Introduction
A single messenger RNA (mRNA) may be regulated by a
number of micro RNAs (miRNAs), and a single miRNA is able to
regulate multiple mRNAs (1). This
finding suggests that the crosstalk between messenger RNAs and
miRNAs is intricate and complex. The canonical role of mRNAs is the
delivery of protein-coding information to sites of protein
synthesis (2). However, during the
past decade, miRNAs, a family of small non-coding RNAs that are
important post-transcriptional regulators of gene expression
through binding to RNAs, have been well characterized. Numerous
researchers have attempted to decipher the crosstalk at this
post-transcriptional regulatory stage. The competing endogenous RNA
(ceRNA) hypothesis states that all types of RNA transcript compete
for miRNAs through a ‘language’ mediated by miRNA response elements
(MREs), and is compelling in this context (3). On the basis of this hypothesis, coding
and non-coding RNA transcripts with shared MREs are able to
actively communicate with each other to regulate their respective
expression levels, thus explaining the consistent expression
patterns of ceRNAs and miRNA targets (4).
miRNA inhibitors, termed ‘microRNA sponges’, are
transcripts expressed from certain promoters, which contain
multiple, tandem binding sites to for a particular miRNA (5). These artificial sponges may be
transiently transfected into cultured cells to derepress miRNA
targets, at least as effectively as chemically altered antisense
oligonucleotides (6). IPS1, the first
endogenous sponge RNA discovered in plants, contains a motif with
sequence complementary to miRNA (miR)-399, and functions as an
miRNA decoy for sequestering miR-399. This mechanism of inhibition
of miRNA activity is termed ‘target mimicry’, thus resulting in
derepression of the miR-399 target PHO2 (7). Artificial target mimics, antisense
oligonucleotides and natural miRNA sponges have all been confirmed
to compete with miRNA targets for miRNAs, and attenuate the
inhibitory activity of miRNAs, indicating the potential ability of
non-coding transcripts to interact with miRNAs and control the
expression of miRNA target genes at the post-transcriptional
regulatory circuit level.
The emerging roles of RNA-RNA crosstalk, as part of
a complex posttranscriptional regulatory circuit, have been
implicated in human development and disease (Table I). Functional analysis of the miRNA
competition and inhibition will likely result in significant
insights, regarding basic physiology and disease progression. The
present review focused on the molecular mechanisms of ceRNAs and
the classification of natural RNA species, as well as the
implication of ceRNAs in the progression of cancer.
 | Table I.Validated non-coding competing
endogenous RNAs. |
Table I.
Validated non-coding competing
endogenous RNAs.
| ceRNA subtypes | Function | Tissues and
species | Reference |
|---|
| Protein-coding
RNAs |
|
|
|
|
ZEB2 and
PTEN | Upregulation of
PTEN by sequestering miRNAs | Melanoma | (11) |
|
DKK1 and
PTEN | Modulation of
PTEN protein levels | Diabetic
cardiomyocytes | (12) |
|
VCAN and PTEN,
Rb1 | Free Rb1 and
PTEN mRNAs for translation, and thus inhibit growth | Breast cancer | (13) |
|
VCAN and CD34,
FN1 | VCAN interacts with
CD34 and FN1 as an miRNA decoy | Hepatocellular
carcinoma | (14,15) |
|
CD44 and
CDC42 | Inhibition of cell
proliferation, colony formation, tumor growth | Breast cancer and
various other types of cancer | (37) |
|
CD44 and
Col1α1, FN1 | Enhancement of
metastasis in vivo | Breast
carcinoma | (57) |
|
PTEN and CNOT6L,
VAPA | Control downstream
PI3K signaling and cell growth | Glioblastoma and
prostate cancer | (51) |
|
HMGA2 and
Tgfβr3 | Competition for
let-7 occupancy with the TGF-β co-receptor
Tgfβr3 | Lung cancer | (40) |
| Long non-coding
RNAs |
|
|
|
|
LINCMD1 and MAML1,
MEF2C | Control of muscle
differentiation through upregulation of MAML1 and
MEF2C transcription factors | Mouse and human
myoblasts | (21) |
| HULC
and PRKACB | Downregulation of
miRNA-mediated repression | Liver cancer | (22) |
|
HOTAIR and
HER2 | Regulation of HER2
expression | Gastric cancer | (47) |
|
LincRNA-RoR, OCT4, | Mediation of
embryonic stem cell | Endometrial cancer
stem cells, | (33,57) |
|
SOX2 and
NANOG | self-renewal and
differentiation | embryonic stem
cells |
|
|
IPS1 and
PHO2 | Downregulation of
miRNA-mediated repression | Arabidopsis
thaliana | (7) |
|
H19 and DICER,
HMGA2 | Modulation of let-7
availability as a molecular sponge | Mus musculus
and Homo sapiens | (23) |
| Pseudogenes |
|
|
|
|
PTENP1 and
PTEN | Upregulation of
PTEN as a ceRNA | Prostate tumor,
colon cancer | (4) |
|
KRAS1P and
KRAS | Increase
KRAS mRNA abundance | Prostate cancer,
neuroblastoma, retinoblastoma and hepatocellular carcimoma | (4) |
| Pbcas4
and BCAS4 | Conserved
competitive endogenous RNA of BCAS4 | Mus musculus
and Homo sapiens | (27) |
| Circular RNAs |
|
|
|
|
CDR1as or ciRS-7
and | Downregulation
of | Human and mouse
brain | (29,30) |
|
SNCA, EGFR,
IRS2 | miRNA-mediated
repression |
|
|
Molecular mechanism of ceRNAs and
classification
mRNAs, transcribed pseudogenes, long non-coding RNAs
(lncRNAs) and circular RNAs (circRNAs) that share common MREs with
mRNAs may be similarly targeted, sequestering miRNAs to inhibit
their interaction with protein-coding mRNAs (Fig. 1A). Therefore, these ceRNAs competing
for common miRNAs are also able to regulate each other in ceRNA
networks. The strength of this cross-talk is determined by various
conditions, including the relative levels of miRNAs and targets,
the number of shared miRNA binding sites and the strength of miRNA
binding to the target or ceRNA (8).
When a given mRNA is upregulated, the repression conferred by its
associated targeting miRNAs is decreased, as the total number of
MREs exceeds that of the miRNAs themselves (9). Similarly, forced expression of ceRNAs
sharing common MREs with protein-coding RNAs sequesters microRNAs
and alters the expression levels of miRNA targets (Fig. 1B and C).
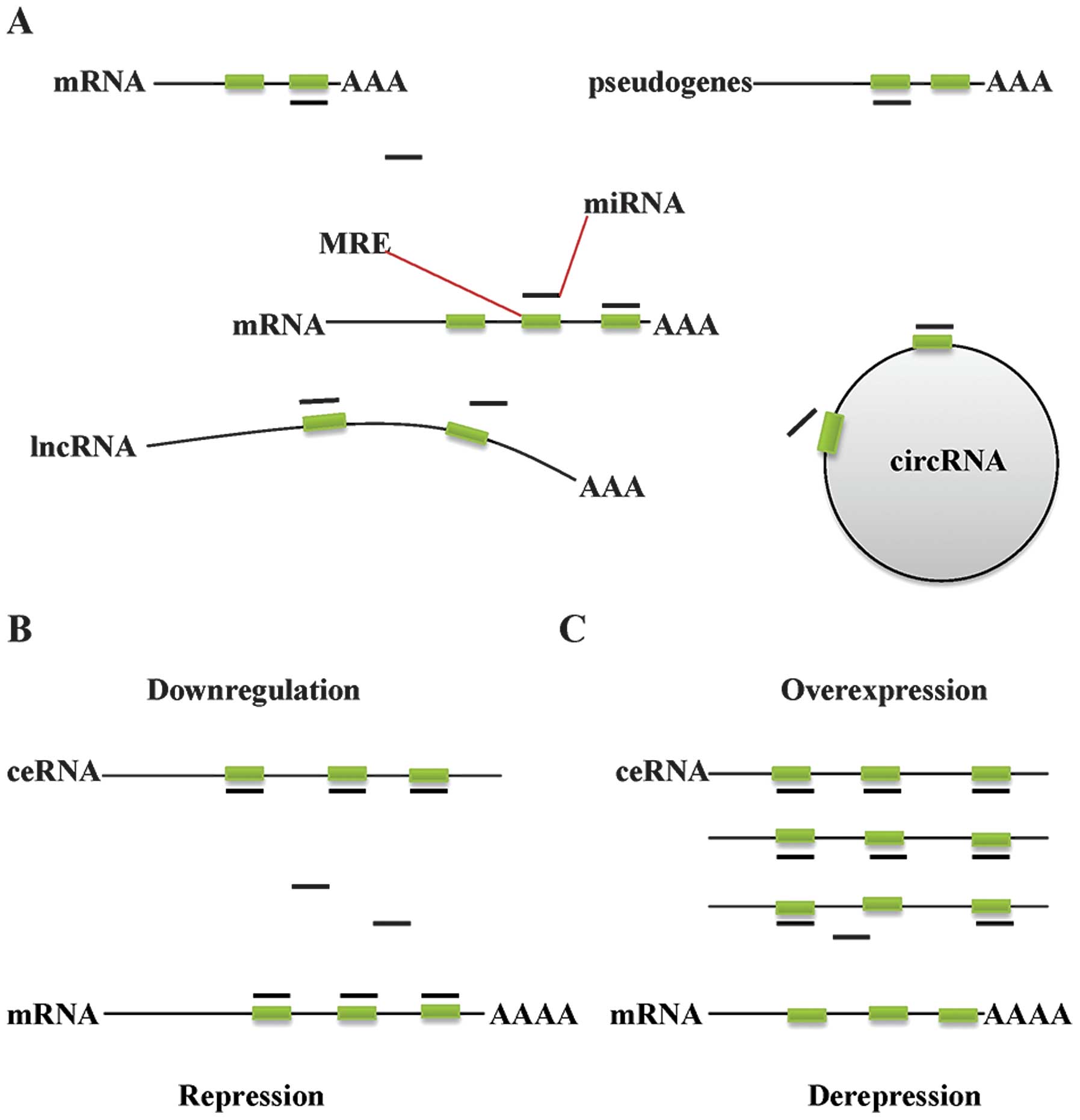 | Figure 1.Molecular mechanism of ceRNAs and
classification. (A) mRNAs, transcribed pseudogenes, lncRNAs and
cirRNAs are able to function as competing endogenous RNAs. (B)
Downregulation of ceRNAs results in increased availability of miRNA
molecules to bind to mRNA, thereby repressing protein translation.
(C) Overexpression of ceRNA results in a reduction in free miRNA
abundance, facilitating the derepression of mRNAs that contain
identical MREs, thus increasing protein expression levels. ceRNA,
competing endogenous RNA; mRNA, messenger RNA; lncRNA, long
non-coding RNA; cirRNA, circular RNA; miRNA, micro RNA; MRE, miRNA
response element. |
mRNA as ceRNAs
It has previously been demonstrated that miRNAs are
able to inhibit protein-coding RNAs through binding to MREs of mRNA
3′ untranslated regions (3′UTR) (10). The majority of validated ceRNAs are
mRNAs, and their ability to compete for miRNA binding and sequester
miRNAs from alternate targets may induce biological functions of
mRNAs independent of those of their encoded proteins (4). One putative phosphatase and tensin
homolog (PTEN) ceRNA, the zinc finger E-box binding homeobox 2
(ZEB2) mRNA, was demonstrated to exert protein-independent and
miRNA-dependent regulation of PTEN expression via sequestration of
shared miRNAs (11). ZEB2 mRNA has
been validated as a PTEN ceRNA and regulates PTEN levels by
sequestering at least four miRNAs (miR-181, miR-200b, miR-25 and
miR-92a). Attenuation of ZEB2 expression results in the repression
of PTEN in human melanomas (11).
Another validated PTEN ceRNA is Dickkopf WNT signaling pathway
inhibitor 1 mRNA which competes with PTEN mRNA for miR-93 and
miR-106a (12). In addition, the
3′UTRs of protein-coding transcripts that typically contain MREs
for multiple miRNAs are critical for mRNAs to function as ceRNAs
for miRNA targets. The versican (VCAN) 3′UTR has been reported to
modulate PTEN levels by sequestering shared miRNAs miR-144 and
miR-136 (13). miR-199a-3p and
miR-144 target cell cycle regulator retinoblastoma 1 (Rb1), which
was also demonstrated to function as a ceRNA for VCAN (13). Thus, the VCAN 3′UTR binds and
modulates miRNA activities, acting as a natural miRNA sponge and
releasing the Rb1 and PTEN mRNAs for translation. CD34 and FN1 have
also been validated as two additional VCAN ceRNAs, which compete
for binding to miR-133a, miR-199a-3p, miR-144 and miR-431 (14,15). In
light of the ceRNA hypothesis, the 3′UTRs of protein-coding
transcripts also likely decoy miRNAs from transcripts with shared
MREs in a protein-independent manner, thereby acting as trans
regulatory elements to regulating such transcripts (3). Overall, these findings suggest that
protein-coding transcripts and 3′UTRs of coding genes may possess
significant biological activity through their ability to function
as endogenous decoys for miRNAs and thereby regulate miRNA
targets.
lncRNAs as ceRNAs
With the ever-expanding number of lncRNAs,
increasing numbers of studies have focused on the roles of lncRNAs
in epigenetic mechanisms and other biological processes (16,17).
Notably, lncRNAs have begun to emerge as natural miRNA decoys,
suggesting that they may function as ceRNAs at the
post-transcriptional regulatory level (18–20). In
particular, the muscle-specific lncRNA LINCMD1 regulates muscle
differentiation by binding and sequestering miR-133 and miR-135
(21). Typically, these miRNAs
negatively regulate expression of the mastermind-like 1 (MAML1) and
myocyte enhancer factor 2C (MEF2C) transcription factors, which
induce muscle-specific gene expression. Therefore, by sequestering
these miRNAs, LINCMD1 functions as an miRNA decoy and activates
MAML1 and MEF2C. In addition, HULC, an lncRNA that has previously
been identified to be highly upregulated in liver cancer, acts as
an endogenous sponge, downregulating the activity of a series of
miRNAs, including miR-372 (22). The
inhibition of miR-372 reduces translational repression of its
target gene, PRKACB. Another lncRNA, H19, which possesses canonical
and non-canonical binding sites for the let-7 family of miRNAs, has
been demonstrated to modulate let-7 availability by competing with
DICER and HMGA2 as a molecular sponge (23). These studies indicated that lncRNAs
may be involved in post-transcriptional regulation by functioning
as ceRNAs. Additional lncRNAs functioning as miRNA decoys will be
discussed in detail below.
Pseudogenes as ceRNAs
Pseudogenes are defined as genomic loci that
resemble functional genes, but are considered biologically inactive
as they posses premature stop codons, deletions, insertions and/or
frameshift mutations that prevent their effective translation into
functional proteins. There are almost as many pseudogenes as there
are coding genes, and these pseudogenes represent a significant
proportion of the ‘transcriptome’ (24). Sequencing has revealed that nucleotide
sequences contained within pseudogenes are well preserved,
suggesting that selective pressure to maintain these genetic
elements may exist (25). Processed
pseudogenes are generated through retrotransposition and therefore
contain no introns; however, they commonly share 5′ and 3′UTR
sequences with their ancestral genes (26). PTENP1 is supressed by several
validated PTEN-targeting miRNAs, and overexpression and RNA
interference experiments confirmed that PTENP1
posttranscriptionally regulates the expression of PTEN via shared
miRNAs (4). The breast carcinoma
amplified sequence 4 (BCAS4) pseudogene, Pbcas4, whose transcripts
compete with BCAS4 mRNAs for binding to the common miR-185, is a
conserved ceRNA in the mouse and human genome (27). In addition, several other pseudogenes,
including the OCT4-pg1, E2F3P1 and CDK4PS, were demonstrated to
share binding sites for common miRNAs with their parental genes
(4), suggesting that gene regulation
by pseudogenes acting as ceRNAs may be a frequent phenomenon.
However, to date, few pseudogenes have been functionally
characterized, and more evidence supporting this potentially common
phenomenon remain to be experimentally validated.
circRNA as ceRNAs
circRNAs were initially described in the 1990s, and
the most well-studied circRNA is that generated from the
sex-determining Y (SRY) gene, although the biological functions of
these RNA circles has remained elusive (28). Recently, SRY, the testis-specific
circRNA, was validated as an miR-138 sponge, with ceRNA activity
(29), suggesting that these RNA
circles may have significant roles in regulatory RNA networks. An
antisense transcript to CDR1, termed CDR1as or circRNA sponge for
miR-7, is highly expressed in the human and mouse brain, where
circRNA acts as an miR-7 sponge (29,30).
CDR1as contains >70 selectively conserved miRNA target sites and
suppresses miR-7 activity, resulting in the enhanced expression of
miR-7 targets (30). circRNAs have
been identified in multiple types of tissue, and were demonstrated
to be resistant to RNase R treatment (which degrades linear RNA
species), and possess longer half-lives compared with that of their
linear RNA transcript counterparts (31,32). Only
the linear forms of these RNAs were present in heavy polyribosome
fractions, suggesting that the circular forms remain untranslated
(31). As a result of their high
expression levels and stability, circRNAs with ceRNA activity may
be particularly effective modulators of the cross-talk between
linear ceRNAs.
ceRNA crosstalk in the progression of
cancer
Cross-talk between ceRNAs through shared miRNAs
represents a novel layer of gene regulation that may have
significant roles in development and disease (12,21,33). In
living organisms, although diverse cells that are structured into
tissues and organs grow, progress and exert their functions
constantly, it is difficult to appreciate the ‘perfect mechanism’
(34). However, tumorous cells
transformed from normal physiological cells that grow beyond their
natural boundaries, provide an approach for elucidation of the
underlying mechanism. Over several decades of cancer research, six
hallmarks of cancer that form the fundamental principle of the
process of malignant transformation were proposed and modified
(35). These six hallmarks of cancer
were: i) sustained proliferative signaling; ii) evasion of growth
suppressors; iii) facilitation of replicative immortality; iv)
activation of invasion and metastasis; v) induction of
angiogenesis; and vi) resistance to cell death. Herein, the present
review proposes to introduce ceRNA post-transcriptional regulation
into cancer biology and focus on the ceRNA networks involved in
these six hallmarks of cancer.
Sustaining proliferative
signaling
The capability of constant proliferation is one of
the most prominent characteristics of cancer cells. Normal cells
tightly regulate the balance of cellular proliferation and death to
maintain strict control of cell number, tissue architecture and
function. Once this balance is broken, tumor cell signaling
cascades that determine their dependence on proliferation signals
are deregulated, which results in unlimited growth (36). Ectopic expression of the CD44 3′UTR
binds and inactivates multiple miRNAs, including miR-216a, miR-330
and miR-608, freeing the target mRNA CDC42 from repression
(37). CDC42, a gene involved in cell
cycle progression, inhibits proliferation, colony formation and
tumor growth following enhanced translation. Upregulation of CD44
expression in epithelial ovarian cancer is associated with a
favorable outcome and is indicative of enhanced survival time
(38). By contrast, the loss of CD44
expression observed in Burkitt's lymphoma, neuroblastoma and
prostate cancer was accompanied by oncogenic transformation
(39). Thus, it was hypothesized that
CD44 mRNA competes with CDC42 for miRNA binding and results in
tumorigenesis following the loss of CD44 expression.
In addition to CD44, HMGA2 has been demonstrated to
promote lung carcinogenesis via two mechanisms: As a protein-coding
gene and as a non-coding RNA (40).
HMGA2 is highly expressed in metastatic lung adenocarcinoma, where
it has previously been demonstrated to contribute to cancer
progression and metastasis (41–43). HMGA2
promotes lung tumor formation by competing with the TGF-β
co-receptor Tgfβr3 for let-7 occupancy, which activates the TGF-β
signaling involved in lung cancer progression. These studies
thereby identified a novel gene-expression pathway, where the
protein-coding gene, HMGA2, largely behaves independently of its
protein-coding function, inducing lung cancer progression as a
ceRNA.
The lncRNA, HOTAIR, was initially known for its
effects in primary breast tumors and breast cancer metastases,
where enhanced HOTAIR expression promoted invasion and metastasis
(44). Recent studies also identified
upregulated HOTAIR expression in gastric cancer (45,46).
HOTAIR functions as a ceRNA, regulating the expression of human
epithelial growth factor receptor 2 (HER2) by competing for
miR-331-3p. HOTAIR thus functions as an oncogene in gastric
pathogenesis by inducing the activation of HER2 cell signaling
networks (47). This finding further
supports the hypothesis that the ceRNA function of RNA transcripts
is of fundamental significance in oncogenic transformation.
A study revealed that overexpression of the KRAS1P
pseudogene 3′UTR resulted in enhanced KRAS mRNA expression and
accelerated cell growth (4). Notably,
KRAS and KRAS1P transcript levels are co-regulated in prostate
cancer, and the KRAS1P locus at 6p11–12 is amplified in various
types of human tumor, including retinoblastoma, neuroblastoma and
hepatocellular carcinoma (48–50). These
findings are indicative of a putative, coding-independent,
proto-oncogenic role for KRAS1P, which may be explained by an miRNA
decoy mechanism. However, the specific miRNAs involved remain to be
elucidated and require further investigation.
Protein-coding RNAs, lncRNAs and pseudogene
transcripts are able to function as ceRNAs and contribute to the
induction of uncontrolled proliferation. Furthermore, invaluable
insight into the function of diverse species may be acquired
following analysis of the ceRNA crosstalk involved in cancer
progression.
Evading growth suppressors
Several tumor suppressive genes that inhibit
cellular growth and proliferation through diverse signaling
pathways have been identified, including Tp53, phosphate and tensin
homolog (PTEN) and Rb1. The complete loss of a tumor suppressor
gene in tumor cells represents one mechanism of achieving constant
proliferative capability, for example Tp53. However, another
mechanism is also used to posttranscriptionally modulate and
inactivate the tumor suppressor genes.
The pseudogene PTENP1 competes with PTEN for miRNA
binding, thereby modulating the derepression of specific miRNA
targets (4). Following the loss of
PTENP1, the decreased translation of PTEN was unable to exert tumor
suppressive functions by inhibiting the phosphoinositide 3-kinase
(PI3K)/AKT signaling pathway. The existence of genomic copy number
losses at the PTENP1 locus supports the hypothesis that PTENP1 is a
tumor suppressor gene, and may be under selective pressure to
undergo copy number losses in cancer. Another two protein-coding
RNAs, CNOT6L and (vesicle-associated membrane protein)-associated
protein A (VAPA), appeared to phenocopy PTEN loss-mediated AKT
activation based on their function as PTEN ceRNAs (51). Thus, silencing of the tumor suppressor
gene PTEN due to loss of CNOT6L and VAPA, resulted in evasion of
growth inhibition. The identification of significant copy number
losses of the VAPA and CNOT6L genomic loci supported the hypothesis
that these protein-coding RNAs exert tumor-suppressive effects.
The ZEB2 protein has previously been established as
an activator of the epithelial-to-mesenchymal transition (EMT) and
was demonstrated to be involved in the promotion of cancer
progression and metastasis in certain instances of epithelial
cancer (52,53). However, the ZEB2 transcript functions
as a PTEN ceRNA and modulates PTEN protein levels in an
miRNA-dependent, protein coding-independent manner (11). ZEB2 and PTEN are co-regulated, and
ZEB2 levels are commonly attenuated in human cancers. PTEN
antagonizes PI3K/AKT signaling, therefore attenuation of ZEB2
expression activates the PI3K/AKT pathway, enhancing cell
transformation.
Expression of the VCAN 3′UTR has been reported to
bind and modulate miRNA activities, releasing PTEN mRNAs for
translation, and resulting in reduced cell proliferation and tumor
growth (13). In addition to its role
as a PTEN ceRNA, VCAN was also demonstrated to act as a ceRNA for
the cell-cycle regulator Rb1 by competing for common miRNAs,
upregulating this crucial tumor suppressor in vitro and
in vivo (13). Expression of
Rb1 and PTEN were synergistically upregulated in vitro and
in vivo, suggesting that VCAN transcripts acting as ceRNAs
contribute to tumorigenesis via the Rb1 and PI3K signaling
pathways.
In addition, an lncRNA tumor suppressor, PTCSC3, was
found to be downregulated in thyroid cancer cells (54). PTCSC3 functions as a ceRNA by
sequestering miR-574-5p, thereby inducing cell cycle arrest.
However, further investigations regarding the miRNA targets
co-regulated with PTCSC3 in thyroid cancer is required.
Collectively, protein-coding RNAs and noncoding
RNAs, including pseudogene transcripts and lncRNAs, attenuate
proliferation inhibition and promote tumorigenesis through the
sequestration of miRNAs as ceRNAs.
Enabling replicative immortality
In contrast to normal cells that are only able to
complete a limited number of cell divisions, tumor cells
extensively self-renew, similarly to embryonic stem cells (ESCs).
Thus, it was hypothesized that tumor cells acquire this ESC-like
property by reprogramming normal cells and enabling replicative
immortality. A large intergenic noncoding RNA, linc-RoR, whose
expression is linked to pluripotency and self-renewal, has been
identified as a key reprogramming regulator (55). Linc-RoR functions as a key ceRNA,
linking the network of miRNAs and core transcription factors (TFs),
including Oct4, Sox2 and Nanog, and protecting these core TFs from
miRNA-mediated suppression in self-renewing human ESCs (56). Linc-RoR also functions as a ‘sponge’
against mediation of the differentiation of endometrial cancer stem
cells by miR-145, suggesting that endogenous linc-RoR may have a
key role in cancer stem cell (CSC) maintenance and pluripotency
(33). The identification of
additional ceRNAs associated with self-renewal of CSCs requires
further exploration.
Activating invasion and
metastasis
Invasion and distant metastases occur in diverse
types of advanced tumor. Notably, the majority of patients with
cancer succumb to these metastases rather than the primary tumor.
Frequently, cancer cells undergo morphological changes and alter
their cell-cell or cell-matrix interactions. Altered expression of
components of the extracellular matrix (ECM) has a significant role
in cancer metastasis. It was previously revealed that
overexpression of the CD44 3′UTR resulted in enhanced cell
motility, invasion and adhesion in the MDA-MB-231 human breast
carcinoma cell line, and also enhanced metastasis in vivo
(57). miRNAs which interact with the
CD44 3′UTR also have binding sites in other matrix-encoding mRNA
3′UTRs, including collagen type 1α1 (Col1α1), which is suppressed
by miR-328, as well as fibronectin type 1 (FN1), which is repressed
by miR-512-3p, miR-491 and miR-671 (57). The involvement of FN1 in cancer cell
migration and metastasis has also been well documented (58). Col1α1 is a major ECM component, which
affects cell behavior and maintains tissue architecture. Protein
analysis demonstrated that, following transfection of the CD44
3′UTR, the expression levels of CD44, Col1α1 and FN1 were
synergistically upregulated in vitro and in vivo
(57). Therefore, the non-coding
3′UTR of CD44 interacts with numerous miRNAs that target ECM
properties and activate invasion and metastasis. Furthermore,
overexpression of the VCAN 3′UTR would compete with FN1 for binding
to three miRNAs, miR-133a, miR-431 and miR-199a*, resulting in
ectopic invasion and metastasis in human hepatocellular carcinoma
(14,15).
In addition, Col1α2 was confirmed to be one of the
ceRNA targets of HMGA2, which also competes for let-7 binding
(44). Let-7 targets Col1α2 and
inhibits cell migration in hepatocellular carcinoma (59). These findings suggest that HMGA2 may
also function as a ceRNA and promote invasion and metastasis. These
invasive and migratory phenotypic consequences remain to be
validated following silencing of HMGA2 or overexpression of HMGA2
3′UTR.
Inducing angiogenesis
Angiogenesis, a process which is typically confined
to embryonic development, may be reactivated under certain
conditions in patients with tumors. The secreted protein, vascular
endothelial growth factor-A (VEGFA), whose expression may be
induced by hypoxia or oncogenic signals, is an activator of
angiogenesis (60). Analysis of gene
expression in glioblastoma, in combination with matched miRNA
profiles, revealed posttranscriptional regulation of notable
magnitude, comprising >248,000 miRNA-mediated interactions
(61). In particular, ectopic
expression of PTEN or Rb1 3′UTRs induced a 1.5-fold upregulation of
VEGFA, in an miRNA-dependent manner. These data indicated that
VEGFA functioned as natural miRNA sponge and was involved in the
posttranscriptional regulation of gene expression. Therefore, the
determination of whether overexpression of PTEN or Rb1 3′UTRs
promotes angiogenesis by competing for miRNA binding with VEGFA in
glioblastoma requires experimental validation.
Resisting cell death
Three major pathways, apoptosis, autophagy and
necrosis result in cell death, and highly malignant types of cancer
are able to attenuate cell death and become therapy resistant. In
cancer, the role of miRNA in drug resistance has been well studied
over the last few years (62,63). ceRNAs which compete for miRNA binding
with RNA transcripts involved in these three pathways remain to be
researched.
Conclusion and perspectives
The emerging ceRNA hypothesis is a novel field of
RNA biology. Studies by several groups have revealed that ceRNAs
function as posttranscriptional regulators of gene expression by
decoying miRNAs from other RNA transcripts (4,11,21). Protein-coding RNAs, pseudogenes,
lncRNAs and circRNAs serve as natural miRNA sponges through
sequestration of miRNAs. Previously, loss of CD44 expression
reduced CDC42 as miRNA decoys and promoted proliferation. In
addition, CD44 competed with FN1 and Col1α1 as a ceRNA leading to
metastasis in human tumors (37,57).
Expression of VCAN 3′UTRs reduced tumor growth by competing with
Rb1 and PTEN mRNAs, and contributed to invasion by serving as an
FN1 ceRNA (13–15). These findings suggest that
tumorigenesis, accompanied by metastasis and other hallmarks of
cancer, may be partially attributed to ceRNAs, which compete with
diverse RNA transcripts for various miRNAs. Furthermore, an
equilibrium exists between ceRNAs and miRNAs in the
post-transcriptional regulatory network, which when perturbed,
contributes to disease pathogenesis.
Although the ceRNA field remains in its infancy, the
rapid discovery of ceRNA-associated miRNAs and miRNA targets has
contributed to the development of miRNA-based therapeutics
(64). However, certain questions
need to be addressed. The developing progress of antisense or small
interfering RNA drugs has been limited by stability, specificity
and delivery problems. In addition, since a single miRNA is able to
recognize and inhibit a large number of target genes, altering a
single miRNA may affect multiple, unintended genes. Conversely, a
single gene may be targeted by numerous miRNAs, and whether varying
one miRNA is sufficient to affect a specific gene target remains to
be elucidated (65). The ceRNA
mechanism provides a novel perspective to gene therapy for cancer.
ceRNAs are natural miRNA sponges and may possess improved
biological stability compared with that of miRNAs, particularly
circRNAs (31,32). In addition, ceRNAs combine and
sequester multiple miRNAs, and these common miRNAs ensure that the
specific ceRNA target harboring the common MREs will be largely
affected. Furthermore, off-target effects are significantly
reduced. Taking into consideration the undesired gene targets
altered by miRNA transduction, the expression of mRNA-associated
ceRNAs may be restored to physiological levels by ceRNAs, so that
miRNAs and mRNAs will be balanced. Extensive studies of miRNAs and
ceRNAs are required, as these may serve as therapeutic targets for
the treatment of cancer to enhance sensitivity and attenuate drug
resistance.
References
|
1
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Katahira J: Nuclear export of messenger
RNA. Genes (Basel). 6:163–184. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: The Rosetta Stone of a hidden RNA
language? Cell. 146:353–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Poliseno L, Salmena L, Zhang J, Carver B,
Haveman WJ and Pandolfi PP: A coding-independent function of gene
and pseudogene mRNAs regulates tumour biology. Nature.
465:1033–1038. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ebert MS, Neilson JR and Sharp PA:
MicroRNA sponges: Competitive inhibitors of small RNAs in mammalian
cells. Nat Methods. 4:721–726. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ebert MS and Sharp PA: Emerging roles for
natural microRNA sponges. Curr Biol. 20:R858–R861. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Franco-Zorrilla JM, Valli A, Todesco M,
Mateos I, Puga MI, Rubio-Somoza I, Leyva A, Weigel D, García JA and
Paz-Ares J: Target mimicry provides a new mechanism for regulation
of microRNA activity. Nat Genet. 39:1033–1037. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ala U, Karreth FA, Bosia C, Pagnani A,
Taulli R, Léopold V, Tay Y, Provero P, Zecchina R and Pandolfi PP:
Integrated transcriptional and competitive endogenous RNA networks
are cross-regulated in permissive molecular environments. Proc Natl
Acad Sci USA. 110:7154–7159. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Arvey A, Larsson E, Sander C, Leslie CS
and Marks DS: Target mRNA abundance dilutes microRNA and siRNA
activity. Mol Syst Biol. 6:3632010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Karreth FA, Tay Y, Perna D, Ala U, Tan SM,
Rust AG, DeNicola G, Webster KA, Weiss D, Perez-Mancera PA, et al:
In vivo identification of tumor-suppressive PTEN ceRNAs in
an oncogenic BRAF-induced mouse model of melanoma. Cell.
147:382–395. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ling S, Birnbaum Y, Nanhwan MK, Thomas B,
Bajaj M, Li Y, Li Y and Ye Y: Dickkopf-1 (DKK1) phosphatase and
tensin homolog on chromosome 10 (PTEN) crosstalk via microRNA
interference in the diabetic heart. Basic Res Cardiol. 108:3522013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lee DY, Jeyapalan Z, Fang L, Yang J, Zhang
Y, Yee AY, Li M, Du WW, Shatseva T and Yang BB: Expression of
versican 3′-untranslated region modulates endogenous microRNA
functions. PLoS One. 5:e135992010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee DY, Shatseva T, Jeyapalan Z, Du WW,
Deng Z and Yang BB: A 3′-untranslated region (3′UTR) induces organ
adhesion by regulating miR-199a* functions. PLoS One. 4:e45272009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fang L, Du WW, Yang X, Chen K, Ghanekar A,
Levy G, Yang W, Yee AJ, Lu WY, Xuan JW, et al: Versican
3′-untranslated region (3′-UTR) functions as a ceRNA in inducing
the development of hepatocellular carcinoma by regulating miRNA
activity. FASEB J. 27:907–919. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nagano T and Fraser P: No-nonsense
functions for long noncoding RNAs. Cell. 145:178–181. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mercer TR and Mattick JS: Structure and
function of long noncoding RNAs in epigenetic regulation. Nat
Struct Mol Biol. 20:300–307. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Griffiths-Jones S, Saini HK, van Dongen S
and Enright AJ: miRBase: Tools for microRNA genomics. Nucleic Acids
Res. 36:D154–D158. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Paraskevopoulou MD, Georgakilas G,
Kostoulas N, Reczko M, Maragkakis M, Dalamagas TM and Hatzigeorgiou
AG: DIANA-LncBase: Experimentally verified and computationally
predicted microRNA targets on long non-coding RNAs. Nucleic Acids
Res. 41:D239–D245. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chiyomaru T, Fukuhara S, Saini S, Majid S,
Deng G, Shahryari V, Chang I, Tanaka Y, Enokida H, Nakagawa M, et
al: Long non-coding RNA HOTAIR is targeted and regulated by miR-141
in human cancer cells. J Biol Chem. 289:12550–12565. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cesana M, Cacchiarelli D, Legnini I,
Santini T, Sthandier O, Chinappi M, Tramontano A and Bozzoni I: A
long noncoding RNA controls muscle differentiation by functioning
as a competing endogenous RNA. Cell. 147:358–369. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang J, Liu X, Wu H, Ni P, Gu Z, Qiao Y,
Chen N, Sun F and Fan Q: CREB up-regulates long non-coding RNA,
HULC expression through interaction with micro RNA-372 in liver
cancer. Nucleic Acids Res. 38:5366–5383. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kallen AN, Zhou XB, Xu J, et al: The
imprinted H19 lncRNA antagonizes Let-7 microRNAs. Mol Cell.
52:101–112. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Harrison PM, Zheng D, Zhang Z, Carriero N
and Gerstein M: Transcribed processed pseudogenes in the human
genome: An intermediate form of expressed retrosequence lacking
protein-coding ability. Nucleic Acids Res. 33:2374–2383. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pink RC, Wicks K, Caley DP, Punch EK,
Jacobs L and Carter DR: Pseudogenes: Pseudo-functional or key
regulators in health and disease? RNA. 17:792–798. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
D'Errico I, Gadaleta G and Saccone C:
Pseudogenes in metazoa: Origin and features. Brief Funct Genomic
Proteomic. 3:157–167. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Marques AC, Tan J, Lee S, Kong L, Heger A
and Ponting CP: Evidence for conserved post-transcriptional roles
of unitary pseudogenes and for frequent bifunctionality of mRNAs.
Genome Biol. 13:R1022012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Capel B, Swain A, Nicolis S, Hacker A,
Walter M, Koopman P, Goodfellow P and Lovell-Badge R: Circular
transcripts of the testis-determining gene Sry in adult mouse
testis. Cell. 73:1019–1030. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hansen TB, Jensen TI, Clausen BH, Bramsen
JB, Finsen B, Damgaard CK and Kjems J: Natural RNA circles function
as efficient microRNA sponges. Nature. 495:384–388. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Memczak S, Jens M, Elefsinioti A, Torti F,
Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer
M, et al: Circular RNAs are a large class of animal RNAs with
regulatory potency. Nature. 495:333–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jeck WR, Sorrentino JA, Wang K, Slevin MK,
Burd CE, Liu J, Marzluff WF and Sharpless NE: Circular RNAs are
abundant, conserved and associated with ALU repeats. RNA.
19:141–157. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Salzman J, Gawad C, Wang PL, Lacayo N and
Brown PO: Circular RNAs are the predominant transcript isoform from
hundreds of human genes in diverse cell types. PLoS One.
7:e307332012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhou X, Gao Q, Wang J, Zhang X, Liu K and
Duan Z: Linc-RNA-RoR acts as a ‘sponge’ against mediation of the
differentiation of endometrial cancer stem cells by microRNA-145.
Gynecol Oncol. 133:333–339. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Slaidina M and Lehmann R: Translational
control in germline stem cell Development. J Cell Biol. 207:13–21.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gutschner T and Diederichs S: The
hallmarks of cancer: A long non-coding RNA point of view. RNA Biol.
9:703–719. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jeyapalan Z, Deng Z, Shatseva T, Fang L,
He C and Yang BB: Expression of CD44 3′-untranslated region
regulates endogenous microRNA functions in tumorigenesis and
angiogenesis. Nucleic Acids Res. 39:3026–3041. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Voutilainen K, Anttila M, Sillanpää S,
Tammi R, Tammi M, Saarikoski S and Kosma VM: Versican in epithelial
ovarian cancer: Relation to hyaluronan, clinicopathologic factors
and prognosis. Int J Cancer. 107:359–364. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Herrlich P, Morrison H, Sleeman J,
Orian-Rousseau V, König H, Weg-Remers S and Ponta H: CD44 acts both
as a growth- and invasiveness-promoting molecule and as a
tumor-suppressing cofactor. Ann NY Acad Sci. 910:106–118. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kumar MS, Armenteros-Monterroso E, East P,
Chakravorty P, Matthews N, Winslow MM and Downward J: HMGA2
functions as a competing endogenous RNA to promote lung cancer
progression. Nature. 505:212–217. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Di Cello F, Hillion J, Hristov A, Wood LJ,
Mukherjee M, Schuldenfrei A, Kowalski J, Bhattacharya R, Ashfaq R
and Resar LM: HMGA2 participates in transformation in human lung
cancer. Mol Cancer Res. 6:743–750. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Meyer B, Loeschke S, Schultze A, Weigel T,
Sandkamp M, Goldmann T, Vollmer E and Bullerdiek J: HMGA2
overexpression in non-small cell lung cancer. Mol Carcinog.
46:503–511. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Winslow MM, Dayton TL, Verhaak RG,
Kim-Kiselak C, Snyder EL, Feldser DM, Hubbard DD, DuPage MJ,
Whittaker CA, Hoersch S, et al: Suppression of lung adenocarcinoma
progression by Nkx2-1. Nature. 473:101–104. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Endo H, Shiroki T, Nakagawa T, Yokoyama M,
Tamai K, Yamanami H, Fujiya T, Sato I, Yamaguchi K, Tanaka N, et
al: Enhanced expression of long non-coding RNA HOTAIR is associated
with the development of gastric cancer. PLoS One. 8:e770702013.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hajjari M, Behmanesh M, Sadeghizadeh M and
Zeinoddini M: Up-regulation of HOTAIR long non-coding RNA in human
gastric adenocarcinoma tissues. Med Oncol. 30:6702013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Liu XH, Sun M, Nie FQ, Ge YB, Zhang EB,
Yin DD, Kong R, Xia R, Lu KH, Li JH, et al: Lnc RNA HOTAIR
functions as a competing endogenous RNA to regulate HER2 expression
by sponging miR-331-3p in gastric cancer. Mol Cancer. 13:922014.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
van der Wal JE, Hermsen MA, Gille HJ,
Schouten-Van Meeteren NY, Moll AC, Imhof SM, Meijer GA, Baak JP and
van der Valk P: Comparative genomic hybridisation divides
retinoblastomas into a high and a low level chromosomal instability
group. J Clin Pathol. 56:26–30. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zimonjic DB, Keck CL, Thorgeirsson SS and
Popescu NC: Novel recurrent genetic imbalances in human
hepatocellular carcinoma cell lines identified by comparative
genomic hybridization. Hepatology. 29:1208–1214. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Plantaz D, Mohapatra G, Matthay KK,
Pellarin M, Seeger RC and Feuerstein BG: Gain of chromosome 17 is
the most frequent abnormality detected in neuroblastoma by
comparative genomic hybridization. Am J Pathol. 150:81–89.
1997.PubMed/NCBI
|
|
51
|
Tay Y, Kats L, Salmena L, Weiss D, Tan SM,
Ala U, Karreth F, Poliseno L, Provero P, Di Cunto F, et al:
Coding-independent regulation of the tumor suppressor PTEN by
competing endogenous mRNAs. Cell. 147:344–357. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Gregory PA, Bert AG, Paterson EL, Barry
SC, Tsykin A, Farshid G, Vadas MA, Khew-Goodall Y and Goodall GJ:
The miR-200 family and miR-205 regulate epithelial to mesenchymal
transition by targeting ZEB1 and SIP1. Nat Cell Biol. 10:593–601.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Vandewalle C, Comijn J, De Craene B,
Vermassen P, Bruyneel E, Andersen H, Tulchinsky E, Van Roy F and
Berx G: SIP1/ZEB2 induces EMT by repressing genes of different
epithelial cell-cell junctions. Nucleic Acids Res. 33:6566–6578.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Fan M, Li X, Jiang W, Huang Y, Li J and
Wang Z: A long non-coding RNA, PTCSC3, as a tumor suppressor and a
target of miRNAs in thyroid cancer cells. Exp Ther Med.
5:1143–1146. 2013.PubMed/NCBI
|
|
55
|
Loewer S, Cabili MN, Guttman M, Loh YH,
Thomas K, Park IH, Garber M, Curran M, Onder T, Agarwal S, et al:
Large intergenic non-coding RNA-RoR modulates reprogramming of
human induced pluripotent stem cells. Nat Genet. 42:1113–1117.
2010. View
Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wang Y, Xu Z, Jiang J, Xu C, Kang J, Xiao
L, Wu M, Xiong J, Guo X and Liu H: Endogenous miRNA sponge
lincRNA-RoR regulates Oct4, Nanog and Sox2 in human embryonic stem
cell self-renewal. Dev Cell. 25:69–80. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Rutnam ZJ and Yang BB: The non-coding
3′UTR of CD44 induces metastasis by regulating extracellular matrix
functions. J Cell Sci. 125:2075–2085. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Waalkes S, Atschekzei F, Kramer MW,
Hennenlotter J, Vetter G, Becker JU, Stenzl A, Merseburger AS,
Schrader AJ, Kuczyk MA and Serth J: Fibronectin 1 mRNA expression
correlates with advanced disease in renal cancer. BMC Cancer.
10:5032010. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ji J, Zhao L, Budhu A, Forgues M, Jia HL,
Qin LX, Ye QH, Yu J, Shi X, Tang ZY and Wang XW: Let-7g targets
collagen type I alpha2 and inhibits cell migration in
hepatocellular carcinoma. J Hepatol. 52:690–697. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Ferrara N: Vascular endothelial growth
factor. Arterioscler Thromb Vasc Biol. 29:789–791. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Sumazin P, Yang X, Chiu HS, Chung WJ, Iyer
A, Llobet-Navas D, Rajbhandari P, Bansal M, Guarnieri P, Silva J
and Califano A: An extensive microRNA-mediated network of RNA-RNA
interactions regulates established oncogenic pathways in
glioblastoma. Cell. 147:370–381. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Giovannetti E, Erozenci A, Smit J, Danesi
R and Peters GJ: Molecular mechanisms underlying the role of
microRNAs (miRNAs) in anticancer drug resistance and implications
for clinical practice. Crit Rev Oncol Hematol. 81:103–122. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Allen KE and Weiss GJ: Resistance may not
be futile: MicroRNA biomarkers for chemoresistance and potential
therapeutics. Mol Cancer Ther. 9:3126–3136. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Cho WC: MicroRNAs: Potential biomarkers
for cancer diagnosis, prognosis and targets for therapy. Int J
Biochem Cell Biol. 42:1273–1281. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Cho WC: MicroRNAs in cancer-from research
to therapy. Biochim Biophys Acta. 1805:209–217. 2010.PubMed/NCBI
|














