Introduction
The p16INK4a protein (p16) is a tumor
suppressor protein that functions as an inhibitor of
cyclin-dependent kinase (CDK) 4 and CDK6, the CDKs that initiate
the phosphorylation of the retinoblastoma protein (pRb) (1). Thus, p16 has the capacity to arrest the
cell cycle at the G1 phase through the cyclin D-CDK4-Rb pathway
(2).
It has been reported that the expression of p16 is
regulated primarily at the transcriptional level, and various types
of transcription factors, including Sp1 and Ets1, are involved in
its transcriptional regulation (2,3). In
previous studies, the histone acetyltransferase p300 has been found
to be involved in the activation of p16 expression through
recruitment by Sp1, while histone deacetylases 3 and 4 inhibited
the activity of the p16 promoter through Yin Yang 1 and
zinc-binding protein-89 (4–6).
In addition, it has been reported that
phosphorylation of p16 protein at Ser152 promotes the association
between p16 and CDK4, while phosphorylation at Ser8 abolished the
CDK4-inhibitory activity of p16 (7).
A previous study identified that hypomethylation of p16 protein
exhibited a potentiated function in preventing cell proliferation
(8). These results indicated that the
post-translational modification of p16 plays an important role in
the regulation of the function of p16.
Arginine residues are methylated by protein arginine
methyltransferases (PRMTs). Type I PRMTs include PRMT1, PRMT3,
coactivator-associated arginine methyltransferase 1, also termed
PRMT4, PRMT6 and PRMT8, which catalyze the formation of
ω-monomethylarginine and asymmetric dimethylarginine (9,10). In a
previous study, methylation at specific arginine residues of the
p16 protein by PRMT6 was found to be critical for the activity of
p16 (8). However, the existence of
crosstalk between arginine methylation and phosphorylation of p16
has not been elucidated.
The present study aimed to determine the role of
phosphorylation and arginine methylation in the functional
regulation of p16, and identify potential crosstalk between these
mechanisms.
Materials and methods
Cell culture and transient
transfection
The HeLa, BT549, 293T and A549 cell lines (Cell Bank
of Shanghai Institutes for Biological Sciences, Chinese Academy of
Sciences, Shanghai, China) were cultured in appropriate media
supplemented with 10% fetal bovine serum, 100 U/ml penicillin and
100 µg/ml streptomycin (Sigma-Aldrich, St Louis, MO, USA), and
maintained in a humidified atmosphere containing 5% CO2
at 37°C. For the transient transfection of HeLa and A549 cells, the
cells were seeded into six-well plates and cultured for 1 day.
Transient transfection was then performed using Lipofectamine 2000
(Invitrogen Life Technologies, Carlsbad, CA, USA). Subsequent to 48
h, the cells were harvested.
Plasmids
The human plasmid expressing p16-EGFP-N1 was
provided by Dr Jun Chen (New York Medical College, Valhalla, NY,
USA). The specific site mutations for the four serine residues were
introduced into the p16 cDNA region using a two-step polymerase
chain reaction (PCR) procedure, as previously described (11). Two simultaneous PCR reactions were
performed using p16-EGFP-N1 as a template. Amplified fragments from
each PCR reaction were purified, mixed and subjected to a second
round of PCR using two external primers. The mutagenic sequence for
the Ser7 and 8 residues was AGC (Serine) mutated to GCC (Alanine).
For the Ser140 residues, the sequence was AGT (Serine) mutated to
GCT (Alanine). For the Ser152 residues, the sequence was TCA
(Serine) mutated to GCA (Alanine). The amplified PCR products were
inserted into the HindIII and BamHI sites of the EGFP-N1 vector,
and the correct insertion was verified by DNA sequencing. The
arginine mutations in the p16 vectors were previously described
(8). The PRMT1-myc plasmid was
provided by Professor Mark A. Wainberg (McGill AIDS Centre, Lady
Davis Institute, Jewish General Hospital, Montréal, QC, Canada)
(12). Plasmids expressing human
PRMT4 were provided by Professor Mark T. Bedford (The University of
Texas MD Anderson Cancer Center, Science Park, Department of
Molecular Carcinogenesis, Smithville, TX, USA) (13). PRMT6 plasmids were provided by Dr
Stephane Richard (Lady Davis Institute) (14).
Western blotting and
co-immunoprecipitation (CoIP)
Exogenous expression of p16 or PRMTs was detected by
western blotting. The A549 cells were harvested 48 h subsequent to
transfection with the relevant plasmids. In total, 1×106
cells were digested and lysed with lysis buffer consisting of 50 mm
Tris/HCl, 1% Nonidet P (NP)-40, 150 mm NaCl, 1 mm EDTA and 1 mm
phenylmethanesulfonyl fluoride for 30 min at 4°C. Total cell
extracts were separated using 12% SDS-PAGE, and then transferred to
polyvinylidene fluoride (PVDF) membranes (EMD Millipore, Billerica,
MA, USA). The membranes were incubated with the following
antibodies: Polyclonal rabbit anti-human PRMT6 (catalog no. P6495;
Sigma-Aldrich, St Louis, MO, USA; dilution, 1:1,000); polyclonal
rabbit anti-human PRMT1 (catalog no. 07-404; EMD Millipore;
dilution, 1:1,000), polyclonal rabbit anti-human PRMT4 (catalog no.
09-818; EMD Millipore; dilution, 1:1,000); monoclonal mouse
anti-human p16 (catalog no. SAB3300036; Sigma-Aldrich; dilution,
1:2,000); monoclonal mouse anti-green fluorescent protein (GFP;
catalog no. ab1218; Abcam, Cambridge, MA, USA; dilution, 1:3,000);
or monoclonal mouse anti-human β-actin (catalog no. A1978;
Sigma-Aldrich; dilution, 1:4,000). The membranes were then
visualized using the chemiluminescent substrate method and the
SuperSignal West Pico kit (Pierce Co, Rockford, IL, USA). β-actin
was used as an internal control for normalizing the loading
materials.
CoIP was performed using the A549 cells. The cells
were lysed in lysis buffer consisting of 50 mM Tris-HCl (pH 8.0),
150 mM NaCl, 0.5% NP-40, 1 mM EDTA and protease inhibitor cocktail.
Total cell extracts were incubated with gentle agitation with the
antibodies against GFP or p16 overnight at 4°C. This was followed
by the addition of 40 µl of Protein A agarose (Upstate
Biotechnology, Lake Placid, NY, USA) and incubation for another 3
h. The pellets were collected by centrifugation at 500 × g for 3
min at 4°C and washed twice with Buffer A consisting of 20 mM
Tris-HCl (pH 8.0), 10 mM NaCl, 0.5% NP-40 and 1 mM EDTA
(Sigma-Aldrich). The beads were suspended in 50 µl of 5X loading
buffer and boiled for 10 min. The proteins were separated on a 12%
SDS-PAGE gel and then transferred to a PVDF membrane for
immunoblotting with antibodies against p16, GFP, phosphoserine
(polyclonal rabbit anti-human; catalog no. AB1603; EMD Millipore;
dilution, 1:1,000) or dimethyl-arginine, asymmetric (ASYM24;
polyclonal rabbit anti-human; catalog no. 07-414; EMD Millipore;
dilution, 1:1,000).
Immunofluorescence and terminal
deoxynucleotidyl transferase-mediated dUTP nick end-labeling
(TUNEL) assay
Apoptosis of A549 cells was measured using TUNEL
stain (Nanjing KeyGen Biotech Co., Ltd., Nanjing, Jiangsu, China).
The treated A549 cells grown on coverslips were washed twice with
phosphate-buffered saline (PBS), fixed in 4% paraformaldehyde for
30 min and endogenous peroxidase activity was blocked with 3%
hydrogen peroxide diluted in methanol for 10 min at room
temperature. The cells were then incubated in 0.1% Triton X-100 for
2 min on ice. Subsequent to washing with PBS, the cells were
covered with 50 µl of TUNEL reaction mixture and incubated in the
solution for 60 min at 37°C in a humidified dark chamber. Finally,
the cells were stained with DAPI prior to visualization under an
Olympus FV1000 confocal microscope (Olympus Corporation, Tokyo,
Japan). Images were then captured using the FV10-ASW 2.1 microscopy
software (Olympus Corporation).
Flow cytometric analysis
The A549 cells were trypsinized and washed once with
cold PBS and then fixed in 70% ethanol and stored at 4°C for 30
min. The fixed cells were washed with PBS and suspended in 100 µl
of PBS, supplemented with 1 µl 10 mg/ml ribonuclease A and 100 µl
propidium iodide (Sigma-Aldrich). The stained cells were incubated
at room temperature for 30 min in the dark. The cellular DNA
content was quantified by flow cytometric analysis using a Epics XL
FACSCalibur flow cytometer (Beckman Coulter, Brea, CA, USA) and the
and the Multicycle AV for Windows software (Beckman Coulter).
Statistical analysis
Data are expressed as the mean ± standard deviation
based on ≥3 individual experiments. Treatment-related differences
were evaluated using one-way analysis of variance. P<0.05 was
considered to indicate a statistically significant difference.
Results
Mutant p16 protein exhibited
hypomethylation and enhanced function
The EGFP-p16 expression plasmids carrying three
arginine-lysine point mutations, consisting of R22/131/138K
(p16KKK), were generated. Firstly, the wild-type p16 and p16KKK
vectors were transfected into A549 cells, then analyzed using CoIP
assays performed using the cell extracts. The extracts from the
A549 cells were immunoprecipitated using antibodies against p16,
and detected in immunoblotting with antibodies against p16 and
asymmetric dimethylarginine (Fig.
1A). The findings of CoIP revealed that the wild-type p16
protein was methylated, while the methylation of p16KKK was
prominently reduced. Next, the cell cycle distribution was detected
using flow cytometric analysis (FACS). The results revealed that
overexpression of the wild-type p16 protein caused an accumulation
of cells in the G1 phase. However, cells overexpressing the p16KKK
protein exhibited an increased accumulation of cells in the G1
phase (Fig. 1B). Finally, the A549
cells were transfected with the wild-type p16 and p16KKK vectors,
and the expression was detected using a confocal laser scanning
microscope. The results revealed that the wild-type p16 and p16KKK
proteins were located in the nucleus (Fig. 1C). These data indicate that the
arginine point mutations in the p16KKK protein led to
hypomethylation of the p16 protein and reinforced the increased the
ability to prevent cell proliferation.
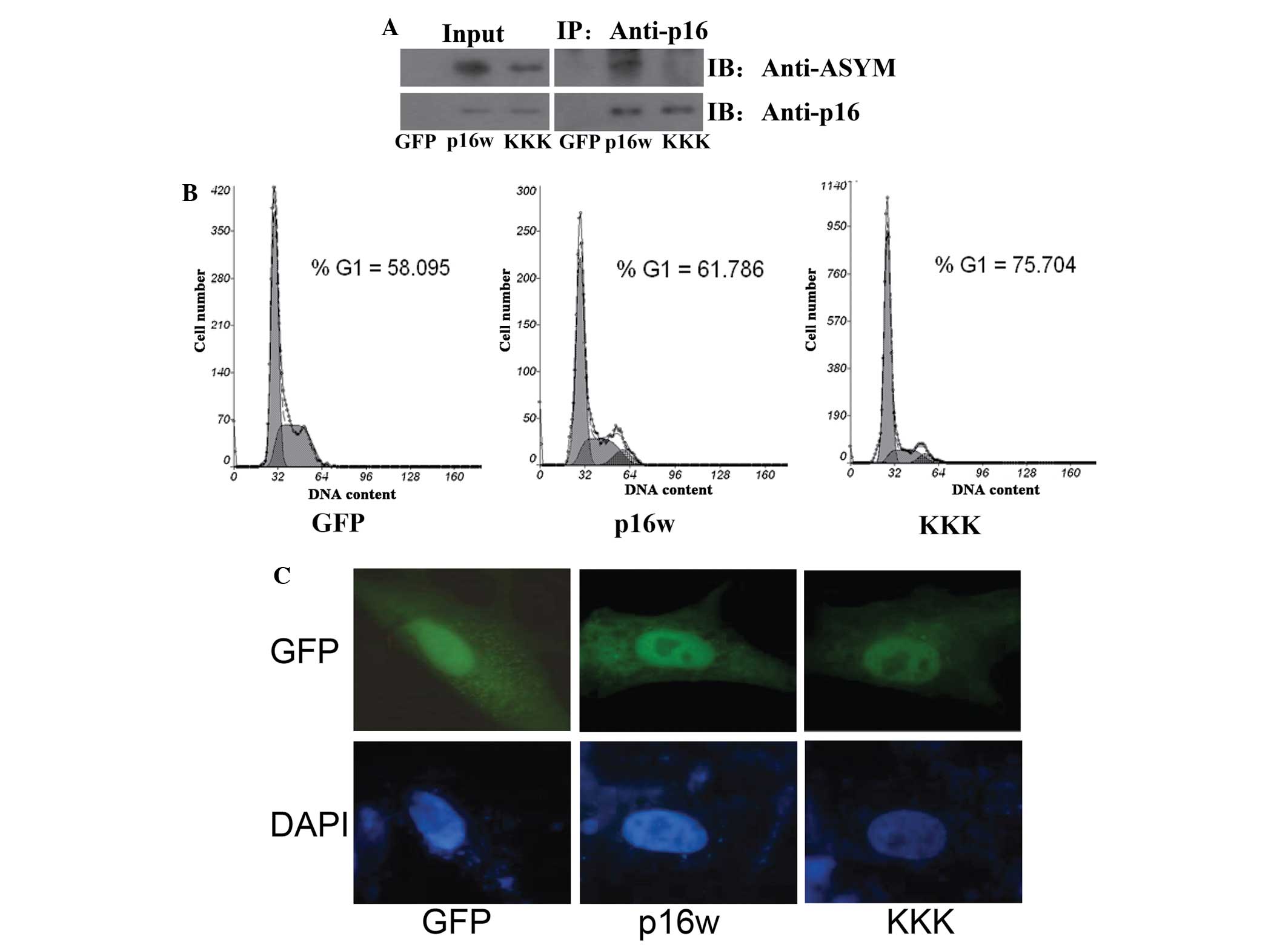 | Figure 1.The mutant p16KKK protein exhibited
hypomethylation and enhanced function. (A) The arginine 22, 131 and
138 residues in the p16 protein were methylation sites. A549 cells
transfected with the p16w and p16KKK expression vectors or control
EGFP-N1 empty vector. Co-immunoprecipitation was performed using
the antibody against p16, and the protein expression was detected
using antibodies against p16 or ASYM. (B) Flow cytometric analysis
of cell cycle changes following the transfection of A549 cells with
GFP, p16w or p16KKK plasmids. The A549 cells were harvested 48 h
subsequent to transfection. (C) The p16KKK and p16w proteins were
located in the nucleus of tbe A549 cells. The cells were
transfected with the p16-wild-EGFP or p16KKK-EGFP vectors. After 48
h, the nuclei were counterstained with DAPI (blue), and then
detected using confocal laser scanning microscopy. p16KKK, p16
protein carrying the R22/131/138K arginine-lysine point mutations;
p16w, wild-type p16; GFP, green fluorescence protein; ASYM,
asymmetric dimethylarginine; Input, proteins prior to
immunoprecipitation; IP: Anti-p16, samples co-immunoprecipitated
with antibodies. |
PRMT6 weakened the ability of p16 to
prevent cell proliferation
Identification of the protein that PRMT involves in
the methylation of the arginine residues of p16 was then attempted.
The present results revealed that p16 was modified with asymmetric
dimethylarginine (Fig. 1A),
suggesting that the type I enzymes are likely to play a role in p16
methylation. Firstly, the exogenous expression of PRMT1, PRMT4 and
PRMT6 was measured by western blotting (Fig. 2A). Next, CoIP assay results revealed
that the exogenous expression of PRMT1, PRMT4 and PRMT6 increased
the methylation level of p16, but only PRMT6 overexpression
inhibited the association of p16 and CDK4 (Fig. 2B). In addition, the results of the
FACS analysis revealed that transfection with PRMT6 counteracted
the arrest of A549 cells in the G1 phase induced by wild-type p16,
while PRMT6 overexpression did not affect the arrest of cell cycle
induced by p16KKK (Fig. 2C). However,
PRMT1 and PRMT4 demonstrated no such effect. Therefore, these data
revealed that PRMT6 inhibited the function of p16 through the
methylation of arginine residues.
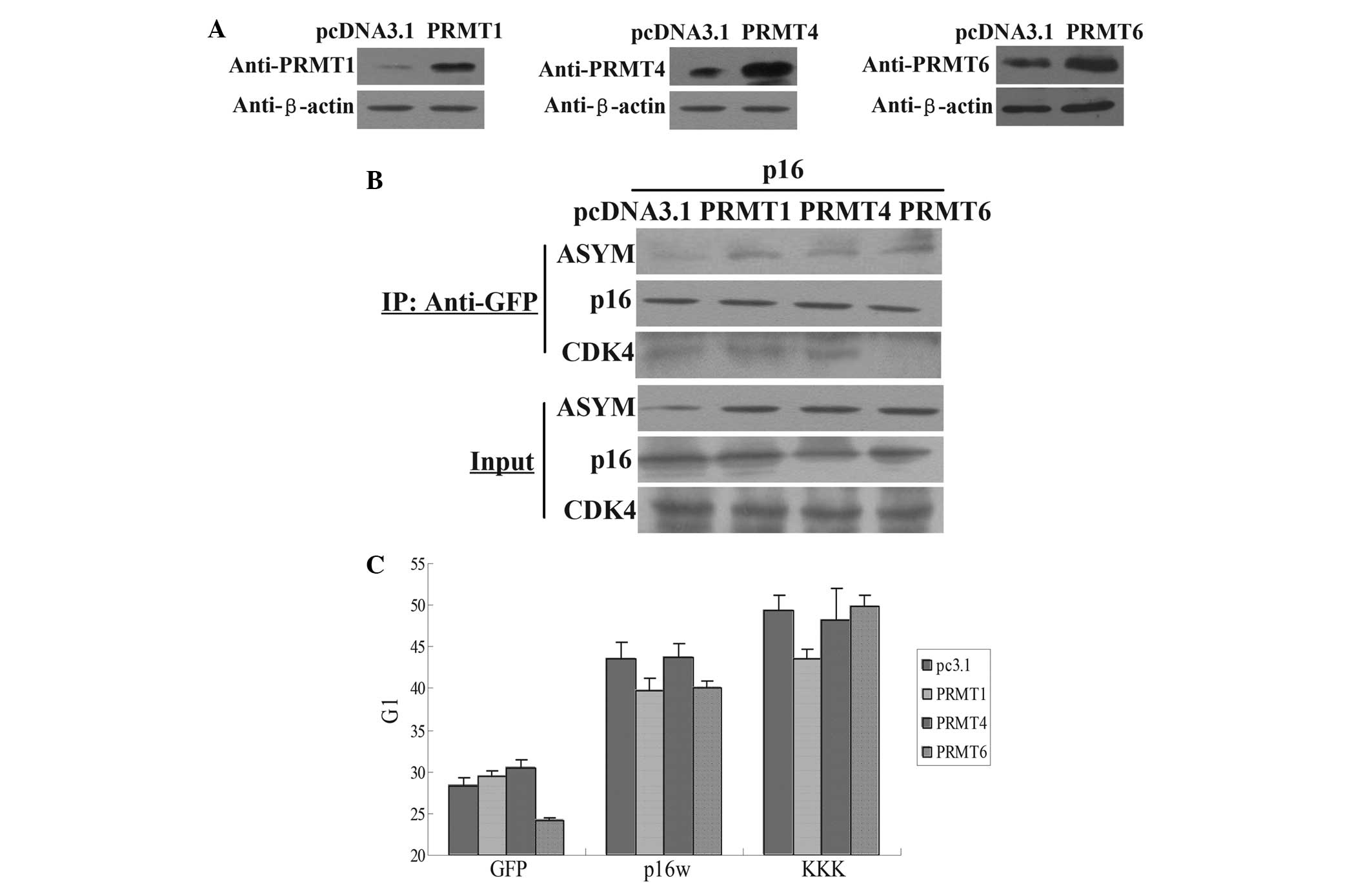 | Figure 2.PRMT6 influences the functional
properties of p16. (A) Western blot analysis of the expression of
the PRMT1, PRMT4 and PRMT6 proteins in A549 cells transfected with
the PRMT1, PRMT4 and PRMT6 expression vectors or the control
pcDNA3.1 empty vector. (B) Co-immunoprecipitation assays for
arginine methylation of p16. The cell extracts were prepared and
precipitated with antibody against GFP, and detected using
immunoblotting with antibodies against p16, CDK4 and ASYM antibody.
The cells were transfected with p16 and pcDNA3.1, PRMT1, PRMT4 or
PRMT6 vectors. (C) Flow cytometric analysis of cell cycle changes
following the transfection of A549 cells. The cells were harvested
48 h subsequent to transfection with wild-type p16 or p16KKK, with
PRMT1, PRMT4 or PRMT6 overexpressed. The bar chart means the G1
phase percentage. Values are means ± standard deviation (n=3).
Input, proteins prior to immunoprecipitation; IP: Anti-GFP, samples
co-immunoprecipitated with antibodies; PRMT, protein argenine
methyltransferase; ASYM, asymmetric dimethylarginine; p16KKK, p16
protein carrying the R22/131/138K arginine-lysine point mutations;
GFP, green fluorescence protein. |
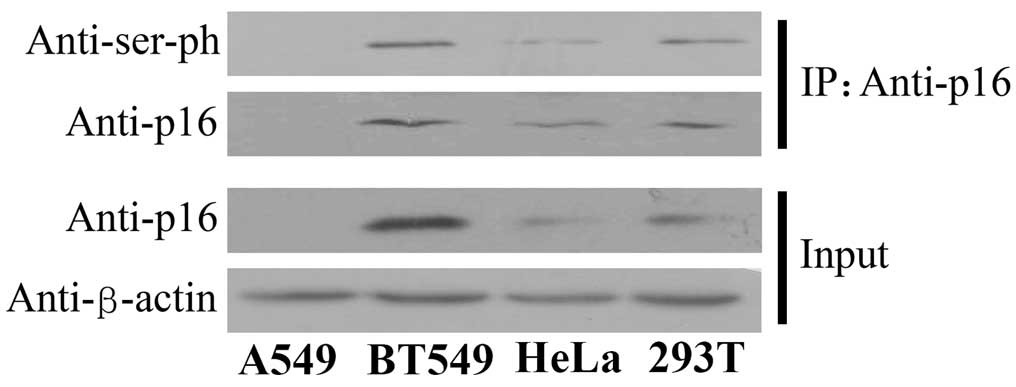
p16 was phosphorylated in a variety of
cells
The present study also investigated the effect of
other post-translational modifications on the function of the p16
protein. Therefore, the status of serine phosphorylation was
examined in four human cell lines derived from various cell types.
CoIP assays were performed using the antibody against p16 and the
protein expression was detected using the antibody against serine
phosphorylation. The findings revealed that the p16 protein was
phosphorylated in the human breast cancer BT549, cervical
epithelioid carcinoma HeLa and embryo kidney 293T cell lines
(Fig. 3). These data indicate that
the serine residues of p16 are universally phosphorylated in
various cell lineages.
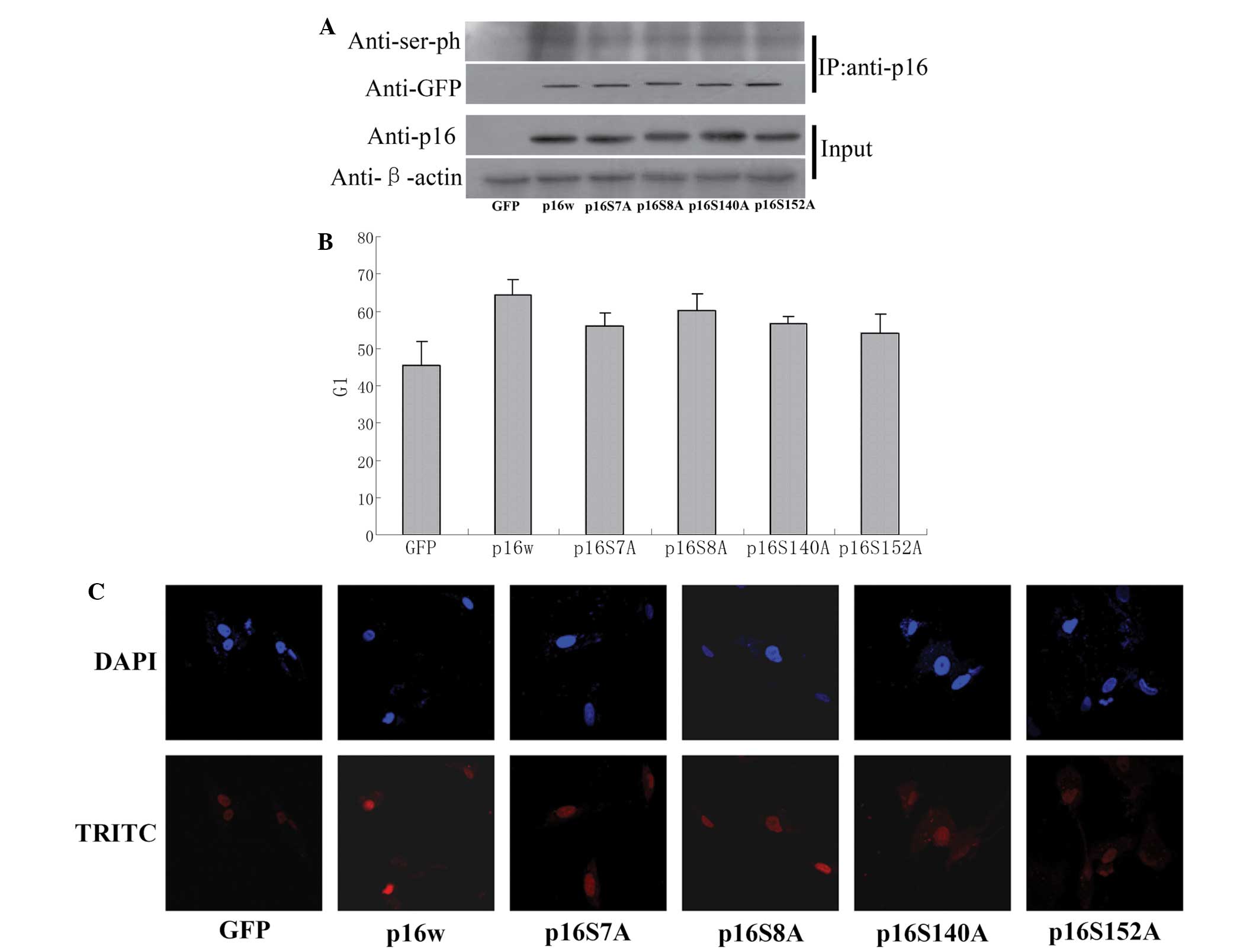
p16 protein with mutated serine
residues demonstrates a decreased ability to prevent A549 cell
proliferation
As a tumor suppressor, p16 prevents cell cycle
progression at the G1/S checkpoint. The present study aimed to
identify whether mutation of the serine residues S7, S8, S140 and
S152 in the p16 protein affect the ability of p16 to arrest the
cell cycle. CoIP assays demonstrated that the wild-type p16 protein
was phosphorylated, while the levels of phosphorylated p16 S7A, p16
S8A, p16 S140A and p16 S152A decreased by varying amounts (Fig. 4A). The FACS analysis performed in the
present study revealed that overexpression of the wild-type p16
protein resulted in an accumulation of cells in the G1 phase
compared with the control (65 vs. 45%). However, cells
overexpressing the mutant p16 protein exhibited a decreased ratio
of cells in the G1 phase (Fig. 4B).
Consequently, TUNEL staining revealed that the p16 mutant induced
decreased apoptosis compared with wild-type p16 in A549 cells
(Fig. 4C). These results indicated
that the phosphorylation of p16 is involved in the regulation of
the ability of p16 to reduce cell apoptosis.
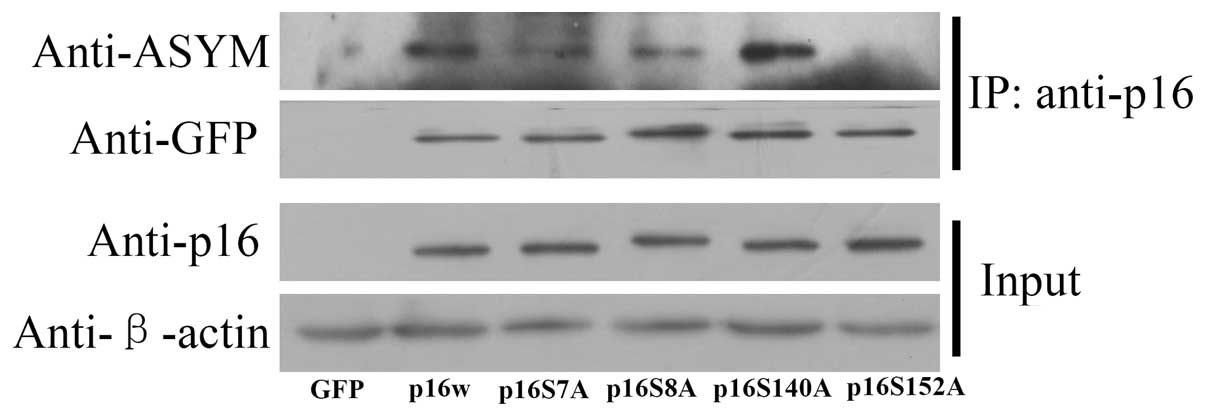
Crosstalk between arginine methylation
and serine phosphorylation in p16
To determine whether serine phosphorylation in p16
affects arginine methylation, CoIP assays were used. The mutated
serine residue and wild-type p16 vectors were transfected into A549
cells, then precipitated with anti-p16 antibody. The deposited
samples were detected with antibodies against p16 and asymmetric
dimethylarginine. The results revealed, as exhibited in Fig. 5, that the wild-type p16 was methylated
and the level of p16 S140A methylation was increased, while p16
S152A methylation was decreased. These results indicated that an
association existed between the methylation of arginine and
phosphorylation of serine in p16.
Discussion
It is well known that various post-translational
modifications modulate the function and activity of target proteins
by inducing changes in the structure and cellular localization of
the protein (15,16). In the present study, the p16 protein
was revealed to undergo arginine methylation and phosphorylation
(Figs. 1 and 3). It has previously been reported that
senescence in human prostatic epithelial cells not only induces an
elevated level of p16 protein, but also promotes the
phosphorylation of p16, which contributes to the binding affinity
between CDK4/6 and p16 and cell cycle arrest in the G1 phase
(17). Additional studies have
demonstrated that the four specific serine sites of p16, consisting
of Ser7, Ser8, Ser140 and Ser152, are phosphorylated, but only
Ser152 is phosphorylated in the CDK4/6-p16 compound (7). The present results suggested that the
phosphorylation of p16 played a role in regulating the ability of
p16 to reduce cell apoptosis, based on the evidence that mutation
of the residues from serine to alanine resulted in a decrease of
the phosphorylation level of p16 and a decreased the apoptosis
ratio compared with wild-type p16 in A549 cells (Fig. 4B and C). These findings support the
hypothesis that phosphorylation of p16 is an important mechanism
for p16 regulation. A previous study described that phosphorylation
of p16 in senescent prostatic epithelial cells may facilitate the
association between p16 and CDK4 and 6 (7). The phosphorylation of p16 evidently
plays a role in the function of the protein, and the findings from
the present study are consistent with previous data (7).
It is also recognized that one post-translational
modification may enhance or prevent another post-translational
modification, resulting in interplay that regulates diverse
molecular processes (18,19). In the present study, hypomethylation
of the p16 protein was found to enhance the ability of the protein
to prevent cell proliferation, while hypophosphorylation of p16
decreased the ability of the protein to reduce cell apoptosis
(Figs. 1 and 4). In addition, the present results
indicated that wild-type p16 was methylated, and the amount of
methylated p16 S140A was increased, while the amount of methylated
p16 S152A was decreased (Fig. 5). The
present results indicated that an association existed between the
methylation of arginine and phosphorylation of serine in p16. A
previous study reported that PRMT1 methylated forkhead box protein
O1 (FOXO1) at Arg248 and Arg250, which blocked Akt-mediated
phosphorylation of FOXO1 at Ser253 (18). In addition, another study revealed
that epidermal growth factor receptor (EGFR) Arg1175 is methylated
by PRMT5, and this modification positively modulates EGF-induced
EGFR trans-autophosphorylation at Tyr1173 (19). These results are similar to the
current research, promoting the further study of the function of
p16.
Overall, the present study hypothesizes that
crosstalk exists between phosphorylation and arginine methylation
of p16. The effects of the corresponding methylation of arginine
and phosphorylation of serine on the function of p16 require
additional investigation.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (grant no., 31271442).
References
|
1
|
Sherr CJ and Roberts JM: CDK inhibitors:
Positive and negative regulators of G1-phase progression. Genes
Dev. 13:1501–1512. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ohtani N, Zebedee Z, Huot TJ, Stinson JA,
Sugimoto M, Ohashi Y, Sharrocks AD, Peters G and Hara E: Opposing
effects of Ets and Id proteins on p16INK4a expression during
cellular senescence. Nature. 409:1067–1070. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gizard F, Amant C, Barbier O, Bellosta S,
Robillard R, Percevault F, Sevestre H, Krimpenfort P, Corsini A,
Rochette J, et al: PPAR alpha inhibits vascular smooth muscle cell
proliferation underlying intimal hyperplasia by inducing the tumor
suppressor p16INK4a. J Clin Invest. 115:3228–3238. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang X, Feng Y, Xu L, Chen Y, Zhang Y, Su
D, Ren G, Lu J and Huang B: YY1 restrained cell senescence through
repressing the transcription of p16. Biochim Biophys Acta.
1783:1876–1883. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang X, Pan L, Feng Y, Wang Y, Han Q, Han
L, Han S, Guo J, Huang B and Lu J: P300 plays a role in p16 (INK4a)
expression and cell cycle arrest. Oncogene. 27:1894–1904. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Feng Y, Wang X, Xu L, Pan H, Zhu S, Liang
Q, Huang B and Lu J: The transcription factor ZBP-89 suppresses p16
expression through a histone modification mechanism to affect cell
senescence. FEBS J. 276:4197–4206. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gump J, Stokoe D and McCormick F:
Phosphorylation of p16INK4A correlates with Cdk4 association. J
Biol Chem. 278:6619–6622. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang X, Huang Y, Zhao J, Zhang Y, Lu J and
Huang B: Suppression of PRMT6-mediated arginine methylation of p16
protein potentiates its ability to arrest A549 cell proliferation.
Int J Biochem Cell Biol. 44:2333–2341. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pahlich S, Zakaryan RP and Gehring H:
Protein arginine methylation: Cellular functions and methods of
analysis. Biochim Biophys Acta. 1764:1890–1903. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Di Lorenzo A and Bedford MT: Histone
arginine methylation. FEBS Lett. 585:2024–2031. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Han L, Lu J, Pan L, Wang X, Shao Y, Han S
and Huang B: Histone acetyltransferase p300 regulates the
transcription of human erythroid-specific 5-aminolevulinate
synthase gene. Biochem Biophys Res Commun. 348:799–806. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xie B, Invernizzi CF, Richard S and
Wainberg MA: Arginine methylation of the human immunodeficiency
virus type 1 Tat protein by PRMT6 negatively affects Tat
Interactions with both cyclin T1 and the Tat transactivation
region. J Virol. 81:4226–4234. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Frankel A, Yadav N, Lee J, Branscombe TL,
Clarke S and Bedford MT: The novel human protein arginine
N-methyltransferase PRMT6 is a nuclear enzyme displaying unique
substrate specificity. J Biol Chem. 277:3537–3543. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Boulanger MC, Liang C, Russell RS, Lin R,
Bedford MT, Wainberg MA and Richard S: Methylation of Tat by PRMT6
regulates human immunodeficiency virus type 1 gene expression. J
Virol. 79:124–131. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Strahl BD and Allis CD: The language of
covalent histone modifications. Nature. 403:41–45. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Berger SL: The complex language of
chromatin regulation during transcription. Nature. 447:407–412.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sandhu C, Peehl DM and Slingerland J:
p16INK4A mediates cyclin dependent kinase 4 and 6 inhibition in
senescent prostatic epithelial cells. Cancer Res. 60:2616–2622.
2000.PubMed/NCBI
|
|
18
|
Yamagata K, Daitoku H, Takahashi Y, Namiki
K, Hisatake K, Kako K, Mukai H, Kasuya Y and Fukamizu A: Arginine
methylation of FOXO transcription factors inhibits their
phosphorylation by Akt. Mol Cell. 32:221–231. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hsu JM, Chen CT, Chou CK, Kuo HP, Li LY,
Lin CY, Lee HJ, Wang YN, Liu M, Liao HW, et al: Crosstalk between
Arg 1175 methylation and Tyr 1173 phosphorylation negatively
modulates EGFR-mediated ERK activation. Nat Cell Biol. 13:174–181.
2011. View
Article : Google Scholar : PubMed/NCBI
|