, ED1+ and
EMAPII+ cells suggests that the observed reactive
microglia/macrophages were CD8+. Moreover, the
presumable CD8+ macrophages/microglia were mainly
observed in areas of the compact tumor growth and concentrated at
the boundary of the pannecrosis. The presumable CD8+
macrophages/microglia have been observed to be limited to regions
of cavitation lesions following cerebral ischemia (26) and spinal cord injury (20). The restricted distribution arrangement
of CD8+ phagocytes in rat C6 glioma indicates a crucial
role in regulating responses in tumor necrosis.
The contributions of the CD8+
microglia/macrophages to the development of CNS damage are not
fully understood. In alveolar macrophages, the ligation of CD8
molecules resulted in the production of the inducible nitric oxide
synthase, nitric oxide, tumor necrosis factor (TNF)-α and
interleukin-1β, and enhanced the cytotoxicity to Leishmania
major (30). Furthermore, the
infiltration of CD8+ microglia/macrophages was mainly
located to severe inflammatory destruction tissue in experimental
autoimmune encephalomyelitis, suggesting their contribution to the
pathological process (26).
In a more traditional view, OX8 is the marker of a
subtype of activated T cells, which can also be observed in glioma
(24). In the present study, the
observation of W3/13+ cells confirmed the existence of T
cells in glioma, which was in accordance with the results of other
studies (24,31). It has been shown that peripheral
blood-derived CD4+ and CD8+ T cells may
mediate the cytotoxicity of human gliomas via a mechanism
independent of TNF in vitro (31). The observation of T cells inside the
necrosis area in the present study suggests its role in the
induction of necrosis. However, OX8 showed neither a similar
distribution nor morphology to the detected W3/13+
cells. This may be attributed to the time course of the innate and
adaptive immune reaction. Further study is required to find the
underling mechanism.
In summary, in the present study, the early
accumulation of diverse subsets of activated macrophages/microglia,
including reactive ED1+ and EMAPII+ cells,
was observed in rat C6 gliomas. Notably, the CD8+ cells
shared a similar distribution and morphology with these two
subpopulations rather than the W3/13+ T cells. The
abundance of this subpopulation and their strategic location
highlights the possible comprehensive roles of these cells in
glioma development, and the orchestration of active macrophages
could be a potential research area for studying the development of
immunotherapeutic antitumor agents in glioma.
Acknowledgements
The authors would like to thank Professor H. J.
Schluesener for providing the C6 glioma rat brain tissue library
and for assistance with immunostaining. This study was supported by
the National Nature Science Foundation of China (grant no.
31170851).
Abbreviations:
|
CNS
|
central nervous system
|
|
EMAPII
|
endothelial monocyte-activaing
polypeptide II
|
|
CD8
|
cluster of differentiation 8
|
References
|
1
|
Jovčevska I, Kočevar N and Komel R: Glioma
and glioblastoma- how much do we (not) know? Mol Clin Oncol.
1:935–941. 2013.PubMed/NCBI
|
|
2
|
Platten M, Ochs K, Lemke D, Opitz C and
Wick W: Microenvironmental clues for glioma immunotherapy. Curr
Neurol Neurosci Rep. 14:4402014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Glass R and Synowitz M: CNS macrophages
and peripheral myeloid cells in brain tumours. Acta Neuropathol.
128:347–362. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jackson CM, Lim M and Drake CG:
Immunotherapy for Brain Cancer: Recent progress and future promise.
Clin Cancer Res. 20:3651–3659. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sloan AE, Dansey R, Zamorano L, Barger G,
Hamm C, Diaz F, Baynes R and Wood G: Adoptive immunotherapy in
patients with recurrent malignant glioma: Preliminary results of
using autologous whole-tumor vaccine plus granulocyte-macrophage
colony-stimulating factor and adoptive transfer of
anti-CD3-activated lymphocytes. Neurosurg Focus. 9:e92000.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Atkinson LL, Merchant RE, Ghatak NR and
Young HF: Sterile abscesses in glioma patients treated by
intraparenchymal injection of lymphokine-activated killer cells and
recombinant interleukin-2: Case reports. Neurosurgery. 25:805–810.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Badie B and Schartner J: Role of microglia
in glioma biology. Microsc Res Tech. 54:106–113. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lerch JK, Puga DA, Bloom O and Popovich
PG: Glucocorticoids and macrophage migration inhibitory factor
(MIF) are neuroendocrine modulators of inflammation and neuropathic
pain after spinal cord injury. Semin Immunol. 26:409–414. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Doens D and Fernández PL: Microglia
receptors and their implications in the response to amyloid β for
Alzheimer's disease pathogenesis. J Neuroinflammation. 11:482014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Onore CE, Schwartzer JJ, Careaga M, Berman
RF and Ashwood P: Maternal immune activation leads to activated
inflammatory macrophages in offspring. Brain Behav Immun.
38:220–226. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou W and Bao S: Reciprocal supportive
interplay between glioblastoma and tumor-associated macrophages.
Cancers (Basel). 6:723–740. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Streit WJ: Microglia as neuroprotective,
immunocompetent cells of the CNS. Glia. 40:133–139. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kempermann G and Neumann H: Neuroscience.
Microglia: The enemy within? Science. 302:1689–1690. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rock RB, Gekker G, Hu S, Sheng WS, Cheeran
M, Lokensgard JR and Peterson PK: Role of microglia in central
nervous system infections. Clin Microbiol Rev. 17:942–964. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ladeby R, Wirenfeldt M, GarciaOvejero D,
Fenger C, DissingOlesen L, Dalmau I and Finsen B: Microglial cell
population dynamics in the injured adult central nervous system.
Brain Res Brain Res Rev. 48:196–206. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mueller CA, Schluesener HJ, Conrad S,
Meyermann R and Schwab JM: Lesional expression of a proinflammatory
and antiangiogenic cytokine EMAP II confined to endothelium and
microglia/macrophages during secondary damage following
experimental traumatic brain injury. J Neuroimmunol. 135:1–9. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Brabeck C, Michetti F, Geloso MC, Corvino
V, Goezalan F, Meyermann R and Schluesener HJ: Expression of
EMAP-II by activated monocytes/microglial cells in different
regions of the rat hippocampus after trimethyltin-induced brain
damage. Exp Neurol. 177:341–346. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fang KM, Wang YL, Huang MC, Sun SH, Cheng
H and Tzeng SF: Expression of macrophage inflammatory protein-1α
and monocyte chemoattractant protein-1 in glioma-infiltrating
microglia: Involvement of ATP and P2X7 receptor. J
Neurosci Res. 89:199–211. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gupta S: Role of dendritic cells in innate
and adaptive immune response in human aging. Exp Gerontol.
54:47–52. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Popovich PG, van Rooijen N, Hickey WF,
Preidis G and McGaughy V: Hematogenous macrophages express CD8 and
distribute to regions of lesion cavitation after spinal cord
injury. Exp Neurol. 182:275–287. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang Z, Artelt M, Burnet M, Trautmann K
and Schluesener HJ: Early infiltration of CD8+
macrophages/microglia to lesions of rat traumatic brain injury.
Neuroscience. 141:637–644. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Grobben B, De Deyn PP and Slegers H: Rat
C6 glioma as experimental model system for the study of
glioblastoma growth and invasion. Cell Tissue Res. 310:257–270.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Damoiseaux JG, Döpp EA, Calame W, Chao D,
MacPherson GG and Dijkstra CD: Rat macrophage lysosomal membrane
antigen recognized by monoclonal antibody ED1. Immunology.
83:140–147. 1994.PubMed/NCBI
|
|
24
|
Perrin G, Schnuriger V, Quiquerez AL, Saas
P, Pannetier C, de Tribolet N, Tiercy JM, Aubry JP, Dietrich PY and
Walker PR: Astrocytoma infiltrating lymphocytes include major T
cell clonal expansions confined to the CD8 subset. Int Immunol.
11:1337–1350. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yamashiro S, Takeya M, Nishi T, Kuratsu J,
Yoshimura T, Ushio Y and Takahashi K: Tumor-derived monocyte
chemoattractant protein-1 induces intratumoral infiltration of
monocyte-derived macrophage subpopulation in transplanted rat
tumors. Am J Pathol. 145:856–867. 1994.PubMed/NCBI
|
|
26
|
Schroeter M, Stoll G, Weissert R, Hartung
HP, Lassmann H and Jander S: CD8+ phagocyte recruitment in rat
experimental autoimmune encephalomyelitis: Association with
inflammatory tissue destruction. Am J Pathol. 163:1517–1524. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Stoll G, Jander S and Myers RR:
Degeneration and regeneration of the peripheral nervous system:
From Augustus Waller's observations to neuroinflammation. J
Peripher Nerv Syst. 7:13–27. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Weissenböck H, Hornig M, Hickey WF and
Lipkin WI: Microglial activation and neuronal apoptosis in
Bornavirus infected neonatal Lewis rats. Brain Pathol. 10:260–272.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Marc T: Brain development and the immune
system: An introduction to inflammatory and infectious diseases of
the child's brain. Handb Clin Neurol. 112:1087–1089. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hirji N, Lin TJ, Bissonnette E, Belosevic
M and Befus AD: Mechanisms of macrophage stimulation through CD8:
Macrophage CD8alpha and CD8beta induce nitric oxide production and
associated killing of the parasite Leishmania major. J Immunol.
160:6004–6011. 1998.PubMed/NCBI
|
|
31
|
Baltuch GH, Villemure JG, McCrea E and
Antel JP: T cell-mediated cytotoxicity of human gliomas: A tumor
necrosis factor-independent mechanism. Neurosurgery. 35:450–456.
1994. View Article : Google Scholar : PubMed/NCBI
|