Introduction
Intrinsic or acquired resistance of tumour cells to
chemotherapy or radiotherapy remains a major obstacle to successful
cancer management. Mechanisms leading to resistance are diverse and
poorly defined; however, recent experimental data support the
concept that cancer stem cells (CSCs) are more radioresistant and
chemoresistant than their non-stem counterparts (1–3). CSCs
display stem-like characteristics and are initially defined as
cells endowed with long-term self-renewal and differentiation
capacity. In solid tumours, CSCs have been proposed to represent a
small proportion of tumour cells; they were also reported to be
capable of forming colonies in an in vitro clonogenic assay
and tumours in an in vivo assay (4). In breast cancer, CSCs were first
described as a population bearing the
ESA+/CD44+/CD24− phenotype, with a
50-fold higher capability to form tumours in immunodeficient mice
and to differentiate into di°stinct cellular subtypes (4,5). In breast
cancer cell lines, CD44+/CD24− cells were
also described as a subpopulation bearing an invasive capacity and
a genetic signature underlying an aggressive phenotype (6,7). Breast
CSCs have been characterised by a number of markers, among which
CD44+/CD24−/low is the most widely used.
However, other markers have also been associated with CSC
characteristics, including the presence of a side population
(Hoechst 33342 dye exclusion), aldehyde dehydrogenase activity and
other prospective markers, including CD133, ESA, PROCR and CXCR4
(8).
DNA damage activates signal transduction pathways
referred to as checkpoints, which delay cell cycle progression and
allow more time for DNA repair (9).
Checkpoints arrest cells in the G1 phase to prevent replication of
damaged DNA and in the G2 phase to prevent the segregation of
damaged chromosomes during mitosis (9). Increased levels of phosphocholine (PC)
is one of the hallmarks of cancer, and numerous studies have
established a strong correlation between increased PC and malignant
progression (9,10). One of the major causes of high PC in
tumours is the increase in the expression and activity of
checkpoint kinase (CHK), a rate-limiting enzyme that phosphorylates
and converts choline to PC (10–12). CHK
has been previously targeted with novel pharmacological inhibitors
(13,14) and posttranscriptional gene silencing
(15). The pharmacological inhibition
of CHK cancer cells results in growth arrest and apoptosis
(13).
Numerous previous studies have investigated the CHK
pathway in breast cancer cell lines. However, few studies have
investigated the CHK pathway in breast cancer stem cells. Bensimon
et al (16) reported that CD24
is associated with the transmission of genomic instability, which
leads tumour cells to acquire more aggressive characteristics. The
present study aimed to investigate the association between the CHK
pathway and the stem cell population of breast cancer cell line,
MCF-7. Curman et al (17)
reported that debromohymenialdisine (DBH) blocks two major branches
of the checkpoint pathway downstream of the serine/threonine kinase
ATM, thereby preventing the activation or inhibition of different
signal transduction proteins and inhibiting a narrow range of
protein kinases in vivo. Therefore, the present study
investigated the DBH-inhibited cell cycle CHKl/2 DNA repair system
signal pathway in MCF-7 cancer stem cells to explore the survival
impact and the molecular mechanisms of radiotherapy.
Materials and methods
Cell culture
The MCF-7 human breast cancer cell line was acquired
from American Type Culture Collection (Manassas, VA, USA) and
cultured in minimal essential media (Sigma-Aldrich, St. Louis, MO,
USA) supplemented with 10% (v/v) fetal bovine serum with 100
units/ml penicillin and 100 µg/ml streptomycin (Thermo Fisher
Scientific, Inc., Atlanta, GA, USA). The cells were cultured in
standard cell culture incubator conditions at 37°C in a humidified
atmosphere containing 5% CO2.
Grouping and cell irradiation
Linear accelerator X-ray (6 MV) at dose rate of 2
Gy/min was administered with a gantry rotation 180°. Irradiation
(IR) was performed through the bottom of the cell culture plate
with the source at a distance of 100 cm (equivalent to 1.5 cm
tissue) in a radiation field size of 10×10 cm. The following
experimental groups were established: Control group, A group (DBH),
B group (2 Gy IR), B1 group (2 Gy IR + DBH), C group (5 Gy IR) and
C1 group (5 Gy IR + DBH). DBH (Enzo Life Sciences, Farmingdale, NY,
USA) was supplemented with 3 µM/l Dulbecco's Modified Eagle's
medium.
Western blot analysis
Total protein from MCF-7 cells was extracted using a
cracking buffer [100 mmol/l Tris (pH 6.7), 2% glycerol] containing
a protease inhibitor (Sigma-Aldrich) at a 1:200 dilution, resolved
on 10% SDS-PAGE for immunoblot analysis and then incubated using
custom-made rabbit polyclonal antibody against human-CHK1/CHK2
(Cell Signalling Technology, Inc., Danvers, MA, USA) at 1:100
dilution in 5% nonfat dry milk overnight at 4°C. A mouse monoclonal
antibody against human-β-actin (Sigma-Aldrich) at 1:10,000 was used
as control. Appropriate horseradish peroxidase-conjugated secondary
antibody, either anti-mouse or anti-rabbit (GE Healthcare Life
Sciences, Chalfont, UK), was used at 1:2,500 dilution in milk.
Immunoblots were developed using the Super Signal West Pico
chemiluminescent substrate kit (Pierce Biotechnology, Inc.,
Rockford, IL, USA) and images were captured using a Digimax i50
digital camera (Samsung, Suwon, South Korea). The density of
immunoblot bands was analyzed using Band Leader software (version
3.0; Band Leader Systems, Inc., Boulder City, NV, USA) as described
previously (18).
Methylthiazyl blue tetrazolium bromide
(MTT) viability assay
The MCF-7 cells were cultured in vitro in 96-well
plates. The concentration was adjusted to 105 cells/ml.
A total of 100 µl of the cell suspension was added to each well
(edge holes were filled with sterile phosphate-buffered solution or
PBS to maintain humidity), and maintained at 4.5% CO2,
37°C. The cells were assigned to the dosing and radiation grouping
as above, and cultured for 24, 48 or 72 h. The OD value of each
well was measured at a detection wavelength of 570 nm using a
microplate reader (Synergy H1 Multi-Mode Reader; BioTek, Shanghai,
China), compared with the blank control well (medium, MTT, dimethyl
sulfoxide).
IR (%) = [(OD control group - OD experimental group)
/ OD of control group] × 100.
Flow cytometry
The proportion of the stem cell-like MCF-7 cells was
investigated following radiotherapy. Cells were treated with 0, 2
and 5 Gy IR. All groups were cultured for 48 h. The MCF-7 cell
culture was digested with 0.25% trypsin to produce a single cell
suspension. Digestion was terminated by adding a culture solution
of 10% fetal calf serum. The cells were centrifuged at 200 × g for
10 min and then washed twice with PBS and the cell concentration
was adjusted to 1×106 cells/ml. Approximately 40 µl of
the cell suspension was placed in a flow cytometry test tube, added
with 0.5% bovine serum albumin (BSA) was added and then the samples
were incubated at room temperature for 30 min. Approximately 20 µl
of CD24-FITC mouse monoclonal conjugated antibody (1:300) was added
to CD44-PE mouse monoclonal antibody (1:300; R&D Systems China
Co., Ltd., Shanghai, China) and then incubated in the dark at room
temperature for 20 min. The cells were washed twice with PBS (3 ml)
to remove excess antibodies. The cells were then re-suspended in 3
ml PBS and analysed using a BD Accuri™ C6 flow cytometer (Becton
Dickinson, Franklin Lakes, NJ, USA) at 488 nm/520 nm. Three
parallel samples were run (Separate blank, CD44-PE, CD24-FITC
control tube).
Direct immunofluorescence
microscopy
MCF-7 cells in the logarithmic growth phase were
seeded on sterilised glass slides in a 24-well cell culture plate
(50,000 cells/well). The following groups were established: Control
group, A group (DBH), B group (2 Gy IR), B1 group (2 Gy IR + DBH),
C group (5 Gy IR) and C1 group (5 Gy IR + DBH). Adherent cells that
survived were subjected to 5 Gy radiation and then cultured for 1
to 8 days. Parallel experiments were performed in triplicate.
Following incubation, the cells were washed twice with PBS, cooled
to 4°C, and then fixed with methanol for 10 min. The slides were
washed 4 times with PBS and then 1% BSA blocking solution was added
dropwise at 4°C for 1 h. The slides were washed 4 times with PBS.
Mouse anti-human PE-CD44-IgG (red) and mouse anti-human
FITC-CD24-IgG (Green) at 1:200 dilution were added dropwise to the
samples. The samples were then incubated in the dark for 60 min.
Subsequently, the samples were washed one to two times with PBS (pH
7.2 to 7.4). The cells were stained with 20 µl DAPI (Sigma-Aldrich)
by dropwise addition at ambient temperature in the dark. After
allowing to stand for 10 min, the slides were washed with PBS.
Anti-fade mounting medium (Beyotime Institute of Biotechnology,
Shanghai, China) was added dropwise. The cells were observed by
fluorescence microscopy (IX51; Olympus Co., Ltd., Shanghai, China).
Three independent experiments were performed, the results were
counted and the means were calculated.
Statistical analysis
Statistical analysis was performed using SPSS
software, version 13.0 (SPSS, Inc., Chicago, IL, USA). All data are
presented as the mean ± standard deviation and one-way analysis of
variance and Dunnett's T3 post test were used to determine the
statistical significance. Differences between groups were analyzed
using two-sided t-tests. P<0.05 and P<0.01 were considered to
indicate a statistical difference and statistically significant
difference, respectively.
Results
Radiation may result in DNA damage and increase the
pCHK1/CHK2 level in breast cancer cells, DBH inhibits the
phosphorylation of cell cycle checkpoint kinase CHK1/2 specifically
(13–15). In the present study, western blot
analysis demonstrated that pCHK1/2 was markedly increased following
24 h of radiotherapy with either low dose radiation at 2 Gy or high
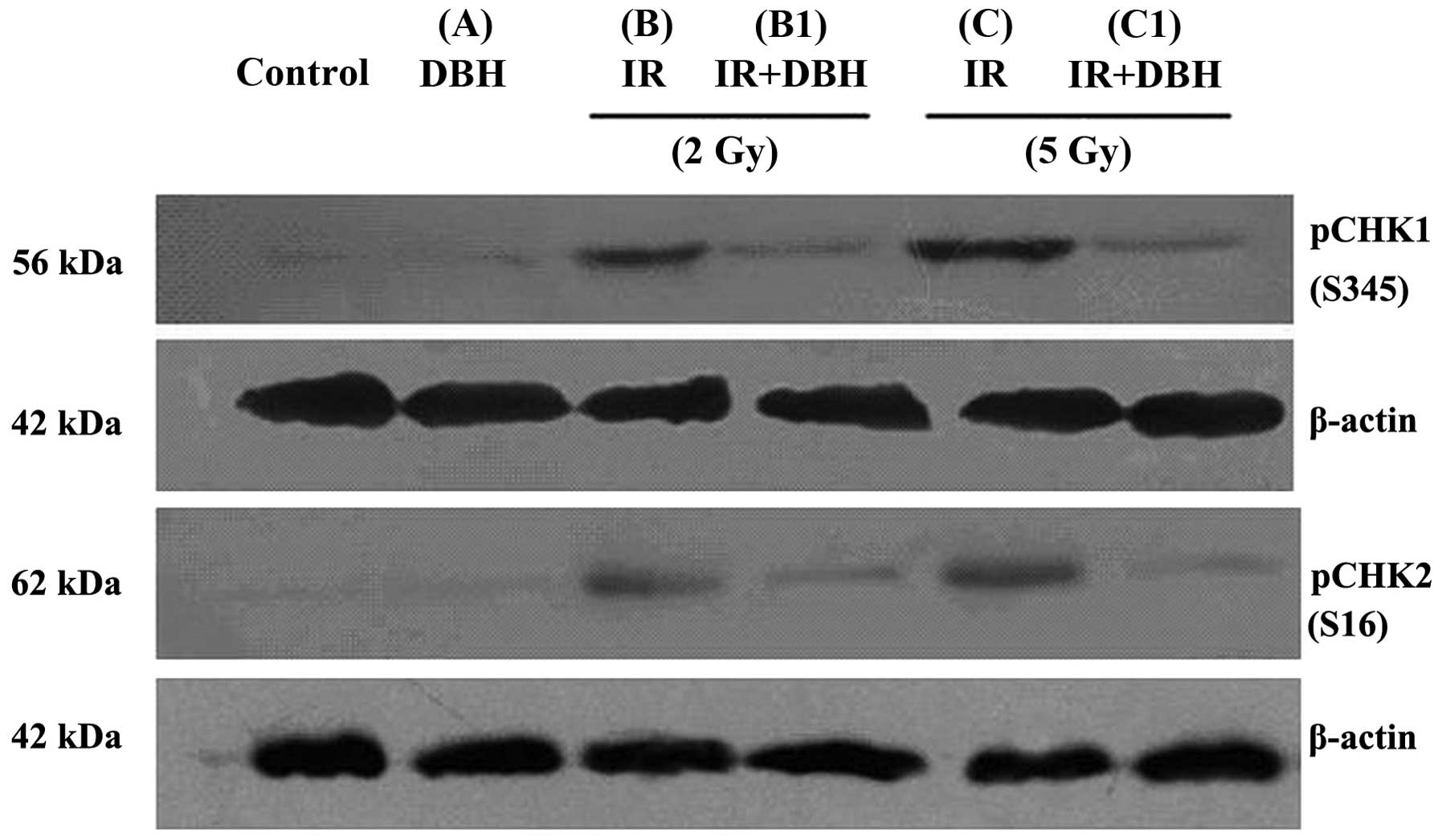
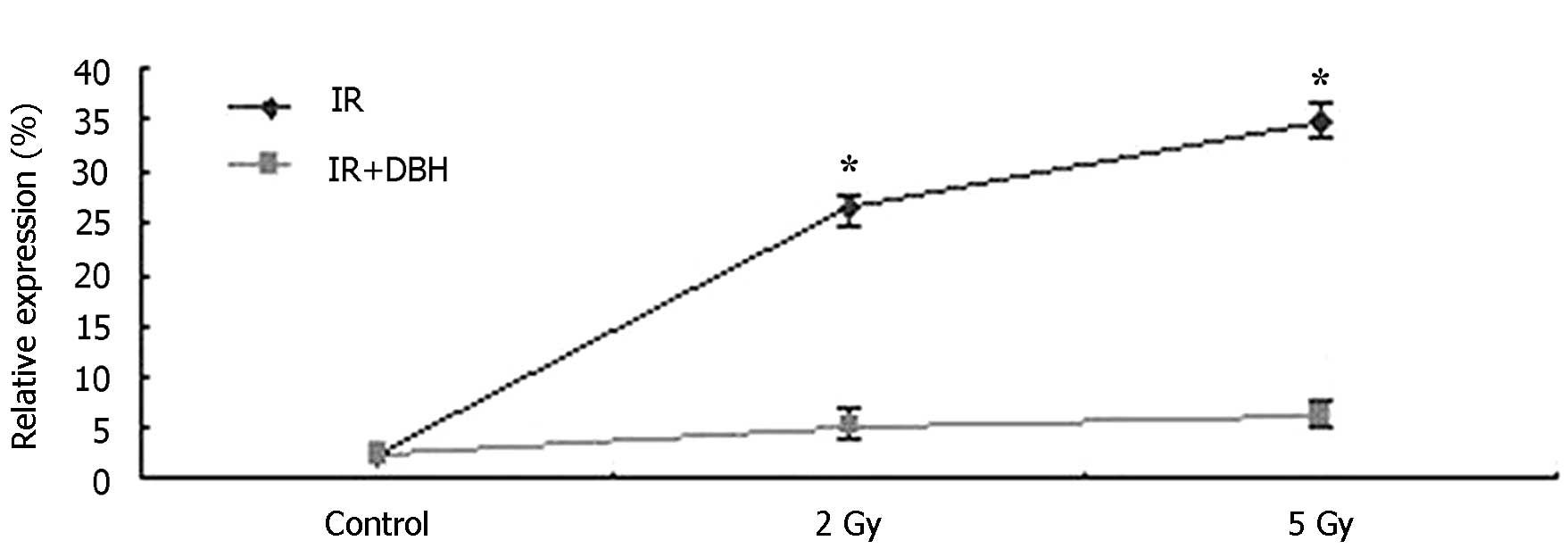
dose radiation at 5 Gy, as presented in Fig. 1 (Bandleader calculation). The pCHK1
protein levels in the control, B (treated with 2 Gy radiation dose)
and C (treated with 5 Gy radiation dose) groups were 1.48±0.11,
26.29±0.24, and 39.72±1.45%, respectively. pCHK2 protein level
exhibited the same trend. CHK protein was therefore significantly
activated in the 24 h following radiation. In addition, the
activation of CHK proteins by radiation was dose-dependent in the
MCF-7 cell line. DBH had no effect on the activation of CHK protein
in the MCF-7 cell line not treated with radiation. However it
appeared to serve a role in the MCF-7 cells which were treated by
radiation. The pCHK1 protein compared with actin in the B1 group (2
Gy radiation and 3 µM DHB application) and C1 group (5 Gy radiation
and 3 µM DHB application) were 5.46±1.45% and 6.02±1.39%,
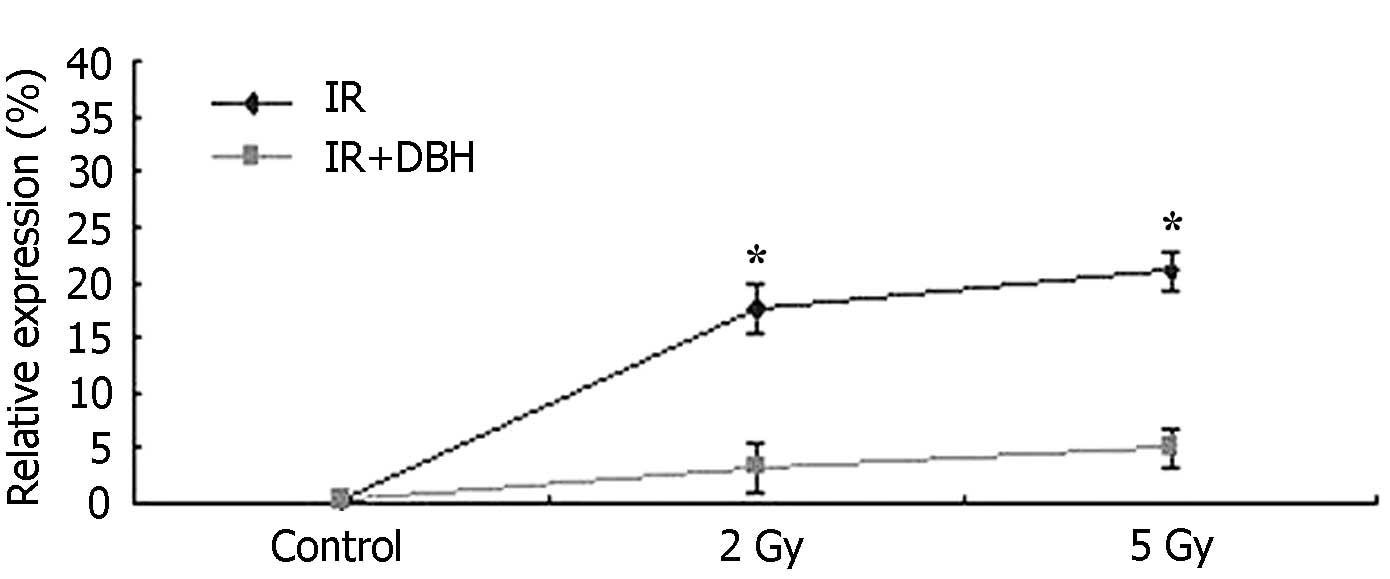
respectively (P<0.05) (Fig. 2).
pCHK1 protein expression markedly reduced following low or high
dose radiation when DHB was also applied. With low dose radiation,
the pCHK1 protein was reduced by 79.23±3.80% (2 Gy) compared with B
group. With high dose radiation, the pCHK1 protein was reduced by
82.67±4.19% (5 Gy) compared with C group (Fig. 3).
DBH inhibits MCF-7 proliferation
following radiotherapy
Methylthiazyl blue tetrazolium bromide (MTT)
viability assay is presented in Table
I: No significant difference was observed in the inhibition
rate of breast cancer MCF-7 cells in the simple dosing DBH group
compared with the control group (P>0.05). This result indicates
that the experimental DBH drug concentration (3 µM) had no
significant cytotoxicity. Following 24, 48, and 72 h of
radiotherapy, the inhibition rates of the B group were 21.43±3.19%,
36.36±5.47%, and 47.79±9.16%, respectively. After 24, 48, and 72 h
of radiotherapy, the inhibition rates of the B1 group were
60.71±5.23, 67.27±3.74, and 73.45±5.72%, respectively. The
inhibition rate of the C1 group was 65.18±6.41, 72.73±10.18,
80.53±9.16%, which was increased compared with the B1 group. These
results indicated that the inhibition rate of MCF-7 was time and
dose dependent. In addition, DBH may increase the sensitivity of
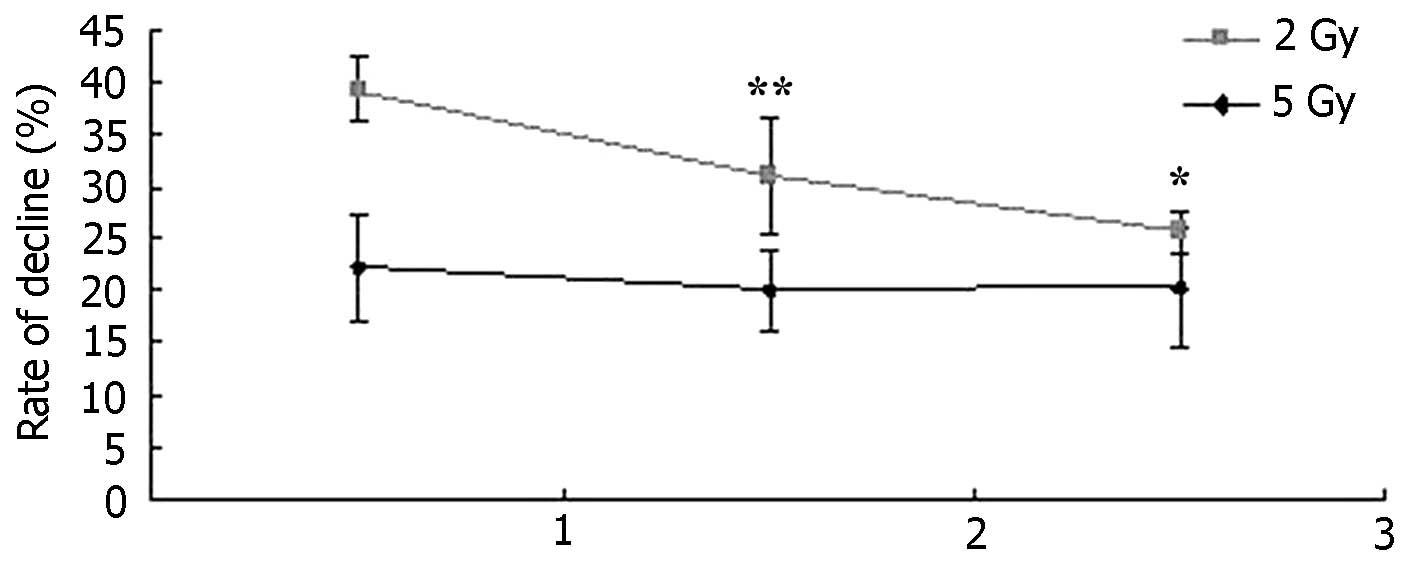
radiotherapy by inhibiting the CHK signal pathway. The percentage
of the downregulation of inhibition of MCF-7 cells between the B1
and C1 groups and their control groups B and C were calculated and
compared at different time periods (Fig.
4). The inhibition rate increased in Group B1, where cells were
simultaneously treated with the low dose radiation and application
of 3 µM DBH. In group B1, increasing the incubation time with DBH
contributed to the increase in inhibition rate following
radiotherapy. However, the same trend was not observed in Group C1,
the inhibition rate in Group C1 was not statistically different at
longer culture time, when cells were simultaneously treated with
high dose radiation and application of 3 µM DBH (P>0.05).
Therefore, DBH inhibited the survival of MCF-7 cells following
low-dose radiation and the inhibition rate becomes more effective
as the incubation time with DBH is increased.
 | Table I.Inhibition of MCF-7 by DBH after
radiotherapy. |
Table I.
Inhibition of MCF-7 by DBH after
radiotherapy.
|
| Exposure time | Inhibitive
proportion (%) |
|---|
|
|
|
|
|---|
| Groups | 24 h | 48 h | 72 h | 24 h | 48 h | 72 h |
|---|
| Control | 1.12±0.05 | 1.10±0.07 | 1.13±0.03 | 0 | 0 | 0 |
| A | 1.08±0.06 | 1.15±0.08 | 1.10±0.03 | 3.57±0.21 | 4.54±0.23 | 2.65±0.35 |
| B | 0.88±0.07 | 0.70±0.03 | 0.59±0.04 | 21.43±3.19 | 36.36±5.47 | 47.79±9.16 |
| B1a | 0.44±0.03 | 0.36±0.02 | 0.30±0.08 | 60.71±5.23 | 67.27±3.74 | 73.45±5.72 |
| C | 0.64±0.05 | 0.52±0.02 | 0.45±0.01 | 42.86±2.09 | 52.72±8.68 | 60.18±7.81 |
| C1b | 0.39±0.02 | 0.30±0.06 | 0.22±0.04 | 65.18±6.41 | 72.73±10.18 | 80.53±9.16 |
Increase in the proportion of
CD44+CD24− MCF-7 stem cells following
radiotherapy
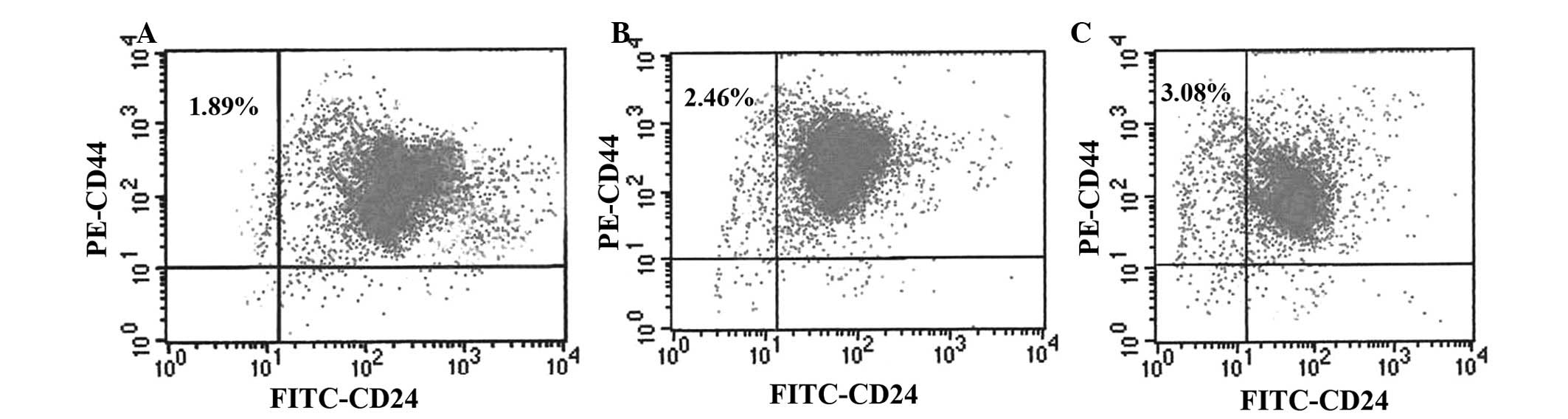
The flow cytometry excitation wavelength was 488 nm.
The PE and FITC emitted light was collected at 525 and 575 nm,
respectively. The results demonstrated that the breast cancer MCF-7
cell line was composed of four subpopulations:
CD44+CD24+ (95.04±2.15%),
CD44+CD24− (1.89±0.20%),
CD44−CD24+ (1.65±0.33%), and
CD44−CD24− (1.41±0.17%). The majority of the
MCF-7 cell line were CD44+CD24+ cells.
CD44+CD24− cells were rare, and may be
regarded as stem cells in MCF-7 cell line (Fig. 5A). Following irradiation, the
CD44+CD24− ratio in the 2 Gy irradiation
group increased to 2.46±0.27% (Fig.
5B), and that of the 5 Gy irradiation group reached 3.08±0.21%
(Fig. 5C). The results demonstrated
that exposure to radiation results in the increase of
CD44+CD24− cell population in the MCF-7 cell
line. The ratio of CD44+CD24− MCF-7 cell line
increased gradually with increasing radiation dose (P<0.05).
Increase in
CD44+CD24− MCF-7 cell population following
radiotherapy was inhibited by DBH
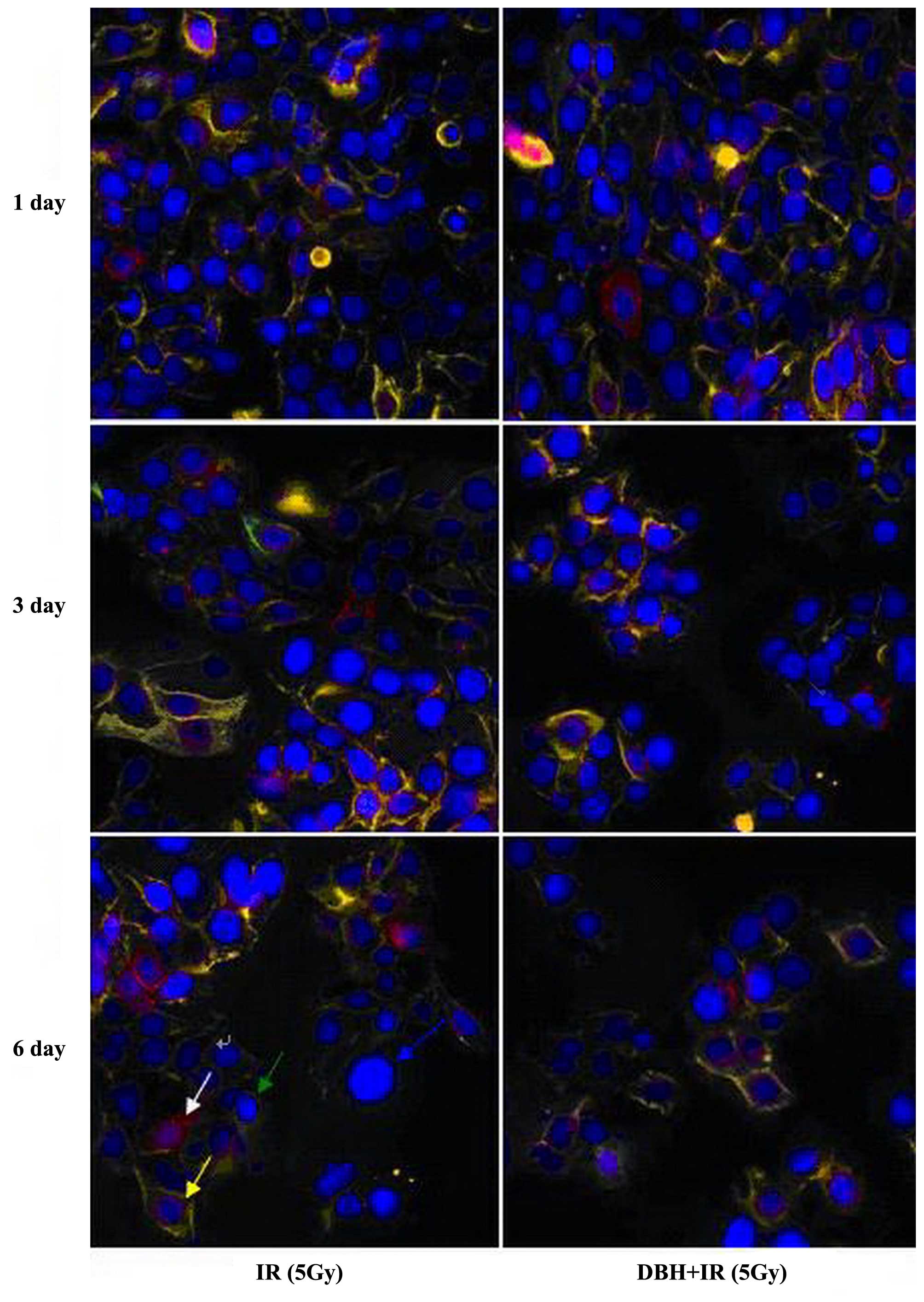
In the direct immunofluorescence microscopy,
PE-CD44-IgG and FITC-CD24-IgG were red and green, respectively. The
strength of CD44 and CD24 expression levels on the cell membrane
can be determined. CD44+CD24+ had yellow
fluorescence, CD44+CD24− had red,
CD44−CD24+ had green, and
CD44−CD24− only showed deep blue nuclear DAPI
fluorescence. In the control group and the dosing group, the
CD44+CD24− cell ratio was 1.89±0.20%, and
CD44+CD24+ cells accounted for 95.04±2.15% of
the total cell population. The ratio of
CD44+CD24− cancer stem cells significantly
increased following 5 Gy irradiation, and the activation of
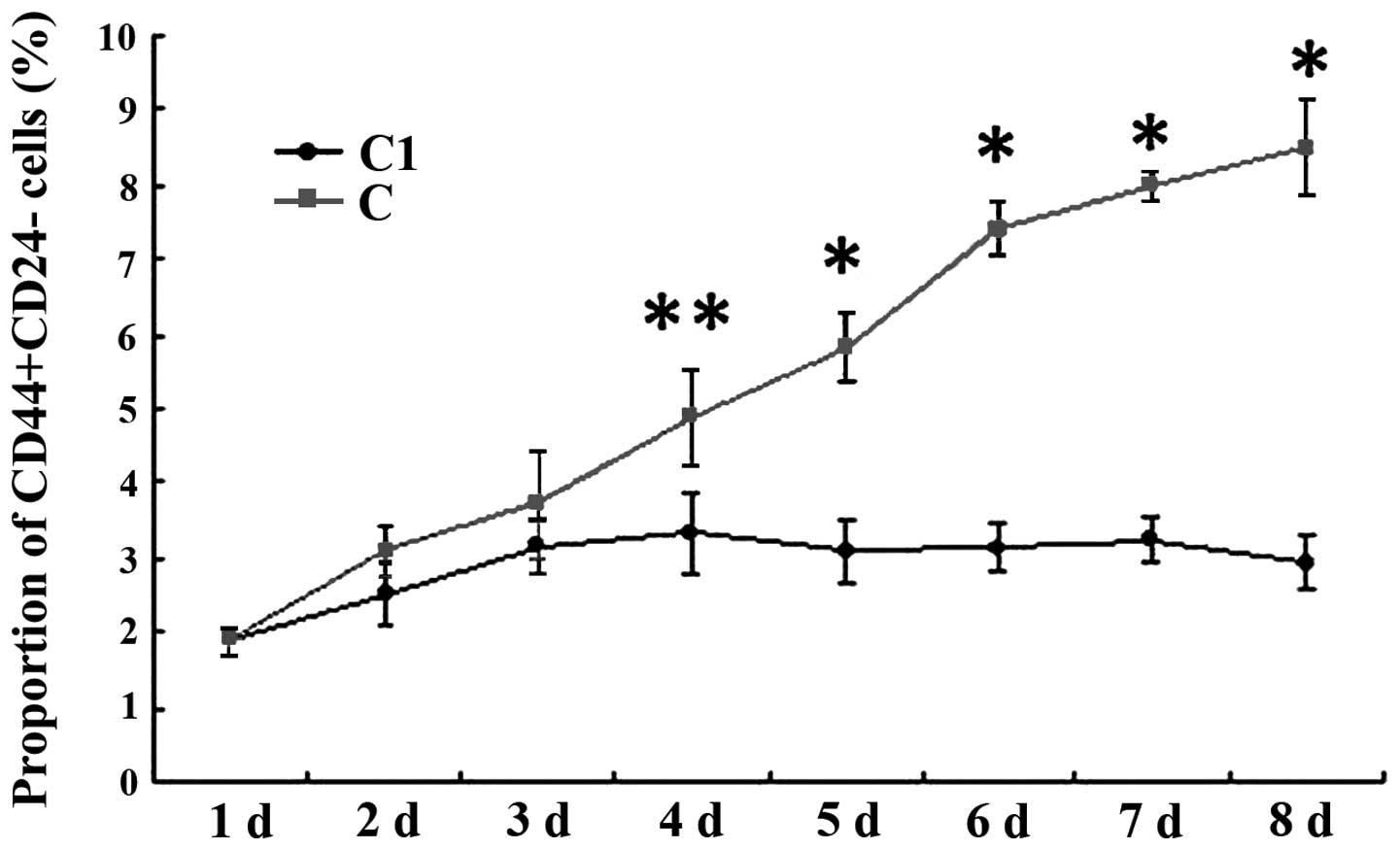
CD44+CD24− cells was time dependent (Fig. 6). In the DBH with irradiation group,
the proportion of CD44+CD24− cancer stem
cells was slightly increased in the first 3 days and then reduced
and remained stable at 3.73±0.35%. However, the proportion of
CD44+CD24− in the DBH with irradiation group
was always reduced compared with the irradiation group alone
(Fig. 7). DBH may inhibit the CHK
signal pathway therefore inhibiting the breast cancer stem cells
from being activated by the radiotherapy.
Discussion
Radiotherapy may result in damaged DNA. It has an
important role in breast cancer treatment. However, radiation
resistance of breast cancer remains a challenge. Previous studies
have demonstrated the mechanism of radiation resistance of cancer
cells in a number of aspects, including the level of reactive
oxygen species, histone H2AX phosphorylation of the EGFR signalling
pathway activation (19), Notch
pathway activation (20),
Wnt-β-catenin of surviving signal activation (21,22), the
hypoxic microenvironment (23) and
the cell cycle checkpoint control of cell proliferation cycle,
which has an important function in radiotherapy response (24). The CHKl/2 downstream effector gene
ATM/ATR, located on the cell cycle checkpoint activation pathway
terminal activates DNA damage detection point when DNA is damaged.
The activation is regulated by cell division cycle protein 25A/B/C
(CDC25A/B/C) and the 14-3-3 proteins. CHKl as a cell cycle
checkpoint kinase has a protective role in the M phase checkpoint;
it also alleviates the segregation of damaged chromosomes during
mitosis, thereby reducing cell death (25). CHK2 is expressed throughout the cell
cycle but only when the double-strand break upstream protein ATM is
activated. The activation of breast cancer gene l (breast cancer 1,
BRCAl) by CHK2, in addition to the promotion of BRCA2 expression,
is involved in homologous recombination, nucleotide excision
repair, and DNA repair (26–28). CHKl and CHK2 are activated by
phosphorylation following DNA damage resulting from drugs, ionising
radiation (IR), and ultraviolet radiation (9). ATM and ATR occupy an important position
in radiotherapy-induced DNA repair.
Focusing radiotherapy of breast cancer cells in the
cell cycle checkpoint resistance results in enhanced DNA repair
capacity. Therefore, agents that block DNA damage-induced cell
cycle arrest and reduce DNA repair efficiency may potentially
sensitize breast cancer cells to radiotherapy. DBH is sponge
extract isolated from marine organisms containing pyrrole
seven-membered ring lactam alkaloid. DBH inhibits the activation of
CHK1 and CHK2 (17). Others, such as
17-DMAG of CHK1 inhibitors (29),
UCN-01 (CHK2 inhibitor) (30) and
other drugs had no for both CHK 1/2. The overlapping functions of
CHK1 and CHK2 enhance the ability of tumour cells to protect
themselves, and both can act on Cdc25C. They have functions on the
S and G2 phase detection point, demonstrating mutually
complementary roles (31,32). Therefore, selecting both CHKl and CHK2
as therapeutic targets is more reasonable than either of the two
alone.
In the present study, pCHK1/2 in the breast cancer
MCF-7 cell line radiotherapy following 24 h was detected by western
blot analysis. pCHK1/2 expression level significantly increased
with increasing radiation dose. Therefore, the breast cancer cell
cycle checkpoint pathway reaction enhanced DNA damage repair,
thereby weakening the sensitivity to radiotherapy. When DBH was
coupled with radiotherapy, pCHK1 and pCHK2 were downregulated
(P<0.01), and the radiation dose was positively correlated with
this effect (P<0.05). The downregulation of pCHK1/2 had no
significant correlation with radiotherapy prior to and following
DBH treatment, which indicates that DBH as an efficacy inhibitor is
relatively stable. MCF-7 cell proliferation was also determined
using an MTT assay following radiotherapy. The inhibition of MCF-7
cell proliferation had the same trend as was observed for pCHK1/2
expression levels. Low-dose radiotherapy combined with DBH achieved
a higher MCF-7 inhibition rate compared with high-dose radiation
alone (P<0.01). This finding indicates that the inhibition of
the CHK1/2 molecule signalling pathway reduces cell DNA damage
repair.
Stem cells have the capacity for self-renewal,
unlimited proliferation and differentiation. By comparing stem
cells and tumour cell subsets in cancer research, similarities were
observed between the two, including self-renewal and proliferation
capacity; Notch, Wnt, Sonic hedgehog (Shh) and Bmi21 signalling
pathways involved in cell growth and development; and their ability
to migrate or transfer (33). As
such, the CSC hypothesis was proposed, which indicated that the
presence of a small proportion of tumour cells in the tumour tissue
has a significant role in initiating tumour formation and
maintaining tumour growth. These cells also have a decisive role,
self-renewal capacity and differentiation potential source of
malignant tumour growth, metastasis and recurrence. Al-Hajj et
al (5) isolated a
CD44+CD24−/low-population of cells
from the tissue of breast cancer patients. Following
transplantation of ~200 of these cells in non-obese diabetic/severe
combined immunodeficient mice formed ~1 cm tumours in 5–6 months.
By contrast, no tumourigenic or low tumourigenic ability was
observed in the other MCF-7 cell subtypes. Compared with the
unsorted cells, the CD44+CD24−/low
and ESA+lin− population cells exhibited a
50-fold increase in tumourigenic ability. The resulting tumour
contained the same separable
CD44+CD24−/low
ESA+lin−cancer cells, with the same
tumourigenic ability, which for the first time confirmed the
existence of breast cancer stem cells. Fillmore et al
(1) reaffirmed the phenotype of
CD44+CD24− MCF-7 cells having CSC
characteristics.
The experiments of the present study further
explored the association between the
CD44+CD24− subgroup of MCF-7 cells following
radiotherapy with the CHK1/2 signal pathway. Radiotherapy increased
the population of CD44+CD24− MCF-7 cells,
which was positively correlated with radiation dose and culture
time (P<0.05). With the application of DBH, the dosing of
CD44+CD24− cells reduced following
radiotherapy from 3.08±0.41% to 2.52±0.34%, which is a reduction of
18.18%. This result indicated that the CHKl/2 inhibitor DBH reduced
the stem cell population of MCF-7. The inhibition by DBH of the
CD44+CD24− stem cell population increased
significantly in a time-dependent manner until the eighth day and
then reduced until 64.45% was reached.
The proliferation of the
CD44+CD24− group cells was suppressed
following the inhibition of CHK1/CHK2. This result indicates that
the ATM/ATR-CHK1/CHK2-CDC 25A/25B/25C cell cycle checkpoint arm may
be important in MCF-7 cancer stem cell population radiation
resistance. It may also be compared with other cell subsets. More
extensive activation of DBH inhibition may reverse its CHKl/2
activation following radiotherapy resistance. It is of note that
CD44+CD24− cells in the experiment may not be
separated completely because of the limited separation methods
used. Combined application of immunomagnetic beads, flow cytometry,
and other marker, such as antibodies, to label the breast stem cell
cluster may results in a more precise conclusion. Studies using
serum plus growth factor CD44+CD24− cells
in vitro have been previously reported; however, separate
cell culture conditions are more demanding.
CSC theory introduced a new tumour formation
mechanism to improve understanding of tumour development and
prognosis. The results of the present study may serve as a novel
theoretical foundation for therapeutic targets, which has important
clinical significance. Focusing on cell cycle checkpoint, the
target CHKl/2 may have a potential to reduce CSC resistance to
radiation. However, CSC research remains in its infancy, and
numerous problems are still encountered. Previous studies have also
investigated the CHK signal pathway with cancer stem lines using
siRNA technique or gene analysis (16,34).
Acknowledgements
The present study was approved by the International
Review Board of Soochow University (Suzhou, China)and funded by the
Second Affiliated Hospital of Soochow University Preponderant
Clinical Discipline Fund (grant no. XKQ2015008; awarded to Dr
Guo-Qin Jiang).
References
|
1
|
Fillmore CM and Kuperwasser C: Human
breast cancer cell lines contain stem-like cells that self-renew,
give rise to phenotypically diverse progeny and survive
chemotherapy. Breast Cancer Res. 10:R252008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nguyen NP, Almeida FS, Chi A, Nguyen LM,
Cohen D, Karlsson U and Vinh-Hung V: Molecular biology of breast
cancer stem cells: Potential clinical applications. Cancer Treat
Rev. 36:485–491. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pajonk F, Vlashi E and McBride WH:
Radiation resistance of cancer stem cells: The 4 R's of
radiobiology revisited. Stem Cells. 28:639–648. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kai K, Arima Y, Kamiya T and Saya H:
Breast cancer stem cells. Breast Cancer. 17:80–85. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ and Clarke MF: Prospective identification of
tumorigenic breast cancer cells. Proc Natl Acad Sci USA.
100:3983–3988. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sheridan C, Kishimoto H, Fuchs RK,
Mehrotra S, Bhat-Nakshatri P, Turner CH, Goulet R Jr, Badve S and
Nakshatri H: CD44+/CD24− breast cancer cells
exhibit enhanced invasive properties: An early step necessary for
metastasis. Breast Cancer Res. 8:R592006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shipitsin M, Campbell LL, Argani P,
Weremowicz S, Bloushtain-Qimron N, Yao J, Nikolskaya T,
Serebryiskaya T, Beroukhim R, Hu M, et al: Molecular definition of
breast tumor heterogeneity. Cancer Cell. 11:259–273. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hwang-Verslues WW, Kuo WH, Chang PH, Pan
CC, Wang HH, Tsai ST, Jeng YM, Shew JY, Kung JT, Chen CH, et al:
Multiple lineages of human breast cancer stem/progenitor cells
identified by profiling with stem cell markers. PLoS One.
4:e83772009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Harper JW and Elledge SJ: The DNA damage
response: Ten years after. Mol Cell. 28:739–745. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ackerstaff E, Glunde K and Bhujwalla ZM:
Choline phospholipid metabolism: A target in cancer cells? J Cell
Biochem. 90:525–533. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
de Ramírez Molina A, Gutiérrez R, Ramos
MA, Silva JM, Silva J, Bonilla F, Sánchez JJ and Lacal JC:
Increased choline kinase activity in human breast carcinomas:
Clinical evidence for a potential novel antitumor strategy.
Oncogene. 21:4317–4322. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Glunde K, Ackerstaff E, Natarajan K,
Artemov D and Bhujwalla ZM: Real-time changes in 1H and 31P NMR
spectra of malignant human mammary epithelial cells during
treatment with the anti-inflammatory agent indomethacin. Magn Reson
Med. 48:819–825. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hernández-Alcoceba R, Saniger L, Campos J,
Núñez MC, Khaless F, Gallo MA, Espinosa A and Lacal JC: Choline
kinase inhibitors as a novel approach for antiproliferative drug
design. Oncogene. 15:2289–2301. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rodríguez-González A, de Ramirez Molina A,
Fernández F and Lacal JC: Choline kinase inhibition induces the
increase in ceramides resulting in a highly specific and selective
cytotoxic antitumoral strategy as a potential mechanism of action.
Oncogene. 23:8247–8259. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Glunde K, Raman V, Mori N and Bhujwalla
ZM: RNA interference-mediated choline kinase suppression in breast
cancer cells induces differentiation and reduces proliferation.
Cancer Res. 65:11034–11043. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bensimon J, Altmeyer-Morel S, Benjelloun
H, Chevillard S and Lebeau J: CD24(-/low) stem-like breast cancer
marker defines the radiation-resistant cells involved in
memorization and transmission of radiation-induced genomic
instability. Oncogene. 32:251–258. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Curman D, Cinel B, Williams DE, Rundle N,
Block WD, Goodarzi AA, Hutchins JR, Clarke PR, Zhou BB, Lees-Miller
SP, et al: Inhibition of the G2 DNA damage checkpoint and of
protein kinases chkl and chk2 by the marine sponge alkaloid
debromohymenialdisine. J Biol Chem. 276:17914–17919. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Su WY and Gordon T: In vivo exposure to
ozone produces an increase in a 72-kDa heat shock protein in guinea
pigs. J Appl Physiol (1985). 83:707–711. 1997.PubMed/NCBI
|
|
19
|
Dittmann K, Mayer C, Fehrenbacher B,
Schaller M, Raju U, Milas L, Chen DJ, Kehlbach R and Rodemann HP:
Radiation-induced epidermal growth factor receptor nuclear import
is linked to activation of DNA-dependent protein kinase. J Biol
Chem. 280:31182–31189. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Marín L, Minguela A, Torío A, Moya-Quiles
MR, Muro M, Montes-Ares O, Parrado A, Alvarez-López DM and
García-Alonso AM: Flow cytometric quantification of apoptosis and
proliferation in mixed lymphocyte culture. Cytometry A. 51:107–118.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen MS, Woodward WA, Behbod F,
Peddibhotla S, Alfaro MP, Buchholz TA and Rosen JM:
Wnt/beta-catenin mediates radiation resistance of Sca1+
progenitors in an immortalized mammary gland cell line. J Cell Sci.
120:468–477. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Woodward WA, Chen MS, Behbod F, Alfaro MP,
Buchholz TA and Rosen JM: WNT/beta-catenin mediates radiation
resistance of mouse mammary progenitor cells. Proc Natl Acad Sci
USA. 104:618–623. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sansone P, Storci G, Giovannini C,
Pandolfi S, Pianetti S, Taffurelli M, Santini D, Ceccarelli C,
Chieco P and Bonafé M: p66Shc/Notch-3 interplay controls
self-renewal and hypoxia survival in human stem/progenitor cells of
the mammary gland expanded in vitro as mammospheres. Stem Cells.
25:807–815. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sancar A, Lindsey-Boltz LA, Unsal-Kaçmaz K
and Linn S: Molecular mechanisms of mammalian DNA repair and the
DNA damage checkpoints. Annu Rev Biochem. 73:39–85. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xiao Z, Xue J, Semizarov D, Sowin TJ,
Rosenberg SH and Zhang H: Novel indication for cancer therapy: Chkl
inhibition sensitizes tumor cells to antimitotice. Int J Cancer.
115:528–538. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang HC, Chou WC, Shieh SY and Shen CY:
Amtaxia telangiectasia mumted and checkpoint kinase 2 regulate
BRCA1 to promote the fidelity of DNA end-joining. Cancer Res.
66:1391–1400. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhuang J, Zhang J, Willers H, Wang H,
Chung JH, van Gent DC, Hallahan DE, Powell SN and Xia F: Checkpoint
kinase 2-mediated Phosphoryiation of BRCAl regulates the fidelity
of nonhomologous end-joining. Cancer Res. 66:1401–1408. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tan Y, Raychaudhuri P and Costa RH: Chk2
medistes stabilization of the FoxM1 transcription factor to
stimulate expression of DNA repair genes. Mol Cell Biol.
27:1007–1016. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Arienti KL, Brunmark A, Axe FU, McClure K,
Lee A, Blevitt J, Neff DK, Huang L, Crawford S, Pandit CR, et al:
Checkpoint kinase inhibitors: SAR and radioprotective properties of
aseries of 2-arylbenzimidazoles. J Med Chem. 48:1873–1885. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bull EE, Dote H, Brady KJ, Burgan WE,
Carter DJ, Cerra MA, Oswald KA, Hollingshead MG, Camphausen K and
Tofilon PJ: Enhanced tumor cell radiosensitivity and a btogation of
G2 and S phase arrest by the Hsp90 inhibitor
17-(dimethylaminoethylamino)-17-demethoxygeldanamycin. Clin Cancer
Res. 10:8077–8084. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kastan MB and Bartek J: Cell-cycle
checkpoints and Cancer. Nature. 432:316–323. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee JH and Paull TT: ATM activation by DNA
double-strand breaks through the Mrell-Rad50-Nbs1 complex. Science.
308:551–554. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer and cancer stem cells. Nature.
414:105–111. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shah T, Wildes F, Penet MF, Winnard PT Jr,
Glunde K, Artemov D, Ackerstaff E, Gimi B, Kakkad S, Raman V and
Bhujwalla ZM: Choline kinase overexpression increases invasiveness
and drug resistance of human breast cancer cells. NMR Biomed.
23:633–642. 2010. View Article : Google Scholar : PubMed/NCBI
|